
Anti-Inflammatory Activity of Cyanobacteria Pigment Extracts: Physiological Free Radical Scavenging and Modulation of iNOS and LOX Activity
 , , and
, , and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Identification of the Strains
2.2. Chemical Characterization
2.2.1. Carotenoid and Chlorophylls
2.2.2. Phycobiliproteins (PBPs)
2.3. Biological Activities
2.3.1. Antioxidant Potential
Superoxide Anion Radical Scavenging
Nitric Oxide Radical (•NO) Scavenging
2.3.2. Anti-Inflammatory Potential
Lipoxygenase Inhibition
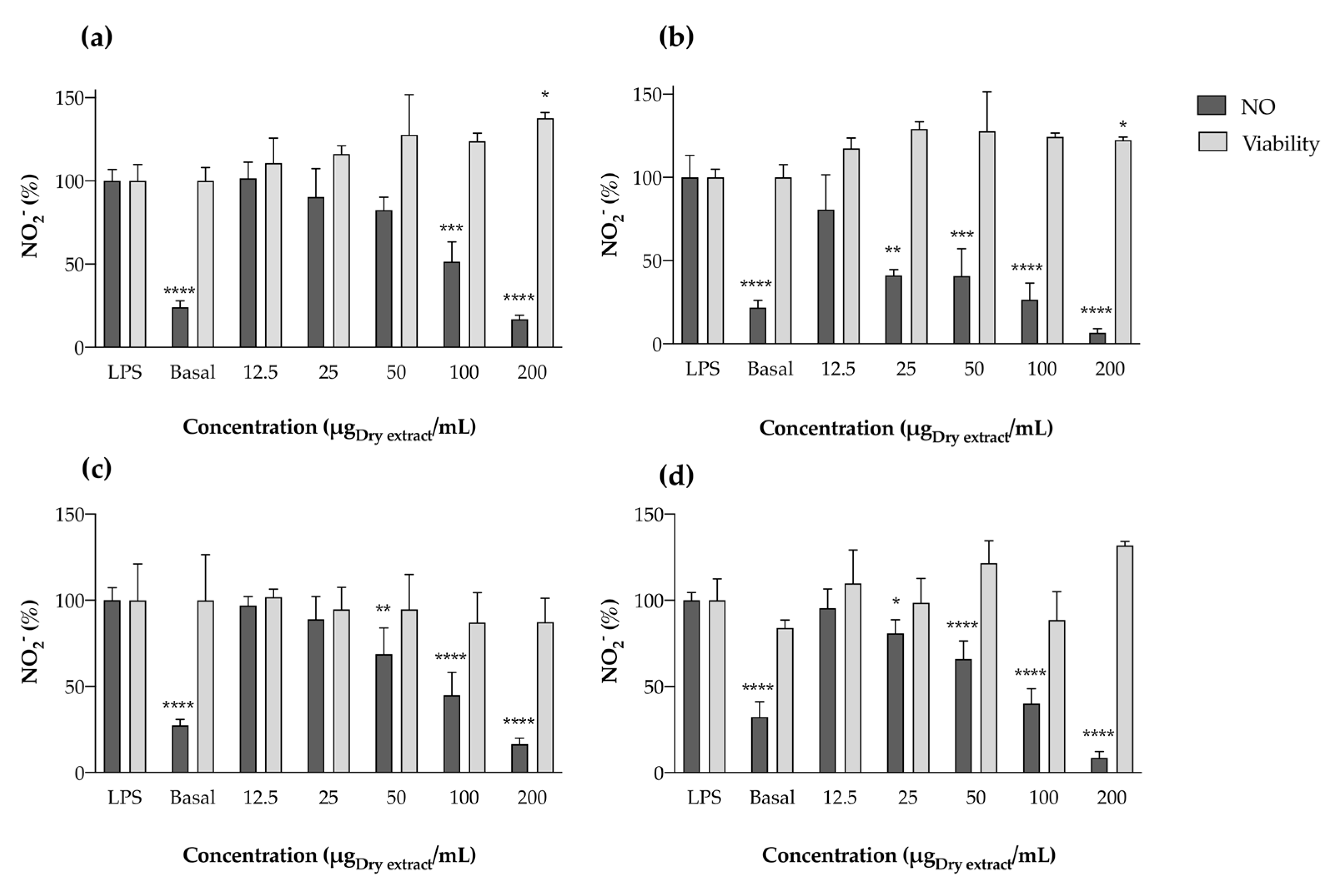
Effect in LPS-Stimulated RAW 264.7 Cells
3. Materials and Methods
3.1. Cyanobacteria Strains
Identification of the Strains
3.2. Culture Conditions and Biomass Collection
3.3. Extracts Preparation
3.4. Chemical Analysis
3.4.1. Carotenoid and Chlorophyll Profiling by HPLC-PDA
3.4.2. PBP Profiles
3.5. Biological Activities
3.5.1. Antioxidant Potential
O2•− Scavenging
•NO Scavenging
3.5.2. Anti-Inflammatory Potential
Enzymatic Assays
- (a)
- LOX Inhibition
Cell Assays
- (a)
- Cell Maintenance
- (b)
- Toxicity to RAW 264.7 Cells
- (c)
- NO Release by RAW 264.7 Cells
- (d)
- Determination of L-Citrulline Levels
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rea, I.M.; Gibson, D.S.; McGilligan, V.; McNerlan, S.E.; Alexander, H.D.; Ross, O.A. Age and age-related diseases: Role of inflammation triggers and cytokines. Front. Immunol. 2018, 9, 586. [Google Scholar] [CrossRef]
- Murakami, M.; Hirano, T. The molecular mechanisms of chronic inflammation development. Front. Immunol. 2012, 3, 323. [Google Scholar] [CrossRef]
- Anavi, S.; Tirosh, O. iNOS as a metabolic enzyme under stress conditions. Free Radic. Biol. Med. 2020, 146, 16–35. [Google Scholar] [CrossRef]
- Korhonen, R.; Lahti, A.; Kankaanranta, H.; Moilanen, E. Nitric oxide production and signaling in inflammation. Curr. Drug Targets-Inflamm. Allergy 2005, 4, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Koeberle, A.; Werz, O. Multi-target approach for natural products in inflammation. Drug Discov. Today 2014, 19, 1871–1882. [Google Scholar] [CrossRef]
- Singh, R.K.; Tiwari, S.P.; Rai, A.K.; Mohapatra, T.M. Cyanobacteria: An emerging source for drug discovery. J. Antibiot. 2011, 64, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Pagels, F.; Guedes, A.C.; Vasconcelos, V.; Lopes, G. Anti-inflammatory compounds from cyanobacteria. In The Pharmacological Potential of Cyanobacteria; Elsevier: Amsterdam, The Netherlands, 2022; pp. 81–105. [Google Scholar]
- Heo, S.-J.; Yoon, W.-J.; Kim, K.-N.; Ahn, G.-N.; Kang, S.-M.; Kang, D.-H.; Oh, C.; Jung, W.-K.; Jeon, Y.-J. Evaluation of anti-inflammatory effect of fucoxanthin isolated from brown algae in lipopolysaccharide-stimulated RAW 264.7 macrophages. Food Chem. Toxicol. 2010, 48, 2045–2051. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.-J.; Lin, J.-T.; Chen, Y.-C.; Liu, S.-C.; Lu, F.-J.; Chang, T.-J.; Wang, M.; Lin, H.-W.; Chang, Y.-Y. Suppressive effect of carotenoid extract of Dunaliella salina alga on production of LPS-stimulated pro-inflammatory mediators in RAW264. 7 cells via NF-κB and JNK inactivation. J. Funct. Foods 2013, 5, 607–615. [Google Scholar] [CrossRef]
- Chini Zittelli, G.; Lauceri, R.; Faraloni, C.; Silva Benavides, A.M.; Torzillo, G. Valuable pigments from microalgae: Phycobiliproteins, primary carotenoids, and fucoxanthin. Photochem. Photobiol. Sci. 2023, 22, 1733–1789. [Google Scholar] [CrossRef]
- Slavich, G.M. Understanding inflammation, its regulation, and relevance for health: A top scientific and public priority. Brain Behav. Immun. 2015, 45, 13. [Google Scholar] [CrossRef]
- Morone, J.; Lopes, G.; Morais, J.; Neves, J.; Vasconcelos, V.; Martins, R. Cosmetic application of cyanobacteria extracts with a sustainable vision to skincare: Role in the antioxidant and antiaging process. Mar. Drugs 2022, 20, 761. [Google Scholar] [CrossRef]
- Morone, J.; Lopes, G.; Preto, M.; Vasconcelos, V.; Martins, R. Exploitation of filamentous and picoplanktonic cyanobacteria for cosmetic applications: Potential to improve skin structure and preserve dermal matrix components. Mar. Drugs 2020, 18, 486. [Google Scholar] [CrossRef]
- Bogacz-Radomska, L.; Harasym, J. β-Carotene—Properties and production methods. Food Qual. Saf. 2018, 2, 69–74. [Google Scholar] [CrossRef]
- Juturu, V.; Bowman, J.P.; Deshpande, J. Overall skin tone and skin-lightening-improving effects with oral supplementation of lutein and zeaxanthin isomers: A double-blind, placebo-controlled clinical trial. Clin. Cosmet. Investig. Dermatol. 2016, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Hashtroudi, M.S.; Shariatmadari, Z.; Riahi, H.; Ghassempour, A. Analysis of Anabaena vaginicola and Nostoc calcicola from Northern Iran, as rich sources of major carotenoids. Food Chem. 2013, 136, 1148–1153. [Google Scholar] [CrossRef] [PubMed]
- Lopes, G.; Clarinha, D.; Vasconcelos, V. Carotenoids from cyanobacteria: A biotechnological approach for the topical treatment of psoriasis. Microorganisms 2020, 8, 302. [Google Scholar] [CrossRef] [PubMed]
- Kusama, Y.; Inoue, S.; Jimbo, H.; Takaichi, S.; Sonoike, K.; Hihara, Y.; Nishiyama, Y. Zeaxanthin and echinenone protect the repair of photosystem II from inhibition by singlet oxygen in Synechocystis sp. PCC 6803. Plant Cell Physiol. 2015, 56, 906–916. [Google Scholar] [CrossRef] [PubMed]
- Palinska, K.A.; Deventer, B.; Hariri, K.; Lotocka, M. A taxonomic study on Phormidium–group (cyanobacteria) based on morphology, pigments, RAPD molecular markers and RFLP analysis of the 16S rRNA gene fragment. Fottea 2011, 11, 41–55. [Google Scholar] [CrossRef]
- Hossain, M.F.; Ratnayake, R.R.; Meerajini, K.; Wasantha Kumara, K. Antioxidant properties in some selected cyanobacteria isolated from fresh water bodies of Sri Lanka. Food Sci. Nutr. 2016, 4, 753–758. [Google Scholar] [CrossRef]
- Favas, R.; Morone, J.; Martins, R.; Vasconcelos, V.; Lopes, G. Cyanobacteria secondary metabolites as biotechnological ingredients in natural anti-aging cosmetics: Potential to overcome hyperpigmentation, loss of skin density and UV radiation-deleterious effects. Mar. Drugs 2022, 20, 183. [Google Scholar] [CrossRef]
- Schipper, K.; Fortunati, F.; Oostlander, P.C.; Al Muraikhi, M.; Al Jabri, H.M.S.; Wijffels, R.H.; Barbosa, M.J. Production of phycocyanin by Leptolyngbya sp. in desert environments. Algal Res. 2020, 47, 101875. [Google Scholar] [CrossRef]
- Pagels, F.; Guedes, A.C.; Amaro, H.M.; Kijjoa, A.; Vasconcelos, V. Phycobiliproteins from cyanobacteria: Chemistry and biotechnological applications. Biotechnol. Adv. 2019, 37, 422–443. [Google Scholar] [CrossRef]
- Basheva, D.; Moten, D.; Stoyanov, P.; Belkinova, D.; Mladenov, R.; Teneva, I. Content of phycoerythrin, phycocyanin, alophycocyanin and phycoerythrocyanin in some cyanobacterial strains: Applications. Eng. Life Sci. 2018, 18, 861–866. [Google Scholar] [CrossRef] [PubMed]
- Assunção, J.; Amaro, H.M.; Lopes, G.; Tavares, T.; Malcata, F.X.; Guedes, A.C. Synechocystis salina: Potential bioactivity and combined extraction of added-value metabolites. J. Appl. Phycol. 2021, 33, 3731–3746. [Google Scholar] [CrossRef]
- Assunção, J.; Amaro, H.M.; Lopes, G.; Tavares, T.; Malcata, F.X.; Guedes, A.C. Exploration of marine genus Chroococcidiopsis sp.: A valuable source for antioxidant industry? J. Appl. Phycol. 2021, 33, 2169–2187. [Google Scholar] [CrossRef]
- Mashima, R.; Okuyama, T. The role of lipoxygenases in pathophysiology; new insights and future perspectives. Redox Biol. 2015, 6, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Serpen, A.; Gökmen, V. Effects of β-carotene on soybean lipoxygenase activity: Kinetic studies. Eur. Food Res. Technol. 2007, 224, 743–748. [Google Scholar] [CrossRef]
- Jensen, G.S.; Attridge, V.L.; Beaman, J.L.; Guthrie, J.; Ehmann, A.; Benson, K.F. Antioxidant and anti-inflammatory properties of an aqueous cyanophyta extract derived from Arthrospira platensis: Contribution to bioactivities by the non-phycocyanin aqueous fraction. J. Med. Food 2015, 18, 535–541. [Google Scholar] [CrossRef]
- Fagundes, M.B.; Alvarez-Rivera, G.; Mendiola, J.A.; Bueno, M.; Sánchez-Martínez, J.D.; Wagner, R.; Jacob-Lopes, E.; Zepka, L.Q.; Ibañez, E.; Cifuentes, A. Phytosterol-rich compressed fluids extracts from Phormidium autumnale cyanobacteria with neuroprotective potential. Algal Res. 2021, 55, 102264. [Google Scholar] [CrossRef]
- da Costa, E.; Amaro, H.M.; Melo, T.; Guedes, A.C.; Domingues, M.R. Screening for polar lipids, antioxidant, and anti-inflammatory activities of Gloeothece sp. lipid extracts pursuing new phytochemicals from cyanobacteria. J. Appl. Phycol. 2020, 32, 3015–3030. [Google Scholar] [CrossRef]
- Pagels, F.; Salvaterra, D.; Amaro, H.M.; Lopes, G.; Sousa-Pinto, I.; Vasconcelos, V.; Guedes, A.C. Bioactive potential of Cyanobium sp. pigment-rich extracts. J. Appl. Phycol. 2020, 32, 3031–3040. [Google Scholar] [CrossRef]
- Reddy, C.M.; Bhat, V.B.; Kiranmai, G.; Reddy, M.N.; Reddanna, P.; Madyastha, K. Selective inhibition of cyclooxygenase-2 by C-phycocyanin, a biliprotein from Spirulina platensis. Biochem. Biophys. Res. Commun. 2000, 277, 599–603. [Google Scholar] [CrossRef]
- Ferreira, M.G. Study of Anti-Inflammatory Bioactivity of Cyanobacterial Strains Using Murine Macrophage RAW 264.7 Cells. 2016. Available online: https://repositorio-aberto.up.pt/bitstream/10216/87536/2/163123.pdf (accessed on 4 January 2024).
- Bai, S.-K.; Lee, S.-J.; Na, H.-J.; Ha, K.-S.; Han, J.-A.; Lee, H.; Kwon, Y.-G.; Chung, C.-K.; Kim, Y.-M. β-Carotene inhibits inflammatory gene expression in lipopolysaccharide-stimulated macrophages by suppressing redox-based NF-κB activation. Exp. Mol. Med. 2005, 37, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Okai, Y.; Higashi-Okai, K. Potent anti-inflammatory activity of pheophytin a derived from edible green alga, Enteromorpha prolifera (Sujiao-nori). Int. J. Immunopharmacol. 1997, 19, 355–358. [Google Scholar] [CrossRef]
- Soontornchaiboon, W.; Joo, S.S.; Kim, S.M. Anti-inflammatory effects of violaxanthin isolated from microalga Chlorella ellipsoidea in RAW 264.7 macrophages. Biol. Pharm. Bull. 2012, 35, 1137–1144. [Google Scholar] [CrossRef]
- Stewart, I.; Schluter, P.J.; Shaw, G.R. Cyanobacterial lipopolysaccharides and human health—A review. Environ. Health 2006, 5, 7. [Google Scholar] [CrossRef] [PubMed]
- Swanson-Mungerson, M.; Incrocci, R.; Subramaniam, V.; Williams, P.; Hall, M.L.; Mayer, A.M. Effects of cyanobacteria Oscillatoria sp. lipopolysaccharide on B cell activation and Toll-like receptor 4 signaling. Toxicol. Lett. 2017, 275, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Tabarzad, M.; Atabaki, V.; Hosseinabadi, T. Anti-inflammatory activity of bioactive compounds from microalgae and cyanobacteria by focusing on the mechanisms of action. Mol. Biol. Rep. 2020, 47, 6193–6205. [Google Scholar] [CrossRef]
- Tarasuntisuk, S.; Palaga, T.; Kageyama, H.; Waditee-Sirisattha, R. Mycosporine-2-glycine exerts anti-inflammatory and antioxidant effects in lipopolysaccharide (LPS)-stimulated RAW 264.7 macrophages. Arch. Biochem. Biophys. 2019, 662, 33–39. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef]
- Kotai, J. Instructions for preparation of modified nutrient solution Z8 for algae. Nor. Inst. Water Res. Oslo 1972, 11, 5. [Google Scholar]
- Bennett, A.; Bogorad, L. Complementary chromatic adaptation in a filamentous blue-green alga. J. Cell Biol. 1973, 58, 419–435. [Google Scholar] [CrossRef]
- Saluri, M.; Kaldmäe, M.; Tuvikene, R. Extraction and quantification of phycobiliproteins from the red alga Furcellaria lumbricalis. Algal Res. 2019, 37, 115–123. [Google Scholar] [CrossRef]
- Lopes, G.; Sousa, C.; Silva, L.R.; Pinto, E.; Andrade, P.B.; Bernardo, J.; Mouga, T.; Valentão, P. Can phlorotannins purified extracts constitute a novel pharmacological alternative for microbial infections with associated inflammatory conditions? PLoS ONE 2012, 7, e31145. [Google Scholar] [CrossRef]
- Fernandes, F.; Barbosa, M.; Pereira, D.M.; Sousa-Pinto, I.; Valentão, P.; Azevedo, I.C.; Andrade, P.B. Chemical profiling of edible seaweed (Ochrophyta) extracts and assessment of their in vitro effects on cell-free enzyme systems and on the viability of glutamate-injured SH-SY5Y cells. Food Chem. Toxicol. 2018, 116, 196–206. [Google Scholar] [CrossRef]
- Barbosa, M.; Lopes, G.; Ferreres, F.; Andrade, P.B.; Pereira, D.M.; Gil-Izquierdo, Á.; Valentão, P. Phlorotannin extracts from Fucales: Marine polyphenols as bioregulators engaged in inflammation-related mediators and enzymes. Algal Res. 2017, 28, 1–8. [Google Scholar] [CrossRef]
- Barbosa, M.; Fernandes, F.; Carlos, M.J.; Valentao, P.; Andrade, P.B. Adding value to marine invaders by exploring the potential of Sargassum muticum (Yendo) Fensholt phlorotannin extract on targets underlying metabolic changes in diabetes. Algal Res. 2021, 59, 102455. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Compound | RT (min) | Cephalothrix lacustris LEGE 15493 | Nodosilinea nodulosa LEGE 06104 | Leptolyngbya boryana LEGE 15486 | Leptothoe sp. LEGE 11479 |
|---|---|---|---|---|---|---|
| 1 | Chlorophyll-a derivative | 9.7 | nd | nd | nq | 2.177 ± 0.066 |
| 2 | Chlorophyll-a derivative | 10.1 | 0.018 ± 0.004 c | 1.269 ± 0.179 a | nd | 0.574 ± 0.059 b |
| 3 | Chlorophyll-a derivative | 11.0 | nd | nd | nd | 0.321 ± 0.028 |
| 4 | Lutein | 12.5 | 0.578 ± 0.033 c | 1.044 ± 0.065 b | 0.381 ± 0.018 c | 1.383 ± 0.097 a |
| 5 | Unidentified Carotenoid | 13.6 | 1.368 ± 0.060 c | 11.996 ± 0.037 a | 2.321 ± 0.013 b | 2.099 ± 0.103 b |
| 6 | Chlorophyll-a | 14.9 | 17.461 ± 0.186 a | 5.031 ± 0.162 c | 10.949 ± 0.604 b | 14.740 ± 1.533 a |
| 7 | Zeaxanthin | 15.2 | 99.553 ± 0.515 c | 123.911 ± 0.865 b | 52.680 ± 0.399 d | 192.908 ± 3.198 a |
| 8 | Chlorophyll-a derivative | 16.6 | 0.532 ± 0.039 c | 1.934 ± 0.050 b | 0.512 ± 0.047 c | 3.663 ± 0.333 a |
| 9 | Unidentified Carotenoid | 17.4 | nd | nd | 7.661 ± 0.128 | nd |
| 10 | Unidentified Carotenoid | 20 | 8.588 ± 0.050 b | 18.944 ± 0.111 a | 1.134 ± 0.030 d | 2.060 ± 0.082 c |
| 11 | Unidentified Carotenoid | 21.4 | 2.219 ± 0.050 b | 3.156 ± 0.012 a | 3.496 ± 0.174 a | 1.068 ± 0.011 c |
| 12 | Unidentified Carotenoid | 22.9 | nd | 5.834 ± <0.001 a | nd | 2.485 ± 0.087 b |
| 13 | β-Cryptoxanthin | 24.0 | 2.033 ± 0.037 b | 2.793 ± 0.188 a | 1.686 ± 0.017b | 2.577 ± 0.085 a |
| 14 | Unidentified Carotenoid | 24.7 | nd | nd | 2.261 ± 0.069 | nd |
| 15 | Echinenone | 25.3 | 1.785 ± 0.005 a | nd | 1.699 ± 0.079 b | nd |
| 16 | Unidentified Carotenoid | 26.9 | 2.969 ± 0.006 b | 2.455 ± 0.136 b | 1.374 ± 0.127 c | 3.901 ± 0.177 a |
| 17 | Unidentified Carotenoid | 28.0 | 10.324 ± 0.087 b | nd | 22.332 ± 0.168 a | nd |
| 18 | β-Carotene | 32.8 | 18.393 ± 0.250 b | 44.315 ± 1.471 a | 20.408 ± 3.198 b | 39.791 ± 8.117 a |
| 19 | β-Carotene derivative | 34.0 | 6.167 ± 0.147 c | 13.747 ± 0.046 a | 3.870 ± 0.026 d | 12.358 ± 0.252 b |
| Total carotenoids | 153.978 ± 1.391 b | 228.194 ± 2.929 a | 121.304 ± 4.447 b | 260.629 ± 12.205 a | ||
| Total chlorophylls | 18.012 ± 0.229 a | 8.234 ± 0.390 b | 11.462 ± 0.651 b | 21.476 ± 2.019 a |
| Strains | Phycobiliproteins | ||
|---|---|---|---|
| Phycocyanin | Allophycocyanin | Phycoerythrin | |
| Cephalothrix lacustris LEGE 15493 | 108.27 ± 0.54 b | 31.98 ± 0.38 b | 16.52 ± 0.61 b |
| Nodosilinea nodulosa LEGE 06104 | 30.83 ± 0.28 d | 4.96 ± 0.44 d | 4.93 ± 0.18 c |
| Leptolyngbya boryana LEGE 15486 | 204.52 ± 0.14 a | 41.61 ± 0.22 a | 14.42 ± 0.39 b |
| Leptothoe sp. LEGE 11479 | 54.02 ± 1.76 c | 22.04 ± 0.87 c | 78.49 ± 3.01 a |
| Strains | IC25 | IC50 |
|---|---|---|
| Cephalothrix lacustris LEGE 15493 | 87.55 ± 1.07 a,b | 178.06 ± 9.82 |
| Nodosilinea nodulosa LEGE 06104 | 66.72 ± 29.60 a,b | 233.56 ± 176.09 |
| Leptolyngbya boryana LEGE 15486 | 25.19 ± 3.06 a | 49.24 ± 4.45 |
| Leptothoe sp. LEGE 11479 | 111.08 ± 30.43 b | nd |
| Strains | Aqueous Extracts | Acetonic Extracts | ||
|---|---|---|---|---|
| IC25 | IC5O | IC25 | IC5O | |
| Cephalothrix lacustris LEGE 15493 | 15.91 ± 10.36 a | 36.68 ± 4.57 | 57.56 ± 2.02 a | 329.627 ± 7.54 a,b |
| Nodosilinea nodulosa LEGE 06104 | 1617.62 ± 186.65 c | nd | 75.16 ± 13.52 a,b | 186.50 ± 47.90 a,b |
| Leptolyngbya boryana LEGE 15486 | 835.31 ± 286.74 b | nd | 95.19 ± 5.24 b | 232.57 ± 36.46 b |
| Leptothoe sp. LEGE 11479 | 472.99 ± 104.64 a,b | 1526.9 * | 46.63 ± 11.40 a | 129.47 ± 23.87 a |
| Strains | Aqueous Extracts | Acetonic Extracts | ||
|---|---|---|---|---|
| IC25 | IC5O | IC25 | IC5O | |
| Cephalothrix lacustris LEGE 15493 | 87.54 ± 26.48 | 375.51 ± 32.97 b | 96.77 ± 44.96 | 251.69 ± 29.54 b |
| Nodosilinea nodulosa LEGE 06104 | 37.43 ± 15.13 | 206.02 ± 70.10 a | 42.44 ± 22.75 | 94.81 ± 40.83 a |
| Leptolyngbya boryana LEGE 15486 | 68.94 ± 23.10 | 142.75 ± 21.91 a | 34.36 ± 13.94 | 196.37 ± 0.21 b |
| Leptothoe sp. LEGE 11479 | 46.98 ± 27.78 | 484.04 ± 27.44 b | 25.92 ± 3.52 | 176.00 ± 3.49 a,b |
| Standards | Calibration Curve | r2 |
|---|---|---|
| Lutein | y = 93629588x + 13548 | 0.9999 |
| Chlorophyll-a | y = 83010783x + 144987 | 0.9985 |
| Zeaxanthin | y = 80009404x + 19635 | 0.9944 |
| β-Cryptoxanthin | y = 28522518x + 8558 | 0.9992 |
| Echinenone | y = 87231499x + 43970 | 0.9996 |
| β-Carotene | y = 29605275x + 36236 | 0.9982 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, L.; Morone, J.; Hentschke, G.S.; Vasconcelos, V.; Lopes, G. Anti-Inflammatory Activity of Cyanobacteria Pigment Extracts: Physiological Free Radical Scavenging and Modulation of iNOS and LOX Activity. Mar. Drugs 2024, 22, 131. https://doi.org/10.3390/md22030131
Rodrigues L, Morone J, Hentschke GS, Vasconcelos V, Lopes G. Anti-Inflammatory Activity of Cyanobacteria Pigment Extracts: Physiological Free Radical Scavenging and Modulation of iNOS and LOX Activity. Marine Drugs. 2024; 22(3):131. https://doi.org/10.3390/md22030131
Chicago/Turabian StyleRodrigues, Lécia, Janaína Morone, Guilherme Scotta Hentschke, Vitor Vasconcelos, and Graciliana Lopes. 2024. "Anti-Inflammatory Activity of Cyanobacteria Pigment Extracts: Physiological Free Radical Scavenging and Modulation of iNOS and LOX Activity" Marine Drugs 22, no. 3: 131. https://doi.org/10.3390/md22030131