Zinc Finger Proteins in Head and Neck Squamous Cell Carcinomas: ZNF540 May Serve as a Biomarker

 , , , ,
, , , ,
Abstract
:1. Introduction
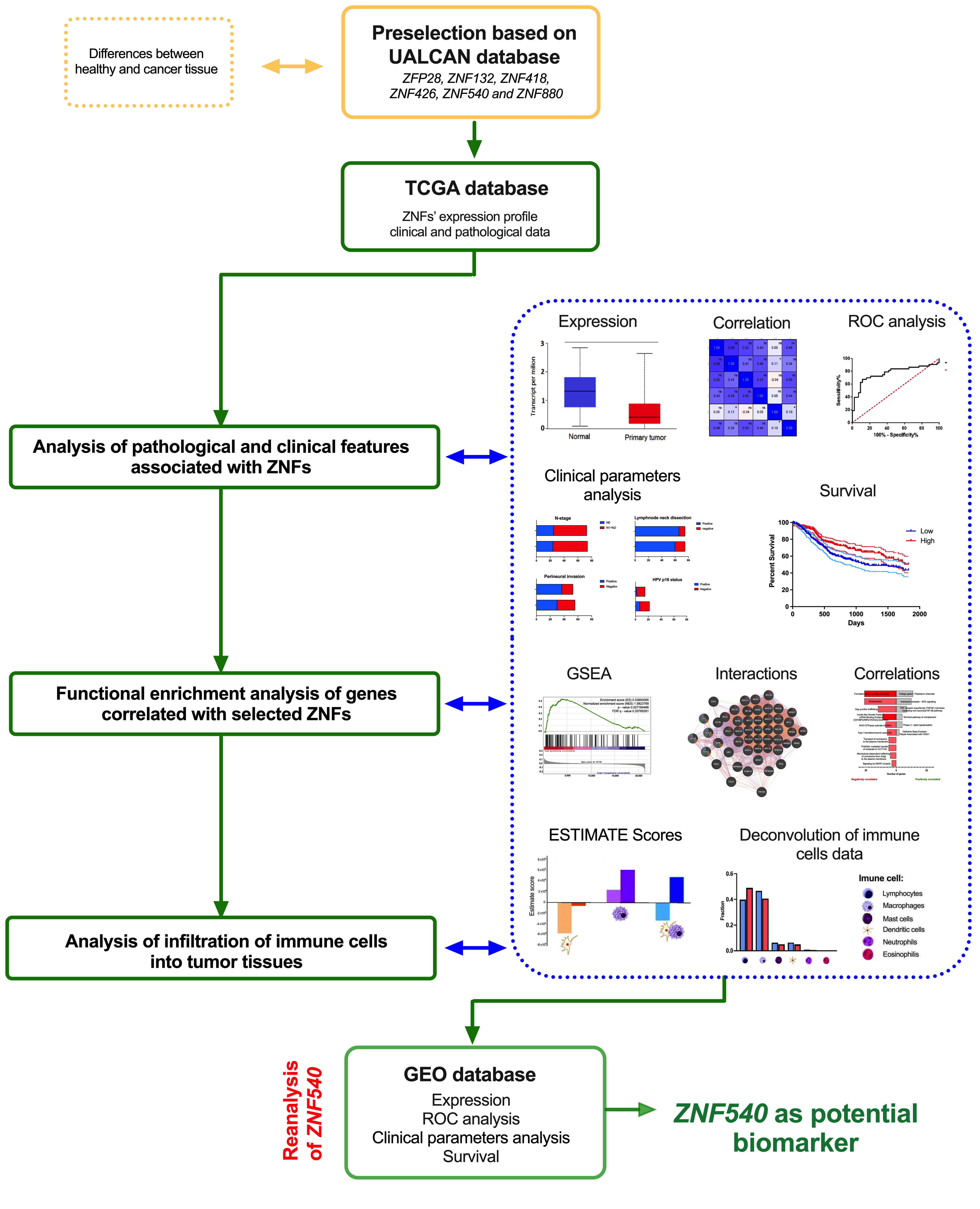
2. Materials and Methods
2.1. TCGA Data
2.2. Pathological and Clinical Analysis
2.3. Functional Enrichment Analysis of Genes Correlated with Selected ZNFs
2.4. Infiltration of Immune Cells into Tumor Tissues
2.5. Statistical Analysis
2.6. Validation of the Results
3. Results
3.1. Expression of the ZNFs Depends on the Tumor Localization and Clinical-Pathological Parameters
3.2. Patients with Low ZNF418 and ZNF540 Expression Display Shorter Overall Survival
3.3. Expression of the ZNFs Is Connected with Critical/Essential Cellular Processes and Pathways
3.4. The Expression Levels of ZNFs Are Associated with the Tumor Immunological Profile in HNSCC Patients
3.5. Validation of ZNF540 as a Potential Biomarker Using GEO Data
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chow, L.Q.M. Head and Neck Cancer. N. Engl. J. Med. 2020, 382, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Young, D.; Xiao, C.C.; Murphy, B.; Moore, M.; Fakhry, C.; Day, T.A. Increase in head and neck cancer in younger patients due to human papillomavirus (HPV). Oral Oncol. 2015, 51, 727–730. [Google Scholar] [CrossRef] [PubMed]
- Rajabi-Moghaddam, M.; Abbaszadeh, H. Gene polymorphisms and prognosis of head and neck squamous cell carcinoma: A systematic review. Rep. Pract. Oncol. Radiother. 2022; ahead of print. [Google Scholar] [CrossRef]
- Rajabi-Moghaddam, M.; Abbaszadeh, H. Gene polymorphisms and risk of head and neck squamous cell carcinoma: A systematic review. Rep. Pract. Oncol. Radiother. 2022; ahead of print. [Google Scholar] [CrossRef]
- Guglas, K.; Kozłowska-Masłoń, J.; Kolenda, T.; Paszkowska, A.; Teresiak, A.; Bliźniak, R.; Lamperska, K. Midsize noncoding RNAs in cancers: A new division that clarifies the world of noncoding RNA or an unnecessary chaos? Rep. Pract. Oncol. Radiother. 2022; ahead of print. [Google Scholar] [CrossRef]
- Kolenda, T.; Guglas, K.; Baranowski, D.; Sobocińska, J.; Kopczyńska, M.; Teresiak, A.; Bliźniak, R.; Lamperska, K. cfRNAs as biomarkers in oncology—still experimental or applied tool for personalized medicine already? Rep. Pract. Oncol. Radiother. 2020, 25, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Krishna, S.S.; Majumdar, I.; Grishin, N.V. Structural classification of zinc fingers: Survey and summary. Nucleic Acids Res. 2003, 31, 532–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamsjaeger, R.; Liew, C.K.; Loughlin, F.E.; Crossley, M.; Mackay, J.P. Sticky fingers: Zinc-fingers as protein-recognition motifs. Trends Biochem. Sci. 2007, 32, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.K.; van Attikum, H. DNA double-strand break repair: Putting zinc fingers on the sore spot. Semin. Cell Dev. Biol. 2021, 113, 65–74. [Google Scholar] [CrossRef]
- Ecco, G.; Imbeault, M.; Trono, D. KRAB zinc finger proteins. Development 2017, 144, 2719–2729. [Google Scholar] [CrossRef]
- Jen, J.; Wang, Y.C. Zinc finger proteins in cancer progression. J. Biomed. Sci. 2016, 23, 53. [Google Scholar] [CrossRef] [Green Version]
- Collins, T.; Stone, J.R.; Williams, A.J. All in the family: The BTB/POZ, KRAB, and SCAN domains. Mol. Cell. Biol. 2001, 21, 3609–3615. [Google Scholar] [CrossRef] [Green Version]
- Najafabadi, H.S.; Mnaimneh, S.; Schmitges, F.W.; Garton, M.; Lam, K.N.; Yang, A.; Albu, M.; Weirauch, M.T.; Radovani, E.; Kim, P.M.; et al. C2H2 zinc finger proteins greatly expand the human regulatory lexicon. Nat. Biotechnol. 2015, 33, 555–562. [Google Scholar] [CrossRef]
- Sobocińska, J.; Molenda, S.; Machnik, M.; Oleksiewicz, U. KRAB-ZFP Transcriptional Regulators Acting as Oncogenes and Tumor Suppressors: An Overview. Int. J. Mol. Sci. 2021, 22, 2212. [Google Scholar] [CrossRef] [PubMed]
- Yajima, I.; Kumasaka, M.; Thang, N.D.; Yanagishita, T.; Ohgami, N.; Kallenberg, D.; Naito, Y.; Yoshikawa, T.; Sakashita, N.; Kato, M. Zinc finger protein 28 as a novel melanoma-related molecule. J. Dermatol. Sci. 2009, 55, 68–70. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.L.; Chen, Y.J.; Chang, W.A.; Jian, S.F.; Fan, H.L.; Wang, J.Y.; Kuo, P.L. Interaction between Tumor-Associated Dendritic Cells and Colon Cancer Cells Contributes to Tumor Progression via CXCL1. Int. J. Mol. Sci. 2018, 19, 2427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.; Hong, H.; Zhang, S.; Zong, Y.; Ma, J.; Lu, A.; Sun, J.; Zheng, M. Identification of key genes and pathways involved in microsatellite instability in colorectal cancer. Mol. Med. Rep. 2019, 19, 2065–2076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arai, E.; Chiku, S.; Mori, T.; Gotoh, M.; Nakagawa, T.; Fujimoto, H.; Kanai, Y. Single-CpG-resolution methylome analysis identifies clinicopathologically aggressive CpG island methylator phenotype clear cell renal cell carcinomas. Carcinogenesis 2012, 33, 1487–1493. [Google Scholar] [CrossRef]
- Hui, H.X.; Hu, Z.W.; Jiang, C.; Wu, J.; Gao, Y.; Wang, X.W. ZNF418 overexpression protects against gastric carcinoma and prompts a good prognosis. Onco Targets Ther. 2018, 11, 2763–2770. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; He, Z.; Wang, C.; Zhou, Y.; Li, F.; Pu, W.; Zhang, X.; Feng, X.; Zhang, M.; Yecheng, X.; et al. Epigenetic silencing of ZNF132 mediated by methylation-sensitive Sp1 binding promotes cancer progression in esophageal squamous cell carcinoma. Cell Death Dis. 2018, 10, 1. [Google Scholar] [CrossRef]
- Machnik, M.; Cylwa, R.; Kiełczewski, K.; Biecek, P.; Liloglou, T.; Mackiewicz, A.; Oleksiewicz, U. The expression signature of cancer-associated KRAB-ZNF factors identified in TCGA pan-cancer transcriptomic data. Mol. Oncol. 2019, 13, 701–724. [Google Scholar] [CrossRef] [Green Version]
- Chandrashekar, D.S.; Karthikeyan, S.K.; Korla, P.K.; Patel, H.; Shovon, A.R.; Athar, M.; Netto, G.J.; Qin, Z.S.; Kumar, S.; Manne, U.; et al. UALCAN: An update to the integrated cancer data analysis platform. Neoplasia 2022, 25, 18–27. [Google Scholar] [CrossRef]
- Lleras, R.A.; Adrien, L.R.; Smith, R.V.; Brown, B.; Jivraj, N.; Keller, C.; Sarta, C.; Schlecht, N.F.; Harris, T.M.; Childs, G.; et al. Hypermethylation of a cluster of Krüppel-type zinc finger protein genes on chromosome 19q13 in oropharyngeal squamous cell carcinoma. Am. J. Pathol. 2011, 178, 1965–1974. [Google Scholar] [CrossRef] [Green Version]
- Lleras, R.A.; Smith, R.V.; Adrien, L.R.; Schlecht, N.F.; Burk, R.D.; Harris, T.M.; Childs, G.; Prystowsky, M.B.; Belbin, T.J. Unique DNA methylation loci distinguish anatomic site and HPV status in head and neck squamous cell carcinoma. Clin. Cancer Res. 2013, 19, 5444–5455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, J.; Wang, L.; Zhong, W.; Chen, Z.; Chen, J.; Yang, H.; Liu, G. Development and Validation of Epigenetic Signature Predict Survival for Patients with Laryngeal Squamous Cell Carcinoma. DNA Cell Biol. 2021, 40, 247–264. [Google Scholar] [CrossRef] [PubMed]
- Tomaszewska, W.; Kozłowska-Masłoń, J.; Baranowski, D.; Perkowska, A.; Szałkowska, S.; Kazimierczak, U.; Severino, P.; Lamperska, K.; Kolenda, T. miR-154 Influences HNSCC Development and Progression through Regulation of the Epithelial-to-Mesenchymal Transition Process and Could Be Used as a Potential Biomarker. Biomedicines 2021, 9, 1894. [Google Scholar] [CrossRef]
- Thorsson, V.; Gibbs, D.L.; Brown, S.D.; Wolf, D.; Bortone, D.S.; Ou Yang, T.H.; Porta-Pardo, E.; Gao, G.F.; Plaisier, C.L.; Eddy, J.A.; et al. The Immune Landscape of Cancer. Immunity 2019, 48, 812–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wichmann, G.; Rosolowski, M.; Krohn, K.; Kreuz, M.; Boehm, A.; Reiche, A.; Scharrer, U.; Halama, D.; Bertolini, J.; Bauer, U.; et al. The role of HPV RNA transcription, immune response-related gene expression and disruptive TP53 mutations in diagnostic and prognostic profiling of head and neck cancer. Int. J. Cancer 2015, 137, 2846–2857. [Google Scholar] [CrossRef]
- Hamada, K.; Tian, Y.; Fujimoto, M.; Takahashi, Y.; Kohno, T.; Tsuta, K.; Watanabe, S.I.; Yoshida, T.; Asamura, H.; Kanai, Y.; et al. DNA hypermethylation of the ZNF132 gene participates in the clinicopathological aggressiveness of ‘pan-negative’-type lung adenocarcinomas. Carcinogenesis 2021, 42, 169–179. [Google Scholar] [CrossRef]
- Zhang, P.; Li, S.; Zhang, T.; Cui, F.; Shi, J.H.; Zhao, F.; Sheng, X. Characterization of Molecular Subtypes in Head and Neck Squamous Cell Carcinoma With Distinct Prognosis and Treatment Responsiveness. Front. Cell Dev. Biol. 2021, 9, 711348. [Google Scholar] [CrossRef]
- Sun, S.C. Non-canonical NF-κB signaling pathway. Cell Res. 2011, 21, 71–85. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.A.; Lee, K.H.; Lee, D.H.; Lee, J.K.; Lim, S.C.; Joo, Y.E.; Chung, I.J.; Noh, M.G.; Yoon, T.M. Receptor tyrosine kinase, RON, promotes tumor progression by regulating EMT and the MAPK signaling pathway in human oral squamous cell carcinoma. Int. J. Oncol. 2019, 55, 513–526. [Google Scholar] [CrossRef]
- Smith, A.; Teknos, T.N.; Pan, Q. Epithelial to mesenchymal transition in head and neck squamous cell carcinoma. Oral Oncol. 2013, 49, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Tomar, S. Differential Gene Expression Patterns in HPV-Positive and HPV-Negative Oropharyngeal Carcinomas. Ph.D. Thesis, University of South Carolina, Columbia, SC, USA, 2013. Available online: https://scholarcommons.sc.edu/cgi/viewcontent.cgi?article=4577&context=etd (accessed on 25 October 2022).
- Cillo, A.R.; Kürten, C.H.L.; Tabib, T.; Qi, Z.; Onkar, S.; Wang, T.; Liu, A.; Duvvuri, U.; Kim, S.; Soose, R.J.; et al. Immune Landscape of Viral- and Carcinogen-Driven Head and Neck Cancer. Immunity 2020, 52, 183–199. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FP28 | ZNF880 | ZNF540 | ZNF418 | ZNF426 | ZNF132 | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameter | Group | Mean ± SEM | p-Val | n | Mean ± SEM | p-Val | n | Mean ± SEM | p-Val | n | Mean ± SEM | p-Val | n | Mean ± SEM | p-Val | n | Mean ± SEM | p-Val | n |
| Age | <61 | −1.666 ± 0.1029 | 0.0284 | 258 | −1.753 ± 0.08617 | 0.4497 | 258 | −2.023 ± 0.07620 | 0.0016 | 258 | −2.436 ± 0.07546 | 0.0429 | 258 | 0.2168 ± 0.09069 | 0.9285 | 258 | −1.024 ± 0.05959 | 0.0020 | 258 |
| >61 | −1.971 ± 0.09452 | 263 | −1.848 ± 0.09072 | 263 | −2.354 ± 0.07172 | 263 | −2.736 ± 0.07637 | 263 | 0.2706 ± 0.07680 | 263 | −1.273 ± 0.05513 | 263 | |||||||
| Gender | Female | −1.878 ± 0.1224 | 0.6950 | 137 | −1.856 ± 0.1226 | 0.5837 | 137 | −2.430 ± 0.09578 | 0.0060 | 137 | −2.655 ± 0.1031 | 0.2337 | 137 | 0.5759 ± 0.10140 | 0.0080 | 137 | −1.328 ± 0.06942 | 0.0043 | 173 |
| Male | −1.796 ± 0.08431 | 385 | −1.778 ± 0.07269 | 385 | −2.102 ± 0.06225 | 385 | −2.513 ± 0.06598 | 385 | 0.1259 ± 0.07089 | 385 | −1.087 ± 0.04916 | 385 | |||||||
| Alcohol | Positive | −1.726 ± 0.08599 | 0.0480 | 348 | −1.752 ± 0.07466 | 0.1381 | 348 | −2.151 ± 0.06220 | 0.2524 | 348 | −2.516 ± 0.06876 | 0.2158 | 348 | 0.2608 ± 0.07361 | 0.1041 | 348 | −1.094 ± 0.04794 | 0.0568 | 348 |
| Negative | −2.027 ± 0.1232 | 163 | −1.952 ± 0.1160 | 163 | −2.283 ± 0.1014 | 163 | −2.683 ± 0.09407 | 163 | 0.2041 ± 0.1056 | 163 | −1.272 ± 0.07984 | 163 | |||||||
| Smoking | No/Ex | −1.994 ± 0.08627 | 0.0004 | 333 | −1.902 ± 0.07998 | 0.0456 | 333 | −2.234 ± 0.06583 | 0.3147 | 333 | −2.664 ± 0.06435 | 0.0091 | 333 | 0.2411 ± 0.07058 | 0.1674 | 333 | −1.232 ± 0.04994 | 0.0063 | 333 |
| Yes | −1.508 ± 0.1173 | 177 | −1.638 ± 0.1004 | 177 | −2.122 ± 0.08948 | 117 | −2.332 ± 0.1080 | 117 | 0.2861 ± 0.1105 | 177 | −0.9967 ± 0.06593 | 177 | |||||||
| Cancer Stage | I + II | −1.744 ± 0.1480 | 0.1890 | 101 | −1.771 ± 0.1227 | 0.6981 | 101 | −2.212 ± 0.1121 | 0.5988 | 101 | −2.579 ± 0.1212 | 0.8857 | 101 | 0.4043 ± 0.12760 | 0.3499 | 101 | −1.167 ± 0.06949 | 0.8409 | 101 |
| III + IV | −1.961 ± 0.08416 | 349 | −1.832 ± 0.07689 | 349 | −2.282 ± 0.06302 | 349 | −2.573 ± 0.06828 | 349 | 0.2019 ± 0.07327 | 349 | −1.211 ± 0.04962 | 349 | |||||||
| T stage | T1 + T2 | −1.719 ± 0.1111 | 0.0325 | 185 | −1.736 ± 0.09599 | 0.3174 | 185 | −2.005 ± 0.08372 | <0.0001 | 185 | −2.470 ± 0.08898 | 0.2256 | 185 | 0.2275 ± 0.09747 | 0.5552 | 185 | −1.057 ± 0.06083 | 0.0317 | 185 |
| T3 + T4 | −2.029 ± 0.09507 | 274 | −1.869 ± 0.08743 | 274 | −2.434 ± 0.06896 | 274 | −2.640 ± 0.07954 | 274 | 0.2760 ± 0.08243 | 274 | −1.266 ± 0.05489 | 274 | |||||||
| N stage | N0 + N1 | −1.907 ± 0.1009 | 0.8130 | 243 | −1.789 ± 0.09103 | 0.7360 | 243 | −2.241 ± 0.06699 | 0.9973 | 243 | −2.598 ± 0.07829 | 0.2244 | 243 | 0.4471 ± 0.08127 | 0.0012 | 243 | −1.275 ± 0.05776 | 0.0064 | 243 |
| N2 + N3 | −1.907 ± 0.1135 | 179 | −1.835 ± 0.1021 | 179 | −2.241 ± 0.09822 | 179 | −2.442 ± 0.1032 | 179 | −0.03371 ± 0.1085 | 179 | −1.042 ± 0.06325 | 179 | |||||||
| Grade | G1 + G2 | −1.968 ± 0.08064 | 0.0033 | 368 | −1.871 ± 0.07577 | 0.1031 | 368 | −2.342 ± 0.06235 | <0.0001 | 368 | −2.692 ± 0.06427 | 0.0001 | 368 | 0.3830 ± 0.06992 | 0.0001 | 368 | −1.262 ± 0.04763 | 0.0012 | 368 |
| G3 + G4 | −1.496 ± 0.1444 | 132 | −1.636 ± 0.1149 | 132 | −1.858 ± 0.09752 | 132 | −2.189 ± 0.1152 | 132 | −0.07011 ± 0.1165 | 132 | −0.9401 ± 0.08017 | 132 | |||||||
| Perineural Invasion | Positive | −2.005 ± 0.1118 | 0.4205 | 169 | −1.857 ± 0.1085 | 0.3722 | 169 | −2.493 ± 0.09021 | 0.0188 | 169 | −2.655 ± 0.09371 | 0.9498 | 169 | 0.3002 ± 0.1025 | 0.2816 | 169 | −1.307 ± 0.05987 | 0.0696 | 169 |
| Negative | −1.796 ± 0.1205 | 195 | −1.727 ± 0.09790 | 195 | −2.138 ± 0.08013 | 195 | −2.575 ± 0.09091 | 195 | 0.1339 ± 0.1063 | 195 | −1.125 ± 0.06886 | 195 | |||||||
| Lymph Node Neck Dissection | Positive | −1.915 ± 0.07603 | 0.0044 | 422 | −1.800 ± 0.06881 | 0.8567 | 422 | −2.255 ± 0.05758 | 0.0085 | 422 | −2.555 ± 0.06305 | 0.8512 | 422 | 0.2269 ± 0.06788 | 0.6087 | 422 | −1.207 ± 0.04532 | 0.0079 | 422 |
| Negative | −1.370 ± 0.1726 | 97 | −1.771 ± 0.1533 | 97 | −1.892 ± 0.1286 | 97 | −2.519 ± 0.1193 | 97 | 0.2672 ± 0.11610 | 97 | −0.8949 ± 0.09270 | 97 | |||||||
| Angio- lymphatic Invasion | Positive | −1.765 ± 0.1361 | 0.2468 | 125 | −1.694 ± 0.1156 | 0.3401 | 125 | −2.139 ± 0.1134 | 0.0426 | 125 | −2.484 ± 0.1129 | 0.0888 | 125 | −0.01257 ± 0.1324 | 0.0161 | 125 | −1.082 ± 0.07789 | 0.0709 | 125 |
| Negative | −1.962 ± 0.1060 | 225 | −1.842 ± 0.09593 | 225 | −2.397 ± 0.07049 | 225 | −2.716 ± 0.07975 | 225 | 0.3853 ± 0.09004 | 225 | −1.283 ± 0.05889 | 225 | |||||||
| HPV p16 status | Positive | −1.060 ± 0.3101 | 0.0785 | 39 | −1.761 ± 0.2281 | 0.2259 | 39 | −0.9781 ± 0.1946 | <0.0001 | 39 | −2.332 ± 0.2203 | 0.8762 | 39 | −0.5185 ± 0.2457 | 0.0002 | 39 | −0.3564 ± 0.1596 | <0.0001 | 39 |
| Negative | −1.544 ± 0.1639 | 73 | −1.416 ± 0.1680 | 73 | −2.173 ± 0.1262 | 73 | −2.374 ± 0.1541 | 73 | 0.4726 ± 0.1646 | 73 | −1.072 ± 0.09748 | 73 | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sobocińska, J.; Nowakowska, J.; Molenda, S.; Olechnowicz, A.; Guglas, K.; Kozłowska-Masłoń, J.; Kazimierczak, U.; Machnik, M.; Oleksiewicz, U.; Teresiak, A.; et al. Zinc Finger Proteins in Head and Neck Squamous Cell Carcinomas: ZNF540 May Serve as a Biomarker. Curr. Oncol. 2022, 29, 9896-9915. https://doi.org/10.3390/curroncol29120779
Sobocińska J, Nowakowska J, Molenda S, Olechnowicz A, Guglas K, Kozłowska-Masłoń J, Kazimierczak U, Machnik M, Oleksiewicz U, Teresiak A, et al. Zinc Finger Proteins in Head and Neck Squamous Cell Carcinomas: ZNF540 May Serve as a Biomarker. Current Oncology. 2022; 29(12):9896-9915. https://doi.org/10.3390/curroncol29120779
Chicago/Turabian StyleSobocińska, Joanna, Joanna Nowakowska, Sara Molenda, Anna Olechnowicz, Kacper Guglas, Joanna Kozłowska-Masłoń, Urszula Kazimierczak, Marta Machnik, Urszula Oleksiewicz, Anna Teresiak, and et al. 2022. "Zinc Finger Proteins in Head and Neck Squamous Cell Carcinomas: ZNF540 May Serve as a Biomarker" Current Oncology 29, no. 12: 9896-9915. https://doi.org/10.3390/curroncol29120779