
New Forms of Neuroactive Phospholipids for DHA Enrichment in Brain

Abstract
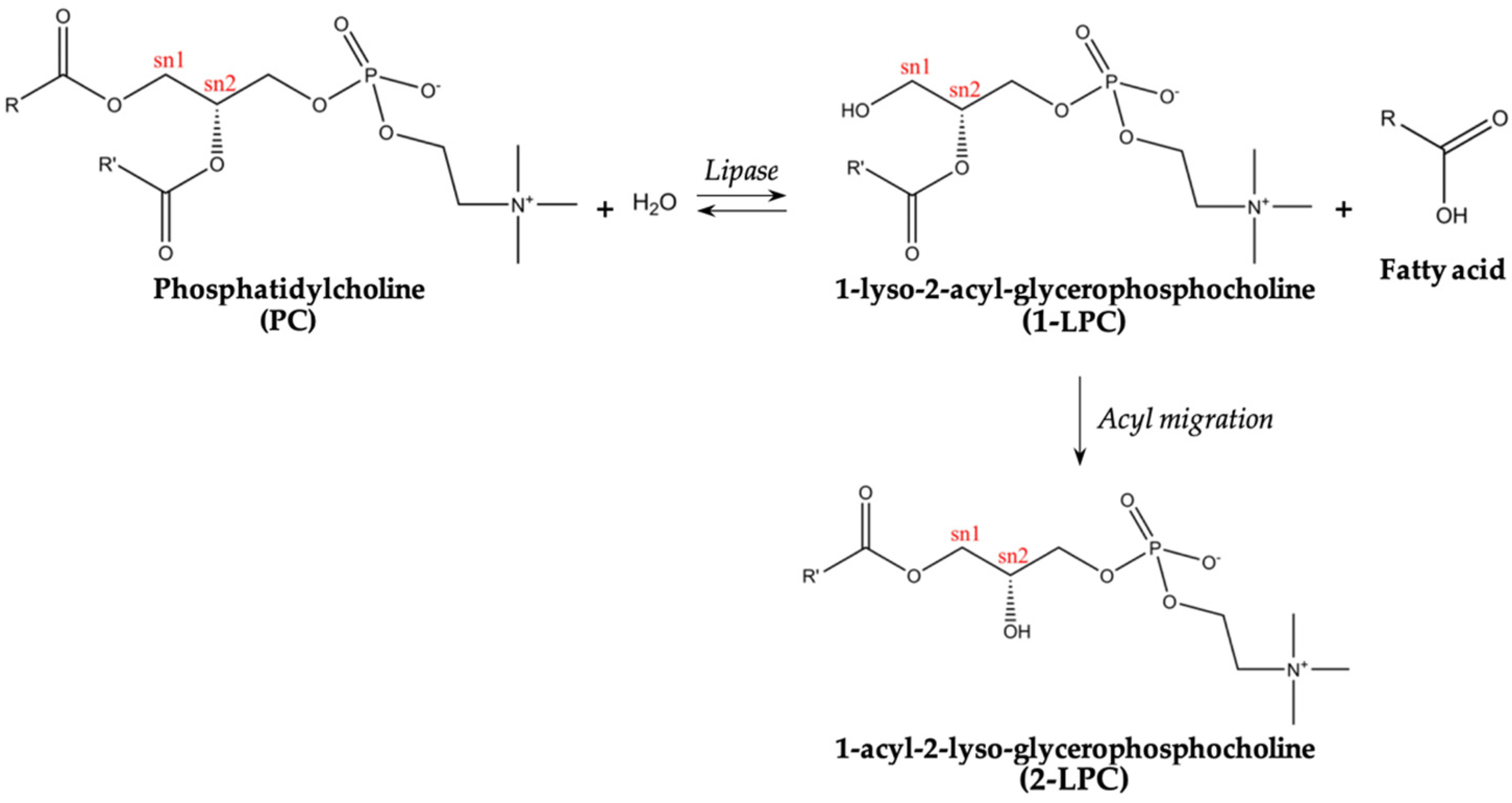
:1. Introduction
2. Results and Discussion
2.1. Characterization of Atlantic Mackerel Lipid Fraction
2.1.1. Total Lipids Content
2.1.2. Fatty Acid Profile
2.1.3. Total Lipid Classes
2.1.4. Fatty Acid Profiles of Phospholipids and Non-Polar Lipid Classes
2.2. Production and Characterization of the Fish Oil Enriched in LPC-DHA
3. Materials and Methods
3.1. Material and Reagents
3.2. DHA-Rich Phospholipids Fish Oil Production
3.3. Total Lipids Content
3.4. Lipid Classes Profile
3.4.1. Total Lipid Classes
3.4.2. Polar Lipid Classes
3.5. Fatty Acid Profile
3.6. Fatty Acid Profile of Lipid Classes
3.6.1. Fish Total Lipid Classes
3.6.2. DHA-Rich Fish Oil Polar Lipid Classes
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wong, L.R.; Wong, P.; Ho, P.C. Metabolic Profiling of Female Tg2576 Mouse Brains Provides Novel Evidence Supporting Intranasal Low-Dose Pioglitazone for Long-Term Treatment at an Early Stage of Alzheimer’s Disease. Biomedicines 2020, 8, 589. [Google Scholar] [CrossRef]
- Patrick, R.P. Role of phosphatidylcholine-DHA in preventing APOE4-associated Alzheimer’s disease. FASEB J. 2019, 33, 1554–1564. [Google Scholar] [CrossRef]
- Reitz, C.; Mayeux, R. Alzheimer disease: Epidemiology, diagnostic criteria, risk factors and biomarkers. Biochem. Pharmacol. 2014, 88, 640–651. [Google Scholar] [CrossRef]
- Gu, Y.; Schupf, N.; Cosentino, S.A.; Luchsinger, J.A.; Scarmeas, N. Nutrient intake and plasma β-amyloid. Neurology 2012, 78, 1832–1840. [Google Scholar] [CrossRef]
- Semba, R.D. Perspective: The Potential Role of Circulating Lysophosphatidylcholine in Neuroprotection against Alzheimer Disease. Adv. Nutr. 2020, 11, 760–772. [Google Scholar] [CrossRef] [PubMed]
- Cunnane, S.C.; Plourde, M.; Pifferi, F.; Begin, M.; Feart, C.; Barberger-Gateau, P. Fish, docosahexaenoic acid and Alzheimer’s disease. Prog. Lipid Res. 2009, 48, 239–256. [Google Scholar] [CrossRef]
- Sugasini, D.; Thomas, R.; Yalagala, P.C.R.; Tai, L.M.; Subbaiah, P.V. Dietary docosahexaenoic acid (DHA) as lysophosphatidylcholine, but not as free acid, enriches brain DHA and improves memory in adult mice. Sci. Rep. 2017, 7, 11263. [Google Scholar] [CrossRef] [PubMed]
- Yin, F. Lipid metabolism and Alzheimer’s disease: Clinical evidence, mechanistic link and therapeutic promise. FEBS J. 2022, 290, 1420–1453. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, C.; Afonso, C.; Bandarra, N.M. Dietary DHA and health: Cognitive function ageing. Nutr. Res. Rev. 2016, 29, 281–294. [Google Scholar] [CrossRef]
- Valentini, K.J.; Pickens, C.A.; Wiesinger, J.A.; Fenton, J.I. The effect of fish oil supplementation on brain DHA and EPA content and fatty acid profile in mice. Int. J. Food Sci. Nutr. 2018, 69, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.Y.; Wu, T.; Huang, S.Y.; Huang, B.X.; Wang, H.L.; Lan, Q.Y.; Li, C.L.; Zhu, H.L.; Fang, A.P. No effect of 6-month supplementation with 300 mg/d docosahexaenoic acid on executive functions among healthy school-aged children: A randomized, double-blind, placebo-controlled trial. Eur. J. Nutr. 2021, 60, 1985–1997. [Google Scholar] [CrossRef]
- Ahmmed, M.K.; Ahmmed, F.; Tian, H.S.; Carne, A.; Bekhit, A.E. Marine omega-3 (n-3) phospholipids: A comprehensive review of their properties, sources, bioavailability, and relation to brain health. Compr. Rev. Food Sci. Food Saf. 2020, 19, 64–123. [Google Scholar] [CrossRef]
- Nguyen, L.N.; Ma, D.; Shui, G.; Wong, P.; Cazenave-Gassiot, A.; Zhang, X.; Wenk, M.R.; Goh, E.L.; Silver, D.L. Mfsd2a is a transporter for the essential omega-3 fatty acid docosahexaenoic acid. Nature 2014, 509, 503–506. [Google Scholar] [CrossRef] [PubMed]
- Bernoud, N.; Fenart, L.; Molière, P.; Dehouck, M.P.; Lagarde, M.; Cecchelli, R.; Lecerf, J. Preferential transfer of 2-docosahexaenoyl-1-lysophosphatidylcholine through an in vitro blood-brain barrier over unesterified docosahexaenoic acid. J. Neurochem. 1999, 71, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Lagarde, M.; Bernoud, N.; Thiès, F.; Brossard, N.; Lemaitre-Delaunay, D.; Croset, M.; Lecerf, J. Lysophosphatidylcholine as a preferred carrier form of docosahexaenoic acid to the brain. J. Mol. Neurosci. 2001, 16, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Thies, F.; Pillon, C.; Moliere, P.; Lagarde, M.; Lecerf, J. Preferential incorporation of sn-2 lysoPC DHA over unesterified DHA in the young rat brain. Am. J. Physiol. 1994, 267, R1273–R1279. [Google Scholar] [CrossRef] [PubMed]
- Law, S.H.; Chan, M.L.; Marathe, G.K.; Parveen, F.; Chen, C.H.; Ke, L.Y. An Updated Review of Lysophosphatidylcholine Metabolism in Human Diseases. Int. J. Mol. Sci. 2019, 20, 1149. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Zhu, W.; Chen, C.; Yan, B.; Zhu, L.; Chen, X.; Peng, C. The mechanisms of lysophosphatidylcholine in the development of diseases. Life Sci. 2020, 247, 117443. [Google Scholar] [CrossRef] [PubMed]
- Akerele, O.A.; Cheema, S.K. Fatty acyl composition of lysophosphatidylcholine is important in atherosclerosis. Med. Hypotheses 2015, 85, 754–760. [Google Scholar] [CrossRef]
- Huang, L.S.; Hung, N.D.; Sok, D.E.; Kim, M.R. Lysophosphatidylcholine containing docosahexaenoic acid at the sn-1 position is anti-inflammatory. Lipids 2010, 45, 225–236. [Google Scholar] [CrossRef]
- Hung, N.D.; Sok, D.E.; Kim, M.R. Prevention of 1-palmitoyl lysophosphatidylcholine-induced inflammation by polyunsaturated acyl lysophosphatidylcholine. Inflamm. Res. 2012, 61, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Mnasri, T.; Hérault, J.; Gauvry, L.; Loiseau, C.; Poisson, L.; Ergan, F.; Pencréac’h, G. Lipase-catalyzed production of lysophospholipids. OCL 2017, 24, D405. [Google Scholar] [CrossRef]
- Bandarra, N.M.; Batista, I.; Nunes, M.L.; Empis, J.M.; Christie, W.W. Seasonal Changes in Lipid Composition of Sardine (Sardina pilchardus). J. Food Sci. 1997, 62, 40–42. [Google Scholar] [CrossRef]
- Bandarra, N.; Batista, I.; Nunes, M.; Empis, J.M. Seasonal variation in the chemical composition of horse-mackerel (Trachurus trachurus). Eur. Food Res. Technol. 2001, 212, 535–539. [Google Scholar] [CrossRef]
- Ferreira, I.; Gomes-Bispo, A.; Lourenco, H.; Matos, J.; Afonso, C.; Cardoso, C.; Castanheira, I.; Motta, C.; Prates, J.A.M.; Bandarra, N.M. The chemical composition and lipid profile of the chub mackerel (Scomber colias) show a strong seasonal dependence: Contribution to a nutritional evaluation. Biochimie 2020, 178, 181–189. [Google Scholar] [CrossRef]
- Costa, A.M.; Gordo, L.S.; Martins, M.M. Growth and distribution of mackerel Scomber scombrus Linnaeus, 1758 from the west coast of Portugal. Cah. Biol. Mar. 2017, 58, 409–421. [Google Scholar] [CrossRef]
- Nøstbakken, O.J.; Rasinger, J.D.; Hannisdal, R.; Sanden, M.; Froyland, L.; Duinker, A.; Frantzen, S.; Dahl, L.M.; Lundebye, A.K.; Madsen, L. Levels of omega 3 fatty acids, vitamin D, dioxins and dioxin-like PCBs in oily fish; a new perspective on the reporting of nutrient and contaminant data for risk-benefit assessments of oily seafood. Environ. Int. 2021, 147, 106322. [Google Scholar] [CrossRef]
- Tang, Y.; Zhao, R.; Pu, Q.; Jiang, S.; Yu, F.; Yang, Z.; Han, T. Investigation of nephrotoxicity on mice exposed to polystyrene nanoplastics and the potential amelioration effects of DHA-enriched phosphatidylserine. Sci. Total Environ. 2023, 892, 164808. [Google Scholar] [CrossRef]
- Ferreira, I.; Rauter, A.P.; Bandarra, N.M. Marine Sources of DHA-Rich Phospholipids with Anti-Alzheimer Effect. Mar. Drugs 2022, 20, 662. [Google Scholar] [CrossRef]
- Wang, Y.-M.; Zhang, Z.; Sheng, Y.; Chi, C.-F.; Wang, B. A systematic review on marine umami peptides: Biological sources, preparation methods, structure-umami relationship, mechanism of action and biological activities. Food Biosci. 2024, 57, 103637. [Google Scholar] [CrossRef]
- Siahaan, E.A.; Agusman; Pangestuti, R.; Shin, K.H.; Kim, S.K. Potential Cosmetic Active Ingredients Derived from Marine By-Products. Mar. Drugs 2022, 20, 734. [Google Scholar] [CrossRef]
- Molversmyr, E. Identification and Quantitation of Lipids in Atlantic Mackerel (Scomber scombrus), Wild and Farmed Atlantic Salmon (Salmo salar), and Salmon Feed by GC-MS. Master’s Thesis, Norwegian University of Life Sciences, As, Norway, 2020. [Google Scholar]
- Petricorena, Z.C. Chemical Composition of Fish and Fishery Products. In Handbook of Food Chemistry; Springer: Berlin/Heidelberg, Germany, 2014; pp. 1–28. [Google Scholar]
- Standal, I.B.; Mozuraityte, R.; Rustad, T.; Alinasabhematabadi, L.; Carlsson, N.G.; Undeland, I. Quality of filleted atlantic mackerel (Scomber scombrus) during chilled and frozen storage: Changes in lipids, vitamin D, proteins, and small metabolites, including biogenic amines. J. Aquat. Food Prod. Technol. 2018, 27, 338–357. [Google Scholar] [CrossRef]
- Guizani, S.E.O.; Moujahed, N. Seasonal Variation of Chemical and Fatty Acids Composition in Atlantic Mackerel from the Tunisian Northern-East Coast. J. Food Process. Technol. 2015, 6, 1–4. [Google Scholar] [CrossRef]
- Romotowska, P.E.; Karlsdóttir, M.G.; Gudjónsdóttir, M.; Kristinsson, H.G.; Arason, S. Seasonal and geographical variation in chemical composition and lipid stability of Atlantic mackerel (Scomber scombrus) caught in Icelandic waters. J. Food Compos. Anal. 2016, 49, 9–18. [Google Scholar] [CrossRef]
- Cretton, M.; Malanga, G.; Mazzuca Sobczuk, T.; Mazzuca, M. Marine lipids as a source of high-quality fatty acids and antioxidants. Food Rev. Int. 2022, 39, 4941–4964. [Google Scholar] [CrossRef]
- Wallace, P.D. Seasonal Variation in Fat Content of Mackerel (Scomber scombrus L.) Caught in the Western English Channel; Directorate of Fisheries Research: Lowestoft, UK, 1991. [Google Scholar]
- Jansen, T.; Gislason, H. Population structure of Atlantic mackerel (Scomber scombrus). PLoS ONE 2013, 8, e64744. [Google Scholar] [CrossRef] [PubMed]
- ICES. ICES Working Group on Mackerel and Horse Mackerel Egg Surveys (WGMEGS: Outputs from 2020 Meeting); ICES: Hong Kong, China, 2021. [Google Scholar]
- Oudiani, S.; Chetoui, I.; Darej, C.; Moujahed, N. Sex and seasonal variation in proximate composition and fatty acid profile of Scomber scombrus (L. 1758) fillets from the Middle East Coast of Tunisia. Grasas Aceites 2019, 70, e285. [Google Scholar] [CrossRef]
- Rustad, T.; Storrø, I.; Slizyte, R. Possibilities for the utilisation of marine by-products. Int. J. Food Sci. Technol. 2011, 46, 2001–2014. [Google Scholar] [CrossRef]
- Setijawati, D.; Aziz Jaziri, A.; Yufidasari, H.S.; Pratomo, M.D.; Wardani, D.W.; Ersyah, D. Effect of incubation time and pH on the protein characterization of the aqueous soluble phase of acidified mackerel by-product. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2020; Volume 493. [Google Scholar] [CrossRef]
- Rustad, T. Physical and chemical properties of protein seafood by-products. In Maximising the Value of Marine By-Products; Woodhead Publishing: Sawston, UK, 2007; pp. 3–21. [Google Scholar]
- Falch, E.; Rustad, T.; Jonsdottir, R.; Shaw, N.B.; Dumay, J.; Berge, J.P.; Arason, S.; Kerry, J.P.; Sandbakk, M.; Aursand, M. Geographical and seasonal differences in lipid composition and relative weight of by-products from gadiform species. J. Food Compos. Anal. 2006, 19, 727–736. [Google Scholar] [CrossRef]
- Selmi, S.; Mbarki, R.; Sadok, S. Seasonal change of lipid and fatty acid composition of little tuna Euthynnus alletteratus-by-prooucts. Nutr. Health 2008, 19, 189–194. [Google Scholar] [CrossRef]
- Jacobsen, C.; Warncke, S.A.; Hansen, S.H.; Sorensen, A.M. Fish Liver Discards as a Source of Long-Chain Omega-3 Polyunsaturated Fatty Acids. Foods 2022, 11, 905. [Google Scholar] [CrossRef]
- Bae, J.H.; Yoon, S.H.; Lim, S.Y. Heavy metal contents and chemical compositions of atlantic (Scomber scombrus), blue (Scomber australasicus), and chub (Scomber japonicus) mackerel muscles. Food Sci. Biotechnol. 2011, 20, 709–714. [Google Scholar] [CrossRef]
- Óskarsson, G.J.; Gudmundsdottir, A.; Sveinbjörnsson, S.; Sigurðsson, Þ. Feeding ecology of mackerel and dietary overlap with herring in Icelandic waters. Mar. Biol. Res. 2015, 12, 16–29. [Google Scholar] [CrossRef]
- Falk-Petersen, S.; Mayzaud, P.; Kattner, G.; Sargent, J.R. Lipids and life strategy of Arctic Calanus. Mar. Biol. Res. 2008, 5, 18–39. [Google Scholar] [CrossRef]
- Aidos, I.; Van der Padt, A.V.; Luten, J.B.; Boom, R.M. Seasonal changes in crude and lipid composition of herring fillets, byproducts, and respective produced oils. J. Agric. Food Chem. 2002, 50, 4589–4599. [Google Scholar] [CrossRef] [PubMed]
- Rincón-Cervera, M.Á.; Suárez-Medina, M.D.; Guil-Guerrero, J.L. Fatty acid composition of selected roes from some marine species. Eur. J. Lipid Sci. Technol. 2009, 111, 920–925. [Google Scholar] [CrossRef]
- Truzzi, C.; Annibaldi, A.; Illuminati, S.; Antonucci, M.; Api, M.; Scarponi, G.; Lombardo, F.; Pignalosa, P.; Carnevali, O. Characterization of the Fatty Acid Composition in Cultivated Atlantic Bluefin Tuna (Thunnus thynnus L.) Muscle by Gas Chromatography-Mass Spectrometry. Anal. Lett. 2018, 51, 2981–2993. [Google Scholar] [CrossRef]
- Laakmann, S.; Stumpp, M.; Auel, H. Vertical distribution and dietary preferences of deep-sea copepods (Euchaetidae and Aetideidae; Calanoida) in the vicinity of the Antarctic Polar Front. Polar Biol. 2009, 32, 679–689. [Google Scholar] [CrossRef]
- Coelho, T. Trophic Connectivity in Coastal Habitats Supporting Fishery Species; Universidade do Algarve: Faro, Portugal, 2017. [Google Scholar]
- Dalsgaard, J.; John, S.M.; Kattner, G.; Müller-Navarra, D.; Hagen, W. Fatty acid trophic markers in pelagic marine environment. Adv. Mar. Biol. 2003, 46, 225–340. [Google Scholar] [CrossRef]
- Hagen, W.; Kattner, G.; Friedrich, C. The lipid compositions of high-Antarctic notothenioid fish species with different life strategies. Polar Biol. 2000, 23, 785–791. [Google Scholar] [CrossRef]
- Saito, H.; Yamashiro, R.; Alasalvar, C.; Konno, T. Influence of diet on fatty acids of three subtropical fish, subfamily Caesioninae (Caesio diagramma and C. tile) and Family Siganidae (Siganus canaliculatus). Lipids 1999, 34, 1073–1082. [Google Scholar] [CrossRef]
- Colaco, A.; Prieto, C.; Martins, A.; Figueiredo, M.; Lafon, V.; Monteiro, M.; Bandarra, N.M. Seasonal variations in lipid composition of the hydrothermal vent mussel Bathymodiolus azoricus from the Menez Gwen vent field. Mar. Environ. Res. 2009, 67, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Farkas, T.; Kitajka, K.; Fodor, E.; Csengeri, I.; Lahdes, E.; Yeo, Y.K.; Krasznai, Z.; Halver, J.E. Docosahexaenoic acid-containing phospholipid molecular species in brains of vertebrates. Proc. Natl. Acad. Sci. USA 2000, 97, 6362–6366. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Li, K.W. Mass spectrometric identification of molecular species of phosphatidylcholine and lysophosphatidylcholine extracted from shark liver. J. Agric. Food Chem. 2007, 55, 9670–9677. [Google Scholar] [CrossRef] [PubMed]
- Linder, M.; Belhaj, N.; Sautot, P.; Arab Tehrany, E. From Krill to Whale: An overview of marine fatty acids and lipid compositions. Oléagineux Corps Gras Lipides 2010, 17, 194–204. [Google Scholar] [CrossRef]
- Sarney, D.B.; Fregapane, G.; Vulfson, E.N. Lipase-catalyzed synthesis of lysophospholipids in a continuous bioreactor. J. Am. Oil Chem. Soc. 1994, 71, 93–96. [Google Scholar] [CrossRef]
- Mnasri, T.; Ergan, F.; Herault, J.; Pencreac’h, G. Lipase-catalyzed Synthesis of Oleoyl-lysophosphatidylcholine by Direct Esterification in Solvent-free Medium without Water Removal. J. Oleo Sci. 2017, 66, 1009–1016. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, S.; Yamamoto, Y. Highly efficient preparation of 1-lysophosphatidylcholine via high proportion of Novozym(R) 435 (lipase B from Candida antarctica)-catalyzed ethanolysis. Biotechnol. Rep. 2020, 27, e00505. [Google Scholar] [CrossRef] [PubMed]
- Kotogan, A.; Furka, Z.T.; Kovacs, T.; Volford, B.; Papp, D.A.; Varga, M.; Huynh, T.; Szekeres, A.; Papp, T.; Vagvolgyi, C.; et al. Hydrolysis of Edible Oils by Fungal Lipases: An Effective Tool to Produce Bioactive Extracts with Antioxidant and Antimicrobial Potential. Foods 2022, 11, 1711. [Google Scholar] [CrossRef]
- Scheinman, S.B.; Sugasini, D.; Zayed, M.; Yalagala, P.C.R.; Marottoli, F.M.; Subbaiah, P.V.; Tai, L.M. LPC-DHA/EPA-Enriched Diets Increase Brain DHA and Modulate Behavior in Mice That Express Human APOE4. Front. Neurosci. 2021, 15, 690410. [Google Scholar] [CrossRef]
- Cravotto, C.; Fabiano-Tixier, A.S.; Claux, O.; Abert-Vian, M.; Tabasso, S.; Cravotto, G.; Chemat, F. Towards Substitution of Hexane as Extraction Solvent of Food Products and Ingredients with No Regrets. Foods 2022, 11, 3412. [Google Scholar] [CrossRef] [PubMed]
- Sicaire, A.-G.; Abert Vian, M.; Fine, F.; Carré, P.; Tostain, S.; Chemat, F. Experimental approachversusCOSMO-RS assisted solvent screening for predicting the solubility of rapeseed oil. OCL 2015, 22, D404. [Google Scholar] [CrossRef]
- Breil, C.; Meullemiestre, A.; Vian, M.; Chemat, F. Bio-Based Solvents for Green Extraction of Lipids from Oleaginous Yeast Biomass for Sustainable Aviation Biofuel. Molecules 2016, 21, 196. [Google Scholar] [CrossRef] [PubMed]
- Marino, D.J. Ethyl acetate. In Encyclopedia of Toxicology; Academic Press: Cambridge, MA, USA, 2005; pp. 277–282. [Google Scholar]
- Haraldsson, G.G.; Thorarensen, A. Preparation of phospholipids highly enriched with n-3 polyunsaturated fatty acids by lipase. J. Am. Oil Chem. Soc. 1999, 76, 1143–1149. [Google Scholar] [CrossRef]
- Haas, M.J.; Cichowicz, D.J.; Jun, W.; Scott, K. The enzymatic hydrolysis of triglyceride-phospholipid mixtures in an organic solvent. J. Am. Oil Chem. Soc. 1995, 72, 519–525. [Google Scholar] [CrossRef]
- Yang, G.; Yang, R.; Hu, J. Lysophosphatidylcholine synthesis by lipase-catalyzed ethanolysis. J. Oleo Sci. 2015, 64, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Ono, M.; Hosokawa, M.; Inoue, Y.; Takahashi, K. Concentration of Docosahexaenoic Phospholipid through Hydrolysis Lipozyme. J. Jpn. Oil Chem. Soc. 1997, 46, 867–872. [Google Scholar] [CrossRef]
- Rodrigues, R.C.; Fernandez-Lafuente, R. Lipase from Rhizomucor miehei as a biocatalyst in fats and oils modification. J. Mol. Catal. B Enzym. 2010, 66, 15–32. [Google Scholar] [CrossRef]
- Vaysse, L.; Ly, A.; Moulin, G.; Dubreucq, E. Chain-length selectivity of various lipases during hydrolysis, esterification and alcoholysis in biphasic aqueous medium. Enzym. Microb. Technol. 2002, 51, 648–655. [Google Scholar] [CrossRef]
- Rivero-Pino, F.; Padial-Dominguez, M.; Guadix, E.M.; Morales-Medina, R. Novozyme 435 and Lipozyme RM IM Preferably Esterify Polyunsaturated Fatty Acids at the sn-2 Position. Eur. J. Lipid Sci. Technol. 2020, 122, 2000115. [Google Scholar] [CrossRef]
- Moharana, T.R.; Byreddy, A.R.; Puri, M.; Barrow, C.; Rao, N.M. Selective Enrichment of Omega-3 Fatty Acids in Oils by Phospholipase A1. PLoS ONE 2016, 11, e0151370. [Google Scholar] [CrossRef]
- Bispo, P.; Batista, I.; Bernardino, R.J.; Bandarra, N.M. Preparation of triacylglycerols rich in omega-3 fatty acids from sardine oil using a Rhizomucor miehei lipase: Focus in the EPA/DHA ratio. Appl. Biochem. Biotechnol. 2014, 172, 1866–1881. [Google Scholar] [CrossRef]
- Mohammadi, M.; Habibi, Z.; Dezvarei, S.; Yousefi, M.; Ashjari, M. Selective enrichment of polyunsaturated fatty acids by hydrolysis of fish oil using immobilized and stabilized Rhizomucor miehei lipase preparations. Food Bioprod. Process. 2015, 94, 414–421. [Google Scholar] [CrossRef]
- Yousefi, M.; Marciello, M.; Guisan, J.M.; Fernandez-Lorente, G.; Mohammadi, M.; Filice, M. Fine Modulation of the Catalytic Properties of Rhizomucor miehei Lipase Driven by Different Immobilization Strategies for the Selective Hydrolysis of Fish Oil. Molecules 2020, 25, 545. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Handloser, D.; Widmer, V.; Reich, E. Separation of Phospholipids by HPTLC–An Investigation of Important Parameters. J. Liq. Chromatogr. Relat. Technol. 2008, 31, 1857–1870. [Google Scholar] [CrossRef]
- Weihrauch, J.L.; Posati, L.P.; Anderson, B.A.; Exler, J. Lipid conversion factors for calculating fatty acid contents of foods. J. Am. Oil Chem. Soc. 1977, 54, 36–40. [Google Scholar] [CrossRef]
- Ulbricht, T.L.V.; Southgate, D.A.T. Coronary heart disease: Seven dietary factors. Lancet 1991, 338, 982–992. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| March | April | May | ||||
|---|---|---|---|---|---|---|
| Muscle | By-products | Muscle | By-products | Muscle | By-products | |
| 14:0 | 4.6 ± 0.3 b,c | 5.3 ± 0.3c | 3.1 ± 0.6 a | 4.1 ± 0.1 b | 2.5 ± 0.2 a | 3.1 ± 0.1 a |
| 16:0 | 11.5 ± 0.4 a | 12.5 ± 0.1 a,b | 13.7 ± 1.1 b,c | 14.8 ± 0.3 c | 18.4 ± 0.3 d | 18.0 ± 0.3 d |
| 18:0 | 2.8 ± 0.1 b | 2.4 ± 0.1 a | 4.6 ± 0.2 d | 3.4 ± 0.1 c | 5.4 ± 0.1 e | 4.6 ± 0.1 d |
| SFA | 20.7 ± 0.8 a | 22.2 ± 0.4 a,b | 23.1 ± 1.7 b | 24.2 ± 0.2 b | 27.7 ± 0.5 c | 27.6 ± 0.3 c |
| 16:1 n-7 | 2.7 ± 0.1 b | 3.6 ± 0.1 c | 2.1 ± 0.3 a | 3.3 ± 0.1 c | 3.6 ± 0.2 c | 4.1 ± 0.1 d |
| 18:1 n-9 | 11.3 ± 0.2 a | 12.2 ± 0.2 a,b | 12.7 ± 0.7 b | 16.1 ± 0.2 c | 20.1 ± 0.2 d | 19.3 ± 0.1 d |
| 18:1 n-7 | 2.1 ± 0.1 a | 2.0 ± 0.1 a | 2.6 ± 0.1 b | 2.6 ± 0.1 b | 3.9 ± 0.4 c | 3.9 ± 0.1 c |
| 20:1 n-9 | 12.3 ± 0.2 f | 10.7 ± 0.2 e | 9.6 ± 0.7 d | 8.6 ± 0.2 c | 1.8 ± 0.1 a | 2.8 ± 0.1 b |
| 22:1 n-11 | 20.9 ± 0.6 c | 15.7 ± 0.4 b | 15.5 ± 3.3 b | 14.2 ± 0.3 b | 0.8 ± 0.1 a | 3.0 ± 0.4 a |
| 22:1 n-9 | 1.4 ± 0.0 c | 0.9 ± 0.1 a,b,c | 1.0 ± 0.6 a,b,c | 1.1 ± 0.0 b,c | 0.4 ± 0.1 a | 0.5 ± 0.1 a,b |
| 24:1 n-9 | 1.6 ± 0.1 b,c | 1.2 ± 0.1 a,b,c | 1.6 ± 0.6 b,c | 1.6 ± 0.1 c | 0.8 ± 0.0 a | 0.9 ± 0.1 a,b |
| MUFA | 53.9 ± 0.7 c | 48.0 ± 0.4 b | 46.7 ± 4.1 b | 49.9 ± 0.4 b,c | 31.8 ± 0.2 a | 35.8 ± 0.5 a |
| 18:2 n-6 | 1.7 ± 0.0 b | 1.6 ± 0.0 a,b | 2.1 ± 0.3 c | 1.9 ± 0.0 b,c | 1.3 ± 0.1 a | 1.3 ± 0.1 a |
| 18:3 n-3 | 0.9 ± 0.0 c | 1.2 ± 0.0 d | 0.7 ± 0.1 a | 0.8 ± 0.0 b,c | 0.8 ± 0.1 a,b | 0.8 ± 0.1 b,c |
| 18:4 n-3 | 2.2 ± 0.1 d | 3.1 ± 0.1 e | 1.1 ± 0.2 a | 1.7 ± 0.1 c | 1.4 ± 0.1 b | 1.4 ± 0.0 b |
| 20:4 n-6 | 0.7 ± 0.0 a | 0.7 ± 0.0 a | 1.0 ± 0.1 b | 0.9 ± 0.1 a,b | 1.2 ± 0.1 c | 1.2 ± 0.1 c |
| 20:5 n-3 | 4.3 ± 0.0 a | 5.6 ± 0.1 b | 4.9 ± 0.8 a,b | 4.8 ± 0.2 a,b | 9.9 ± 0.6 c | 9.6 ± 0.2 c |
| 22:5 n-3 | 1.3 ± 0.0 a | 1.3 ± 0.0 a | 1.4 ± 0.1 a | 1.3 ± 0.1 a | 1.9 ± 0.1 b | 1.8 ± 0.1 b |
| 22:6 n-3 | 8.8 ± 0.1 a | 9.7 ± 0.1 a | 14.2 ± 2.0 b | 8.8 ± 0.3 a | 20.2 ± 1.2 c | 15.8 ± 0.2 b |
| PUFA | 22.6 ± 0.2 a | 26.1 ± 0.3 a,b | 28.1 ± 3.5 b | 22.9 ± 0.5 a | 39.4 ± 1.5 d | 34.9 ± 0.4 c |
| n-3 PUFA | 18.9 ± 0.2 a | 22.6 ± 0.3 a,b | 23.4 ± 3.1 b | 18.8 ± 0.5 a | 35.5 ± 1.6 d | 30.9 ± 0.3 c |
| n-6 PUFA | 3.3 ± 0.0 a | 3.3 ± 0.0 a | 4.3 ± 0.4 c | 3.8 ± 0.0 b | 3.5 ± 0.1 a,b | 3.7 ± 0.1 a,b |
| n-3/n-6 | 5.8 ± 0.1 a | 6.9 ± 0.1 b | 5.4 ± 0.4 a | 4.9 ± 0.1 a | 10.1 ± 0.8 d | 8.4 ± 0.3 c |
| AI | 0.4 | 0.5 | 0.4 | 0.4 | 0.4 | 0.4 |
| TI | 0.2 | 0.2 | 0.2 | 0.3 | 0.2 | 0.2 |
| March | April | May | ||||
|---|---|---|---|---|---|---|
| Muscle | By-products | Muscle | By-products | Muscle | By-products | |
| 14:0 | 176.2 ± 12.3 c | 385.2 ± 16.0 e | 32.9 ± 6.4 a | 255.3 ± 7.0 d | 65.6 ± 4.6 b | 194.5 ± 2.7 c |
| 16:0 | 435.9 ± 14.3 b | 910.4 ± 10.0 d | 145.8 ± 12.2 a | 923.8 ± 14.6 d | 485.9 ± 9.0 c | 1112.3 ± 15.1 e |
| 18:0 | 103.9 ± 0.8 b | 179.0 ± 2.1 d | 48.9 ± 2.2 a | 214.7 ± 1.8 e | 142.6 ± 2.4 c | 282.5 ± 5.6 f |
| SFA | 782.4 ± 29.9 b | 1616.2 ± 23.7 d | 246.2 ± 17.7 a | 1505.6 ± 9.9 c | 729.5 ± 12.6 b | 1697.8 ± 17.3 e |
| 16:1 n-7 | 100.5 ± 3.2 b | 262.3 ± 4.3 e | 22.9 ± 3.2 a | 205.4 ± 5.3 c | 94.3 ± 3.4 b | 249.6 ± 6.1 d |
| 18:1 n-9 | 426.6 ± 5.7 b | 886.8 ± 7.9 d | 134.9 ± 7.7 a | 1004.8 ± 14.9 e | 530.8 ± 4.1 c | 1192.7 ± 3.9 f |
| 18:1 n-7 | 77.6 ± 0.7 b | 148.6 ± 1.5 d | 27.6 ± 1.4 a | 159.0 ± 0.2 d | 102.5 ± 9.4 c | 242.1 ± 2.8 e |
| 20:1 n-9 | 461.9 ± 5.5 d | 780.3 ± 9.9 f | 101.5 ± 7.6 b | 537.0 ± 10.2 e | 48.4 ± 0.4 a | 174.6 ± 9.0 c |
| 22:1 n-11 | 789.7 ± 23.3 c | 1143.0 ± 29.7 e | 164.8 ± 35.6 b | 883.4 ± 17.4 d | 20.1 ± 1.1 a | 186.0 ± 22.3 b |
| 24:1 n-9 | 58.4 ± 2.6 b | 91.4 ± 3.0 c | 16.7 ± 6.7 a | 102.6 ± 3.4 c | 21.1 ± 1.0 a | 53.6 ± 5.6 b |
| MUFA | 2033.8 ± 25.2 c | 3496.0 ± 32.3 f | 496.9 ± 43.9 a | 3107.0 ± 23.5 e | 839.0 ± 5.3 b | 2203.7 ± 32.8 d |
| 18:2 n-6 | 63.0 ± 0.8 c | 116.6 ± 1.7 e | 22.3 ± 2.5 a | 119.5 ± 2.0 e | 35.1 ± 1.1 b | 82.9 ± 2.7 d |
| 18:3 n-3 | 33.1 ± 0.7 c | 85.5 ± 1.3 e | 6.9 ± 0.7 a | 50.2 ± 0.9 d | 20.2 ± 0.7 b | 49.8 ± 3.1 d |
| 18:4 n-3 | 84.4 ± 1.5 c | 224.2 ± 4.2 e | 11.6 ± 1.5 a | 108.4 ± 3.6 d | 36.4 ± 2.1 b | 86.9 ± 1.7 c |
| 20:4 n-6 | 25.8 ± 1.4 b | 49.5 ± 0.8 d | 10.8 ± 1.1 a | 53.7 ± 1.2 d | 31.2 ± 1.7 c | 78.3 ± 3.3 e |
| 20:5 n-3 | 161.5 ± 0.5 b | 410.7 ± 6.8 e | 52.3 ± 8.1 a | 300.4 ± 8.0 d | 262.7 ± 15.6 c | 588.4 ± 12.2 f |
| 22:5 n-3 | 49.9 ± 0.8 b | 97.4 ± 0.6 d | 14.2 ± 1.3 a | 77.5 ± 1.1 c | 50.7 ± 2.4 b | 110.6 ± 4.4 e |
| 22:6 n-3 | 333.5 ± 4.5 b | 706.9 ± 8.6 d | 150.9 ± 21.0 a | 546.1 ± 14.9 c | 534.2 ± 31.1 c | 975.5 ± 11.5 e |
| PUFA | 851.7 ± 7.8 b | 1904.9 ± 26.5 e | 299.5 ± 37.1 a | 1409.5 ± 31.1 d | 1040.6 ± 40.1 c | 2134.2 ± 25.3 f |
| n-3 PUFA | 713.1 ± 7.2 b | 1646.1 ± 24.1 e | 248.3 ± 33.3 a | 1170.6 ± 28.7 d | 937.9 ± 42.9 c | 1903.2 ± 21.9 f |
| n-6 PUFA | 124.0 ± 1.2 c | 240.0 ± 2.2 e | 45.9 ± 4.0 a | 236.5 ± 2.0 e | 92.8 ± 3.7 b | 225.6 ± 6.8 d |
| March | April | May | ||||
|---|---|---|---|---|---|---|
| Muscle | By-products | Muscle | By-products | Muscle | By-products | |
| PL | 22.46 ± 1.07 b,c | 19.07 ± 3.93 a,b | 32.20 ± 2.43 e | 17.29 ± 0.61 a | 26.95 ± 4.13 d | 23.29 ± 0.63 c |
| NL | 77.54 ± 1.07 c,d | 80.93 ± 3.93 d,e | 67.88 ± 2.43 a | 82.71 ± 0.61 e | 73.05 ± 4.13 b | 76.71 ± 0.63 c |
| Muscle | By-products | |||
|---|---|---|---|---|
| PL | TAG | PL | TAG | |
| 14:0 | 1.0 ± 0.1 a | 6.1 ± 0.2 b | 1.1 ± 0.2 a | 5.9 ± 0.1 b |
| 16:0 | 16.6 ± 0.5 b | 12.3 ± 0.3 a | 17.8 ± 1.0 b | 13.1 ± 0.0 a |
| 18:0 | 8.1 ± 0.5 b | 2.3 ± 0.0 a | 7.8 ± 0.2 b | 2.4 ± 0.0 a |
| SFA | 27.1 ± 1.1 b | 23.0 ± 0.5 a | 28.4 ± 1.3 b | 23.2 ± 0.1 a |
| 16:1 n-7 | 0.8 ± 0 a | 3.3 ± 0.1 c | 1.4 ± 0.1 b | 3.9 ± 0.0 d |
| 18:1 n-9 | 5.0 ± 0.1 a | 13.2 ± 0.2 d | 9.2 ± 0.2 b | 12.6 ± 0.2 c |
| 18:1 n-7 | 2.6 ± 0.0 b | 2.1 ± 0.1 a | 2.2 ± 0.1 a | 2.2 ± 0.1 a |
| 20:1 n-9 | 3.3 ± 0.1 b | 13.3 ± 0.2 d | 2.6 ± 0.2 a | 11.7 ± 0.2 c |
| 22:1 n-11 | 1.2 ± 0.1 a | 21.1 ± 1 c | 1.4 ± 0.3 a | 18.4 ± 0.6 b |
| 24:1 n-9 | 1.3 ± 0.0 a | 1.3 ± 0.1 a | 2.0 ± 0.3 b | 1.3 ± 0.1 a |
| MUFA | 15.2 ± 0.3 a | 57.4 ± 1 d | 20.1 ± 0.7 b | 52.7 ± 0.7 c |
| 18:2 n-6 | 2.3 ± 0.0 c | 1.7 ± 0.1 b | 1.4 ± 0.1 a | 1.6 ± 0.0 b |
| 18:3 n-3 | 0.8 ± 0.0 b | 0.9 ± 0.1 b | 0.5 ± 0.0 a | 1.1 ± 0.0 c |
| 18:4 n-3 | 0.6 ± 0.0 a | 2.6 ± 0.2 b | 0.6 ± 0.0 a | 3.0 ± 0.1 c |
| 20:4 n-6 | 1.6 ± 0.0 b | 0.5 ± 0.0 a | 1.7 ± 0.1 c | 0.5 ± 0.0 a |
| 20:5 n-3 | 10.1 ± 0.3 d | 3.3 ± 0.3 a | 9.3 ± 0.3 c | 4.6 ± 0.2 b |
| 22:5 n-3 | 1.9 ± 0.0 b | 1.1 ± 0.1 a | 1.9 ± 0.2 b | 1.1 ± 0.0 a |
| 22:6 n-3 | 32.9 ± 1.0 c | 4.0 ± 0.2 a | 27.6 ± 2.4 b | 6.3 ± 0.2 a |
| PUFA | 53.9 ± 1.5 d | 16.8 ± 1.0 a | 47.0 ± 2.3 c | 21.4 ± 0.7 b |
| n-3 PUFA | 48.0 ± 1.4 d | 13.1 ± 0.9 a | 41.9 ± 2.9 c | 17.7 ± 0.6 b |
| n-6 PUFA | 5.7 ± 0.2 c | 3 ± 0.1 a | 4.7 ± 0.6 b | 2.9 ± 0.1 a |
| Before hydrolysis (PreH_extract) | After hydrolysis (PostH_extract) | |
|---|---|---|
| PL (%) | 50.79 ± 4.92 b | 21.16 ± 2.74 a |
| NL (%) | 49.21 ± 4.92 a | 78.84 ± 2.74 b |
| MAG | ND | ND |
| 1,2 DAG | ND | ND |
| 1,3 DAG + CH (%) | 6.96 ± 1.01 | 6.03 ± 0.79 |
| FFA (%) | 11.50 ± 0.29 a | 46.04 ± 2.39 b |
| TAG (%) | 30.76 ± 4.13 | 26.76 ± 1.23 |
| Before hydrolysis (PreH_extract) | After hydrolysis (PostH_extract) | |
|---|---|---|
| LPC (%) | ND | 25.19 ± 3.38 |
| LPE (%) | ND | 14.90 ± 3.70 |
| PC (%) | 57.76 ± 1.85 b | 21.69 ± 4.09 a |
| PE (%) | 42.24 ± 1.85 | 38.23 ± 4.52 |
| Before hydrolysis (PreH_extract) | After hydrolysis (PostH_extract) | |||||||
|---|---|---|---|---|---|---|---|---|
| PC | PE | LPC | LPE | PC | PE | LPC | LPE | |
| 14:0 | 1.4 ± 0.3 b | 0.3 ± 0.3 a | - | - | 0.2 ± 0.1 a | 0.1 ± 0.1 a | 0.2 ± 0.1 a | 0.4 ± 0.1 a |
| 16:0 | 19.5 ± 2.5 c | 9.4 ± 1.2 b | - | - | 1.3 ± 0.1 a | 1.2 ± 0.0 a | 0.6 ± 0.1 a | 1.0 ± 0.2 a |
| 18:0 | 5.8 ± 1 d | 7.9 ± 0.3 e | - | - | 1.1 ± 0.0 a,b | 1.4 ± 0.1 b,c | 0.5 ± 0.2 a,b | 2.5 ± 0.8 c |
| SFA | 28.3 ± 2.7 d | 19.7 ± 1.9 c | - | - | 8.5 ± 0.8 b | 9.1 ± 0.9 b | 1.6 ± 0.4 a | 6.1 ± 0.6 b |
| 16:1 n-7 | 0.9 ± 0.1 d | 0.6 ± 0.0 c | - | - | 0.2 ± 0.0 b | 0.3 ± 0.0 b | 0.1 ± 0.1 a | 0.2 ± 0.0 b |
| 18:1 n-9 | 6.1 ± 0.4 e | 4.0 ± 0.2 d | - | - | 3.2 ± 0.1 c,d | 3.1 ± 0.3 c | 0.7 ± 0.1 a,b | 1.1 ± 0.6 b |
| 18:1 n-7 | 1.6 ± 0.2 b | 3.5 ± 0.2 c | - | - | 1.0 ± 0.1 a | 1.0 ± 0.0 a | 1.2 ± 0.1 a,b | 0.6 ± 0.4 a |
| 20:1 n-9 | 2.4 ± 0.1 c | 3.9 ± 0.2 d | - | - | 0.6 ± 0.1 b | 0.2 ± 0.2 a,b | 0.6 ± 0.1 b,c | 0.4 ± 0.3 a,b |
| 22:1 n-11 | 0.2 ± 0.0 a,b | 0.1 ± 0.0 a,b | - | - | 0.1 ± 0.1 a | 0.2 ± 0.2 a,b | 0.1 ± 0.1 a,b | 0.4 ± 0.2 b |
| 22:1 n-9 | 0.0 ± 0.0 | ND | - | - | ND | ND | ND | 0.1 ± 0.1 |
| 24:1 n-9 | 0.0 ± 0.0 | ND | - | - | ND | ND | ND | 0.0 ± 0.0 |
| MUFA | 12.0 ± 0.6 c | 12.7 ± 0.3 c | - | - | 5.6 ± 0.0 b | 5.9 ± 0.2 b | 2.8 ± 0.6 a | 3.1 ± 1.1 a |
| 18:2 n-6 | 2.0 ± 0.2 c | 2.5 ± 0.1 d | - | - | 0.4 ± 0.0 b | 0.2 ± 0.2 a,b | 0.2 ± 0.2 a,b | 0.2 ± 0.2 a,b |
| 18:3 n-3 | 0.7 ± 0.0 c | 1.1 ± 0.0 d | - | - | 0.2 ± 0.0 b | 0.1 ± 0.1 a,b | 0.1 ± 0.1 a | 0.0 ± 0.1 a |
| 18:4 n-3 | 0.7 ± 0.0 a | 0.4 ± 0.0 a | - | - | 0.3 ± 0.0 a | 0.6 ± 0.4 a | 1.5 ± 0.1 b | 0.7 ± 0.6 a |
| 20:4 n-6 | 1.9 ± 0.2 d | 1.7 ± 0.1 c,d | - | - | 1.7 ± 0.1 c,d | 1.2 ± 0.1 a,b | 1.5 ± 0.1 b,c | 0.9 ± 0.2 a |
| 20:5 n-3 | 12.0 ± 0.7 d | 9.2 ± 0.4 c | - | - | 9.4 ± 0.3 c | 6.8 ± 0.6 b | 10.9 ± 0.2 c,d | 3.6 ± 1.6 a |
| 22:5 n-3 | 2.0 ± 0.2 c | 2.1 ± 0.0 c | - | - | 2.3 ± 0.0 c | 2.3 ± 0.2 c | 1.4 ± 0.2 b | 0.8 ± 0.4 a |
| 22:6 n-3 | 33.6 ± 2.6 a | 41.4 ± 1.5 b | - | - | 55.6 ± 1.4 c | 49.9 ± 0.8 c | 73.6 ± 1.8 d | 73.4 ± 4.5 d |
| PUFA | 56.3 ± 3.2 a | 63.8 ± 1.4 b | - | - | 78.3 ± 1.2 d | 73.1 ± 0.6 c | 94.1 ± 0.7 f | 86.6 ± 0.6 e |
| n-3 PUFA | 50.4 ± 3.2 a | 57.1 ± 1.3 b | - | - | 74.1 ± 1.0 c | 69.6 ± 1.1 c | 89.1 ± 1.1 e | 80.3 ± 3.2 d |
| n-6 PUFA | 5.4 ± 0.1 b | 6.2 ± 0.2 b | - | - | 3.5 ± 0.4 a,b | 2.7 ± 0.3 a,b | 4.9 ± 0.2 b | 6.2 ± 3.8 b |
| n-3/n-6 | 9.3 ± 0.6 a | 9.2 ± 0.1 a | - | - | 21.2 ± 1.8 b,c | 26.4 ± 3.6 c | 18.4 ± 1.1 b,c | 16.1 ± 7.9 a,b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, R.; Mendes, I.; Duarte, M.P.; Bandarra, N.M.; Gomes-Bispo, A. New Forms of Neuroactive Phospholipids for DHA Enrichment in Brain. Mar. Drugs 2024, 22, 116. https://doi.org/10.3390/md22030116
Gomes R, Mendes I, Duarte MP, Bandarra NM, Gomes-Bispo A. New Forms of Neuroactive Phospholipids for DHA Enrichment in Brain. Marine Drugs. 2024; 22(3):116. https://doi.org/10.3390/md22030116
Chicago/Turabian StyleGomes, Romina, Inês Mendes, Maria Paula Duarte, Narcisa M. Bandarra, and Ana Gomes-Bispo. 2024. "New Forms of Neuroactive Phospholipids for DHA Enrichment in Brain" Marine Drugs 22, no. 3: 116. https://doi.org/10.3390/md22030116