
Antioxidant, Collagenase Inhibitory, and Antibacterial Effects of Bioactive Peptides Derived from Enzymatic Hydrolysate of Ulva australis

Abstract
:
1. Introduction
2. Results
2.1. Properties of Hydrolysate from U. australis
2.2. Purification of Bioactive Peptides
2.3. Amino Acid Sequence of Bioactive Peptides from Hydrolysate
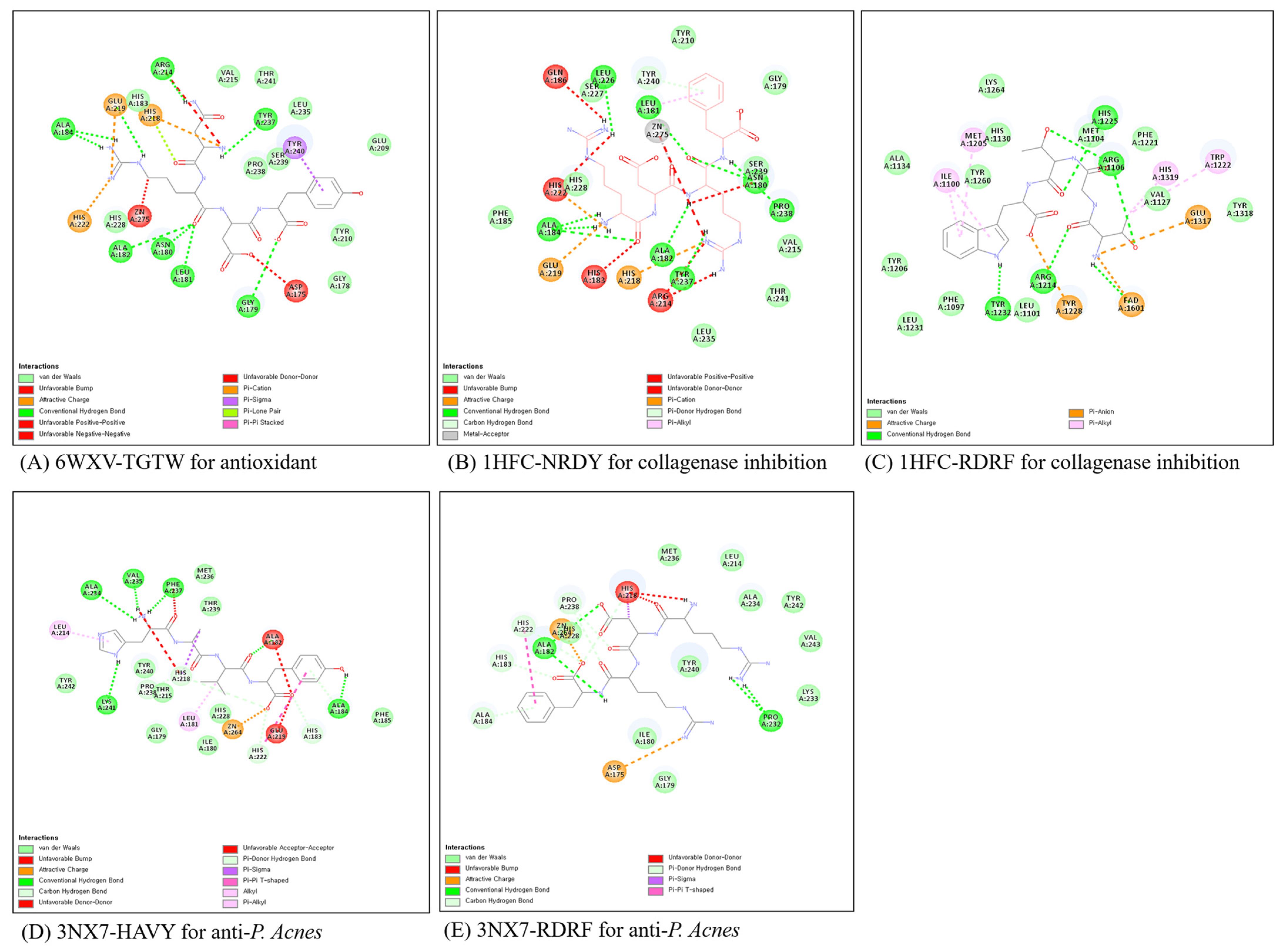
2.4. Molecular Docking for Screening Bioactive Peptides
2.5. In Vitro Validation of Bioactive Peptides
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of Hydrolysate from U. australis
4.3. Solubility of Hydrolysate
4.4. Analysis of Total and Free Amino Acid Composition
4.5. Purification of Bioactive Peptides from Hydrolysate
4.6. Cell Toxicity of the Hydrolysate
4.7. ABTS Radical Scavenging Activity and Total Antioxidant Capacity (TAC)
4.8. Inhibitory Activity of NO Production
4.9. COX-2 Inhibitory Activity
4.10. Collagenase Inhibitory Activity
4.11. Antibacterial Effect on P. Acnes
4.12. MALDI-TOF Mass and Q-TOF Mass/Mass Spectrometry
4.13. Molecular Docking
4.14. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, F.; Chen, N.; Wang, H.; Li, J.; Wang, J.; Qu, F. Novel insights into chloroplast genome evolution in the green macroalgal genus Ulva (Ulvophyceae, Chlorophyta). Front. Plant Sci. 2023, 14, 1126175. [Google Scholar] [CrossRef]
- Tabarsa, M.; Rezaei, M.; Ramezanpour, Z.; Waaland, J.R. Chemical compositions of the marine algae Gracilaria salicornia (Rhodophyta) and Ulva lactuca (Chlorophyta) as a potential food source. J. Sci. Food Agric. 2012, 92, 2500–2506. [Google Scholar] [CrossRef] [PubMed]
- Harnedy, P.A.; FitzGerald, R.J. Extraction and enrichment of protein from red and green macroalgae. In Natural Products from Marine Algae, 1st ed.; Stengel, D.B., Connan, S., Eds.; Humana Press: Totowa, NJ, USA; Springer Science: New York, NY, USA, 2015; pp. 103–108. [Google Scholar]
- Yalçın, S.; Karakaş, Ö.; Okudan, E.Ş.; Başkan, K.S.; Çekiç, S.D.; Apak, R. HPLC detection and antioxidant capacity determination of brown, red and green algal pigments in seaweed extracts. J. Chromatogr. Sci. 2021, 59, 325–337. [Google Scholar] [CrossRef] [PubMed]
- McDermid, K.L.; Stuercke, B. Nutritioanl composition of edible Hawaiian seaweeds. J. Appl. Phycol. 2003, 15, 513–524. [Google Scholar] [CrossRef]
- Rengasamy, K.R.R.; Mahomoodally, M.F.; Aumeeeruddy, M.Z.; Zengin, G.; Xiao, J.; Kim, D.H. Bioactive compounds in seweeds: An overview of their biological properties and safety. Food Chem. Toxicol. 2020, 135, 111013. [Google Scholar] [CrossRef]
- Echave, J.; Fraga-Cocrral, M.; Garcia-Perez, P.; Popović-Djordjević, J.; Avdović, E.H.; Radulović, M.; Xiao, J.; Prieto, M.P.; Simal-Gandara, J. Seaweed protein hydrolysates and bioactive peptides: Extraction, purification, and applications. Mar. Drugs 2021, 19, 500. [Google Scholar] [CrossRef] [PubMed]
- Aneiros, A.; Garateix, A. Bioactive peptides from marine sources: Pharmacological properties and isolation procedures. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2004, 803, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, C.; Gallagher, E.; Tasdemir, D.; Hayes, M. Heart health peptides from macroalgae and their potential use in functional foods. J. Agric. Food Chem. 2011, 59, 6829–6836. [Google Scholar] [CrossRef]
- Gupt, S.; Sabu-Ghannam, N. Bioactive potential and possible health effects of edible brown seaweeds. Trends Food Sci. Technol. 2011, 22, 315–326. [Google Scholar] [CrossRef]
- Fan, X.; Bai, L.; Zhu, L.; Yang, L.; Zhang, X. Marine algae-derived bioactive peptides for human nutrition and health. J. Agric. Food Chem. 2014, 62, 9211–9222. [Google Scholar] [CrossRef]
- Amin, M.A.; Chondra, U.; Mostafa, E.; Alam, M.M. Green seaweed Ulva lactuca, a potential source of bioactive peptides revealed by in silico analysis. Inform. Med. Unloacked 2022, 33, 101099. [Google Scholar] [CrossRef]
- Garcia-Vaquero, M.; Mora, L.; Hayes, M. In vitro and in silico approaches to generating and identifying angiotensin-converting enzyme I inhibitory peptides from green Macroalga Ulva lactuca. Mar. Drugs 2019, 17, 204. [Google Scholar] [CrossRef] [PubMed]
- Cian, R.E.; Hernadez-Chirlaque, C.; Gamez-Belmonte, R.; Drago, S.R.; de Medina, F.S.; Martínez-Augustin, O. Green alga Ulva spp. hydrolysates and their peptide fractions regulate cytokine production in splenic macrophages and lymphocytes involving the TLR-NFkB/MAPK oathways. Mar. Drugs 2018, 16, 235. [Google Scholar] [CrossRef] [PubMed]
- Masomian, M.; Lalani, S.; Poh, C.L. Molecular Docking of SP40 Peptide towards cellular receptors for enterovirus 71 (EV-A71). Molecules 2021, 26, 6576. [Google Scholar] [CrossRef] [PubMed]
- Petsalaki, E.; Russell, R.B. Peptide-mediated interactions in biological systems: New discoveries and applications. Curr. Opin. Biotechnol. 2008, 19, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Pyeun, J.H.; Jeon, J.G. Seafood Chemistry; Suhaksa: Seoul, Republic of Korea, 1994; pp. 283–355. [Google Scholar]
- Neduva, V.; Russell, R.B. Peptides mediating interaction networks: New leads at last. Curr. Opin. Biotechnol. 2006, 17, 465–471. [Google Scholar] [CrossRef]
- Saint-Maur, P.D. Is malignant mesenchymoma a mildly aggressive tumor? Ann. Pathol. 1992, 12, 146. [Google Scholar]
- Nevola, L.; Giralt, E. Modulating protein–protein interactions: The potential of peptides. Chem. Commun. 2015, 16, 3302–3315. [Google Scholar] [CrossRef]
- Kumar, V.; Agrawal, P.; Kumar, R.; Bhalla, S.; Usmani, S.S.; Varshney, G.C.; Raghava, G.P.S. Prediction of cell-penetrating potential of modified peptides containing natural and chemically modified residues. Front. Microbiol. 2018, 9, 725. [Google Scholar] [CrossRef]
- Usmani, S.S.; Bedi, G.; Samuel, J.S.; Singh, S.; Kalra, S.; Kumar, P.; Ahuja, A.A.; Sharma, M.; Gautam, A.; Raghava, G.P.S. THPdb: Database of FDA-approved peptide and protein therapeutics. PLoS ONE 2017, 12, e0181748. [Google Scholar] [CrossRef]
- Fosgerau, K.; Hoffmann, T. Peptide therapeutics: Current status and future directions. Drug Discov. Today 2015, 20, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Doucet, D.; Otter, D.E.; Gauthier, S.F.; Fodgeding, E.A. Enzyme-induced gelation of extensively hydrolyzed whey proteins by Alcalase: Peptide identification and determination of enzyme specificity. J. Agric. Food Chem. 2003, 51, 6300–6308. [Google Scholar] [CrossRef]
- Sun, J. Structure of mouse DUOX1-DOUXA1 provide mechanistic insights into enzyme activation and regulation. Nat. Struct. Mol. Biol. 2020, 27, 1086–1093. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial peptides: Classification, design, application and research progress in multiple fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar] [CrossRef]
- Le, C.F.; Fang, C.M.; Sekaran, S.D. Intracelluar targeting mechanisms by antimicrobial peptides. Antimicrob. Agents Chemother. 2017, 61, e02340-16. [Google Scholar] [CrossRef] [PubMed]
- Cerny, J.; Hobza, P. Non-covalent interactions in biomacromolecules. Phys. Chem. Chem. Phys. 2007, 9, 5291–5303. [Google Scholar] [CrossRef] [PubMed]
- Dawnay, A.B.; Hirst, A.D.; Perry, D.E.; Chambers, R.E. A critical assessment of current analytical methods for the routine assay of serum total protein and recommendations for their improvement. Ann. Clin. Biochem. 1991, 28, 556–567. [Google Scholar] [CrossRef]
- Jenssen, H.; Hamill, P.; Hancock, R.E.W. Peptide antimicrobial agents. Clin. Microbiol. Rev. 2006, 49, 491–511. [Google Scholar] [CrossRef]
- Cascales, J.J.L.; Zenak, S.; de la Torre, J.G.; Lezama, O.G.; Garro, A.; Daniel Enriz, R.D. Small cationic peptides: Influence of charge on their antimicrobial activity. ACS Omega 2018, 3, 5390–5398. [Google Scholar] [CrossRef]
- Bortolotii, A.; Troiano, C.; Bobone, S.; Konai, M.M.; Ghosh, C.; Bocchinfuso, G.; Acharya, Y.; Santucci, V.; Bonacorsi, S.; Di Stefano, C.; et al. Mechanism of lipid bilayer perturbation by bactericidal membrane active small molecules. Biochim. Biophys. Acta (BBA)-Biomembr. 2023, 1865, 184079. [Google Scholar] [CrossRef] [PubMed]
- Ennaas, N.; Hammami, R.; Beaulieu, L.; Fliss, I. Purification a characterization of four antibacterial peptides from protamex hydrolysate of Atlantic mackerel (Scomber scombrus) by-products. Biochem. Biophys. Res. Commun. 2015, 462, 195–200. [Google Scholar] [CrossRef]
- Choi, J.H.; Kim, K.T.; Kim, S.M. Biofunctional properties of enzymatic squid meat hydrolysate. Prev. Nutr. Food Sci. 2015, 20, 67–72. [Google Scholar]
- Jun, S.Y.; Park, P.J.; Jung, W.K.; Kim, S.K. Purification and characterization of an antioxidative peptide from enzymatic hydrolysate of yellowfin sole (Limanda aspera) frame protein. Eur. Food Res. Technol. 2004, 219, 20–26. [Google Scholar]
- Ames, B.N.; Shigenaga, M.K.; Hagen, T.M. Oxidants, antioxidants, and the degenerative diseases of aging. Proc. Natl. Acad. Sci. USA 1993, 90, 7915–7922. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Botta, J.R. Seafoods: Chemistry, Processing Technology and Quality; Springer: New York, NY, USA, 1994; pp. 331–356. [Google Scholar]
- Ganesan, K.; Kumar, K.S.; Rao, P.V.S.; Tsukui, Y.; Bhaskar, N.; Hosokawa, M.; Miyashita, K. Studies on chemical composition of three species of Enteromorpha. Biomed. Prev. Nutr. 2014, 4, 365–369. [Google Scholar] [CrossRef]
- Thiviya, P.; Gamage, A.; Gama-Arachchige, N.S.; Merah, O.; Madhujith, T. Seaweeds as a source of functional proteins. Phycology 2022, 2, 216–243. [Google Scholar] [CrossRef]
- Oliveira, A.P.; Pereira, D.M.; Andrade, P.B.; Valentão, P.; Sousa, C.; Pereira, J.A.; Bento, A.; Rodrigues, M.A.; Seabra, R.M.; Silva, B.M. Free amino acids of tronchuda cabbage (Brassica oleracea L. Var. costata DC): Influence of leaf position (internal or external) and collection time. J. Agric. Food Chem. 2008, 56, 5216–5221. [Google Scholar] [CrossRef] [PubMed]
- Lim, A.S.; Jeong, H.J.; Kim, S.J.; Ok, J.H. Amino acids profiles of six dinoflagellate species belonging to diverse families: Possible use as animal feeds in aquaculture. Algae 2018, 33, 279–290. [Google Scholar] [CrossRef]
- Wang, X.; Yu, H.; Xing, R.; Liu, S.; Chen, X.; Li, P. Optimization of oyster (Crassostrea talienwhanensis) protein hydrolysates using response surface methodology. Molecules 2020, 25, 2844. [Google Scholar] [CrossRef]
- Agrawal, P.; Singh, H.; Srivastava, H.K.; Singh, S.; Kishore, G.; Raghava, G.P.S. Benchmarking of different molecular docking methods for protein-peptide docking. BMC Bioinform. 2019, 19, 426. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Jin, B.; Li, H.; Huang, S.Y. HPEPDOCK: A web server for blind peptide-protein docking based on a hierarchical algorithm. Nucleic Acids Res. 2018, 46, W443–W450. [Google Scholar] [CrossRef] [PubMed]
- Vanvi, A.; Tsopmo, A. Pepsin digested oat bran proteins: Separation, antioxidant activity, and identification of new peptides. J. Chem. 2016, 2016, 8216378. [Google Scholar] [CrossRef]
- Lyskov, S.; Gray, J.J. The RosettaDock server for local protein-protein docking. Nucleic Acids Res. 2008, 36, W233–W238. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Total Amino Acid 1 (g/100 g) | Free Amino Acid 2 (mg/100 g-Hydrolysate) | |
|---|---|---|---|
| Powder of Dried U. australis | Hydrolysate | ||
| Asp | 1.70 ± 0.14 | 2.47 ± 0.03 | 63.6 ± 0.6 |
| Thr | 0.88 ± 0.08 | 1.03 ± 0.02 | 285.6 ± 3.8 |
| Ser | 1.08 ± 0.00 | 0.92 ± 0.03 | 45.7 ± 0.8 |
| Glu | 2.31 ± 0.09 | 3.03 ± 0.04 | 147.3 ± 4.6 |
| Pro | 0.57 ± 0.08 | 1.06 ± 0.01 | 35.9 ± 0.8 |
| Gly | 1.31 ± 0.09 | 1.14 ± 0.08 | 56.6 ± 0.4 |
| Ala | 1.86 ± 0.00 | 1.57 ± 0.07 | 63.6 ± 0.3 |
| Cys | 0.09 ± 0.01 | 0.14 ± 0.01 | 21.0 ± 1.1 |
| Val | 1.07 ± 0.09 | 1.31 ± 0.03 | 30.5 ± 0.3 |
| Met | 0.34 ± 0.02 | 0.53 ± 0.03 | 22.6 ± 6.6 |
| Ile | 0.74 ± 0.04 | 0.87 ± 0.03 | 9.0 ± 1.8 |
| Leu | 1.36 ± 0.06 | 1.56 ± 0.03 | 81.2 ± 4.2 |
| Tyr | 0.59 ± 0.05 | 1.20 ± 0.08 | 42.1 ± 9.2 |
| Phe | 0.95 ± 0.04 | 1.27 ± 0.08 | 33.3 ± 9.9 |
| His | 0.10 ± 0.00 | 0.14 ± 0.02 | 0 |
| Lys | 0.59 ± 0.01 | 0.62 ± 0.04 | 32.8 ± 0.2 |
| NH3 | 0.31 ± 0.02 | 0.31 ± 0.01 | 17.9 ± 0.1 |
| Arg | 1.30 ± 0.04 | 1.06 ± 0.05 | 113.3 ± 0.8 |
| Taurine | ND 3 | ND | 40.6 ± 0.5 |
| Sarcosine | ND | ND | 15.4 ± 1.6 |
| Citrulline | ND | ND | 15.4 ± 0.2 |
| α-amino butyric acid | ND | ND | 5.0 ± 0.1 |
| Cystathionine | ND | ND | 7.3 ± 2.7 |
| β-Alanine | ND | ND | 38.1 ± 14.7 |
| Homocysteine | ND | ND | 17.7 ± 3.4 |
| Hydroxylysine | ND | ND | 12.8 ± 0.4 |
| Ornithine | ND | ND | 32.8 ± 0.2 |
| Total | 17.16 ± 0.05 | 20.52 ± 1.01 | 1296.3 ± 60.2 |
| Fraction | Protein (mg/mL) | TAC (μM Trolox Equivalent/ mg-Protein) | NO Production (%) | COX-2 Inhibition (%) | Collagenase Inhibition (%) | Survival of P. Acnes (%) | |
|---|---|---|---|---|---|---|---|
| Hydrolysate | 14.85 | 382.0 ± 0.0 | 115.4 ± 3.5 | 66.0 ± 0.6 | 87.1 ± 7.4 | 112.0 ± 0.3 | |
| Cation-Chrom. | SP5 | 1.57 | 1177.5 ± 29.3 * | 127.4 ± 2.5 * | 0 | 27.4 ± 0.3 | 19.1 ± 1.6 * |
| SP7 | 2.81 | 674.1 ± 7.8 * | 130.8 ± 3.4 * | 0 | 4.2 ± 1.4 | 18.6 ± 1.1 * | |
| Anion-Chrom. | Q3 | 6.76 | 489.1 ± 10.8 * | 119.8 ± 4.5 | 11.2 ± 3.2 | 98.9 ± 0.1 * | 5.5 ± 0.3 * |
| Q4 | 6.74 | 24.7 ± 8.0 | 99.3 ± 5.1 * | 17.3 ± 0.9 | 0 | 123.7 ± 14.8 | |
| Q15 | 6.6 | 56.5 ± 1.2 | 124.2 ± 2.3 * | 0 | 0 | 98.2 ± 6.7 | |
| Q16 | 5.92 | 33.6 ± 0.3 | 118.9 ± 5.4 | 4.3 ± 3.2 | 0 | 97.8 ± 2.6 | |
| Q17 | 6.6 | 38.3 ± 0.6 | 125.2 ± 7.0 | 0 | 0 | 102.0 ± 4.8 | |
| Q23 | 7.02 | 13.4± 0.5 | 127.1 ± 8.0 | 18.1 ± 2.4 | 0 | 56.2 ± 4.3 * | |
| MS | Fraction (1) | RT (min) | z | m/z | Molecular Weight, Da | Sequence | Database |
|---|---|---|---|---|---|---|---|
| MALDI-TOF/MS | SP7/S9 | 439.818 | 438.52 | HVIA | lectin | ||
| 464.863 | 463.48 | TGTW | ribulose | ||||
| 488.901 | 488.53 | HAVY | ferritin | ||||
| 567.912 | 566.56 | NRDY | lectin | ||||
| 593.005 | 592.64 | RDRF | ribulose | ||||
| Micro Q-TOF III MS/MS | SP7/S9 | 11.64 | 1 | 231.18 | 230.17 | VL | ribulose |
| 14.89 | 1 | 245.18 | 244.18 | LL | ferritin ribulose apoprotein A1 | ||
| SP7/S19-20 | 14 | 1 | 445.23 | 444.23 | DPTL | ND | |
| 15.7 | 1 | 493.23 | 492.23 | PETF | ND | ||
| 17.1 | 1 | 546.29 | 545.29 | LPYPG | ND | ||
| 17.4 | 1 | 675.33 | 674.33 | LPYPGE | ND | ||
| 18.2 | 1 | 578.31 | 577.31 | FTPLT | ND | ||
| 18.5 | 1 | 489.27 | 488.27 | LPYP | ND | ||
| 20.3 | 1 | 344.24 | 343.24 | LVL | ND | ||
| 20.4 | 1 | 399.26 | 398.26 | PLGL | ND | ||
| 20.9 | 1 | 491.25 | 490.25 | FDPL | ND | ||
| 24.6 | 1 | 560.34 | 559.34 | PALLF | ND | ||
| SP7/S21 | 13.1 | 1 | 338.17 | 337.17 | VGY | ND | |
| 17.1 | 1 | 546.29 | 545.29 | LPYPG | ND | ||
| 17.3 | 1 | 463.25 | 462.25 | IAYP | ribulose | ||
| 17.4 | 1 | 352.19 | 351.19 | VSF | ND | ||
| 20.9 | 1 | 378.24 | 377.24 | MF | apoprotein A1 | ||
| Q3/S20 | 2.11 | 1 | 265.123 | 264.123 | VF | ferritin ribulose lectin plastocyanin apoprotein A1 | |
| 3.35 | 1 | 233.15 | 232.15 | LT | ribulose apoprotein A1 | ||
| 3.9 | 1 | 279.138 | 278.138 | LF | ribulose | ||
| 5.57 | 1 | 217.156 | 216.156 | VV | ribulose lectin plastocyanin apoprotein A1 | ||
| 6.92 | 1 | 228.193 | 227.193 | VGL | ferrtin | ||
| 7.6 | 1 | 274.177 | 273.177 | ALA | lectin apoprotein A1 | ||
| 8.02 | 1 | 286.177 | 287.177 | PLG | ribulose | ||
| 14.34 | 1 | 318.204 | 317.204 | SVL | ND | ||
| 14.39 | 2 | 265.19 | 528.38 | AVKVL | ND | ||
| 17.8 | 1 | 316.224 | 315.224 | ALL | ND | ||
| 19.77 | 2 | 286.664 | 571.328 | GVHLF | ND | ||
| 20.02 | 1 | 344.256 | 343.256 | VLL | ND | ||
| 20.96 | 1 | 502.326 | 501.326 | LSGLL | ND | ||
| 21.06 | 1 | 399.261 | 398.261 | PLGL | ND | ||
| 22.61 | 1 | 502.326 | 501.326 | LGVTL | ND | ||
| Q3/S21 | 7.05 | 1 | 203.14 | 202.14 | AL | lectin | |
| 9.26 | 1 | 231.171 | 230.171 | LV | apoprotein A1 | ||
| 9.59 | 1 | 229.156 | 228.156 | PL | ferritin |
| Peptide/Target Protein | Antioxidant Effect | Collagenase Inhibition | Antibacterial Effect | |||
|---|---|---|---|---|---|---|
| 6WXV | 1HFC | 3NX7 | ||||
| Cluster | Gold Score | Cluster | Gold Score | Cluster | Gold Score | |
| DPTL | 14 | 70.4 | 12 | 60.2 | 50 | 84 |
| FDPL | 17 | 85.4 | 8 | 57.3 | 39 | 86.9 |
| FTPLT | 14 | 82.1 | 16 | 60.8 | 14 | 80.5 |
| HAVY | 9 | 79.5 | 32 | 68.5 | 37 | 93.8 |
| HVIA | 27 | 76.6 | 29 | 62.4 | 46 | 85.3 |
| IAYP | 9 | 79.6 | 28 | 65.7 | 17 | 81 |
| LPYP | 7 | 74.3 | 8 | 55.4 | 14 | 59.3 |
| LPYPG | 4 | 71.2 | 21 | 58.4 | 6 | 61.3 |
| LVL | 31 | 68.6 | 16 | 60.5 | 18 | 76.2 |
| MF | 9 | 65.6 | 38 | 55 | 30 | 70.5 |
| NRDY | 8 | 100.5 | 29 | 85.9 | 14 | 97.1 |
| PALLF | 4 | 85.5 | 15 | 67.8 | 7 | 76.3 |
| PETE | 7 | 87.4 | 36 | 67.4 | 28 | 83 |
| PLGL | 8 | 68.9 | 44 | 67.8 | 48 | 76.8 |
| RDRF | 3 | 101.3 | 26 | 83.3 | 23 | 99.7 |
| TGTW | 13 | 89.7 | 27 | 67.4 | 16 | 85.3 |
| VGY | 11 | 54.1 | 42 | 64 | 50 | 77.8 |
| VSF | 29 | 70 | 23 | 63.1 | 39 | 80.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, Y.-A.; Kim, Y.-J.; Jin, S.-K.; Choi, H.-J. Antioxidant, Collagenase Inhibitory, and Antibacterial Effects of Bioactive Peptides Derived from Enzymatic Hydrolysate of Ulva australis. Mar. Drugs 2023, 21, 469. https://doi.org/10.3390/md21090469
Kang Y-A, Kim Y-J, Jin S-K, Choi H-J. Antioxidant, Collagenase Inhibitory, and Antibacterial Effects of Bioactive Peptides Derived from Enzymatic Hydrolysate of Ulva australis. Marine Drugs. 2023; 21(9):469. https://doi.org/10.3390/md21090469
Chicago/Turabian StyleKang, You-An, Ye-Jin Kim, Sang-Keun Jin, and Hwa-Jung Choi. 2023. "Antioxidant, Collagenase Inhibitory, and Antibacterial Effects of Bioactive Peptides Derived from Enzymatic Hydrolysate of Ulva australis" Marine Drugs 21, no. 9: 469. https://doi.org/10.3390/md21090469