Low Water Activity Induces the Production of Bioactive Metabolites in Halophilic and Halotolerant Fungi



Abstract
:1. Introduction
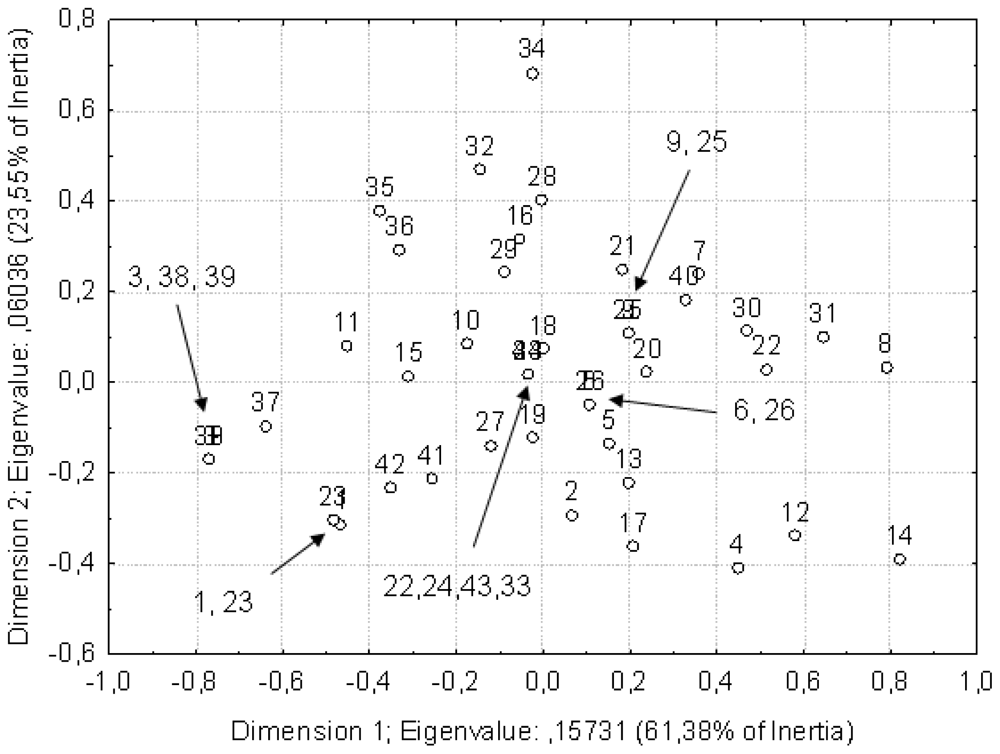
2. Results and Discussion
Biological Activities of the Tested Fungal Strains
3. Experimental Section
3.1. Fungal Sources
3.2. Cultivation of Fungi
3.3. Preparation of Extracts
3.4. Estimation of Antimicrobial Activity
3.5. Estimation of Hemolytic Activity
3.6. Inhibition of Acetylcholinesterase
3.7. Multivariate Statistical Analyses
4. Conclusions
Acknowledgements
- Samples Availability: Available from the authors.
References
- Gunde-Cimerman, N; Zalar, P; de Hoog, S; Plemenitas, A. Hypersaline waters in salterns-natural ecological niches for halophilic black yeasts. FEMS Microbiol Ecol 2000, 32, 235–240. [Google Scholar]
- Samson, RA; Hoekstra, ES; Frisvad, JC. Introduction to Food- and Airborne Fungi, 7th ed; Centraalbureau voor Schimmelcultures: Utrecht, The Netherlands, 2004; p. 389. [Google Scholar]
- Brock, TD. Ecology of saline lakes. Strategies of Microbial Life in Extreme Environments: Report of the Dahlem Workshop on Strategy of Life in Extreme Environments, Berlin, 1978, November 20–24, Shilo, M, Hirsch, P, Eds.; Verlag Chemie: Weinheim, Germany, 1979; p. 513. [Google Scholar]
- Ramos-Cormenzana, AR. Rodriguez-Valera, F, Ed.; Plenum Press: New York NY, USA, 1991; p. 402.
- Ventosa, A; Nieto, JJ; Oren, A. Biology of Moderately Halophilic Aerobic Bacteria. Microbiol Mol Biol Rev 1998, 62, 504–544. [Google Scholar]
- Oren, A. A hundred years of Dunaliella research: 1905–2005. Saline Syst 2005, 1, 2. [Google Scholar]
- Javor, BJ. Hypersaline Environments: Microbiology and Biogeochemistry; Springer: Berlin, Germany, 1989; p. 195. [Google Scholar]
- Zalar, P; de Hoog, GS; Schroers, H-J; Crous, PW; Groenewald, JZ; Gunde-Cimerman, N. Phylogeny and ecology of the ubiquitous saprobe Cladosporium sphaerospermum, with descriptions of seven new species from hypersaline environments. Stud Mycol 2007, 58, 157–183. [Google Scholar]
- Zalar, P; Sybren de Hoog, G; Schroers, H-J; Frank, JM; Gunde-Cimerman, N. Taxonomy and phylogeny of the xerophilic genus Wallemia (Wallemiomycetes and Wallemiales, cl. et ord. nov.). Antonie Van Leeuwenhoek 2005, 87, 311–328. [Google Scholar]
- Zalar, P; Frisvad, JC; Gunde-Cimerman, N; Varga, J; Samson, RA. Four new species of Emericella from the Mediterranean region of Europe. Mycologia 2008, 100, 779–795. [Google Scholar]
- Butinar, L; Santos, S; Spencer-Martins, I; Oren, A; Gunde-Cimerman, N. Yeast diversity in hypersaline habitats. FEMS Microbiol Lett 2005, 244, 229–234. [Google Scholar]
- Zalar, P; de Hoog, GS; Gunde-Cimerman, N. Ecology of halotolerant dothideaceous black yeasts. Stud Mycol 1999, 43, 38–48. [Google Scholar]
- Zalar, P; Gostincar, C; de Hoog, GS; Ursic, V; Sudhadham, M; Gunde-Cimerman, N. Redefinition of Aureobasidium pullulans and its varieties. Stud Mycol 2008, 61, 21–38. [Google Scholar]
- Butinar, L; Spencer-Martins, I; Gunde-Cimerman, N. Yeasts in high Arctic glaciers: the discovery of a new habitat for eukaryotic microorganisms. Antonie Van Leeuwenhoek 2007, 91, 277–289. [Google Scholar]
- Northolt, MD; Frisvad, JC; Samson, RA. Occurrence of food-borne fungi and factors for growth. In Introduction to Food-Borne Fungi; Samson, RA, Hoekstra, ES, Frisvad, JC, Filtenborg, O, Eds.; Centraalbureau voor schimmelcultures: Baarn, The Netherlands, 1995; pp. 243–250. [Google Scholar]
- Oren, A. Halophilic Microorganisms and Their Environments; Kluwer Academic: Dordrecht, The Netherlands, 2002; p. 575. [Google Scholar]
- Plemenitas, A; Vaupotic, T; Lenassi, M; Kogej, T; Gunde-Cimerman, N. Adaptation of extremely halotolerant black yeast Hortaea werneckii to increased osmolarity: A molecular perspective at a glance. Stud Mycol 2008, 61, 67–75. [Google Scholar]
- Gunde-Cimerman, N; Sonjak, S; Zalar, P; Frisvad, JC; Diderichsen, B; Plemenitas, A. Extremophilic fungi in arctic ice: a relationship between adaptation to low temperature and water activity. Phys Chem Earth 2003, 28, 1273–1278. [Google Scholar]
- Sonjak, S; Ursic, V; Frisvad, JC; Gunde-Cimerman, N. Penicillium svalbardense, a new species from Arctic glacial ice. Antonie Van Leeuwenhoek 2007, 92, 43–51. [Google Scholar]
- Ainsworth & Bisby’s Dictionary of the Fungi, 9th ed; Kirk, PM; Cannon, PF; David, JC; Stalpers, JS. C.A.B. International: Oxon, UK, 2001; p. 771. [Google Scholar]
- Butinar, L; Sonjak, S; Zalar, P; Plemenitaš, A; Gunde-Cimerman, N. Melanized halophilic fungi are eukaryotic members of microbial communities in hypersaline waters of solar salterns. Bot Mar 2005, 48, 73–79. [Google Scholar]
- Kralj Kuncic, M; Kogej, T; Drobne, D; Gunde-Cimerman, N. The morphological response of halophilic fungal genus Wallemia to high salinity. Appl Environ Microbiol 2010, 76, 329–337. [Google Scholar]
- Gunde-Cimerman, N; Ramos, J; Plemenitas, A. Halotolerant and halophilic fungi. Mycol Res 2009, 113, 1231–1241. [Google Scholar]
- Singh, BK. Exploring microbial diversity for biotechnology: The way forward. Trends Biotechnol 2010, 28, 111–116. [Google Scholar]
- Tudzynski, B. Gibberellin biosynthesis in fungi: Genes, enzymes, evolution, and impact on biotechnology. Appl Microbiol Biotechnol 2005, 66, 597–611. [Google Scholar]
- Ghorai, S; Banik, SP; Verma, D; Chowdhury, S; Mukherjee, S; Khowala, S. Fungal biotechnology in food and feed processing. Food Res Int 2009, 42, 577–587. [Google Scholar]
- Bennett, JW; Klich, M; Moselio, S. Mycotoxins. In Encyclopedia of Microbiology; Academic Press: Oxford, UK, 2009; pp. 559–565. [Google Scholar]
- Evidente, A; Conti, L; Altomare, C; Bottalico, A; Sindona, G; Segre, AL; Logrieco, A. Fusapyrone and deoxyfusapyrone, two antifungal alpha-Pyrones From Fusarium semitectum. Nat Toxins 1994, 2, 4–13. [Google Scholar]
- Kobayashi, H; Sunaga, R; Furihata, K; Morisaki, N; Iwasaki, S. Isolation and structures of an antifungal antibiotic, fusarielin A, and related compounds produced by a Fusarium sp. J Antibiot 1995, 48, 42–52. [Google Scholar]
- Overy, D; Calati, K; Kahn, JN; Hsu, M-J; Martín, J; Collado, J; Roemer, T; Harris, G; Parish, CA. Isolation and structure elucidation of parnafungins C and D, isoxazolidinone-containing antifungal natural products. Bioorg Med Chem Lett 2009, 19, 1224–1227. [Google Scholar]
- Meca, G; Soriano, JM; Gaspari, A; Ritieni, A; Moretti, A; Manes, J. Antifungal effects of the bioactive compounds enniatins A, A1, B, B1. Toxicon 2010, 56, 480–485. [Google Scholar]
- Renner, MK; Jensen, PR; Fenical, W. Neomangicols: Structures and Absolute Stereochemistries of Unprecedented Halogenated Sesterterpenes from a Marine Fungus of the Genus Fusarium. J Org Chem 1998, 63, 8346–8354. [Google Scholar]
- Jiang, Z; Barret, M-O; Boyd, KG; Adams, DR; Boyd, ASF; Burgess, JG. JM47, a cyclic tetrapeptide HC-toxin analogue from a marine Fusarium species. Phytochemistry 2002, 60, 33–38. [Google Scholar]
- Meca, G; Sospedra, I; Soriano, JM; Ritieni, A; Moretti, A; Mańes, J. Antibacterial effect of the bioactive compound beauvericin produced by Fusarium proliferatum on solid medium of wheat. Toxicon 2010, 56, 349–354. [Google Scholar]
- Belofsky, GN; Jensen, PR; Fenical, W. Sansalvamide: A new cytotoxic cyclic depsipeptide produced by a marine fungus of the genus Fusarium. Tetrahedron Lett 1999, 40, 2913–2916. [Google Scholar]
- Renner, MK; Jensen, PR; Fenical, W. Mangicols: Structures and Biosynthesis of a New Class of Sesterterpene Polyols from a Marine Fungus of the Genus Fusarium. J Org Chem 2000, 65, 4843–4852. [Google Scholar]
- Cueto, M; Jensen, PR; Fenical, W. N-Methylsansalvamide, a cytotoxic cyclic depsipeptide from a marine fungus of the genus Fusarium. Phytochemistry 2000, 55, 223–226. [Google Scholar]
- Hyun, U; Lee, D-H; Lee, C; Shin, C-G. Apoptosis induced by enniatins H and MK1688 isolated from Fusarium oxysporum FB1501. Toxicon 2009, 53, 723–728. [Google Scholar]
- Jadulco, R; Proksch, P; Wray, V; Sudarsono Berg, A; Grafe, U. New Macrolides and Furan Carboxylic Acid Derivative from the Sponge-Derived Fungus Cladosporium herbarum. J Nat Prod 2001, 64, 527–530. [Google Scholar]
- Shigemori, H; Kasai, Y; Komatsu, K; Tsuda, M; Mikami, Y; Kobayashi, JI. Sporiolides A and B, New Cytotoxic Twelve-Membered Macrolides from a Marine-Derived Fungus Cladosporium Species. Mar Drugs 2004, 2, 164–169. [Google Scholar]
- Wood, GM; Mann, PJ; Lewis, DF; Reid, WJ; Moss, MO. Studies on a toxic metabolite from the mould Wallemia. Food Addit Contam 1990, 7, 69–77. [Google Scholar]
- Takashi, I; Maruta, R; Ando, K; Yoshida, M; Iwasaki, T; Kanazawa, J; Okabe, M; Tamaoki, T. UCA-1064-B, a new antitumor antibiotic isolated from Wallemia sebi: Production, isolation and structural determination. J Antibiot 1993, 46, 1312–1314. [Google Scholar]
- Chi, Z; Wang, F; Chi, Z; Yue, L; Liu, G; Zhang, T. Bioproducts from Aureobasidium pullulans, a biotechnologically important yeast. Appl Microbiol Biotechnol 2009, 82, 793–804. [Google Scholar]
- Brauers, G; Ebel, R; Edrada, R; Wray, V; Berg, A; Grafe, U; Proksch, P. Hortein, a new natural product from the fungus Hortaea werneckii associated with the sponge Aplysina aerophoba. J Nat Prod 2001, 64, 651–652. [Google Scholar]
- Belmonte, G; Pederzolli, C; Maček, P; Menestrina, G. Pore formation by the sea anemone cytolysin equinatoxin II in red blood cells and model lipid membranes. J Membr Biol 1993, 131, 11–22. [Google Scholar]
- Ellman, GL; Courtney, KD; Andres, V, Jr; Featherstone, RM. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem Pharmacol 1961, 7, 88–95. [Google Scholar]


{kind=link}
| Strain no. * | Species | Strain accession no. | Origin | Temperature (°C) | Glucose (w/v%) | NaCl (w/v%) | |||
|---|---|---|---|---|---|---|---|---|---|
| ↓Temp | ↑Temp | ↓Glc | ↑Glc | ↓NaCl | ↑NaCl | ||||
| 1 | Acremonium strictum | EXF-2277 | Hypersaline water from salterns; Sečovlje, SI | 4 | 30 | 2 | 30 | 0 | 10 |
| 2 | Alternaria arborescens | EXF-2340 | Soil from salterns covered with gypsum; Sečovlje, SI | 4 | 30 | 2 | 40 | 0 | 10 |
| 3 | Alternaria infectoria | EXF-2332 | Hypersaline water from salterns; Sečovlje, SI | 10 | 30 | 2 | 30 | 0 | 10 |
| 4 | Alternaria tenuissima | EXF-2318 | Hypersaline water from salterns; Sečovlje, SI | 4 | 30 | 2 | 40 | 0 | 10 |
| 5 | Alternaria tenuissima | EXF-2329 | Hypersaline water from salterns; Sečovlje, SI | 4 | 30 | 2 | 40 | 0 | 10 |
| 6 | Alternaria tenuissima | EXF-2338 | Hypersaline water from salterns; Sečovlje, SI | 4 | 30 | 2 | 40 | 0 | 10 |
| 7 | Aureobasidium pullulans var. pullulans | EXF-150 | Hypersaline water from salterns; Sečovlje, SI | 4 | 30 | 2 | 40 | 0 | 10 |
| 8 | Aureobasidium pullulans var. melanogenum | EXF-3382 | Deep-sea (4500 m depth); Sea of Japan, JP | 4 | 30 | 2 | 40 | 0 | 10 |
| 9 | Aureobasidium sp. | EXF-922 | Subglacial ice from sea water; Svalbard, NO | 4 | 30 | 2 | 40 | 0 | 10 |
| 10 | Candida parapsilosis | EXF-517 | Hypersaline water from salterns; Sečovlje, SI | 10 | 30 | 2 | 40 | 0 | 0 |
| 11 | Candida parapsilosis | EXF-1574 | Glacial ice; Svalbard, NO | 10 | 30 | 2 | 40 | 0 | 10 |
| 12 | Cladosporium cladosporioides | EXF-381 | Hypersaline water from salterns; Sečovlje, SI | 10 | 30 | 2 | 40 | 0 | 10 |
| 13 | Cladosporium dominicanum | EXF-732 T | Hypersaline water; lake Enriquilo, DO | 10 | 30 | 2 | 40 | 0 | 10 |
| 14 | Cladosporium fusiforme | EXF-449 T | Hypersaline water from salterns; Sečovlje, SI | 4 | 30 | 2 | 40 | 0 | 10 |
| 15 | Cladosporium halotolerans | EXF-572 T | Hypersaline water from salterns; salterns; NA | 10 | 30 | 2 | 40 | 0 | 10 |
| 16 | Cladosporium halotolerans | EXF-2513 | Glacial ice, Svalbard, NO | 10 | 30 | 2 | 40 | 0 | 10 |
| 17 | Cladosporium langeronii | CBS 189.54 NT | Man, mycosis; BR | 10 | 30 | 2 | 40 | 0 | 10 |
| 18 | Cladosporium oxysporum | EXF-2246 | Ice, Staubaier glacier, AT | 4 | 30 | 2 | 40 | 0 | 10 |
| 19 | Cladosporium psychrotolerans | EXF-391 T | Hypersaline water from salterns; Sečovlje, SI Slovenia | 4 | 30 | 2 | 40 | 0 | 10 |
| 20 | Cladosporium salinae | EXF-335 T | Hypersaline water from salterns; Sečovlje, SI | 10 | 30 | 2 | 40 | 0 | 10 |
| 21 | Cladosporium sphaerospermum | EXF-385 | Hypersaline water from salterns; Sečovlje, SI | 10 | 30 | 2 | 40 | 0 | 10 |
| 22 | Cladosporium spinulosum | EXF-334 T | Hypersaline water from salterns; Sečovlje, SI | 10 | 30 | 2 | 40 | 0 | 10 |
| 23 | Cladosporium velox | EXF-466 | Hypersaline water from salterns; Sečovlje, SI | 4 | 30 | 2 | 40 | 0 | 10 |
| 24 | Cryptococcus albidus | EXF-3363 | Glacial ice; Svalbard, NO | 4 | 30 | 2 | 40 | 0 | 10 |
| 25 | Cryptococcus liquefaciens | EXF-3409 | Glacial ice; Svalbard, NO | 4 | 30 | 2 | 40 | 0 | 10 |
| 26 | Cryptococcus magnus | EXF-3360 | Glacial ice; Svalbard, NO | 4 | 30 | 2 | 30 | 0 | 10 |
| 27 | Fusarium aff. equiseti | EXF-2275 | Hypersaline water from salterns; Sečovlje, SI | 10 | 30 | 2 | 40 | 0 | 10 |
| 28 | Fusarium graminearum | EXF-2254 | Hypersaline water from salterns; Sečovlje, SI | 4 | 30 | 2 | 40 | 0 | 10 |
| 29 | Fusarium verticilloides | EXF-2276 | Hypersaline water from salterns; Sečovlje, SI | 10 | 30 | 2 | 40 | 0 | 10 |
| 30 | Hortaea werneckii | EXF-225 | Hypersaline water from salterns; Sečovlje, SI | 10 | 30 | 2 | 40 | 0 | 10 |
| 31 | Phaeotheca triangularis | EXF-206 | Hypersaline water from salterns; Sečovlje, SI | 4 | 30 | 2 | 40 | 0 | 10 |
| 32 | Pichia guilliermondii | EXF-518 | Hypersaline water from salterns; Sečovlje, SI | 4 | 30 | 2 | 40 | 0 | 10 |
| 33 | Pichia guilliermondii | EXF-1496 | Seawater; Svalbard, NO | 4 | 30 | 2 | 40 | 0 | 10 |
| 34 | Rhodosporidium babjevae | EXF-513 | Hypersaline water from salterns; Sečovlje, SI | 4 | 30 | 2 | 40 | 0 | 10 |
| 35 | Rhodosporidium diobovatum | EXF-3361 | Glacial ice; Svalbard, NO | 4 | 30 | 2 | 40 | 0 | 10 |
| 36 | Rhodotorula mucilaginosa | EXF-1630 | Glacial ice; Svalbard, NO | 4 | 30 | 2 | 40 | 0 | 10 |
| 37 | Saccharomyces cerevisiae | EXF-531 | Unknown | 10 | 30 | 2 | 40 | 0 | |
| 38 | Trichosporon mucoides | EXF-1444 | Hypersaline water from salterns; Eilat, IL | 10 | 30 | 2 | 30 | 0 | 10 |
| 39 | Trichosporon mucoides | EXF-3366 | Glacial ice; Svalbard, NO | 10 | 30 | 2 | 40 | 0 | 10 |
| 40 | Trimmatostroma salinum | EXF-295 T | Hypersaline water from salterns; Sečovlje, SI | 10 | 30 | 2 | 40 | 0 | 10 |
| 41 | Wallemia ichthyophaga | EXF-994 NT | Hypersaline water; Sečovlje salterns, SI | 10 | 30 | 55 | 55 | 10 | 17 |
| 42 | Wallemia muriae | EXF-951 NT | Hypersaline water; Sečovlje salterns, SI | 10 | 30 | 40 | 40 | 5 | 17 |
| 43 | Wallemia sebi | CBS 818.96 NT (EXF-958) | Sunflower seed; SE | 22 | 30 | 2 | 40 | 0 | 10 |
| Species | Strain | Acetone extracts | Methanolic extracts | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Control | 10% NaCl | Low T | 40% glucose | Control | 10% NaCl | Low T | 40% glucose | ||
| Acremonium strictum | EXF-2277 | − | − | − | − | − | − | − | |
| Alternaria arborescens | EXF-2340 | − | − | − | − | − | − | − | + |
| Alternaria infectoria | EXF-2332 | − | − | − | − | − | − | − | − |
| Alternaria tenuissima | EXF-2318 | − | − | − | +++ | − | − | ++ | + |
| Alternaria tenuissima | EXF-2329 | ++ | − | − | − | − | − | +++ | ++ |
| Alternaria tenuissima | EXF-2338 | − | − | +++ | − | − | − | +++ | ++ |
| Aureobasidium pullulans var. pullulans | EXF-150 | +++ | − | − | ++ | ++ | − | +++ | ++ |
| Aureobasidium pullulans var. melanogenum | EXF-3382 | +++ | +++ | − | − | ++ | − | ++ | ++ |
| Aureobasidium sp. | EXF-922 | − | +++ | − | + | − | +++ | +++ | ++ |
| Candida parapsilosis | EXF-517 | − | − | +++ | ++ | − | − | +++ | − |
| Candida parapsilosis | EXF-1574 | − | − | +++ | − | − | − | +++ | + |
| Cladosporium cladosporioides | EXF-381 | +++ | − | − | − | ++ | − | + | ++ |
| Cladosporium dominicanum | EXF-732 | − | − | + | − | ++ | − | ++ | ++ |
| Cladosporium fusiforme | EXF-449 | +++ | − | − | + | ++ | − | +++ | − |
| Cladosporium halotolerans | EXF-572 | − | − | +++ | − | + | − | +++ | − |
| Cladosporium halotolerans | EXF-2513 | ++ | ND | − | ND | ND | +++ | ++ | |
| Cladosporium langeronii | CBS 189.54 | + | − | − | − | − | − | ++ | ++ |
| Cladosporium oxysporum | EXF-2246 | − | − | − | ++ | − | − | +++ | ++ |
| Cladosporium psychrotolerans | EXF-391 | + | − | − | − | − | − | +++ | +++ |
| Cladosporium salinae | EXF-335 | +++ | − | − | − | ++ | − | +++ | ++ |
| Cladosporium sphaerospermum | EXF-385 | +++ | − | +++ | − | ++ | − | +++ | ++ |
| Cladosporium spinulosum | EXF-334 | ++ | − | − | − | + | − | − | ++ |
| Cladosporium velox | EXF-466 | − | − | − | − | − | − | − | − |
| Cryptococcus albidus | EXF-3363 | − | − | + | − | − | +++ | +++ | ++ |
| Cryptococcus liquefaciens | EXF-3409 | − | − | +++ | − | − | − | +++ | +++ |
| Cryptococcus magnus | EXF-3360 | ++ | − | − | − | +++ | − | +++ | − |
| Fusarium aff. equiseti | EXF-2275 | − | − | − | − | − | − | +++ | ++ |
| Fusarium graminearum | EXF-2254 | − | − | +++ | +++ | − | − | +++ | +++ |
| Fusarium verticilloides | EXF-2276 | − | − | − | − | − | − | +++ | +++ |
| Hortea werneckii | EXF-225 | − | +++ | − | − | − | ++ | +++ | +++ |
| Phaeotheca triangularis | EXF-206 | +++ | +++ | − | − | ++ | ++ | ++ | − |
| Pichia guilliermondii | EXF-518 | +++ | +++ | +++ | − | + | − | +++ | − |
| Pichia guilliermondii | EXF-1496 | − | − | +++ | − | − | − | − | +++ |
| Rhodosporidium babjevae | EXF-513 | +++ | +++ | +++ | +++ | ++ | − | ++ | ++ |
| Rhodosporidium diobovatum | EXF-3361 | − | +++ | +++ | − | − | ++ | +++ | − |
| Rhodotorula mucilaginosa | EXF-1630 | − | − | ++ | ++ | − | − | +++ | +++ |
| Saccharomyces cerevisiae | EXF-531 | − | − | − | − | − | − | ++ | − |
| Trichosporon mucoides | EXF-1444 | − | − | − | − | − | − | − | − |
| Trichosporon mucoides | EXF-3366 | − | − | − | − | − | − | − | − |
| Trimmatostroma salinum | EXF-295 | − | +++ | − | − | − | +++ | +++ | ++ |
| Wallemia ichthyophaga | EXF-994 | − | − | − | +++ | − | − | − | + |
| Wallemia muriae | EXF-951 | − | − | − | − | − | − | ++ | − |
| Wallemia sebi | EXF-958 | − | − | +++ | − | − | − | +++ | − |
| Species | Strain | Acetone extracts | Methanolic extracts | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Control | 10% NaCl | Low T | 40% glucose | Control | 10% NaCl | Low T | 40% glucose | ||
| Acremonium strictum | EXF-2277 | − | − | − | + | − | − | + | − |
| Alternaria aborescens | EXF-2340 | ++ | ++ | − | − | + | − | + | + |
| Alternaria infectoria | EXF-2332 | − | − | − | − | − | − | − | − |
| Alternaria tenuissima | EXF-2329 | + | + | − | − | + | + | − | − |
| Alternaria tenuissima | EXF-2318 | + | + | − | + | + | + | − | + |
| Alternaria tenuissima | EXF-2338 | − | + | − | − | + | + | − | + |
| Aureobasidium pullulans var. pullulans | EXF-150 | ++ | + | − | − | + | + | − | − |
| Aureobasidium pullulans var. melanogenum | EXF-3382 | + | + | ++ | − | + | + | + | + |
| Aureobasidium sp. | EXF-922 | − | + | − | − | − | + | − | + |
| Candida parapsilosis | EXF-517 | − | + | − | + | − | − | − | − |
| Candida parapsilosis | EXF-1574 | − | − | − | − | − | − | − | − |
| Cladosporium cladosporioides | EXF-381 | + | + | − | + | + | + | − | + |
| Cladosporium dominicanum | EXF-732 | + | − | − | − | + | − | − | + |
| Cladosporium fusiforme | EXF-449 | + | + | + | + | + | + | + | + |
| Cladosporium halotolerans | EXF-572 | − | − | − | − | − | − | − | + |
| Cladosporium halotolerans | EXF-2513 | − | − | − | − | − | − | − | + |
| Cladosporium langeronii | CBS 189.54 | + | + | − | − | + | − | − | + |
| Cladosporium oxysporum | EXF-2246 | ++ | − | − | − | − | + | − | + |
| Cladosporium psychrotolerans | EXF-391 | − | + | + | − | + | − | − | − |
| Cladosporium salinae | EXF-335 | + | + | + | − | + | − | − | − |
| Cladosporium sphaerospermum | EXF-385 | + | + | − | − | + | − | − | − |
| Cladosporium spinulosum | EXF-334 | + | ++ | +++ | + | − | + | +++ | + |
| Cladosporium velox | EXF-466 | − | − | − | − | − | − | + | + |
| Cryptococcus albidus | EXF-3363 | − | − | − | − | − | + | + | − |
| Cryptococcus liquefaciens | EXF-3409 | + | ++ | + | − | − | + | + | − |
| Cryptococcus magnus | EXF-3360 | + | − | + | − | + | − | − | + |
| Fusarium aff. equiseti | EXF-2275 | + | − | − | − | + | − | + | − |
| Fusarium graminearum | EXF-2254 | − | − | ++ | + | − | − | + | − |
| Fusarium verticilloides | EXF-2276 | ++ | − | − | + | + | − | +++ | − |
| Hortea werneckii | EXF-225 | + | ++ | − | − | + | + | + | + |
| Phaeotheca triangularis | EXF-206 | + | ++ | − | − | + | + | + | + |
| Pichia guilliermondii | EXF-518 | ++ | − | − | − | − | − | − | − |
| Pichia guilliermondii | EXF-1496 | + | ++ | − | − | + | + | − | − |
| Rhodosporidium babjevae | EXF-513 | − | − | − | − | − | − | − | − |
| Rhodosporidium diobovatum | EXF-3361 | − | − | − | − | − | − | − | − |
| Rhodotorula mucilaginosa | EXF-1630 | − | − | − | − | − | − | − | − |
| Saccharomyces cerevisiae | EXF-531 | − | − | − | − | − | − | − | − |
| Trichosporon mucoides | EXF-1444 | − | − | − | − | − | − | − | − |
| Trichosporon mucoides | EXF-3366 | − | − | − | − | − | − | − | − |
| Trimmatostroma salinum | EXF-295 | ++ | + | − | + | − | + | + | − |
| Wallemia ichthyophaga | EXF-994 | − | + | − | + | − | − | − | − |
| Wallemia muriae | EXF-951 | − | − | + | + | − | − | − | − |
| Wallemia sebi | EXF-958 | + | ++ | − | − | + | − | + | − |
© 2011 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sepcic, K.; Zalar, P.; Gunde-Cimerman, N. Low Water Activity Induces the Production of Bioactive Metabolites in Halophilic and Halotolerant Fungi. Mar. Drugs 2011, 9, 43-58. https://doi.org/10.3390/md9010043
Sepcic K, Zalar P, Gunde-Cimerman N. Low Water Activity Induces the Production of Bioactive Metabolites in Halophilic and Halotolerant Fungi. Marine Drugs. 2011; 9(1):43-58. https://doi.org/10.3390/md9010043
Chicago/Turabian StyleSepcic, Kristina, Polona Zalar, and Nina Gunde-Cimerman. 2011. "Low Water Activity Induces the Production of Bioactive Metabolites in Halophilic and Halotolerant Fungi" Marine Drugs 9, no. 1: 43-58. https://doi.org/10.3390/md9010043