
Anti-Protozoan Activities of Polar Fish-Derived Polyalanine Synthetic Peptides

 , , , , and
, , , , and
Abstract
:
1. Introduction
2. Results
2.1. Peptide Design Strategy
2.2. Anti-Protozoan Activities of Peptides
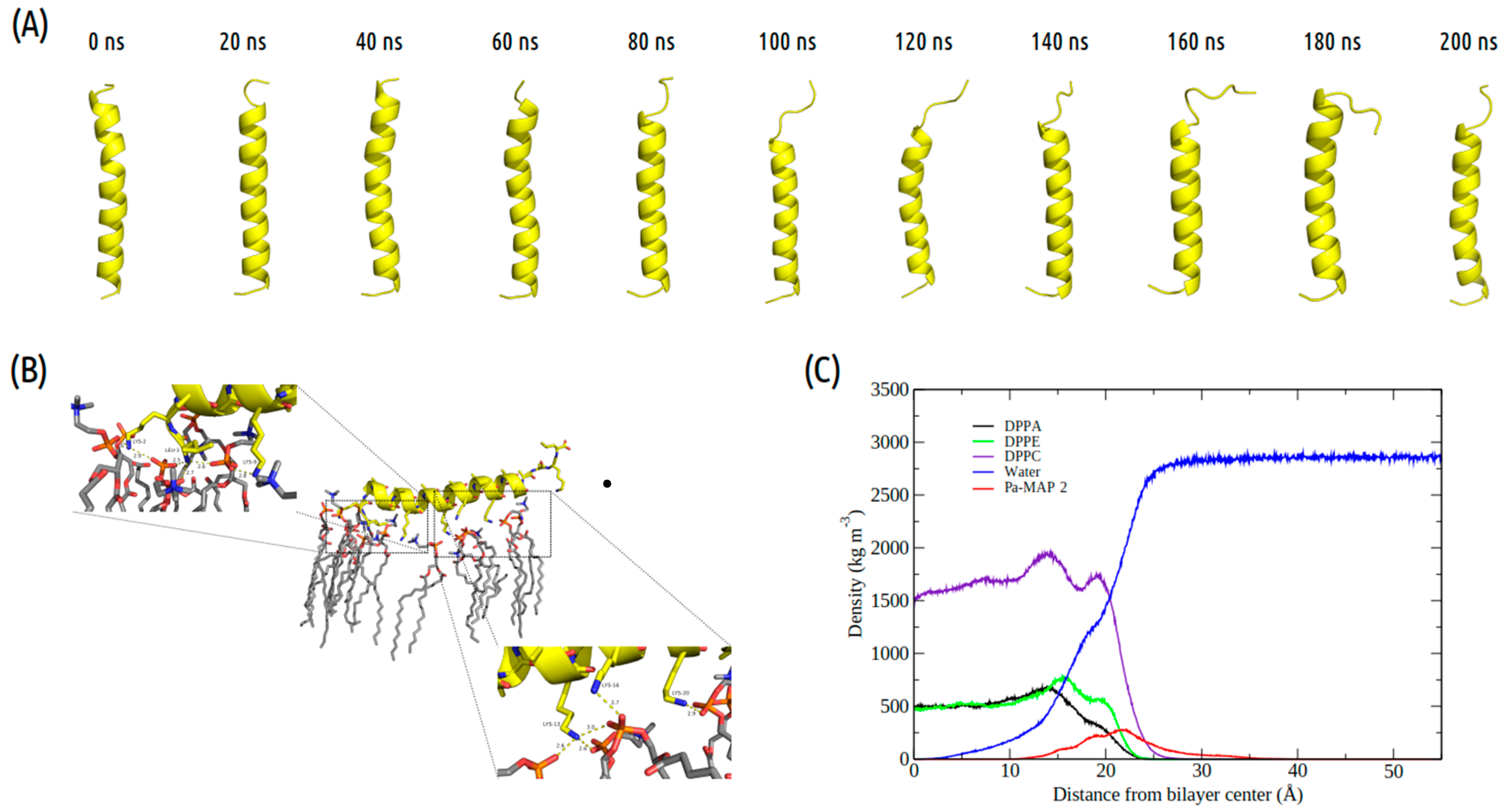
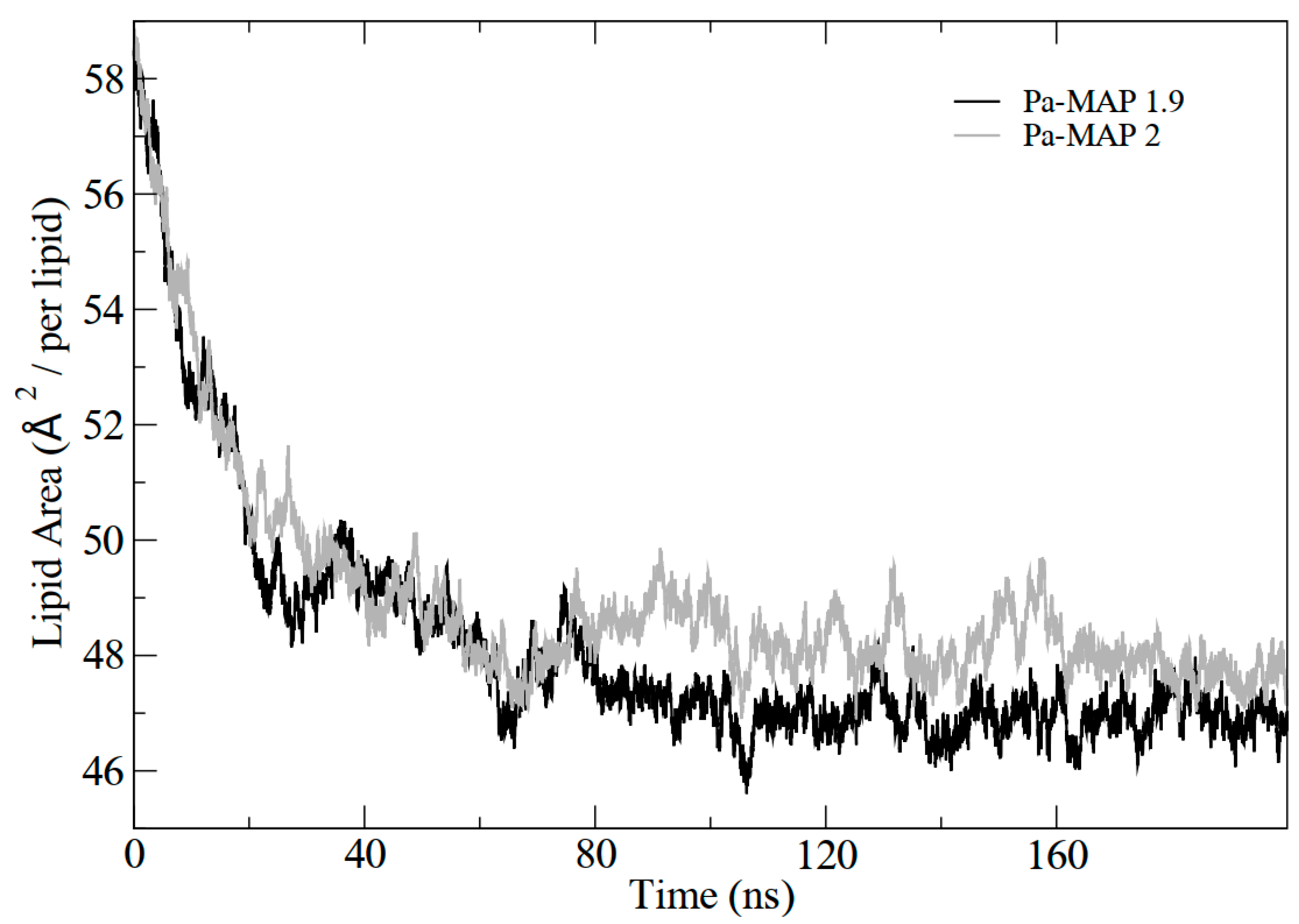
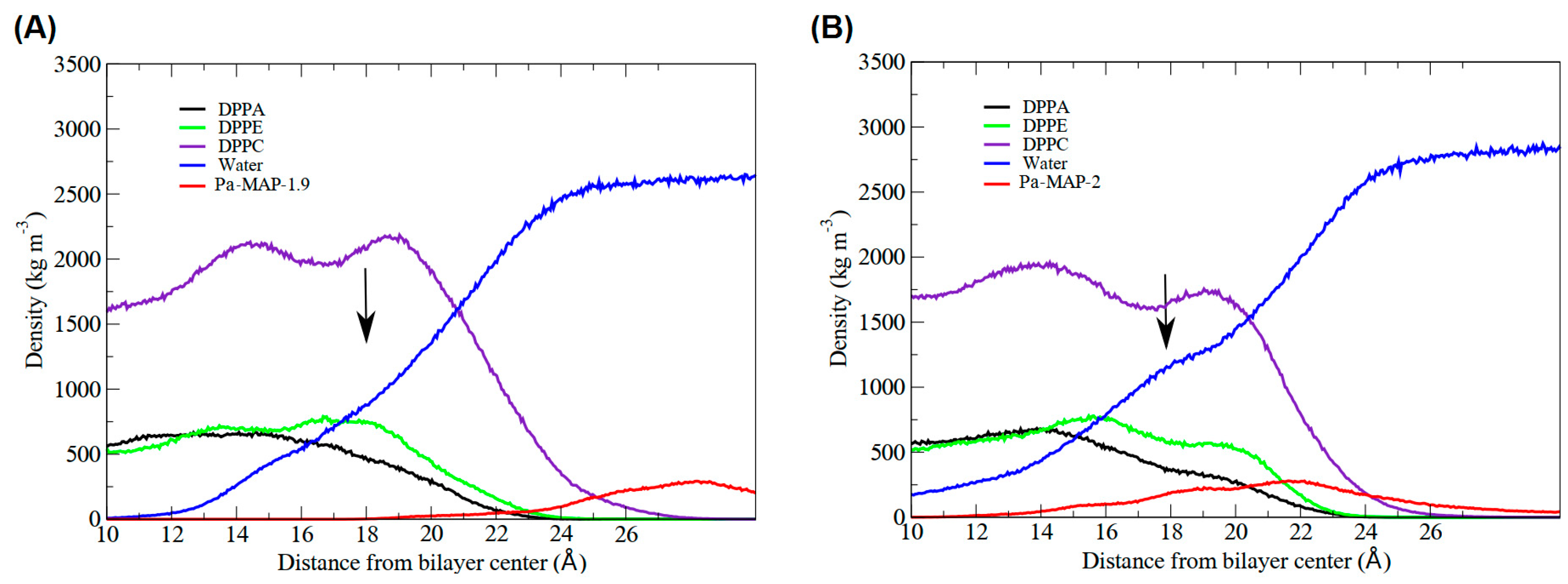
2.3. Molecular Dynamics Structural Analyses
3. Discussion
3.1. Peptides Design Strategy
3.2. Anti-Protozoan Activities of Peptides
3.3. Molecular Dynamics Structural Analyses
4. Materials and Methods
4.1. Peptide Synthesis and Preparation
4.2. In Vitro Bioassay of Antiplasmodial Activity
4.3. In Vitro Bioassay of Anti-Trypanosoma cruzi Activity
4.4. In Vitro Bioassay of Anti-Trypanosoma brucei gambiense Activity
4.5. Cytotoxic Activity on L6 Myoblasts
4.6. In Vitro Hemolysis Assay
4.7. In Silico Studies with a Trypomastigote Membrane
4.8. In Silico Structural Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hotez, P.J.; Fenwick, A.; Savioli, L.; Molyneux, D.H. Rescuing the bottom billion through control of neglected tropical diseases. Lancet 2009, 373, 1570–1575. [Google Scholar] [CrossRef] [PubMed]
- Rassi, A.; de Rezende, M. Leishmaniasis worldwide and global estimates of its incidence. Infect. Dis. Clin. N. Am. 2012, 26, 275–291. [Google Scholar] [CrossRef] [PubMed]
- Simarro, P.P.; Cecchi, G.; Paone, M.; Franco, J.R.; Diarra, A.; A Ruiz, J.; Fèvre, E.M.; Courtin, F.; Mattioli, R.C.; Jannin, J.G. The Atlas of human African trypanosomiasis: A contribution to global mapping of neglected tropical diseases. Int. J. Health Geogr. 2010, 9, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Echeverria, L.E.; Morillo, C.A. American Trypanosomiasis (Chagas Disease). Infect. Dis. Clin. 2019, 33, 119–134. [Google Scholar] [CrossRef]
- Maya, J.D.; Cassels, B.K.; Iturriaga-Vásquez, P.; Ferreira, J.; Faúndez, M.; Galanti, N.; Ferreira, A.; Morello, A. Mode of action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the mammalian host. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 146, 601–620. [Google Scholar] [CrossRef] [PubMed]
- Viotti, R.; Vigliano, C.; Lococo, B.; Alvarez, M.G.; Petti, M.; Bertocchi, G.; Armenti, A. Side effects of benznidazole as treatment in chronic Chagas disease: Fears and realities. Expert Rev. Anti-Infect. Ther. 2009, 7, 157–163. [Google Scholar] [CrossRef]
- Mejia, A.M.; Hall, B.S.; Taylor, M.C.; Gómez-Palacio, A.; Wilkinson, S.R.; Triana-Chávez, O.; Kelly, J.M. Benznidazole-Resistance in Trypanosoma cruzi Is a Readily Acquired Trait That Can Arise Independently in a Single Population. J. Infect. Dis. 2012, 206, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Büscher, P.; Cecchi, G.; Jamonneau, V.; Priotto, G. Human african trypanosomiasis. Lancet 2017, 390, 2397–2409. [Google Scholar] [CrossRef]
- Na-Bangchang, K.; Karbwang, J. Current status of malaria chemotherapy and the role of pharmacology in antimalarial drug research and development. Fundam. Clin. Pharmacol. 2009, 23, 387–409. [Google Scholar] [CrossRef]
- Nsanzabana, C.; Djalle, D.; Guérin, P.J.; Ménard, D.; González, I.J. Tools for surveillance of anti-malarial drug resistance: An assessment of the current landscape. Malar. J. 2018, 17, 1–16. [Google Scholar] [CrossRef]
- Nunes, E.; Frihling, B.; Barros, E.; De Oliveira, C.; Verbisck, N.; Flores, T.; Júnior, A.D.F.; Franco, O.; De Macedo, M.; Migliolo, L.; et al. Antibiofilm Activity of Acidic Phospholipase Isoform Isolated from Bothropserythromelas Snake Venom. Toxins 2020, 12, 606. [Google Scholar] [CrossRef] [PubMed]
- Frihling, B.E.F.; Boleti, A.P.d.A.; de Oliveira, C.F.R.; Sanches, S.C.; Cardoso, P.H.d.O.; Verbisck, N.; Macedo, M.L.R.; Rita, P.H.S.; Carvalho, C.M.E.; Migliolo, L. Purification, Characterization and Evaluation of the Antitumoral Activity of a Phospholipase A2 from the Snake Bothropsmoojeni. Pharmaceuticals 2022, 15, 724. [Google Scholar] [CrossRef]
- Migliolo, L.; Felício, M.R.; Cardoso, M.H.; Silva, O.N.; Xavier, M.-A.E.; Nolasco, D.O.; de Oliveira, A.S.; Roca-Subira, I.; Estape, J.V.; Teixeira, L.D.; et al. Structural and functional evaluation of the palindromic alanine-rich antimicrobial peptide Pa-MAP2. Biochim. Biophys. Acta-Biomembr. 2016, 1858, 1488–1498. [Google Scholar] [CrossRef]
- Cardoso, M.H.; Ribeiro, S.M.; Nolasco, D.O.; De La Fuente-Núñez, C.; Felício, M.R.; Gonçalves, S.; Matos, C.O.; Liao, L.M.; Santos, N.C.; Hancock, R.E.W.; et al. A polyalanine peptide derived from polar fish with anti-infectious activities. Sci. Rep. 2016, 6, 21385. [Google Scholar] [CrossRef] [Green Version]
- Migliolo, L.; Silva, O.N.; Silva, P.A.; Costa, M.P.; Costa, C.R.; Nolasco, D.O.; Barbosa, J.A.R.G.; Silva, M.R.R.; Bemquerer, M.P.; Lima, L.M.P.; et al. Structural and Functional Characterization of a Multifunctional Alanine-Rich Peptide Analogue from Pleuronectes americanus. PLoS ONE 2012, 7, e47047. Available online: https://guides.lib.monash.edu/ld.php?content_id=48260115 (accessed on 12 April 2023). [CrossRef] [PubMed]
- Nascimento, J.M.; Oliveira, M.D.; Franco, O.L.; Migliolo, L.; de Melo, C.P.; Andrade, C.A. Elucidation of mechanisms of interaction of a multifunctional peptide Pa-MAP with lipid membranes. Biochim. Biophys. Acta-Biomembr. 2014, 1838, 2899–2909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delarue-Cochin, S.; Grellier, P.; Maes, L.; Mouray, E.; Sergheraert, C.; Melnyk, P. Synthesis and antimalarial activity of carbamate and amide derivatives of 4-anilinoquinoline. Eur. J. Med. Chem. 2008, 43, 2045–2055. [Google Scholar] [CrossRef]
- Freire, K.A.; Torres MD, T.; Lima, D.B.; Monteiro, M.L.; Martins, A.M.C.; Oliveira, V.X., Jr. Wasp venom peptide as a new antichagasic agent. Toxicon 2020, 181, 71–78. [Google Scholar] [CrossRef]
- Takahashi, H.; Iwasaki, A.; Kurisawa, N.; Suzuki, R.; Jeelani, G.; Matsubara, T.; Sato, T.; Nozaki, T.; Suenaga, K. Motobamide, an antitrypanosomal cyclic peptide from a Leptolyngbya sp. marine cyanobacterium. J. Nat. Prod. 2021, 84, 1649–1655. [Google Scholar] [CrossRef]
- Sweeney-Jones, A.M.; Gagaring, K.; Antonova-Koch, J.; Zhou, H.; Mojib, N.; Soapi, K.; Skolnick, J.; McNamara, C.W.; Kubanek, J. Antimalarial Peptide and Polyketide Natural Products from the Fijian Marine Cyanobacterium Moorea producens. Mar. Drugs 2020, 18, 167. [Google Scholar] [CrossRef]
- Chatelain, E. Chagas Disease Drug Discovery: Toward a New Era. J. Biomol. Screen. 2015, 20, 22–35. [Google Scholar] [CrossRef] [Green Version]
- McGwire, B.S.; Olson, C.L.; Tack, B.F.; Engman, D.M. Killing of African Trypanosomes by Antimicrobial Peptides. J. Infect. Dis. 2003, 188, 146–152. [Google Scholar] [CrossRef] [Green Version]
- Arrighi, R.B.G.; Ebikeme, C.; Jiang, Y.; Ranford-Cartwright, L.; Barrett, M.P.; Langel, U.; Faye, I. Cell-Penetrating Peptide TP10 Shows Broad-Spectrum Activity against both Plasmodium falciparum and Trypanosoma brucei brucei. Antimicrob. Agents Chemother. 2008, 52, 3414–3417. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.J.N.; Pavanelli, W.R.; Gutierrez, F.R.S.; Lima, F.C.A.; da Silva, A.B.F.; Silva, J.S.; Franco, D.W. Complexation of the anti-Trypanosoma cruzi Drug Benznidazole Improves Solubility and Efficacy. J. Med. Chem. 2008, 51, 4104–4114. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Travkova, O.G.; Moehwald, H.; Brezesinski, G. The interaction of antimicrobial peptides with membranes. Adv. Colloid Interface Sci. 2017, 247, 521–532. [Google Scholar] [CrossRef]
- Gong, Z.; Ewart, K.V.; Hu, Z.; Fletcher, G.L.; Hew, C.L. Skin Antifreeze Protein Genes of the Winter Flounder, Pleuronectesamericanus, Encode Distinct and Active Polypeptides without the Secretory Signal and Prosequences (∗). J. Biol. Chem. 1996, 271, 4106–4112. [Google Scholar] [CrossRef] [Green Version]
- Desjardins, E.R.; Canfield, C.J.; Haynes, J.D.; Chulay, J.D. Quantitative assessment of antimalarial activity in vitro by a semiautomated microdilution technique. Antimicrob. Agents Chemother. 1979, 16, 710–718. [Google Scholar] [CrossRef] [Green Version]
- Guillon, J.; Grellier, P.; Labaied, M.; Sonnet, P.; Léger, J.-M.; Déprez-Poulain, R.; Forfar-Bares, I.; Dallemagne, P.; Lemaître, N.; Péhourcq, F.; et al. Synthesis, antimalarial activity, and molecular modeling of new pyrrolo[1,2-a]quinoxalines, bispyrrolo[1,2-a]quinoxalines, bispyrido[3,2-e]pyrrolo[1,2-a]pyrazines, and bispyrrolo[1,2-a]thieno[3,2-e]pyrazines. J. Med. Chem. 2004, 47, 1997–2009. [Google Scholar] [CrossRef] [PubMed]
- Charneau, S.; de Mesquita, M.L.; Bastos IM, D.; Santana, J.M.; de Paula, J.E.; Grellier, P.; Espindola, L.S. In vitro investigation of Brazilian Cerrado plant extract activity against Plasmodium falciparum, Trypanosoma cruzi and T. brucei gambiense. Nat. Prod. Res. 2016, 30, 1320–1326. [Google Scholar] [CrossRef] [PubMed]
- Triana-Vidal, L.E.; Castro, M.S.; Pires Júnior, O.R.; Álvares, A.C.M.; de Freitas, S.M.; Fontes, W.; Vargas, J.A.G.; Zúñiga-Baos, J.A.; de Fátima Correia Batista, I.; Grellier, P.; et al. Dendropsophin 1, a novel antimicrobial peptide from the skin secretion of the endemic Colombian frog Dendropsophuscolumbianus. Nat. Prod. Res. 2018, 32, 1383–1389. [Google Scholar] [CrossRef]
- Trager, W.; Jensen, J.B. Human malaria parasites in continuous culture. Science 1976, 193, 673–675. [Google Scholar] [CrossRef]
- Buckner, F.S.; Verlinde, C.L.; La Flamme, A.C.; Van Voorhis, W.C. Efficient technique for screening drugs for activity against Trypanosoma cruzi using parasites expressing beta-galactosidase. Antimicrob. Agents Chemother. 1996, 40, 2592–2597. [Google Scholar] [CrossRef]
- Bosc, D.; Mouray, E.; Cojean, S.; Franco, C.H.; Loiseau, P.M.; Freitas-Junior, L.H.; Moraes, C.B.; Grellier, P.; Dubois, J. Highly improved antiparasitic activity after introduction of an N-benzylimidazole moiety on protein farnesyltransferase inhibitors. Eur. J. Med. Chem. 2016, 109, 173–186. [Google Scholar] [CrossRef]
- Grellier, P.; Vendeville, S.; Joyeau, R.; Bastos, I.M.D.; Drobecq, H.; Frappier, F.; Teixeira, A.R.L.; Schrével, J.; Davioud-Charvet, E.; Sergheraert, C.; et al. Trypanosoma cruzi Prolyl Oligopeptidase Tc80 Is Involved in Nonphagocytic Mammalian Cell Invasion by Trypomastigotes. J. Biol. Chem. 2001, 276, 47078–47086. [Google Scholar] [CrossRef] [Green Version]
- Hirumi, H.; Hirumi, K. Axenic culture of African trypanosome bloodstream forms. Parasitol. Today 1994, 10, 80–84. [Google Scholar] [CrossRef]
- Loiseau, P.M.; Dreyfuss, G.; Daulouède, S.; Lachâtre, G.; Vincendeau, P.; Craciunescu, D.G. Trypanocidal effect of Ir-(COD)-pentamidine tetraphenylborate on Trypanosoma brucei and T. b. gambiense rodent models and serum kinetics in sheep. Trop. Med. Int. Health 1997, 2, 19–27. [Google Scholar] [CrossRef]
- Räz, B.; Iten, M.; Grether-Bühler, Y.; Kaminsky, R.; Brun, R. The Alamar Blue® assay to determine drug sensitivity of African trypanosomes (T. b. rhodesiense and T. b. gambiense) in vitro. Acta Trop. 1997, 68, 139–147. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Jo, S.; Lim, J.B.; Klauda, J.B.; Im, W. CHARMM-GUI Membrane Builder for Mixed Bilayers and Its Application to Yeast Membranes. Biophys. J. 2009, 97, 50–58. [Google Scholar] [CrossRef] [Green Version]
- Kaneda, Y.; Nagakura, K.; Goutsu, T. Lipid composition of three morphological stages of Trypanosoma cruzi. Comp. Biochem. Physiol. 1986, 83, 533–536. [Google Scholar] [CrossRef] [PubMed]
- Sicheri, F.; Yang, D.S.C. Ice-binding structure and mechanism of an antifreeze protein from winter flounder. Nature 1995, 375, 427–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šali, A.; Blundell, T.L. Comparative Protein Modelling by Satisfaction of Spatial Restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef]
- Wiederstein, M.; Sippl, M.J. ProSA-web: Interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Res. 2007, 35 (Suppl. S2), W407–W410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Leimkuhler, B.J.; Skeel, R.D. Symplectic Numerical Integrators in Constrained Hamiltonian Systems. J. Comput. Phys. 1994, 112, 117–125. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Anti-Plasmodial Activity | Anti-T. cruzi Activity | Anti-T. bruceigambiense Activity | Cytotoxicity against L6 Cell | |||
|---|---|---|---|---|---|---|---|
| (µM) | |||||||
| IC50 ± SD | SI | IC50 ± SD | SI | IC50 ± SD | SI | TC50 ± SD | |
| Pa-MAP | >45.2 | 1.0 | >45.2 | 1.0 | >45.2 | 1.0 | >45.2 |
| Pa-MAP1.9 | 35.0 ± 1.2 | 1.1 | >37.2 | 1.0 | 1.2 ± 0.1 | 31.0 | >37.2 |
| Pa-MAP2 | 47.0 ± 6.9 | >1 | >46.0 | 1.0 | 5.7 ± 1.2 | >8.1 | >46.0 |
| Chloroquine | 0.2 ± 0.1 | >1887 | >434.8 | ||||
| Nifurtimox | 0.6 ± 0.1 | 35.9 | 22.3 ± 8.0 | ||||
| Pentamidine | 0.004 ± 0.000 | 25,000 | >100 | ||||
| DMSO | <2%: w/o significant effect | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nunes, E.A.C.; da Silva, M.C.; Cardoso, M.H.; Preza, S.L.E.; de Oliveira, L.S.; Frihling, B.E.F.; Charneau, S.O.; Grellier, P.; Franco, O.L.; Migliolo, L. Anti-Protozoan Activities of Polar Fish-Derived Polyalanine Synthetic Peptides. Mar. Drugs 2023, 21, 434. https://doi.org/10.3390/md21080434
Nunes EAC, da Silva MC, Cardoso MH, Preza SLE, de Oliveira LS, Frihling BEF, Charneau SO, Grellier P, Franco OL, Migliolo L. Anti-Protozoan Activities of Polar Fish-Derived Polyalanine Synthetic Peptides. Marine Drugs. 2023; 21(8):434. https://doi.org/10.3390/md21080434
Chicago/Turabian StyleNunes, Ellynes Amancio Correia, Maria Cláudia da Silva, Marlon Henrique Cardoso, Sergio Leandro Espíndola Preza, Lucas Silva de Oliveira, Breno Emanuel Farias Frihling, Sébastien Olivier Charneau, Philippe Grellier, Octávio Luiz Franco, and Ludovico Migliolo. 2023. "Anti-Protozoan Activities of Polar Fish-Derived Polyalanine Synthetic Peptides" Marine Drugs 21, no. 8: 434. https://doi.org/10.3390/md21080434