High-Fat Diet Exposure in Early Life Alters Mammary Metabolic and Inflammatory Microenvironment in Favor of Breast Tumorigenesis Later in Life in Mice




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
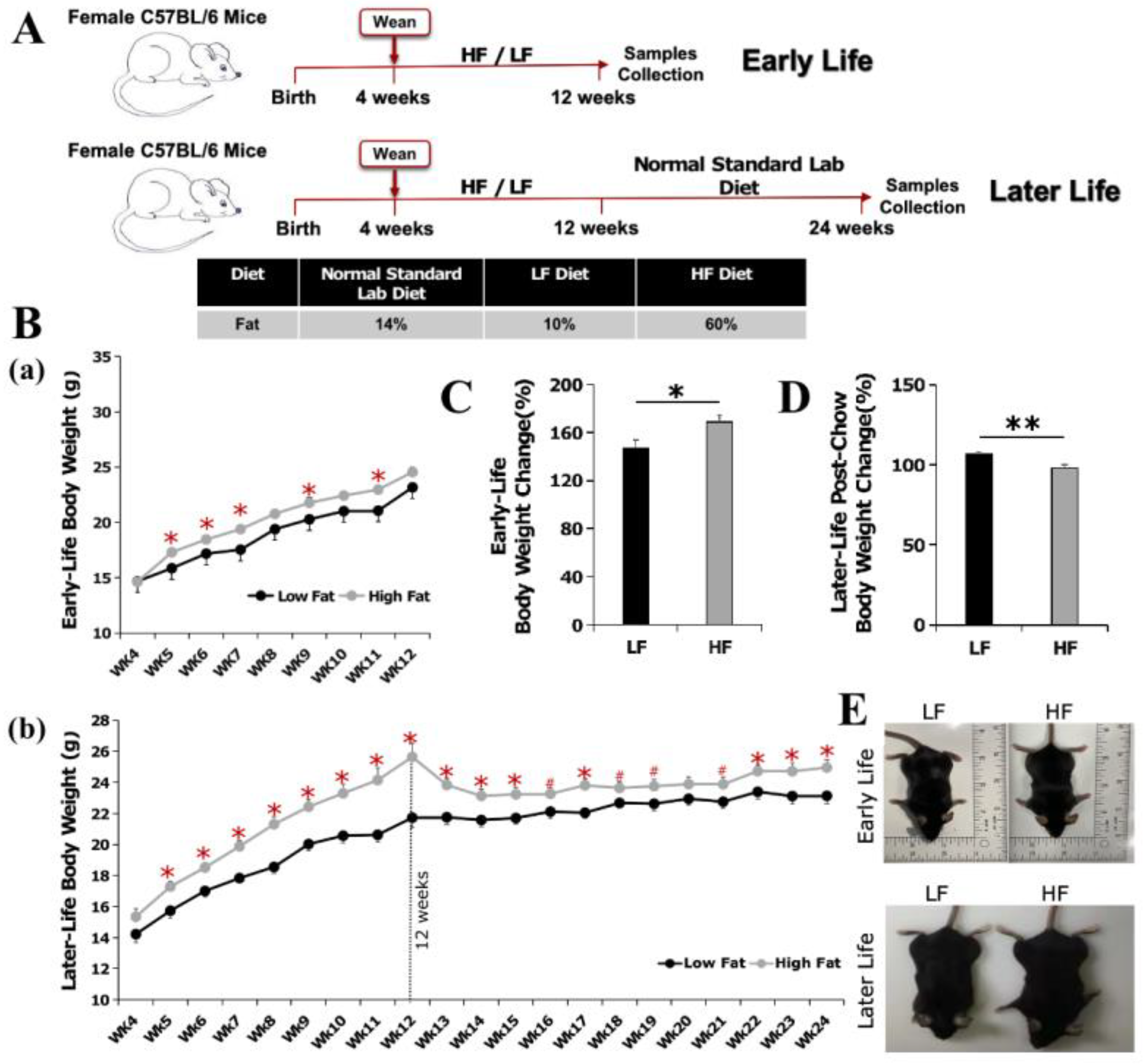
2.1. Animals and Experimental Design
2.2. Real-Time PCR for Gene Expression
2.3. Western Blot Analyses
2.4. Plasma and Mammary Inflammatory Cytokine and Metabolic Hormone Assays
2.5. Statistical Analysis
3. Results
3.1. High-Fat Diet Feeding in Early Life Increased Body Weight Gain, Which Is Retained at a Reduced Magnitude in Later Life of Young Adulthood
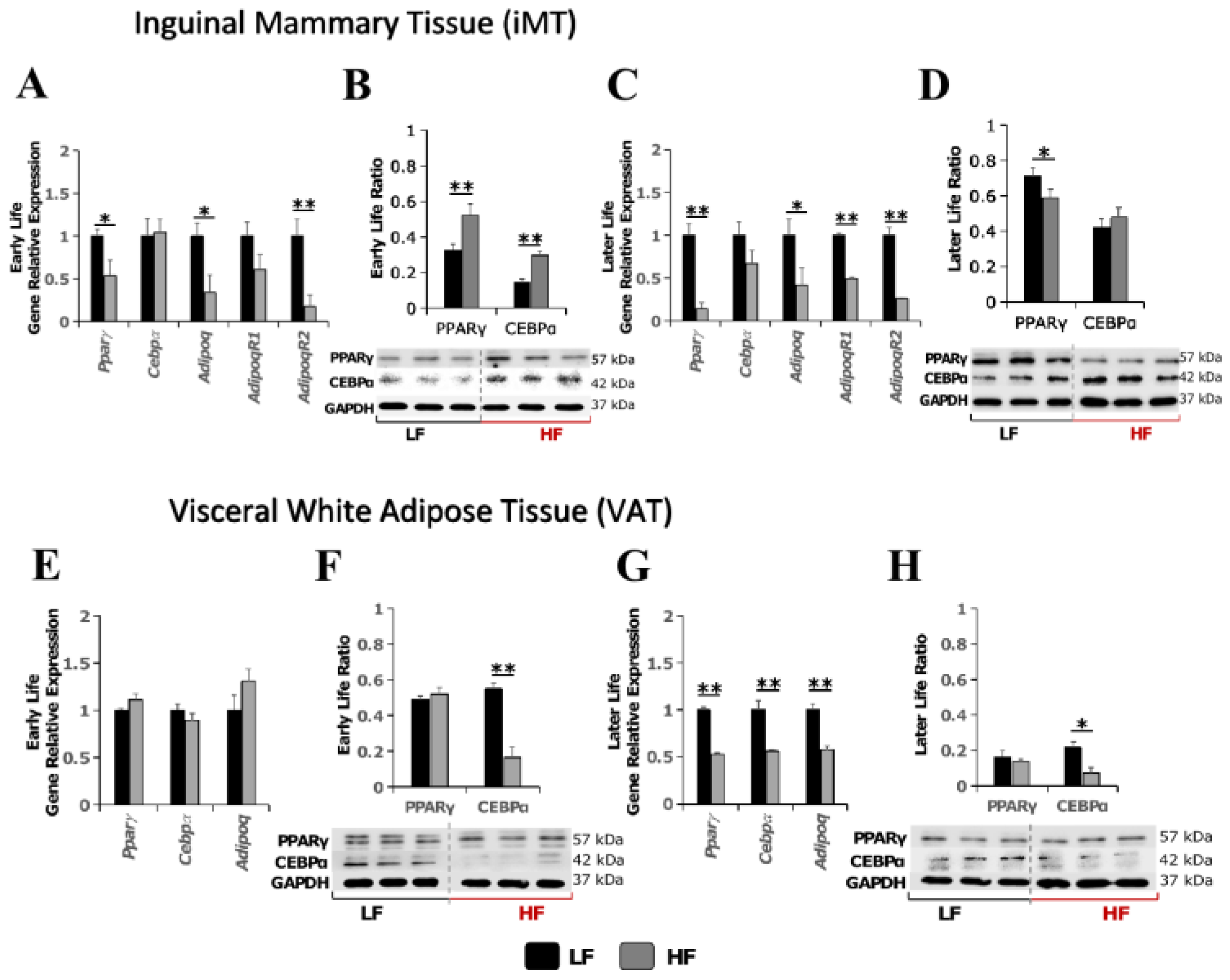
3.2. Influence of Early-Life High-Fat Diet Feeding on Mammary Metabolic Microenvironment
3.2.1. High Fat Diet in Early Life Promoted Adipocyte Dysfunction in Later Life
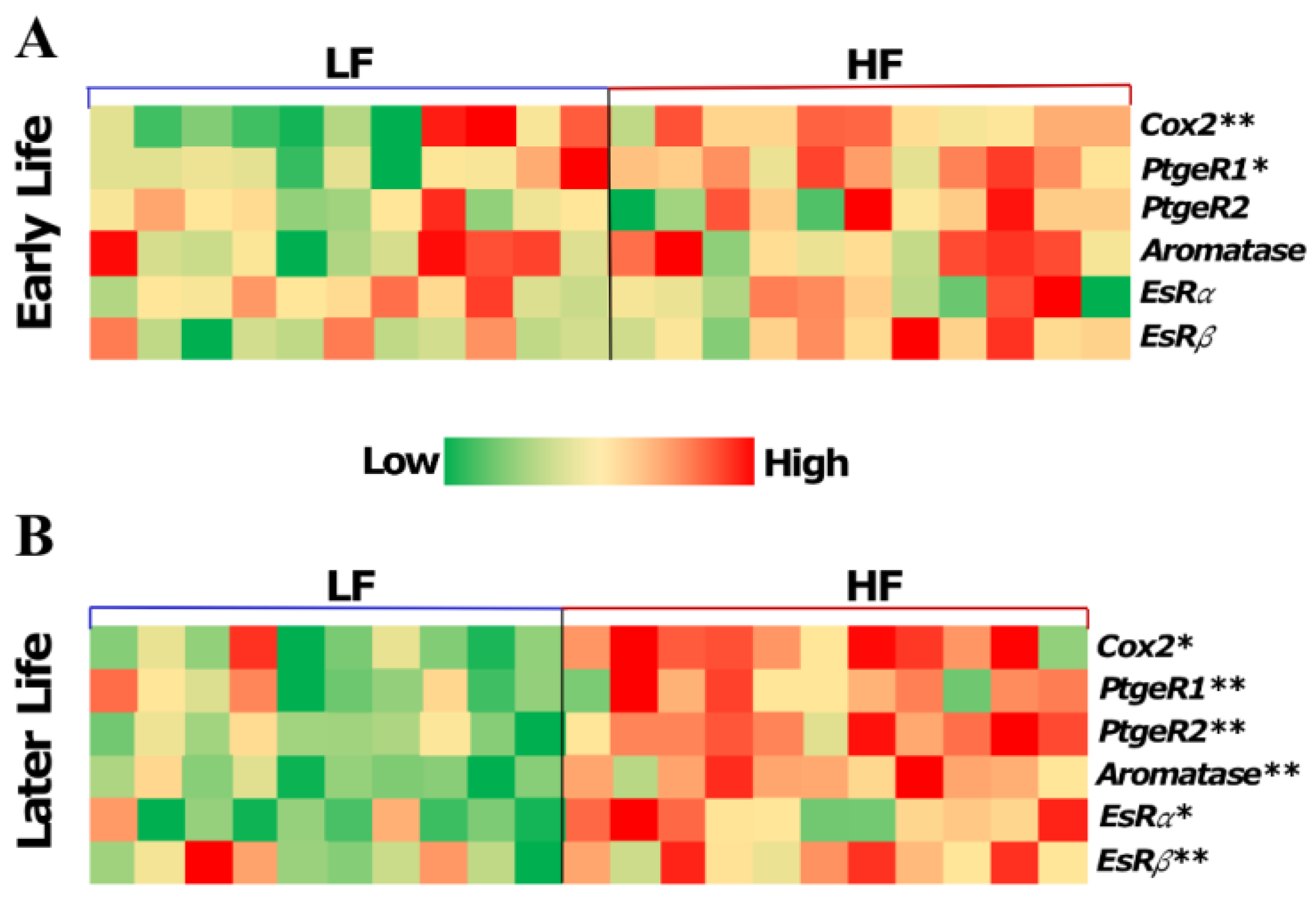
3.2.2. High-Fat Diet in Early Life Promoted Pro-Estrogenic Microenvironment in Later Life
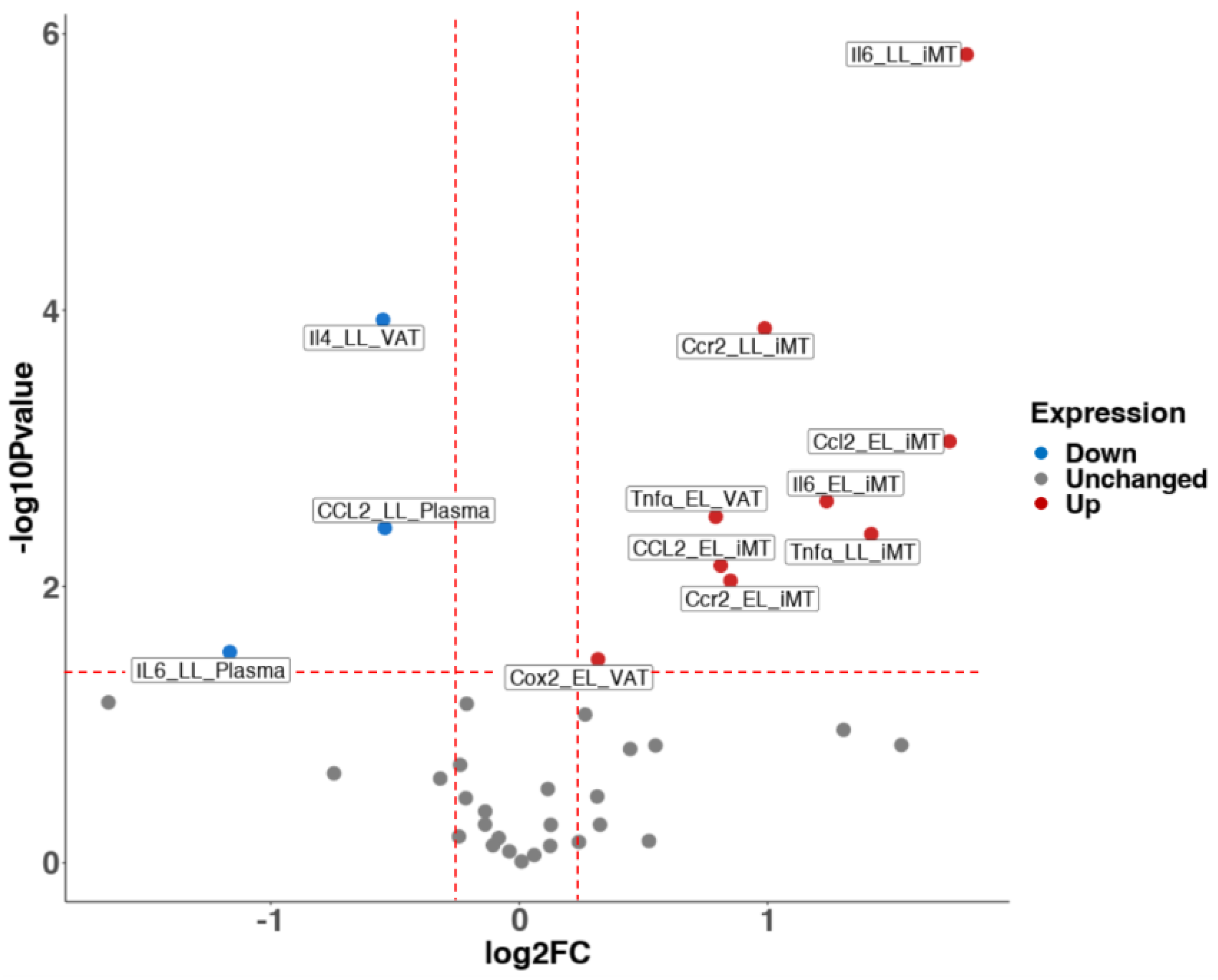
3.3. High-Fat Diet in Early Life Instantly Induced a Pro-Inflammatory Microenvironment, whereas Switching to a Normal Diet Alleviated the Inflammatory Response
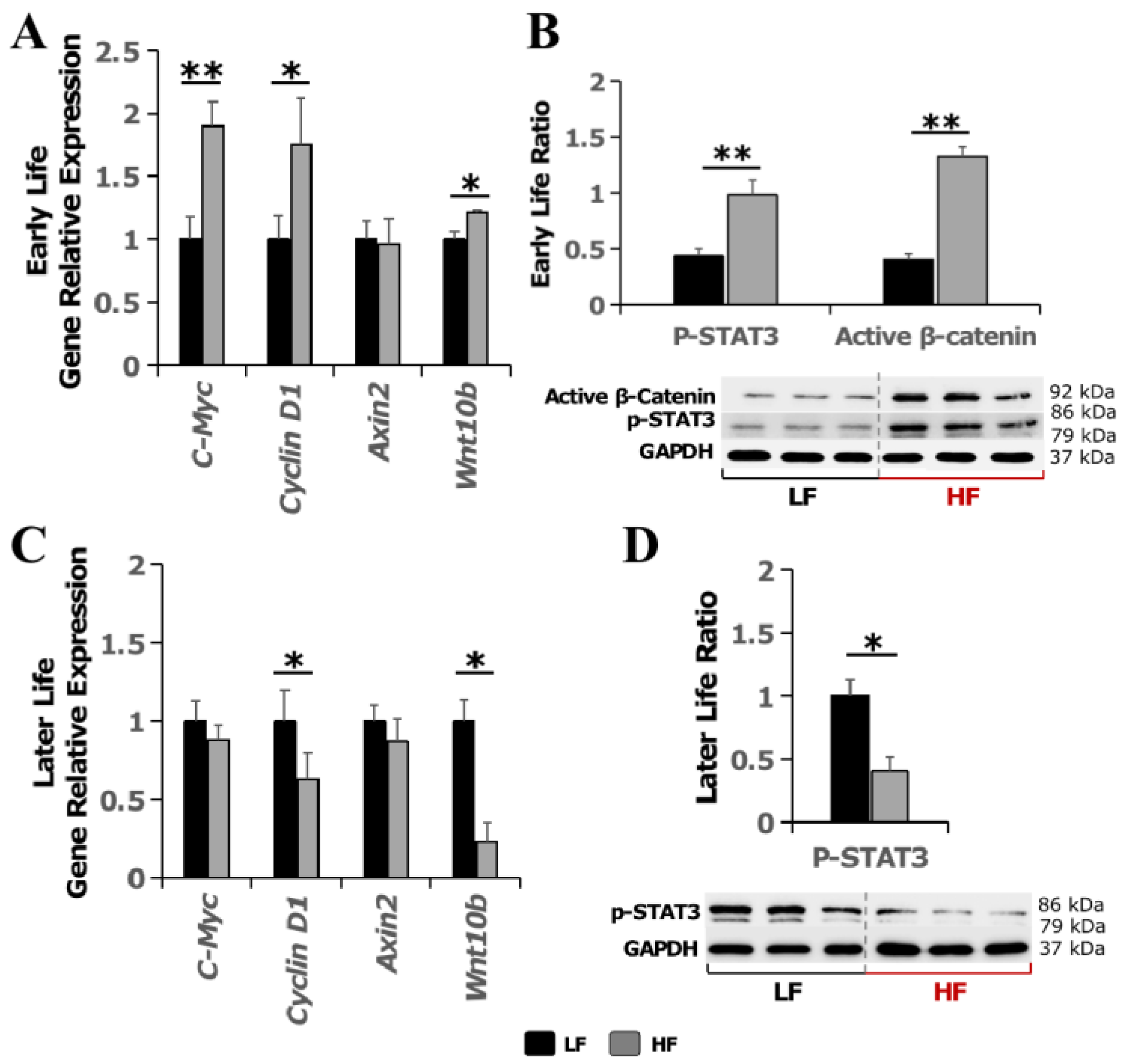
3.4. High-Fat Diet Merely Elevated the Expression of Wnt-Signaling Downstream Genes and Active Β-Catenin in Early Life
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arnold, M.; Morgan, E.; Rumgay, H.; Mafra, A.; Singh, D.; Laversanne, M.; Vignat, J.; Gralow, J.R.; Cardoso, F.; Siesling, S.; et al. Current and future burden of breast cancer: Global statistics for 2020 and 2040. Breast 2022, 66, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Giaquinto, A.N.; Sung, H.; Miller, K.D.; Kramer, J.L.; Newman, L.A.; Minihan, A.; Jemal, A.; Siegel, R.L. Breast Cancer Statistics, 2022. CA Cancer J. Clin. 2022, 72, 524–541. [Google Scholar] [CrossRef] [PubMed]
- Rossi, L.; Mazzara, C.; Pagani, O. Diagnosis and Treatment of Breast Cancer in Young Women. Curr. Treat. Opt. Oncol. 2019, 20, 86. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, L.N. Clinical and Psychosocial Challenges of Breast Cancer in Adolescent and Young Adult Women Under the Age of 40 Years. JCO Oncol. Pract. 2021, 17, 317–319. [Google Scholar] [CrossRef]
- Kehm, R.D.; Yang, W.; Tehranifar, P.; Terry, M.B. 40 Years of Change in Age- and Stage-Specific Cancer Incidence Rates in US Women and Men. JNCI Cancer Spectr. 2019, 3, pkz038. [Google Scholar] [CrossRef]
- Cathcart-Rake, E.J.; Ruddy, K.J.; Bleyer, A.; Johnson, R.H. Breast Cancer in Adolescent and Young Adult Women Under the Age of 40 Years. JCO Oncol. Pract. 2021, 17, 305–313. [Google Scholar] [CrossRef]
- Kashyap, D.; Pal, D.; Sharma, R.; Garg, V.K.; Goel, N.; Koundal, D.; Zaguia, A.; Koundal, S.; Belay, A. Global Increase in Breast Cancer Incidence: Risk Factors and Preventive Measures. BioMed Res. Int. 2022, 2022, 9605439. [Google Scholar] [CrossRef]
- Munsell, M.F.; Sprague, B.L.; Berry, D.A.; Chisholm, G.; Trentham-Dietz, A. Body mass index and breast cancer risk according to postmenopausal estrogen-progestin use and hormone receptor status. Epidemiol. Rev. 2014, 36, 114–136. [Google Scholar] [CrossRef]
- Neuhouser, M.L.; Aragaki, A.K.; Prentice, R.L.; Manson, J.E.; Chlebowski, R.; Carty, C.L.; Ochs-Balcom, H.M.; Thomson, C.A.; Caan, B.J.; Tinker, L.F.; et al. Overweight, Obesity, and Postmenopausal Invasive Breast Cancer Risk: A Secondary Analysis of the Women’s Health Initiative Randomized Clinical Trials. JAMA Oncol. 2015, 1, 611–621. [Google Scholar] [CrossRef]
- Boutari, C.; Mantzoros, C.S. Inflammation: A key player linking obesity with malignancies. Metab. Clin. Exp. 2018, 81, A3–A6. [Google Scholar] [CrossRef]
- Picon-Ruiz, M.; Morata-Tarifa, C.; Valle-Goffin, J.J.; Friedman, E.R.; Slingerland, J.M. Obesity and adverse breast cancer risk and outcome: Mechanistic insights and strategies for intervention. CA Cancer J. Clin. 2017, 67, 378–397. [Google Scholar] [CrossRef] [PubMed]
- Maccio, A.; Madeddu, C. Obesity, inflammation, and postmenopausal breast cancer: Therapeutic implications. Sci. World J. 2011, 11, 806787. [Google Scholar] [CrossRef] [PubMed]
- Key, T.J.; Appleby, P.N.; Reeves, G.K.; Travis, R.C.; Brinton, L.A.; Helzlsouer, K.J.; Dorgan, J.F.; Gapstur, S.M.; Gaudet, M.M.; Kaaks, R.; et al. Steroid hormone measurements from different types of assays in relation to body mass index and breast cancer risk in postmenopausal women: Reanalysis of eighteen prospective studies. Steroids 2015, 99 Pt A, 49–55. [Google Scholar] [CrossRef]
- Stierman, B.; Afful, J.; Carroll, M.D.; Chen, T.-C.; Davy, O.; Fink, S.; Fryar, C.D.; Gu, Q.; Hales, C.M.; Hughes, J.P. National Health and Nutrition Examination Survey 2017–March 2020 Prepandemic Data Files Development of Files and Prevalence Estimates for Selected Health Outcomes. 2021. Available online: https://stacks.cdc.gov/view/cdc/106273 (accessed on 18 January 2023).
- Simmonds, M.; Llewellyn, A.; Owen, C.G.; Woolacott, N. Predicting adult obesity from childhood obesity: A systematic review and meta-analysis. Obes. Rev. 2016, 17, 95–107. [Google Scholar] [CrossRef]
- Colditz, G.A.; Bohlke, K.; Berkey, C.S. Breast cancer risk accumulation starts early: Prevention must also. Breast Cancer Res. Treat. 2014, 145, 567–579. [Google Scholar] [CrossRef] [PubMed]
- Clarke, M.A.; Joshu, C.E. Early Life Exposures and Adult Cancer Risk. Epidemiol. Rev. 2017, 39, 11–27. [Google Scholar] [CrossRef]
- Oh, H.; Boeke, C.E.; Tamimi, R.M.; Smith-Warner, S.A.; Wang, M.L.; Willett, W.C.; Eliassen, A. The interaction between early-life body size and physical activity on risk of breast cancer. Int. J. Cancer 2015, 137, 571–581. [Google Scholar] [CrossRef]
- Baer, H.J.; Tworoger, S.S.; Hankinson, S.E.; Willett, W.C. Body Fatness at Young Ages and Risk of Breast Cancer Throughout Life. Am. J. Epidemiol. 2010, 171, 1183–1194. [Google Scholar] [CrossRef]
- Flurkey, K.; Currer, J.M.; Harrison, D.E. Mouse models in aging research. In The Mouse in Biomedical Research; Academic Press: Cambridge, MA, USA, 2007. [Google Scholar]
- Dutta, S.; Sengupta, P. Men and mice: Relating their ages. Life Sci. 2016, 152, 244–248. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, M.; Xu, F.; Jiang, S. Wnt signaling in breast cancer: Biological mechanisms, challenges and opportunities. Mol. Cancer 2020, 19, 165. [Google Scholar] [CrossRef]
- Abreu de Oliveira, W.A.; El Laithy, Y.; Bruna, A.; Annibali, D.; Lluis, F. Wnt Signaling in the Breast: From Development to Disease. Front. Cell Dev. Biol. 2022, 10, 884467. [Google Scholar] [CrossRef] [PubMed]
- To, S.Q.; Dmello, R.S.; Richards, A.K.; Ernst, M.; Chand, A.L. STAT3 Signaling in Breast Cancer: Multicellular Actions and Therapeutic Potential. Cancers 2022, 14, 429. [Google Scholar] [CrossRef] [PubMed]
- Hruby, A.; Hu, F.B. The Epidemiology of Obesity: A Big Picture. Pharmacoeconomics 2015, 33, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Hales, C.; Fryar, C.D. Prevalence of Obesity and Severe Obesity Among Persons Aged 2–19 Years—National Health and Nutrition Examination Survey, 1999–2000 through 2017–2018. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 390. [Google Scholar]
- Nuotio, J.; Laitinen, T.T.; Sinaiko, A.R.; Woo, J.G.; Urbina, E.M.; Jacobs, D.R.; Steinberger, J.; Prineas, R.J.; Sabin, M.A.; Burgner, D.P.; et al. Obesity during childhood is associated with higher cancer mortality rate during adulthood: The i3C Consortium. Int. J. Obes. 2022, 46, 393–399. [Google Scholar] [CrossRef]
- Lefterova, M.I.; Zhang, Y.; Steger, D.J.; Schupp, M.; Schug, J.; Cristancho, A.; Feng, D.; Zhuo, D.; Stoeckert, C.J., Jr.; Liu, X.S.; et al. PPARgamma and C/EBP factors orchestrate adipocyte biology via adjacent binding on a genome-wide scale. Genes Dev. 2008, 22, 2941–2952. [Google Scholar] [CrossRef]
- Hernandez-Quiles, M.; Broekema, M.F.; Kalkhoven, E. PPARgamma in Metabolism, Immunity, and Cancer: Unified and Diverse Mechanisms of Action. Front. Endocrinol. 2021, 12, 624112. [Google Scholar] [CrossRef]
- Augimeri, G.; Giordano, C.; Gelsomino, L.; Plastina, P.; Barone, I.; Catalano, S.; Ando, S.; Bonofiglio, D. The Role of PPARgamma Ligands in Breast Cancer: From Basic Research to Clinical Studies. Cancers 2020, 12, 2623. [Google Scholar] [CrossRef]
- Pham, D.V.; Park, P.H. Adiponectin triggers breast cancer cell death via fatty acid metabolic reprogramming. J. Exp. Clin. Cancer Res. 2022, 41, 9. [Google Scholar] [CrossRef]
- Margalit, O.; Wang, D.; DuBois, R.N. PPARγ Agonists Target Aromatase Via Both PGE2 and BRCA1. Cancer Prev. Res. 2012, 5, 1169–1172. [Google Scholar] [CrossRef]
- Fan, W.; Yanase, T.; Morinaga, H.; Mu, Y.M.; Nomura, M.; Okabe, T.; Goto, K.; Harada, N.; Nawata, H. Activation of peroxisome proliferator-activated receptor-gamma and retinoid X receptor inhibits aromatase transcription via nuclear factor-kappaB. Endocrinology 2005, 146, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Subbaramaiah, K.; Morris, P.G.; Zhou, X.K.; Morrow, M.; Du, B.; Giri, D.; Kopelovich, L.; Hudis, C.A.; Dannenberg, A.J. Increased levels of COX-2 and prostaglandin E2 contribute to elevated aromatase expression in inflamed breast tissue of obese women. Cancer Discov. 2012, 2, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, P.; Au, C.C.; Benito-Martin, A.; Ladumor, H.; Oshchepkova, S.; Moges, R.; Brown, K.A. Estrogens and breast cancer: Mechanisms involved in obesity-related development, growth and progression. J. Steroid Biochem. Mol. Biol. 2019, 189, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Kolb, R.; Zhang, W. Obesity and Breast Cancer: A Case of Inflamed Adipose Tissue. Cancers 2020, 12, 1686. [Google Scholar] [CrossRef]
- Han, J.M.; Levings, M.K. Immune regulation in obesity-associated adipose inflammation. J. Immunol. 2013, 191, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Danforth, D.N. The Role of Chronic Inflammation in the Development of Breast Cancer. Cancers 2021, 13, 3918. [Google Scholar] [CrossRef]
- Nusse, R.; Varmus, H.E. Many tumors induced by the mouse mammary tumor virus contain a provirus integrated in the same region of the host genome. Cell 1982, 31, 99–109. [Google Scholar] [CrossRef]
- Lin, S.Y.; Xia, W.; Wang, J.C.; Kwong, K.Y.; Spohn, B.; Wen, Y.; Pestell, R.G.; Hung, M.C. Beta-catenin, a novel prognostic marker for breast cancer: Its roles in cyclin D1 expression and cancer progression. Proc. Natl. Acad. Sci. USA 2000, 97, 4262–4266. [Google Scholar] [CrossRef]
- Wulf, G.M.; Ryo, A.; Wulf, G.G.; Lee, S.W.; Niu, T.; Petkova, V.; Lu, K.P. Pin1 is overexpressed in breast cancer and cooperates with Ras signaling in increasing the transcriptional activity of c-Jun towards cyclin D1. EMBO J. 2001, 20, 3459–3472. [Google Scholar] [CrossRef]
- Roubert, A.; Gregory, K.; Li, Y.; Pfalzer, A.C.; Li, J.; Schneider, S.S.; Wood, R.J.; Liu, Z. The influence of tumor necrosis factor-alpha on the tumorigenic Wnt-signaling pathway in human mammary tissue from obese women. Oncotarget 2017, 8, 36127–36136. [Google Scholar] [CrossRef]
- Frederick, A.M.; Guo, C.; Meyer, A.; Yan, L.; Schneider, S.S.; Liu, Z. The influence of obesity on folate status, DNA methylation and cancer-related gene expression in normal breast tissues from premenopausal women. Epigenetics 2021, 16, 458–467. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Y.; Lin, T.-C.; Kim, Y.-C.; Chung, S.; Liu, Z. High-Fat Diet Exposure in Early Life Alters Mammary Metabolic and Inflammatory Microenvironment in Favor of Breast Tumorigenesis Later in Life in Mice. Curr. Oncol. 2023, 30, 4197-4207. https://doi.org/10.3390/curroncol30040320
Tang Y, Lin T-C, Kim Y-C, Chung S, Liu Z. High-Fat Diet Exposure in Early Life Alters Mammary Metabolic and Inflammatory Microenvironment in Favor of Breast Tumorigenesis Later in Life in Mice. Current Oncology. 2023; 30(4):4197-4207. https://doi.org/10.3390/curroncol30040320
Chicago/Turabian StyleTang, Ying, Ting-Chun Lin, Young-Cheul Kim, Soonkyu Chung, and Zhenhua Liu. 2023. "High-Fat Diet Exposure in Early Life Alters Mammary Metabolic and Inflammatory Microenvironment in Favor of Breast Tumorigenesis Later in Life in Mice" Current Oncology 30, no. 4: 4197-4207. https://doi.org/10.3390/curroncol30040320