

Harnessing the Value of Tripolium pannonicum and Crithmum maritimum Halophyte Biomass through Integrated Green Biorefinery


Abstract
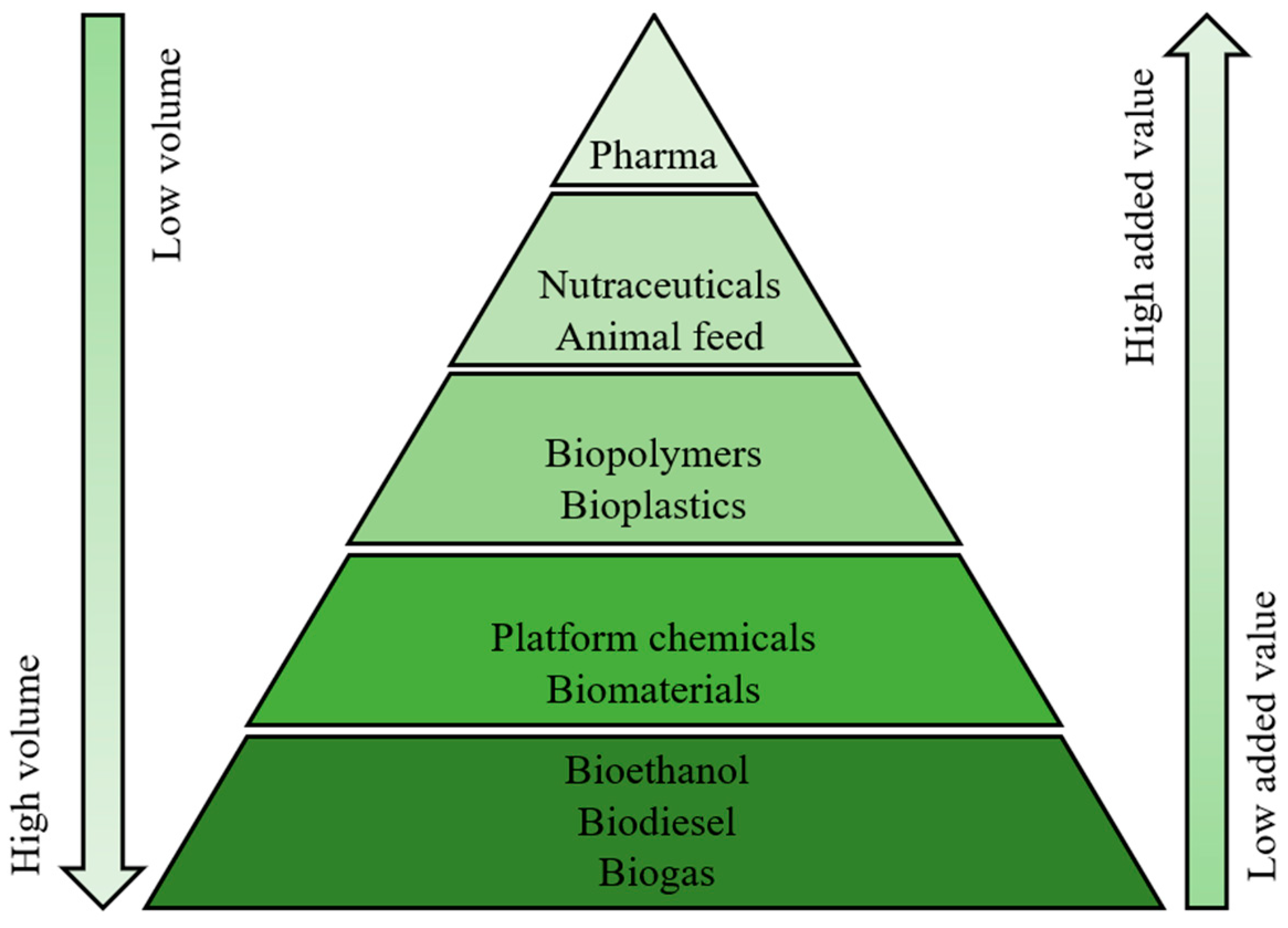
:1. Introduction
2. Results and Discussion
2.1. Fractionation and Extraction Yields
2.2. Fatty Acids in Non-Polar Extracts
2.3. Bioactive Compounds
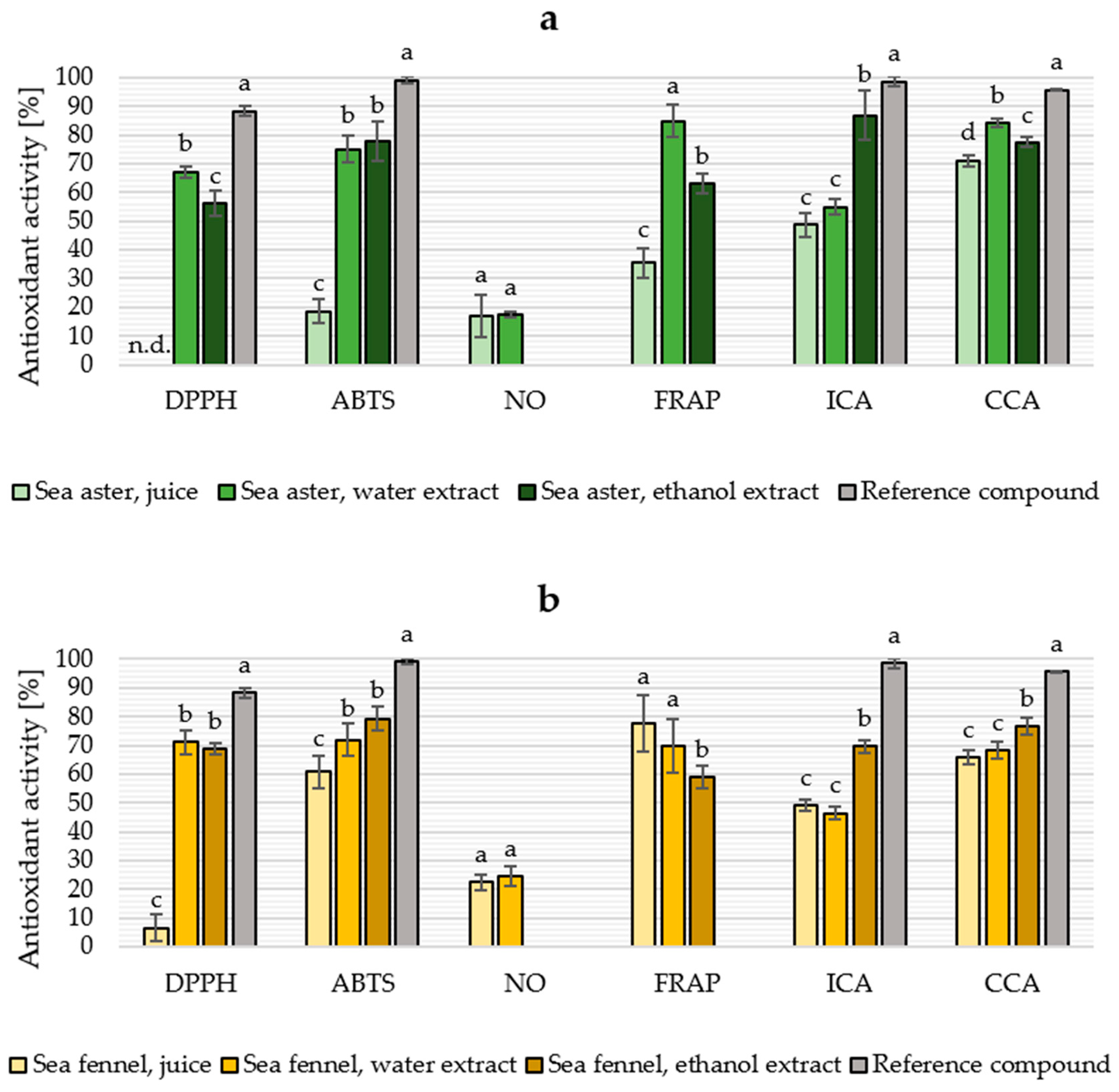
2.4. Antioxidant Properties
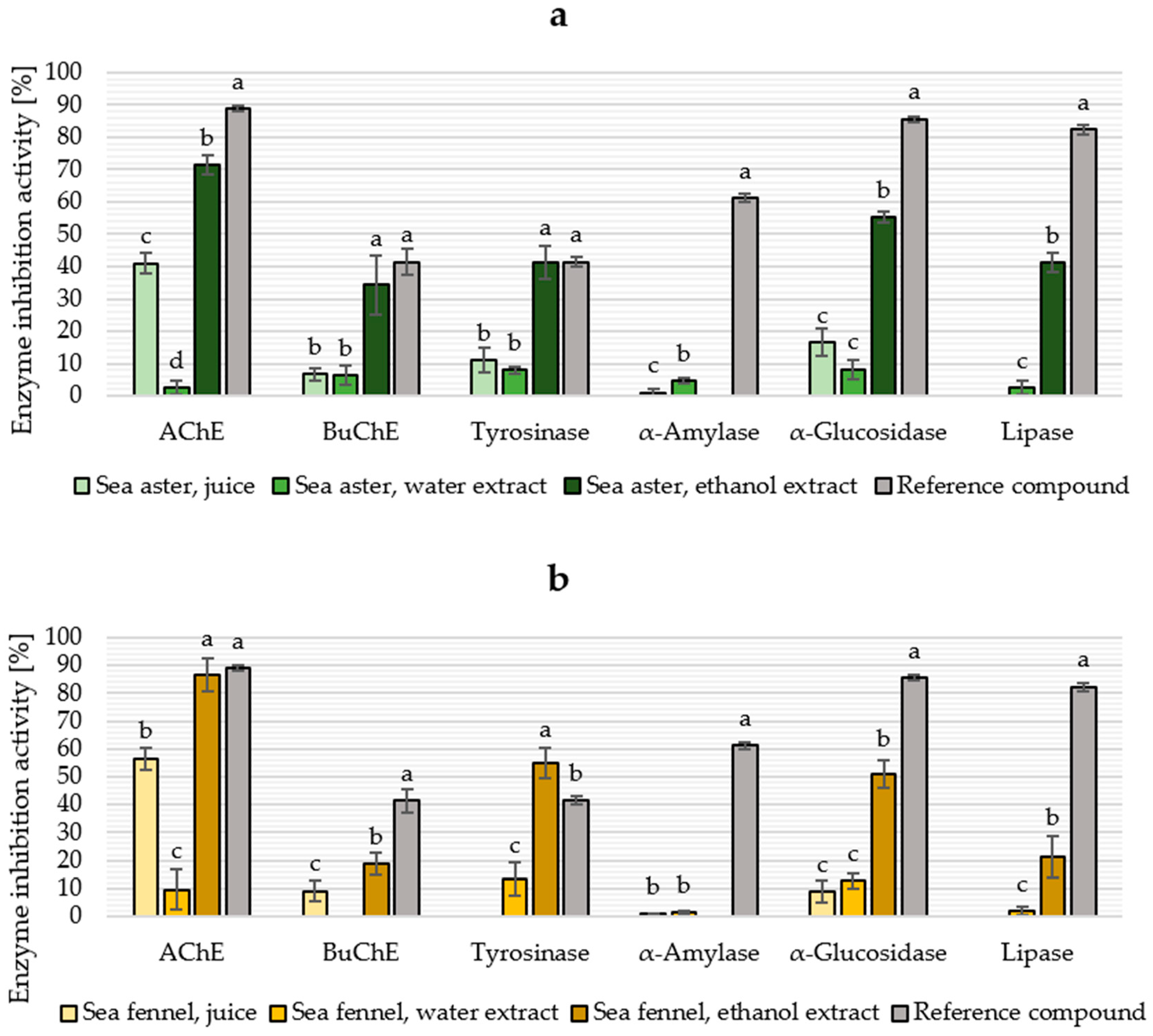
2.5. Enzyme Inhibition
3. Materials and Methods
3.1. Raw Material
3.2. Biomass Fractionation and Extraction
3.3. Transesterification and Determination of Fatty Acids
3.4. Total Photosynthetic Pigments
3.5. Phenolic Compounds in Plant Fractions
3.6. In Vitro Radical Scavenging Activity
3.7. In Vitro Antioxidant Activity by Metal-Based Assays
3.8. In Vitro Enzyme Inhibition Assays
3.9. Statistical Methods
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Intergovernmental Panel on Climate Change (IPCC). Synthesis Report of the IPCC Sixth Assessment Report—Summary for Policymakers; IPCC: Geneva, Switzerland, 2023. [Google Scholar]
- Zabaniotou, A. Redesigning a bioenergy sector in EU in the transition to circular waste-based Bioeconomy—A multidisciplinary review. J. Clean. Prod. 2018, 177, 197–206. [Google Scholar] [CrossRef]
- Mauser, W.; Klepper, G.; Zabel, F.; Delzeit, R.; Hank, T.; Putzenlechner, B.; Calzadilla, A. Global biomass production potentials exceed expected future demand without the need for cropland expansion. Nat. Commun. 2015, 6, 8946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muscat, A.; de Olde, E.M.; Ripoll-Bosch, R.; Van Zanten, H.H.E.; Metze, T.A.P.; Termeer, C.J.A.M.; van Ittersum, M.K.; de Boer, I.J.M. Principles, drivers and opportunities of a circular bioeconomy. Nat. Food 2021, 2, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Hulkko, L.S.S.; Chaturvedi, T.; Thomsen, M.H. Extraction and Quantification of Chlorophylls, Carotenoids, Phenolic Compounds, and Vitamins from Halophyte Biomasses. Appl. Sci. 2022, 12, 840. [Google Scholar] [CrossRef]
- Chaturvedi, T.; Torres, A.I.; Stephanopoulos, G.; Thomsen, M.H.; Schmidt, J.E. Developing process designs for biorefineries-definitions, categories, and unit operations. Energies 2020, 13, 1493. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Lopez, C.; Fraga-Corral, M.; Carpena, M.; García-Oliveira, P.; Echave, J.; Pereira, A.G.; Lourenço-Lopes, C.; Prieto, M.A.; Simal-Gandara, J. Agriculture waste valorisation as a source of antioxidant phenolic compounds within a circular and sustainable bioeconomy. Food Funct. 2020, 11, 4853–4877. [Google Scholar] [CrossRef]
- Stegmann, P.; Londo, M.; Junginger, M. The circular bioeconomy: Its elements and role in European bioeconomy clusters. Resour. Conserv. Recycl. X 2020, 6, 100029. [Google Scholar] [CrossRef]
- Yaashikaa, P.R.; Senthil Kumar, P.; Varjani, S. Valorization of agro-industrial wastes for biorefinery process and circular bioeconomy: A critical review. Bioresour. Technol. 2022, 343, 126126. [Google Scholar] [CrossRef]
- Fermoso, F.G.; Serrano, A.; Alonso-Fariñas, B.; Fernández-Bolaños, J.; Borja, R.; Rodríguez-Gutiérrez, G. Valuable Compound Extraction, Anaerobic Digestion, and Composting: A Leading Biorefinery Approach for Agricultural Wastes. J. Agric. Food Chem. 2018, 66, 8451–8468. [Google Scholar] [CrossRef]
- Manhongo, T.T.; Chimphango, A.F.A.; Thornley, P.; Röder, M. Current status and opportunities for fruit processing waste biorefineries. Renew. Sustain. Energy Rev. 2022, 155, 111823. [Google Scholar] [CrossRef]
- Caldeira, C.; Vlysidis, A.; Fiore, G.; De Laurentiis, V.; Vignali, G.; Sala, S. Sustainability of food waste biorefinery: A review on valorisation pathways, techno-economic constraints, and environmental assessment. Bioresour. Technol. 2020, 312, 123575. [Google Scholar] [CrossRef]
- Uzir, B.; Saha, A. Biorefinery Approach for Production of Biofuel from Algal Biomass. In Biorefinery Production Technologies for Chemicals and Energy; Wiley: Hoboken, NJ, USA, 2020; pp. 335–345. [Google Scholar]
- EIP-AGRI Focus Group Soil Salinisation: Final Report. Available online: https://ec.europa.eu/eip/agriculture/en/publications/eip-agri-focus-group-soil-salinisation-final (accessed on 20 January 2022).
- Nikalje, G.C.; Srivastava, A.K.; Pandey, G.K.; Suprasanna, P. Halophytes in biosaline agriculture: Mechanism, utilisation, and value addition. Land Degrad. Dev. 2018, 29, 1081–1095. [Google Scholar] [CrossRef]
- Duarte, B.; Caçador, I. Iberian Halophytes as Agroecological Solutions for Degraded Lands and Biosaline Agriculture. Sustainability 2021, 13, 1005. [Google Scholar] [CrossRef]
- Thomas, J.E.; Sinha, R.; Strand, Å.; Söderqvist, T.; Stadmark, J.; Franzén, F.; Ingmansson, I.; Gröndahl, F.; Hasselström, L. Marine biomass for a circular blue-green bioeconomy? A life cycle perspective on closing nitrogen and phosphorus land-marine loops. J. Ind. Ecol. 2022, 26, 2136–2153. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Ksouri, R.; Ksouri, W.M.; Jallali, I.; Debez, A.; Magné, C.; Hiroko, I.; Abdelly, C. Medicinal halophytes: Potent source of health promoting biomolecules with medical, nutraceutical and food applications. Crit. Rev. Biotechnol. 2012, 32, 289–326. [Google Scholar] [CrossRef]
- Faustino, M.V.; Faustino, M.A.F.; Pinto, D.C.G.A. Halophytic grasses, a new source of nutraceuticals? A review on their secondary metabolites and biological activities. Int. J. Mol. Sci. 2019, 20, 1067. [Google Scholar] [CrossRef] [Green Version]
- Giordano, R.; Saii, Z.; Fredsgaard, M.; Hulkko, L.S.S.; Poulsen, T.B.G.; Thomsen, M.E.; Henneberg, N.; Zucolotto, S.M.; Arendt-Nielsen, L.; Papenbrock, J.; et al. Pharmacological insights into halophyte bioactive extract action on anti-inflammatory, pain relief and antibiotics-type mechanisms. Molecules 2021, 26, 3140. [Google Scholar] [CrossRef]
- Giordano, R.; Aliotta, G.E.; Johannesen, A.S.; Voetmann-Jensen, D.; Laustsen, F.H.; Andersen, L.A.; Rezai, A.; Fredsgaard, M.; Lo Vecchio, S.; Arendt-Nielsen, L.; et al. Effects of Salicornia-Based Skin Cream Application on Healthy Humans’ Experimental Model of Pain and Itching. Pharmaceuticals 2022, 15, 150. [Google Scholar] [CrossRef]
- Jdey, A.; Falleh, H.; Ben Jannet, S.; Mkadmini Hammi, K.; Dauvergne, X.; Ksouri, R.; Magné, C. Phytochemical investigation and antioxidant, antibacterial and anti-tyrosinase performances of six medicinal halophytes. S. Afr. J. Bot. 2017, 112, 508–514. [Google Scholar] [CrossRef]
- Souid, A.; Della Croce, C.M.; Frassinetti, S.; Gabriele, M.; Pozzo, L.; Ciardi, M.; Abdelly, C.; Hamed, K.B.; Magné, C.; Longo, V. Nutraceutical Potential of Leaf Hydro-Ethanolic Extract of the Edible Halophyte Crithmum maritimum L. Molecules 2021, 26, 5380. [Google Scholar] [CrossRef] [PubMed]
- Ventura, Y.; Eshel, A.; Pasternak, D.; Sagi, M. The development of halophyte-based agriculture: Past and present. Ann. Bot. 2014, 115, 529–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Accogli, R.; Tomaselli, V.; Direnzo, P.; Perrino, E.V.; Albanese, G.; Urbano, M.; Laghetti, G. Edible Halophytes and Halo-Tolerant Species in Apulia Region (Southeastern Italy): Biogeography, Traditional Food Use and Potential Sustainable Crops. Plants 2023, 12, 549. [Google Scholar] [CrossRef] [PubMed]
- Biscotti, N.; Pieroni, A.; Luczaj, L. The hidden Mediterranean diet: Wild vegetables traditionally gathered and consumed in the Gargano area, Apulia, SE Italy. Acta Soc. Bot. Pol. 2015, 84, 327–338. [Google Scholar] [CrossRef]
- Vlahos, N.; Levizou, E.; Stathopoulou, P.; Berillis, P.; Antonopoulou, E.; Bekiari, V.; Krigas, N.; Kormas, K.; Mente, E. An Experimental Brackish Aquaponic System Using Juvenile Gilthead Sea Bream (Sparus aurata) and Rock Samphire (Crithmum maritimum). Sustainability 2019, 11, 4820. [Google Scholar] [CrossRef] [Green Version]
- Custódio, M.; Villasante, S.; Cremades, J.; Calado, R.; Lillebø, A. Unravelling the potential of halophytes for marine integrated multi-trophic aquaculture (IMTA)—A perspective on performance, opportunities and challenges. Aquac. Environ. Interact. 2017, 9, 445–460. [Google Scholar] [CrossRef] [Green Version]
- Zenobi, S.; Fiorentini, M.; Ledda, L.; Deligios, P.; Aquilanti, L.; Orsini, R. Crithmum maritimum L. Biomass Production in Mediterranean Environment. Agronomy 2022, 12, 926. [Google Scholar] [CrossRef]
- Hulkko, L.S.S.; Turcios, A.E.; Kohnen, S.; Chaturvedi, T.; Papenbrock, J.; Thomsen, M.H. Cultivation and characterisation of Salicornia europaea, Tripolium pannonicum and Crithmum maritimum biomass for green biorefinery applications. Sci. Rep. 2022, 12, 20507. [Google Scholar] [CrossRef]
- Jallali, I.; Zaouali, Y.; Missaoui, I.; Smeoui, A.; Abdelly, C.; Ksouri, R. Variability of antioxidant and antibacterial effects of essential oils and acetonic extracts of two edible halophytes: Crithmum maritimum L. and Inula crithmoïdes L. Food Chem. 2014, 145, 1031–1038. [Google Scholar] [CrossRef]
- Park, J.C.; Hur, J.M.; Park, J.G.; Hatano, T.; Yoshida, T.; Miyashiro, H.; Min, B.S.; Hattori, M. Inhibitory effects of Korean medicinal plants and camelliatannin H from Camellia japonica on human immunodeficiency virus type 1 protease. Phyther. Res. 2002, 16, 422–426. [Google Scholar] [CrossRef]
- Pereira, C.G.; Barreira, L.; da Rosa Neng, N.; Nogueira, J.M.F.; Marques, C.; Santos, T.F.; Varela, J.; Custódio, L. Searching for new sources of innovative products for the food industry within halophyte aromatic plants: In vitro antioxidant activity and phenolic and mineral contents of infusions and decoctions of Crithmum maritimum L. Food Chem. Toxicol. 2017, 107, 581–589. [Google Scholar] [CrossRef]
- Meot-Duros, L.; Magné, C. Antioxidant activity and phenol content of Crithmum maritimum L. leaves. Plant Physiol. Biochem. 2009, 47, 37–41. [Google Scholar] [CrossRef]
- Houta, O.; Akrout, A.; Amri, H. Phenolic amounts, antioxidant and antimicrobial potential of Crithmum maritimum cultivated in Tunisian arid zones. Planta Med. 2011, 77, 1416. [Google Scholar] [CrossRef]
- Generalić Mekinić, I.; Blažević, I.; Mudnić, I.; Burčul, F.; Grga, M.; Skroza, D.; Jerčić, I.; Ljubenkov, I.; Boban, M.; Miloš, M.; et al. Sea fennel (Crithmum maritimum L.): Phytochemical profile, antioxidative, cholinesterase inhibitory and vasodilatory activity. J. Food Sci. Technol. 2016, 53, 3104–3112. [Google Scholar] [CrossRef] [Green Version]
- Montero, M.P.; Sánchez-Faure, A.; Rico, D.; Martín-Diana, A.B.; de Palencia, P.F.; Sánchez-Guisado, M.J.; Gómez-Guillén, C.; López-Caballero, M.E.; Martínez-Alvarez, O. Sea Aster (Aster tripolium) as a Source of Bioactive Molecules. In Proceedings of the 49th WEFTA Conference, Tórshavn, Faroe Islands, 15–17 October 2019. [Google Scholar]
- Chai, W.; Chen, L.; Lian, X.Y.; Zhang, Z. Anti-glioma Efficacy and Mechanism of Action of Tripolinolate A from Tripolium pannonicum. Planta Med. 2018, 84, 786–794. [Google Scholar] [CrossRef] [PubMed]
- Wubshet, S.G.; Schmidt, J.S.; Wiese, S.; Staerk, D. High-resolution screening combined with HPLC-HRMS-SPE-NMR for identification of potential health-promoting constituents in sea aster and searocket—New nordic food ingredients. J. Agric. Food Chem. 2013, 61, 8616–8623. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.; Rodrigues, M.J.; Pereira, C.; Oliveira, M.; Barreira, L.; Varela, J.; Trampetti, F.; Custódio, L. Natural products from extreme marine environments: Searching for potential industrial uses within extremophile plants. Ind. Crops Prod. 2016, 94, 299–307. [Google Scholar] [CrossRef]
- Renna, M.; Gonnella, M.; Caretto, S.; Mita, G.; Serio, F. Sea fennel (Crithmum maritimum L.): From underutilised crop to new dried product for food use. Genet. Resour. Crop Evol. 2017, 64, 205–216. [Google Scholar] [CrossRef]
- Pavela, R.; Maggi, F.; Lupidi, G.; Cianfaglione, K.; Dauvergne, X.; Bruno, M.; Benelli, G. Efficacy of sea fennel (Crithmum maritimum L., Apiaceae) essential oils against Culex quinquefasciatus Say and Spodoptera littoralis (Boisd.). Ind. Crops Prod. 2017, 109, 603–610. [Google Scholar] [CrossRef]
- Tsoukatou, M.; Tsitsimpikou, C.; Vagias, C.; Roussis, V. Chemical intra-mediterranean variation and insecticidal activity of Crithmum maritimum. Z. Naturforsch.—Sect. C J. Biosci. 2001, 56, 211–215. [Google Scholar] [CrossRef]
- Polatoğlu, K.; Karakoç, Ö.C.; Yücel Yücel, Y.; Gücel, S.; Demirci, B.; Başer, K.H.C.; Demirci, F. Insecticidal activity of edible Crithmum maritimum L. essential oil against Coleopteran and Lepidopteran insects. Ind. Crops Prod. 2016, 89, 383–389. [Google Scholar] [CrossRef]
- Höltinger, S.; Schmidt, J.; Schönhart, M.; Schmid, E. A spatially explicit techno-economic assessment of green biorefinery concepts. Biofuels Bioprod. Biorefining 2014, 8, 325–341. [Google Scholar] [CrossRef]
- Mandl, M.G. Status of green biorefining in Europe. Biofuels Bioprod. Biorefining 2010, 4, 268–274. [Google Scholar] [CrossRef]
- Prieler, M.; Lindorfer, J.; Steinmüller, H. Life-cycle assessment of green biorefinery process options. Biofuels Bioprod. Biorefining 2019, 13, 1391–1401. [Google Scholar] [CrossRef]
- Novalin, S.; Zweckmair, T. Renewable resources—Green biorefinery: Separation of valuable substances from fluid-fractions by means of membrane technology. Biofuels Bioprod. Biorefining 2009, 3, 20–27. [Google Scholar] [CrossRef]
- Masters, D.G.; Benes, S.E.; Norman, H.C. Biosaline agriculture for forage and livestock production. Agric. Ecosyst. Environ. 2007, 119, 234–248. [Google Scholar] [CrossRef]
- Chaturvedi, T.; Hulkko, L.S.S.; Fredsgaard, M.; Thomsen, M.H. Extraction, Isolation, and Purification of Value-Added Chemicals from Lignocellulosic Biomass. Processes 2022, 10, 1752. [Google Scholar] [CrossRef]
- De Bhowmick, G.; Sarmah, A.K.; Sen, R. Lignocellulosic biorefinery as a model for sustainable development of biofuels and value added products. Bioresour. Technol. 2018, 247, 1144–1154. [Google Scholar] [CrossRef]
- Turcios, A.E.; Weichgrebe, D.; Papenbrock, J. Effect of salt and sodium concentration on the anaerobic methanisation of the halophyte Tripolium pannonicum. Biomass Bioenergy 2016, 87, 69–77. [Google Scholar] [CrossRef]
- Abideen, Z.; Ansari, R.; Khan, M.A. Halophytes: Potential source of ligno-cellulosic biomass for ethanol production. Biomass Bioenergy 2011, 35, 1818–1822. [Google Scholar] [CrossRef]
- Turcios, A.E.; Cayenne, A.; Uellendahl, H.; Papenbrock, J. Halophyte Plants and Their Residues as Feedstock for Biogas Production—Chances and Challenges. Appl. Sci. 2021, 11, 2746. [Google Scholar] [CrossRef]
- Turcios, A.E.; Weichgrebe, D.; Papenbrock, J. Uptake and biodegradation of the antimicrobial sulfadimidine by the species Tripolium pannonicum acting as biofilter and its further biodegradation by anaerobic digestion and concomitant biogas production. Bioresour. Technol. 2016, 219, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Cristina, C.; Lucia, P.; Sara, S.; Francesco, S.; Nobile, D.; Alessandro, M.; Conte, A.; Amalia, C. Study of the Efficacy of Two Extraction Techniques from Crithmum maritimum and Salicornia europaea. J. Food Nutr. Res. 2018, 6, 456–463. [Google Scholar] [CrossRef] [Green Version]
- Pedreiro, S.; Figueirinha, A.; Cavaleiro, C.; Cardoso, O.; Donato, M.M.; Salgueiro, L.; Ramos, F. Exploiting the Crithmum maritimum L. Aqueous Extracts and Essential Oil as Potential Preservatives in Food, Feed, Pharmaceutical and Cosmetic Industries. Antioxidants 2023, 12, 252. [Google Scholar] [CrossRef] [PubMed]
- Duarte, B.; Cabrita, M.T.; Gameiro, C.; Matos, A.R.; Godinho, R.; Marques, J.C.; Caçador, I. Disentangling the photochemical salinity tolerance in Aster tripolium L.: Connecting biophysical traits with changes in fatty acid composition. Plant Biol. 2017, 19, 239–248. [Google Scholar] [CrossRef]
- Ben Hamed, K.; Ben Youssef, N.; Ranieri, A.; Zarrouk, M.; Abdelly, C. Changes in content and fatty acid profiles of total lipids and sulfolipids in the halophyte Crithmum maritimum under salt stress. J. Plant Physiol. 2005, 162, 599–602. [Google Scholar] [CrossRef]
- Guil-Guerrero, J.L.; Rodríguez-García, I. Lipids classes, fatty acids and carotenes of the leaves of six edible wild plants. Eur. Food Res. Technol. 1999, 209, 313–316. [Google Scholar] [CrossRef]
- Atia, A.; Debez, A.; Barhoumi, Z.; Abdelly, C.; Smaoui, A. Localization and composition of seed oils of Crithmum maritimum L. (Apiaceae). Afr. J. Biotechnol. 2010, 9, 6482–6485. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Omega-6/Omega-3 Essential Fatty Acid Ratio and Chronic Diseases. Food Rev. Int. 2004, 20, 77–90. [Google Scholar] [CrossRef]
- Castañeda-Loaiza, V.; Oliveira, M.; Santos, T.; Schüler, L.; Lima, A.R.; Gama, F.; Salazar, M.; Neng, N.R.; Nogueira, J.M.F.; Varela, J.; et al. Wild vs. cultivated halophytes: Nutritional and functional differences. Food Chem. 2020, 333, 127536. [Google Scholar] [CrossRef]
- Hoste, H.; Jackson, F.; Athanasiadou, S.; Thamsborg, S.M.; Hoskin, S.O. The effects of tannin-rich plants on parasitic nematodes in ruminants. Trends Parasitol. 2006, 22, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Politeo, O.; Popović, M.; Veršić Bratinčević, M.; Kovačević, K.; Urlić, B.; Generalić Mekinić, I. Chemical Profiling of Sea Fennel (Crithmum maritimum L., Apiaceae) Essential Oils and Their Isolation Residual Waste-Waters. Plants 2023, 12, 214. [Google Scholar] [CrossRef] [PubMed]
- Stanković, M.S.; Petrović, M.; Godjevac, D.; Stevanović, Z.D. Screening inland halophytes from the central balkan for their antioxidant activity in relation to total phenolic compounds and flavonoids: Are there any prospective medicinal plants? J. Arid Environ. 2015, 120, 26–32. [Google Scholar] [CrossRef]
- Kapadia, P.; Newell, A.S.; Cunningham, J.; Roberts, M.R.; Hardy, J.G. Extraction of High-Value Chemicals from Plants for Technical and Medical Applications. Int. J. Mol. Sci. 2022, 23, 10334. [Google Scholar] [CrossRef] [PubMed]
- Boestfleisch, C.; Wagenseil, N.B.; Buhmann, A.K.; Seal, C.E.; Wade, E.M.; Muscolo, A.; Papenbrock, J. Manipulating the antioxidant capacity of halophytes to increase their cultural and economic value through saline cultivation. AoB Plants 2014, 6, plu046. [Google Scholar] [CrossRef] [PubMed]
- Duarte, B.; Goessling, J.W.; Marques, J.C.; Caçador, I. Ecophysiological constraints of Aster tripolium under extreme thermal events impacts: Merging biophysical, biochemical and genetic insights. Plant Physiol. Biochem. 2015, 97, 217–228. [Google Scholar] [CrossRef]
- Ventura, Y.; Myrzabayeva, M.; Alikulov, Z.; Cohen, S.; Shemer, Z.; Sagi, M. The importance of iron supply during repetitive harvesting of Aster tripolium. Funct. Plant Biol. 2013, 40, 968–976. [Google Scholar] [CrossRef]
- Wiszniewska, A.; Koźmińska, A.; Hanus-Fajerska, E.; Dziurka, M.; Dziurka, K. Insight into mechanisms of multiple stresses tolerance in a halophyte Aster tripolium subjected to salinity and heavy metal stress. Ecotoxicol. Environ. Saf. 2019, 180, 12–22. [Google Scholar] [CrossRef]
- Geissler, N.; Hussin, S.; Koyro, H.-W. Elevated atmospheric CO2 concentration ameliorates effects of NaCl salinity on photosynthesis and leaf structure of Aster tripolium L. J. Exp. Bot. 2008, 60, 137–151. [Google Scholar] [CrossRef] [Green Version]
- Sarrou, E.; Siomos, A.S.; Riccadona, S.; Aktsoglou, D.C.; Tsouvaltzis, P.; Angeli, A.; Franceschi, P.; Chatzopoulou, P.; Vrhovsek, U.; Martens, S. Improvement of sea fennel (Crithmum maritimum L.) nutritional value through iodine biofortification in a hydroponic floating system. Food Chem. 2019, 296, 150–159. [Google Scholar] [CrossRef]
- Siracusa, L.; Kulisic-Bilusic, T.; Politeo, O.; Krause, I.; Dejanovic, B.; Ruberto, G. Phenolic composition and antioxidant activity of aqueous infusions from Capparis spinosa L. and Crithmum maritimum L. before and after submission to a two-step in vitro digestion model. J. Agric. Food Chem. 2011, 59, 12453–12459. [Google Scholar] [CrossRef]
- Zafeiropoulou, V.; Tomou, E.-M.; Karioti, A.; Skaltsa, H.; Ioannidou, O. Sea fennel: Phytochemical analysis of Greek wild and cultivated Crithmum maritimum L. populations, based on HPLC-PDA-MS and NMR methods. J. Pharmacogn. Phytochem. 2020, 9, 998–1004. [Google Scholar]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- García-Sánchez, A.; Miranda-Díaz, A.G.; Cardona-Muñoz, E.G. The Role of Oxidative Stress in Physiopathology and Pharmacological Treatment with Pro- and Antioxidant Properties in Chronic Diseases. Oxid. Med. Cell. Longev. 2020, 2020, 2082145. [Google Scholar] [CrossRef]
- Kamm, B.; Schönicke, P.; Kamm, M. Biorefining of green biomass—Technical and energetic considerations. Clean—Soil Air Water 2009, 37, 27–30. [Google Scholar] [CrossRef]
- Thang, V.H.; Novalin, S. Green Biorefinery: Separation of lactic acid from grass silage juice by chromatography using neutral polymeric resin. Bioresour. Technol. 2008, 99, 4368–4379. [Google Scholar] [CrossRef] [PubMed]
- Mekinic, I.G.; Burčul, F.; Blaževic, I.; Skroza, D.; Kerum, D.; Katalinic, V. Antioxidative/acetylcholinesterase inhibitory activity of some Asteraceae plants. Nat. Prod. Commun. 2013, 8, 471–474. [Google Scholar] [CrossRef] [Green Version]
- Aghraz, A.; Gonçalves, S.; Rodríguez-Solana, R.; Dra, L.A.; Di Stefano, V.; Dugo, G.; Cicero, N.; Larhsini, M.; Markouk, M.; Romano, A. Antioxidant activity and enzymes inhibitory properties of several extracts from two Moroccan Asteraceae species. S. Afr. J. Bot. 2018, 118, 58–64. [Google Scholar] [CrossRef]
- Nguir, A.; Besbes, M.; Ben Jannet, H.; Flamini, G.; Harzallah-Skhiri, F.; Ali Hamza, M. Chemical Composition, Antioxidant and Anti-acetylcholinesterase activities of Tunisian Crithmum maritimum L. Essential oils. Mediterr. J. Chem. 2011, 1, 173–179. [Google Scholar] [CrossRef]
- Kubo, I.; Chen, Q.X.; Nihei, K.I. Molecular design of antibrowning agents: Antioxidative tyrosinase inhibitors. Food Chem. 2003, 81, 241–247. [Google Scholar] [CrossRef]
- Guillerme, J.B.; Couteau, C.; Coiffard, L. Applications for marine resources in cosmetics. Cosmetics 2017, 4, 35. [Google Scholar] [CrossRef] [Green Version]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvo, M.M.; Tzamourani, A.; Martínez-Alvarez, O. Halophytes as a potential source of melanosis-inhibiting compounds. Mechanism of inhibition of a characterised polyphenol extract of purslane (Portulaca oleracea). Food Chem. 2021, 355, 129649. [Google Scholar] [CrossRef]
- Georgiev, V.; Slavov, A.; Vasileva, I.; Pavlov, A. Plant cell culture as emerging technology for production of active cosmetic ingredients. Eng. Life Sci. 2018, 18, 779–798. [Google Scholar] [CrossRef]
- Ma, C.M.; Hattori, M.; Daneshtalab, M.; Wang, L. Chlorogenic acid derivatives with alkyl chains of different lengths and orientations: Potent α-glucosidase inhibitors. J. Med. Chem. 2008, 51, 6188–6194. [Google Scholar] [CrossRef]
- Wang, S.; Li, Y.; Huang, D.; Chen, S.; Xia, Y.; Zhu, S. The inhibitory mechanism of chlorogenic acid and its acylated derivatives on α-amylase and α-glucosidase. Food Chem. 2022, 372, 131334. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Q.; Zhou, F.C.; Gao, F.; Bian, J.S.; Shan, F. Comparative evaluation of quercetin, isoquercetin and rutin as inhibitors of α-glucosidase. J. Agric. Food Chem. 2009, 57, 11463–11468. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.T.; Liu, X.T.; Chen, Q.X.; Shi, Y. Lipase Inhibitors for Obesity: A Review. Biomed. Pharmacother. 2020, 128, 110314. [Google Scholar] [CrossRef]
- Rajan, L.; Palaniswamy, D.; Mohankumar, S.K. Targeting obesity with plant-derived pancreatic lipase inhibitors: A comprehensive review. Pharmacol. Res. 2020, 155, 104681. [Google Scholar] [CrossRef]
- Hulkko, L.S.S.; Rocha, R.M.; Trentin, R.; Fredsgaard, M.; Chaturvedi, T.; Custódio, L.; Thomsen, M.H. Bioactive Extracts from Salicornia ramosissima J. Woods Biorefinery as a Source of Ingredients for High-Value Industries. Plants 2023, 12, 1251. [Google Scholar] [CrossRef]
- Baysal, I.; Ekizoglu, M.; Ertas, A.; Temiz, B.; Agalar, H.G.; Yabanoglu-Ciftci, S.; Temel, H.; Ucar, G.; Turkmenoglu, F.P. Identification of Phenolic Compounds by LC-MS/MS and Evaluation of Bioactive Properties of Two Edible Halophytes: Limonium effusum and L. sinuatum. Molecules 2021, 26, 4040. [Google Scholar] [CrossRef]
- Marrelli, M.; Argentieri, M.P.; Avato, P.; Conforti, F. Lobularia maritima (L.) Desv. Aerial Parts Methanolic Extract: In Vitro Screening of Biological Activity. Plants 2020, 9, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinet, A.; Hostettmann, K.; Schutz, Y. Thermogenic effects of commercially available plant preparations aimed at treating human obesity. Phytomedicine 1999, 6, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Gan, R.Y.; Li, S.; Zhou, Y.; Li, A.N.; Xu, D.P.; Li, H.B.; Kitts, D.D. Antioxidant phytochemicals for the prevention and treatment of chronic diseases. Molecules 2015, 20, 21138–21156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D. NREL/TP-510-42622 Analytical Procedure—Determination of Ash in Biomass; National Renewable Energy Laboratory: Golden, CO, USA, 2005.
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Crocker, D. NREL/TP-510-42618 Analytical Procedure—Determination of Structural Carbohydrates and Lignin in Biomass; National Renewable Energy Laboratory: Golden, CO, USA, 2012.
- Sluiter, A.; Hames, B.; Hyman, D.; Payne, C.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Wolfe, J. NREL/TP-510-42621 Analytical Procedure—Determination of Total Solids in Biomass and Total Dissolved Solids in Liquid Process Samples; National Renewable Energy Laboratory: Golden, CO, USA, 2008; pp. 3–5.
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Velioglu, Y.S.; Mazza, G.; Gao, L.; Oomah, B.D. Antioxidant Activity and Total Phenolics in Selected Fruits, Vegetables, and Grain Products. J. Agric. Food Chem. 1998, 46, 4113–4117. [Google Scholar] [CrossRef]
- Pirbalouti, A.G.; Siahpoosh, A.; Setayesh, M.; Craker, L. Antioxidant activity, total phenolic and flavonoid contents of some medicinal and aromatic plants used as herbal teas and condiments in Iran. J. Med. Food 2014, 17, 1151–1157. [Google Scholar] [CrossRef]
- Li, Y.G.; Tanner, G.; Larkin, P. The DMACA-HCl protocol and the threshold proanthocyanidin content for bloat safety in forage legumes. J. Sci. Food Agric. 1996, 70, 89–101. [Google Scholar] [CrossRef]
- Mazza, G.; Fukumoto, L.; Delaquis, P.; Girard, B.; Ewert, B. Anthocyanins, phenolics, and color of Cabernet Franc, Merlot, and Pinot Noir wines from British Columbia. J. Agric. Food Chem. 1999, 47, 4009–4017. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT—Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorisation assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Baliga, M.S.; Jagetia, G.C.; Rao, S.K.; Babu, K. Evaluation of nitric oxide scavenging activity of certain spices in vitro: A preliminary study. Nahrung/Food 2003, 47, 261–264. [Google Scholar] [CrossRef]
- Megías, C.; Pastor-Cavada, E.; Torres-Fuentes, C.; Girón-Calle, J.; Alaiz, M.; Juan, R.; Pastor, J.; Vioque, J. Chelating, antioxidant and antiproliferative activity of Vicia sativa polyphenol extracts. Eur. Food Res. Technol. 2009, 230, 353–359. [Google Scholar] [CrossRef]
- Xiao, Z.; Storms, R.; Tsang, A. A quantitative starch-iodine method for measuring alpha-amylase and glucoamylase activities. Anal. Biochem. 2006, 351, 146–148. [Google Scholar] [CrossRef]
- Custódio, L.; Patarra, J.; Alberício, F.; da Rosa Neng, N.; Nogueira, J.M.F.; Romano, A. Phenolic composition, antioxidant potential and in vitro inhibitory activity of leaves and acorns of Quercus suber on key enzymes relevant for hyperglycemia and Alzheimer’s disease. Ind. Crops Prod. 2015, 64, 45–51. [Google Scholar] [CrossRef]
- Trentin, R.; Custódio, L.; Rodrigues, M.J.; Moschin, E.; Sciuto, K.; da Silva, J.P.; Moro, I. Exploring Ulva australis Areschoug for possible biotechnological applications: In vitro antioxidant and enzymatic inhibitory properties, and fatty acids contents. Algal Res. 2020, 50, 101980. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef] [PubMed]
- McDougall, G.J.; Kulkarni, N.N.; Stewart, D. Berry polyphenols inhibit pancreatic lipase activity in vitro. Food Chem. 2009, 115, 193–199. [Google Scholar] [CrossRef]
- AAT Bioquest Inc. Quest GraphTM IC50 Calculator. Available online: https://www.aatbio.com/tools/ic50-calculator (accessed on 1 April 2022).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction | DM [w/w %] | Water Extract [g/100 gDM] | Ethanol Extract [g/100 gDM] | n-hexane Extract [g/100 gDM] | Sugars [g/100 gDM] | Lignin [g/100 gDM] | Ash [g/100 gDM] |
|---|---|---|---|---|---|---|---|
| Sea aster (Tripolium pannonicum) | |||||||
| Juice | 4.01 (0.04) | n/a | n/a | n/a | 22.98 (0.71) | n/a | 47.71 (0.76) |
| Fibres | n/a | 18.73 (0.50) | 25.88 (2.47) | 2.87 (0.04) | 37.64 (4.56) | 16.98 (1.18) | 6.30 (0.01) |
| Sea fennel (Crithmum maritimum) | |||||||
| Juice | 7.26 (0.01) | n/a | n/a | n/a | 13.24 (3.49) | n/a | 36.56 (0.09) |
| Fibres | 35.67 (0.84) | 16.54 (0.33) | 13.56 1 | 2.20 (0.10) | 34.62 (0.67) | 18.76 (4.78) | 10.19 (0.07) |
| Fatty Acids | Sea Aster [% FA/Total FA] | Sea Fennel [% FA/Total FA] |
|---|---|---|
| Myristic acid | n.d. | 1.5 (2.2) |
| Palmitic acid | 19.0 (0.2) | 28.9 (1.8) |
| Palmitoleic acid | n.d. | 1.9 (2.7) |
| Stearic acid | 1.6 (0.0) | 6.3 (0.3) |
| Oleic acid | 1.2 (0.5) | 11.5 (0.2) |
| Linoleic acid | 24.9 (0.2) | 34.4 (1.8) |
| α-Linolenic acid | 53.2 (0.3) | 15.5 (0.7) |
| Arachidic acid | n.d. | n.d. |
| Behenic acid | n.d. | n.d. |
| Lignoceric acid | n.d. | n.d. |
| Σ SFA | 20.6 (0.2) | 36.7 (4.3) |
| Σ MUFA | 1.2 (0.5) | 13.4 (2.9) |
| Σ PUFA | 78.2 (0.5) | 49.9 (1.2) |
| ω-6/ω-3 | 0.5 (0.0) | 2.2 (0.2) |
| Fraction | TPC [mgGAE/gDM] | TFC [mgQE/gDM] | TCT [mgCE/gDM] | TAC [mgCCE/gDM] | CHL a [µg/gDM] | CHL b [µg/gDM] | TCA [µg/gDM] |
|---|---|---|---|---|---|---|---|
| Sea aster (Tripolium pannonicum) | |||||||
| Juice | n.d. | n.d. | n.d. | n.d. | 89.42 (14.56) c | 148.30 (24.23) c | 125.75 (8.85) c |
| Water extract | 32.34 (6.80) c | 5.41 (0.68) ab | n.d. | 4.37 (0.74) a | n/a | n/a | n/a |
| Ethanol extract | 45.20 (5.27) b | 6.58 (1.49) a | 0.43 (0.60) | n/a | 2614.08 (12.19) a | 1017.87 (17.94) a | 299.05 (3.79) a |
| Sea fennel (Crithmum maritimum) | |||||||
| Juice | 14.97 (4.56) d | n.d. | n.d. | n.d. | 13.62 (0.69) d | 44.28 (0.73) d | 85.34 (3.20) d |
| Water extract | 33.53 (2.53) c | 4.90 (0.81) b | n.d. | 1.90 (0.21) b | n/a | n/a | n/a |
| Ethanol extract | 64.70 (9.01) a | 1.84 (1.15) c | n.d. | n/a | 1008.44 (14.74) b | 430.92 (9.31) b | 261.79 (2.03) b |
| Fraction | DPPH | ABTS | NO | FRAP | ICA | CCA |
|---|---|---|---|---|---|---|
| Sea aster (Tripolium pannonicum) | ||||||
| Juice | n.d. | >10 | >10 | >10 | >10 | 3.18 |
| Water extract | <10 | 2.24 | >10 | 1.91 | <10 | 3.40 |
| Ethanol extract | 7.67 | 4.86 | n/a | 1.66 | 1.15 | 2.37 |
| Sea fennel (Crithmum maritimum) | ||||||
| Juice | >10 | 4.59 | >10 | 3.46 | >10 | 6.79 |
| Water extract | 3.53 | 4.36 | >10 | 1.13 | >10 | 7.03 |
| Ethanol extract | 2.84 | 3.95 | n/a | <10 | 3.10 | 3.32 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hulkko, L.S.S.; Chaturvedi, T.; Custódio, L.; Thomsen, M.H. Harnessing the Value of Tripolium pannonicum and Crithmum maritimum Halophyte Biomass through Integrated Green Biorefinery. Mar. Drugs 2023, 21, 380. https://doi.org/10.3390/md21070380
Hulkko LSS, Chaturvedi T, Custódio L, Thomsen MH. Harnessing the Value of Tripolium pannonicum and Crithmum maritimum Halophyte Biomass through Integrated Green Biorefinery. Marine Drugs. 2023; 21(7):380. https://doi.org/10.3390/md21070380
Chicago/Turabian StyleHulkko, Laura Sini Sofia, Tanmay Chaturvedi, Luísa Custódio, and Mette Hedegaard Thomsen. 2023. "Harnessing the Value of Tripolium pannonicum and Crithmum maritimum Halophyte Biomass through Integrated Green Biorefinery" Marine Drugs 21, no. 7: 380. https://doi.org/10.3390/md21070380