
Cyanobacteria: A Promising Source of Antifungal Metabolites

 , , and
, , and
Abstract
:1. Introduction
2. Methodology
Antifungal Metabolites from Cyanobacteria
3. Chemical Classes of Metabolites
3.1. Peptides
3.1.1. Peptides with β-Amino Acids
3.1.2. Peptides with Thiazole and Oxazole Rings
3.1.3. Lipoglycopeptides
3.1.4. Extracellular Peptides
3.2. Phycobiliproteins
3.3. Enzymes
3.4. Carbohydrates and Their Derivatives
3.5. Fatty Acids and Their Derivatives
3.6. Alkaloids
3.6.1. Hapalindole-Type Alkaloids
3.6.2. Tjipanazoles
3.6.3. Individual Compounds
Carriebowlinol
Norharmane
Nostocarboline
3.7. Polyketides
3.8. Macrolides
3.9. Phenolic Compounds
3.10. Other
4. Major Targets
4.1. Cell Membrane
4.2. Cytoskeleton
4.3. Other Targets
5. Regulation
5.1. Light
5.2. Nutrients
5.3. pH
5.4. Temperature
6. Final Considerations
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kendrick, B. Fungi: Ecological Importance and Impact on Humans. eLS 2011, 1–5. [Google Scholar] [CrossRef]
- Friedman, D.Z.P.; Schwartz, I.S. Emerging Fungal Infections: New Patients, New Patterns, and New Pathogens. J. Fungi 2019, 5, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lockhart, S.R.; Guarner, J. Emerging and Reemerging Fungal Infections. Semin. Diagn. Pathol. 2019, 36, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Jenks, J.D.; Cornely, O.A.; Chen, S.C.A.; Thompson, G.R.; Hoenigl, M. Breakthrough Invasive Fungal Infections: Who Is at Risk? Mycoses 2020, 63, 1021–1032. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Solache, M.A.; Casadevall, A. Global Warming Will Bring New Fungal Diseases for Mammals. mBio 2010, 1, e00061-10. [Google Scholar] [CrossRef] [Green Version]
- Fausto, A.; Rodrigues, M.L.; Coelho, C. The Still Underestimated Problem of Fungal Diseases Worldwide. Front. Microbiol. 2019, 10, 214. [Google Scholar] [CrossRef] [Green Version]
- Pianalto, K.M.; Alspaugh, J.A. New Horizons in Antifungal Therapy. J. Fungi. 2016, 2, 26. [Google Scholar] [CrossRef] [Green Version]
- Rauseo, A.M.; Coler-reilly, A.; Larson, L.; Spec, A. Hope on the Horizon: Novel Fungal Treatments in Development. Open. Forum. Infect. Dis. 2020, 7, ofaa016. [Google Scholar] [CrossRef] [Green Version]
- Perera, I.; Subashchandrabose, S.R.; Venkateswarlu, K.; Naidu, R.; Megharaj, M. Consortia of Cyanobacteria/Microalgae and Bacteria in Desert Soils: An Underexplored Microbiota. Appl. Microbiol. Biotechnol. 2018, 102, 7351–7363. [Google Scholar] [CrossRef]
- Patel, A.; Matsakas, L.; Rova, U.; Christakopoulos, P. A Perspective on Biotechnological Applications of Thermophilic Microalgae and Cyanobacteria. Bioresour. Technol. 2019, 278, 424–434. [Google Scholar] [CrossRef]
- Welker, M.; Von Döhren, H. Cyanobacterial Peptides—Nature’s Own Combinatorial Biosynthesis. FEMS Microbiol. Rev. 2006, 30, 530–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morone, J.; Lopes, G.; Preto, M.; Vasconcelos, V.; Martins, R. Exploitation of Filamentous and Picoplanktonic Cyanobacteria for Cosmetic Applications: Potential to Improve Skin Structure and Preserve Dermal Matrix Components. Mar. Drugs 2020, 18, 486. [Google Scholar] [CrossRef] [PubMed]
- Calteau, A.; Fewer, D.P.; Latifi, A.; Coursin, T.; Laurent, T.; Jokela, J.; Kerfeld, C.A.; Sivonen, K.; Piel, J.; Gugger, M. Phylum-Wide Comparative Genomics Unravel the Diversity of Secondary Metabolism in Cyanobacteria. BMC Genom. 2014, 15, 977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shishido, T.K.; Humisto, A.; Jokela, J.; Liu, L.; Wahlsten, M.; Tamrakar, A.; Fewer, D.P.; Permi, P.; Andreote, A.P.D.; Fiore, M.F.; et al. Antifungal Compounds from Cyanobacteria. Mar. Drugs 2015, 13, 2124–2140. [Google Scholar] [CrossRef] [Green Version]
- Nowruzi, B.; Porzani, S.J. Toxic Compounds Produced by Cyanobacteria Belonging to Several Species of the Order Nostocales: A Review. J. Appl. Toxicol. 2021, 41, 510–548. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Singh, P.; Tripathi, J.; Srivastava, A.; Tripathi, M.K.; Ravi, A.K.; Asthana, R.K. Identification and structure elucidation of antimicrobial compounds from Lyngbya aestuarii and Aphanothece bullosa. Cell. Mol. Biol. 2014, 60, 82–89. [Google Scholar] [PubMed]
- Moosmann, P.; Ueoka, R.; Gugger, M.; Piel, J. Aranazoles: Extensively Chlorinated Nonribosomal Peptide-Polyketide Hybrids from the Cyanobacterium Fischerella sp. PCC 9339. Org. Lett. 2018, 20, 5238–5241. [Google Scholar] [CrossRef]
- Zervou, S.K.; Gkelis, S.; Kaloudis, T.; Hiskia, A.; Mazur-Marzec, H. New Microginins from Cyanobacteria of Greek Freshwaters. Chemosphere 2020, 248, 125961. [Google Scholar] [CrossRef]
- Swain, S.S.; Paidesetty, S.K.; Padhy, R.N. Antibacterial, Antifungal and Antimycobacterial Compounds from Cyanobacteria. Biomed. Pharmacother. 2017, 90, 760–776. [Google Scholar] [CrossRef]
- Ali Shah, S.A.; Akhter, N.; Auckloo, B.N.; Khan, I.; Lu, Y.; Wang, K.; Wu, B.; Guo, Y.W. Structural Diversity, Biological Properties and Applications of Natural Products from Cyanobacteria. A Review. Mar. Drugs 2017, 15, 354. [Google Scholar] [CrossRef] [Green Version]
- Asimakis, E.; Shehata, A.A.; Eisenreich, W.; Acheuk, F.; Lasram, S.; Basiouni, S.; Emekci, M.; Ntougias, S.; Taner, G.; May-Simera, H.; et al. Algae and Their Metabolites as Potential Bio-Pesticides. Microorganisms 2022, 10, 307. [Google Scholar] [CrossRef] [PubMed]
- Fewer, D.P.; Jokela, J.; Heinilä, L.; Aesoy, R.; Sivonen, K.; Galica, T.; Hrouzek, P.; Herfindal, L. Chemical Diversity and Cellular Effects of Antifungal Cyclic Lipopeptides from Cyanobacteria. Physiol. Plant. 2021, 173, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Leão, P.N.; Engene, N.; Antunes, A.; Gerwick, W.H.; Vasconcelos, V. The Chemical Ecology of Cyanobacteria. Nat. Prod. Rep. 2012, 29, 372–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerphagnon, M.; Latour, D.; Colombet, J.; Sime-Ngando, T. Fungal Parasitism: Life Cycle, Dynamics and Impact on Cyanobacterial Blooms. PLoS ONE 2013, 8, e60894. [Google Scholar] [CrossRef] [Green Version]
- Amaral, S.C.D.; Monteiro, P.R.; Neto, J.d.S.P.; Serra, G.M.; Gonçalves, E.C.; Xavier, L.P.; Santos, A.V. Current Knowledge on Microviridin from Cyanobacteria. Mar. Drugs 2021, 19, 17. [Google Scholar] [CrossRef] [PubMed]
- Rohrlack, T.; Christiansen, G.; Kurmayer, R. Putative Antiparasite Defensive System Involving Ribosomal and Nonribosomal Oligopeptides in Cyanobacteria of the Genus Planktothrix. Appl. Environ. Microbiol. 2013, 79, 2642–2647. [Google Scholar] [CrossRef] [Green Version]
- Demay, J.; Bernard, C.; Reinhardt, A.; Marie, B. Natural Products from Cyanobacteria: Focus on Beneficial Activities. Mar. Drugs 2019, 17, 320. [Google Scholar] [CrossRef] [Green Version]
- Vasas, G.; Borbely, G.; Nanasi, P.; Nanasi, P.P. Alkaloids from Cyanobacteria with Diverse Powerful Bioactivities. Mini. Rev. Med. Chem. 2010, 10, 946–955. [Google Scholar] [CrossRef]
- Chen, M.Y.; Teng, W.K.; Zhao, L.; Hu, C.X.; Zhou, Y.K.; Han, B.P.; Song, L.R.; Shu, W.S. Comparative Genomics Reveals Insights into Cyanobacterial Evolution and Habitat Adaptation. ISME J. 2021, 15, 211–227. [Google Scholar] [CrossRef]
- Jones, M.R.; Pinto, E.; Torres, M.A.; Dörr, F.; Mazur-Marzec, H.; Szubert, K.; Tartaglione, L.; Dell’Aversano, C.; Miles, C.O.; Beach, D.G.; et al. CyanoMetDB, a Comprehensive Public Database of Secondary Metabolites from Cyanobacteria. Water Res. 2021, 196, 117017. [Google Scholar] [CrossRef]
- Monteiro, P.R.; Do Amaral, S.C.; Siqueira, A.S.; Xavier, L.P.; Santos, A.V. Anabaenopeptins: What We Know so Far. Toxins 2021, 13, 522. [Google Scholar] [CrossRef]
- Cabrele, C.; Martinek, T.A.; Reiser, O.; Berlicki, Ł. Peptides Containing β-Amino Acid Patterns: Challenges and Successes in Medicinal Chemistry. J. Med. Chem. 2014, 57, 9718–9739. [Google Scholar] [CrossRef] [PubMed]
- Pergament, I.; Carmeli, S. Schizotrin A; a Novel Antimicrobial Cyclic Peptide from a Cyanobacterium. Tetrahedron Lett. 1994, 35, 8473–8476. [Google Scholar] [CrossRef]
- An, T.; Kumar, T.K.S.; Wang, M.; Liu, L.; Lay, J.O.; Liyanage, R.; Berry, J.; Gantar, M.; Marks, V.; Gawley, R.E.; et al. Structures of Pahayokolides A and B, Cyclic Peptides from a Lyngbya sp. J. Nat. Prod. 2007, 70, 730–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leão, P.N.; Pereira, A.R.; Liu, W.T.; Ng, J.; Pevzner, P.A.; Dorrestein, P.C.; König, G.M.; Vasconcelos, V.M.; Gerwick, W.H. Synergistic Allelochemicals from a Freshwater Cyanobacterium. Proc. Natl. Acad. Sci. USA 2010, 107, 11183–11188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, S.S.; Lu Chen, J.; Moore, R.E.; Patterson, G.M.L. Calophycin, a Fungicidal Cyclic Decapeptide from the Terrestrial Blue-Green Alga Calothrix fusca. J. Org. Chem. 1992, 57, 1097–1103. [Google Scholar] [CrossRef]
- Gregson, J.M.; Chen, J.; Patterson, G.M.L.; Moore, R.E. Structures of Puwainaphycins A–E. Tetrahedron 1992, 48, 3727–3734. [Google Scholar] [CrossRef]
- Berry, J.P.; Gantar, M.; Gawley, R.E.; Wang, M.; Rein, K.S. Pharmacology and Toxicology of Pahayokolide A, a Bioactive Metabolite from a Freshwater Species of Lyngbya Isolated from the Florida Everglades. Comp. Biochem. Physiol. C. Toxicol. Pharmacol. 2004, 139, 231–238. [Google Scholar] [CrossRef] [Green Version]
- Tomek, P.; Hrouzek, P.; Kuzma, M.; Sýkora, J.; Fišer, R.; Černý, J.; Novák, P.; Bártová, S.; Šimek, P.; Hof, M.; et al. Cytotoxic Lipopeptide Muscotoxin A, Isolated from Soil Cyanobacterium Desmonostoc muscorum, Permeabilizes Phospholipid Membranes by Reducing Their Fluidity. Chem. Res. Toxicol. 2016, 28, 216–224. [Google Scholar] [CrossRef]
- Cheel, J.; Hájek, J.; Kuzma, M.; Saurav, K.; Smýkalová, I.; Ondráčková, E.; Urajová, P.; Vu, D.L.; Faure, K.; Kopecký, J.; et al. Application of HPCCC Combined with Polymeric Resins and HPLC for the Separation of Cyclic Lipopeptides Muscotoxins A-C and Their Antimicrobial Activity. Molecules 2018, 23, 2653. [Google Scholar] [CrossRef] [Green Version]
- Mareš, J.; Jek, J.H.; Urajová, P.; Kopecký, J.; Hrouzek, P. A Hybrid Non-Ribosomal Peptide/Polyketide Synthetase Containing Fatty-Acyl Ligase (Faal) Synthesizes the b- Amino Fatty Acid Lipopeptides Puwainaphycins in the Cyanobacterium Cylindrospermum alatosporum. PLoS ONE 2014, 9, e111904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, L.; Morais, J.; Preto, M.; Silva, R.; Urbatzka, R.; Vasconcelos, V.; Reis, M. Uncovering the Bioactive Potential of a Cyanobacterial Natural Products Library Aided by Untargeted Metabolomics. Mar. Drugs 2021, 19, 633. [Google Scholar] [CrossRef] [PubMed]
- Hájek, J.; Bieringer, S.; Voráčová, K.; Macho, M.; Saurav, K.; Delawská, K.; Divoká, P.; Fišer, R.; Mikušová, G.; Cheel, J.; et al. Semi-Synthetic Puwainaphycin/Minutissamide Cyclic Lipopeptides with Improved Antifungal Activity and Limited Cytotoxicity. RSC Adv. 2021, 11, 30873–30886. [Google Scholar] [CrossRef]
- Kang, H.S.; Krunic, A.; Shen, Q.; Swanson, S.M.; Orjala, J. Minutissamides A-D, Antiproliferative Cyclic Decapeptides from the Cultured Cyanobacterium Anabaena minutissima. J. Nat. Prod. 2011, 74, 1597–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kajiyama, S.I.; Kanzaki, H.; Kawazu, K.; Kobayashi, A. Nostofungicidine, an Antifungal Lipopeptide from the Field-Grown Terrestrial Blue-Green Alga Nostoc commune. Tetrahedron Lett. 1998, 39, 3737–3740. [Google Scholar] [CrossRef]
- Jokela, J.; Oftedal, L.; Herfindal, L.; Permi, P.; Wahlsten, M.; Døskeland, S.O.; Sivonen, K. Anabaenolysins, Novel Cytolytic Lipopeptides from Benthic Anabaena Cyanobacteria. PLoS ONE 2012, 7, e41222. [Google Scholar] [CrossRef] [Green Version]
- Shishido, T.K.; Jokela, J.; Kolehmainen, C.T.; Fewer, D.P.; Wahlsten, M.; Wang, H.; Rouhiainen, L.; Rizzi, E.; De Bellis, G.; Permic, P.; et al. Antifungal Activity Improved by Coproduction of Cyclodextrins and Anabaenolysins in Cyanobacteria. Proc. Natl. Acad. Sci. USA 2015, 112, 13669–13674. [Google Scholar] [CrossRef] [Green Version]
- Frankmölle, W.P.; Knübel, G.; Moore, R.E.; Patterson, G.M. Antifungal Cyclic Peptides from the Terrestrial Blue-Green Alga Anabaena laxa. J. Antibiot. 1992, 45, 1458–1466. [Google Scholar] [CrossRef] [Green Version]
- Cai, W.; Matthew, S.; Chen, Q.; Paul, V.J.; Luesch, H. Discovery of New A- and B-Type Laxaphycins with Synergistic Anticancer Activity. Bioorg. Med. Chem. 2018, 26, 2310–2319. [Google Scholar] [CrossRef]
- Bonnard, I.; Rolland, M.; Francisco, C.; Banaigs, B. Total Structure and Biological Properties of Laxaphycins A and B, Cyclic Lipopeptides from the Marine Cyanobacterium Lyngbya majuscula. Int. J. Pept. Res. Ther. 1997, 4, 289–292. [Google Scholar] [CrossRef]
- Bonnard, I.; Rolland, M.; Salmon, J.; Debiton, E.; Barthomeuf, C.; Banaigs, B. Total Structure and Inhibition of Tumor Cell Proliferation of Laxaphycins. J. Med. Chem. 2007, 50, 1266–1279. [Google Scholar] [CrossRef]
- Heinilä, L.M.P.; Fewer, D.P.; Jokela, J.K.; Wahlsten, M.; Jortikka, A.; Sivonen, K. Shared PKS Module in Biosynthesis of Synergistic Laxaphycins. Front. Microbiol. 2020, 11, 578878. [Google Scholar] [CrossRef] [PubMed]
- Heinilä, L.M.P.; Fewer, D.P.; Jokela, J.K.; Wahlsten, M.; Ouyang, X.; Permi, P.; Jortikka, A.; Sivonen, K. The Structure and Biosynthesis of Heinamides A1–A3 and B1–B5, Antifungal Members of the Laxaphycin Lipopeptide Family. Org. Biomol. Chem. 2021, 19, 5577–5588. [Google Scholar] [CrossRef]
- Gerwick, W.H.; Jiang, Z.D.; Agarwal, S.K.; Farmer, B.T. Total Structure of Hormothamnin A, A Toxic Cyclic Undecapeptide from the Tropical Marine Cyanobacterium Hormothamnion enteromorphoides. Tetrahedron 1992, 48, 2313–2324. [Google Scholar] [CrossRef]
- Meickle, T.; Matthew, S.; Ross, C.; Luesch, H.; Paul, V. Bioassay-Guided Isolation and Identification of Desacetylmicrocolin B from Lyngbya Cf. Polychroa. Planta Med. 2009, 75, 1427–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koehn, F.E.; Longley, R.E.; Reed, J.K. Microcolins A and B, New Immunosuppressive Peptides from the Blue-Green Alga Lyngbya majuscula. J. Nat. Prod. 1992, 55, 613–619. [Google Scholar] [CrossRef]
- Yu, H.B.; Glukhov, E.; Li, Y.; Iwasaki, A.; Gerwick, L.; Dorrestein, P.C.; Jiao, B.H.; Gerwick, W.H. Cytotoxic Microcolin Lipopeptides from the Marine Cyanobacterium Moorea producens. J. Nat. Prod. 2019, 82, 2608–2619. [Google Scholar] [CrossRef]
- Engene, N.; Choi, H.; Esquenazi, E.; Byrum, T.; Villa, F.A.; Cao, Z.; Murray, T.F.; Dorrestein, P.C.; Gerwick, L.; Gerwick, W.H. Phylogeny-Guided Isolation of Ethyl Tumonoate A from the Marine Cyanobacterium Cf. Oscillatoria Margaritifera. J. Nat. Prod. 2011, 74, 1737–1743. [Google Scholar] [CrossRef] [Green Version]
- Macmillan, J.B.; Ernst-russell, M.A.; Ropp, J.S.D.; Molinski, T.F. Lobocyclamides A–C, Lipopeptides from a Cryptic Cyanobacterial Mat Containing Lyngbya confervoides. J. Org. Chem. 2002, 271, 8210–8215. [Google Scholar] [CrossRef] [PubMed]
- Macmillan, J.B.; Molinski, T.F. Lobocyclamide B from Lyngbya confervoides. Configuration and Asymmetric Synthesis of -Hydroxy- r -Amino Acids by (−)-Sparteine-Mediated Aldol Addition. Org. Lett. 2002, 4, 1883–1886. [Google Scholar] [CrossRef]
- Jüttner, F.; Todorova, A.K.; Walch, N.; von Philipsborn, W. Nostocyclamide M: A Cyanobacterial Cyclic Peptide with Allelopathic Activity from Nostoc sp. 31. Phytochemistry 2001, 57, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Todorova, A.K.; Juettner, F.; Linden, A.; Pluess, T.; von Philipsborn, W. Nostocyclamide: A New Macrocyclic, Thiazole-Containing Allelochemical from Nostoc sp. 31 (Cyanobacteria). J. Org. Chem. 1995, 60, 7891–7895. [Google Scholar] [CrossRef]
- Ramaswamy, A.V.; Sorrels, C.M.; Gerwick, W.H. Cloning and Biochemical Characterization of the Hectochlorin Biosynthetic Gene Cluster from the Marine Cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2007, 70, 1977–1986. [Google Scholar] [CrossRef] [PubMed]
- Kleigrewe, K.; Gerwick, L.; Sherman, D.H.; Gerwick, W.H. Unique Marine Derived Cyanobacterial Biosynthetic Genes for Chemical Diversity. Nat. Prod. Rep. 2016, 33, 348–364. [Google Scholar] [CrossRef] [Green Version]
- Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J.; Mooberry, S.L. Isolation, Structure Determination, and Biological Activity of Lyngbyabellin A from the Marine Cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2000, 63, 611–615. [Google Scholar] [CrossRef]
- Thornburg, C.C.; Cowley, E.S.; Sikorska, J.; Shaala, L.A.; Ishmael, J.E.; Youssef, D.T.A.; McPhail, K.L. Apratoxin H and Apratoxin A Sulfoxide from the Red Sea Cyanobacterium Moorea producens. J. Nat. Prod. 2013, 76, 1781–1788. [Google Scholar] [CrossRef] [Green Version]
- Neuhof, T.; Schmieder, P.; Seibold, M.; Preussel, K.; von Döhren, H. Hassallidin B-Second Antifungal Member of the Hassallidin Family. Bioorg. Med. Chem. Lett. 2006, 16, 4220–4222. [Google Scholar] [CrossRef]
- Neuhof, T.; Schmieder, P.; Preussel, K.; Dieckmann, R.; Pham, H.; Bartl, F.; Von Döhren, H. Hassallidin A, a Glycosylated Lipopeptide with Antifungal Activity from the Cyanobacterium Hassallia Sp. J. Nat. Prod. 2005, 68, 695–700. [Google Scholar] [CrossRef]
- Vestola, J.; Shishido, T.K.; Jokela, J.; Fewer, D.P.; Aitio, O.; Permi, P.; Wahlsten, M.; Wang, H.; Rouhiainen, L.; Sivonen, K. Hassallidins, Antifungal Glycolipopeptides, are Widespread among Cyanobacteria and are the End-Product of a Nonribosomal Pathway. Proc. Natl. Acad. Sci. USA 2014, 111, 1909–1917. [Google Scholar] [CrossRef] [Green Version]
- Pancrace, C.; Jokela, J.; Sassoon, N.; Ganneau, C.; Desnos-Ollivier, M.; Wahlsten, M.; Humisto, A.; Calteau, A.; Bay, S.; Fewer, D.P.; et al. Rearranged Biosynthetic Gene Cluster and Synthesis of Hassallidin E in Planktothrix serta PCC 8927. ACS Chem. Biol. 2017, 12, 1796–1804. [Google Scholar] [CrossRef] [Green Version]
- Bui, T.H.; Wray, V.; Nimtz, M.; Fossen, T.; Preisitsch, M.; Schröder, G.; Wende, K.; Heiden, S.E.; Mundt, S. Balticidins A-D, Antifungal Hassallidin-like Lipopeptides from the Baltic Sea Cyanobacterium Anabaena cylindrica Bio33. J. Nat. Prod. 2014, 77, 1287–1296. [Google Scholar] [CrossRef] [PubMed]
- Jaki, B.; Zerbe, O.; Heilmann, J.; Sticher, O. Two Novel Cyclic Peptides with Antifungal Activity from the Cyanobacterium Tolypothrix byssoidea (EAWAG 195). J. Nat. Prod. 2001, 64, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Weiss, C.; Figueras, E.; Borbely, A.N.; Sewald, N. Cryptophycins: Cytotoxic Cyclodepsipeptides with Potential for Tumor Targeting. J. Pept. Sci. 2017, 23, 514–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, C.D.; Zhang, X.; Mooberry, S.L.; Patterson, G.M.L.; Moore, R.E. Cryptophycin: A New Antimicrotubule Agent Active against Drug-Resistant Cells. Cancer Res. 1994, 54, 3779–3784. [Google Scholar] [PubMed]
- Neuhof, T.; Seibold, M.; Thewes, S.; Laue, M.; Han, C.O.; Hube, B.; von Döhren, H. Comparison of Susceptibility and Transcription Profile of the New Antifungal Hassallidin A with Caspofungin. Biochem. Biophys. Res. Commun. 2006, 349, 740–749. [Google Scholar] [CrossRef]
- Marquez, B.L.; Watts, K.S.; Yokochi, A.; Roberts, M.A.; Verdier-Pinard, P.; Jimenez, J.I.; Hamel, E.; Scheuer, P.J.; Gerwick, W.H. Structure and Absolute Stereochemistry of Hectochlorin, a Potent Stimulator of Actin Assembly. J. Nat. Prod. 2002, 65, 866–871. [Google Scholar] [CrossRef]
- Gerwick, W.H.; Mrozek, C.; Moghaddam, M.F.; Agarwal, S.K. Novel Cytotoxic Peptides from the Tropical Marine CyanobacteriumHormothamnion Enteromorphoides 1. Discovery, Isolation and Initial Chemical and Biological Characterization of the Hormothamnins from Wild and Cultured Material. Experientia 1989, 45, 115–121. [Google Scholar] [CrossRef]
- Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J. Isolation and Structure of the Cytotoxin Lyngbyabellin B and Absolute Configuration of Lyngbyapeptin a from the Marine Cyanobaeterium Lyngbya majuscula. J. Nat. Prod. 2000, 63, 1437–1439. [Google Scholar] [CrossRef]
- Mareš, J.; Hájek, J.; Urajová, P.; Kust, A.; Jokela, J.; Saurav, K.; Galica, T.; Čapková, K.; Mattila, A.; Haapaniemi, E.; et al. Alternative Biosynthetic Starter Units Enhance the Structural Diversity of Cyanobacterial Lipopeptides. Appl. Environ. Microbiol. 2019, 85, e02675-18. [Google Scholar] [CrossRef] [Green Version]
- Brilisauer, K.; Rapp, J.; Rath, P.; Schöllhorn, A.; Bleul, L.; Weiß, E.; Stahl, M.; Grond, S.; Forchhammer, K. Cyanobacterial Antimetabolite 7-Deoxy-Sedoheptulose Blocks the Shikimate Pathway to Inhibit the Growth of Prototrophic Organisms. Nat. Commun. 2019, 10, 545. [Google Scholar] [CrossRef] [Green Version]
- Belhaj, D.; Frikha, D.; Athmouni, K.; Jerbi, B.; Ahmed, M.B.; Bouallagui, Z.; Kallel, M.; Maalej, S.; Zhou, J.; Ayadi, H. Box-Behnken Design for Extraction Optimization of Crude Polysaccharides from Tunisian Phormidium versicolor Cyanobacteria (NCC 466): Partial Characterization, in Vitro Antioxidant and Antimicrobial Activities. Int. J. Biol. Macromol. 2017, 105, 1501–1510. [Google Scholar] [CrossRef] [PubMed]
- Righini, H.; Baraldi, E.; Fernández, Y.G.; Quintana, A.M.; Roberti, R. Different Antifungal Activity of Anabaena sp., Ecklonia sp., and Jania sp. Against Botrytis Cinerea. Mar. Drugs 2019, 17, 15–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diao, Y.; Han, H.; Li, Y.; Zhou, J.; Yang, Z. Extraction, Infrared Spectral Analysis and the Antimicrobial Activity on Polysaccharide within Nostoc commune Vauch. Int. Proc. Chem. Biol. Environ. Eng. 2013, 51, 6. [Google Scholar] [CrossRef]
- Gutiérrez-Del-Río, I.; de Fraissinette, N.B.; Castelo-Branco, R.; Oliveira, F.; Morais, J.; Redondo-Blanco, S.; Villar, C.J.; JoséIglesias, M.; Soengas, R.; Cepas, V.; et al. Chlorosphaerolactylates A−D: Natural Lactylates of Chlorinated Fatty Acids Isolated from the Cyanobacterium Sphaerospermopsis sp. Lege 00249. J. Nat. Prod. 2020, 83, 1885–1890. [Google Scholar] [CrossRef] [PubMed]
- MacMillan, J.B.; Molinski, T.F. Majusculoic Acid, a Brominated Cyclopropyl Fatty Acid from a Marine Cyanobacterial Mat Assemblage. J. Nat. Prod. 2005, 68, 604–606. [Google Scholar] [CrossRef]
- Carmeli, S.; Moore, R.E.; Patterson, G.M.L.; Mori, Y.; Suzuki, M. Isonitriles from the Blue-Green Alga Scytonema mirabile. J. Org. Chem. 1990, 55, 4431–4438. [Google Scholar] [CrossRef]
- Soares, A.R.; Engene, N.; Gunasekera, S.P.; Sneed, J.M.; Paul, V.J. Carriebowlinol, an Antimicrobial Tetrahydroquinolinol from an Assemblage of Marine Cyanobacteria Containing a Novel Taxon. J. Nat. Prod. 2015, 78, 534–538. [Google Scholar] [CrossRef]
- Singh, I.P.; Milligan, K.E.; Gerwick, W.H. Tanikolide, a Toxic and Antifungal Lactone from the Marine Cyanobacterium Lyngbya majuscula. J. Nat. Prod. 1999, 62, 1333–1335. [Google Scholar] [CrossRef]
- Smitka, T.A.; Bonjouklian, R.; Doolin, L.; Jones, N.D.; Deeter, J.B.; Yoshida, W.Y.; Prinsep, M.R.; Moore, R.E.; Patterson, G.M.L. Ambiguine Isonitriles, Fungicidal Hapalindole-Type Alkaloids from Three Genera of Blue-Green Algae Belonging to the Stigonemataceae. J. Org. Chem. 1992, 57, 857–861. [Google Scholar] [CrossRef]
- Raveh, A.; Carmeli, S. Antimicrobial Ambiguines from the Cyanobacterium Fischerella sp. Collected in Israel. J. Nat. Prod. 2007, 70, 196–201. [Google Scholar] [CrossRef]
- Mo, S.; Krunic, A.; Santarsiero, B.D.; Franzblau, S.G.; Orjala, J. Hapalindole-Related Alkaloids from the Cultured Cyanobacterium Fischerella ambigua. Phytochemistry 2010, 71, 2116–2123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, S.; Krunic, A.; Chlipala, G.; Orjala, J. Antimicrobial Ambiguine Isonitriles from the Cyanobacterium Fischerella ambigua. J. Nat. Prod. 2009, 72, 894–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Lantvit, D.; Hwang, C.H.; Kroll, D.J.; Swanson, S.M.; Franzblau, S.G.; Orjala, J. Indole Alkaloids from Two Cultured Cyanobacteria, Westiellopsis sp. and Fischerella muscicola. Bioorg. Med. Chem. 2012, 20, 5290–5295. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Hillwig, M.L.; Doi, Y.; Liu, X. Aliphatic Halogenase Enables Late-Stage C-H Functionalization: Selective Synthesis of a Brominated Fischerindole Alkaloid with Enhanced Antibacterial Activity. ChemBioChem 2016, 17, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Moore, Richard Elliott Patterson Leon, G.M. Hapalindoles. EP 0171283A2, 6 August 1985.
- Locher, H.H.; Ritz, D.; Pfaff, P.; Gaertner, M.; Knezevic, A.; Sabato, D.; Schroeder, S.; Barbaras, D.; Gademann, K. Dimers of Nostocarboline with Potent Antibacterial Activity. Chemotherapy 2010, 56, 318–324. [Google Scholar] [CrossRef] [Green Version]
- Volk, R.B.; Furkert, F.H. Antialgal, Antibacterial and Antifungal Activity of Two Metabolites Produced and Excreted by Cyanobacteria during Growth. Microbiol. Res. 2006, 161, 180–186. [Google Scholar] [CrossRef]
- Bonjouklian, R.; Smitka, T.A.; Doolin, L.E.; Molloy, R.M.; Debono, M.; Shaffer, S.A.; Moore, R.E.; Stewart, J.B.; Patterson, G.M.L. Tjipanazoles, New Antifungal Agents from the Blue-Green Alga Tolypothrix tjipanasensis. Tetrahedron 1991, 47, 7739–7750. [Google Scholar] [CrossRef]
- Stratmann, K.; Moore, R.E.; Patterson, G.M.L.; Bonjouklian, R.; Deeter, J.B.; Shaffer, S.; Smitka, T.A.; Smith, C.D. Welwitindolinones, Unusual Alkaloids from the Blue-Green Algae Hapalosiphon welwitschii and Westiella intricata. Relationship to Fischerindoles and Hapalinodoles. J. Am. Chem. Soc. 1994, 116, 9935–9942. [Google Scholar] [CrossRef]
- Gross, E.M.; Wolk, C.P.; Juttner, F. Fischerellin, A New Allelochemical From The Freshwater Cyanobacterium Fischerella muscicola. J. Phycol. 1991, 27, 686–692. [Google Scholar] [CrossRef] [Green Version]
- Bertin, M.J.; Demirkiran, O.; Navarro, G.; Moss, N.A.; Lee, J.; Goldgof, G.M.; Vigil, E.; Winzeler, E.A.; Valeriote, F.A.; Gerwick, W.H. Kalkipyrone B, a Marine Cyanobacterial γ-Pyrone Possessing Cytotoxic and Anti-Fungal Activities. Phytochemistry 2016, 122, 113–118. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.; Kang, H.S.; Krunic, A.; Chlipala, G.E.; Cai, G.; Chen, W.L.; Franzblau, S.G.; Swanson, S.M.; Orjala, J. Carbamidocyclophanes F and G with Anti-Mycobacterium Tuberculosis Activity from the Cultured Freshwater Cyanobacterium Nostoc sp. Tetrahedron Lett. 2014, 55, 686–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishibashi, M.; Moore, R.E.; Patterson, G.M.; Xu, C.; Clardy, J. Scytophycins, Cytotoxic and Antimycotic Agents from the Cyanophyte Scytonema pseudohofmanni. J. Org. Chem. 1986, 51, 5300–5306. [Google Scholar] [CrossRef]
- Patterson, G.M.L.; Carmeli, S. Biological Effects of Tolytoxin (6-Hydroxy-7-O-Methyl-Scytophycin b), a Potent Bioactive Metabolite from Cyanobacteria. Arch. Microbiol. 1992, 157, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Carmely, S.; Kashman, Y. Structure of Swinholide-a, a New Macrolide from the Marine Sponge Theonella swinhoei. Tetrahedron Lett. 1985, 26, 511–514. [Google Scholar] [CrossRef]
- Salvador-Reyes, L.A.; Sneed, J.; Paul, V.J.; Luesch, H. Amantelides A and B, Polyhydroxylated Macrolides with Differential Broad-Spectrum Cytotoxicity from a Guamanian Marine Cyanobacterium. J. Nat. Prod. 2015, 78, 1957–1962. [Google Scholar] [CrossRef] [Green Version]
- Oku, N.; Hana, S.; Matsumoto, M.; Yonejima, K.; Tansei, K.; Isogai, Y.; Igarashi, Y. Two New Sacrolide-Class Oxylipins from the Edible Cyanobacterium Aphanothece sacrum. J. Antibiot. 2017, 70, 708–709. [Google Scholar] [CrossRef] [Green Version]
- Oku, N.; Matsumoto, M.; Yonejima, K.; Tansei, K.; Igarashi, Y. Sacrolide A, a New Antimicrobial and Cytotoxic Oxylipin Macrolide from the Edible Cyanobacterium Aphanothece sacrum. Beilstein J. Org. Chem. 2014, 10, 1808–1816. [Google Scholar] [CrossRef]
- Wright, A.D.; Papendorf, O.; König, G.M. Ambigol C and 2,4-Dichlorobenzoic Acid, Natural Products Produced by the Terrestrial Cyanobacterium Fischerella ambigua. J. Nat. Prod. 2005, 68, 459–461. [Google Scholar] [CrossRef]
- Ghasemi, Y.; Tabatabaei Yazdi, M.; Shafiee, A.; Amini, M.; Shokravi, S.; Zarrini, G. Parsiguine, a Novel Antimicrobial Substance from Fischerella ambigua. Pharm. Biol. 2004, 42, 318–322. [Google Scholar] [CrossRef]
- Mo, S.; Krunic, A.; Pegan, S.D.; Franzblau, S.G.; Orjala, J. An Antimicrobial Guanidine-Bearing Sesterterpene from the Cultured Cyanobacterium Scytonema sp. J. Nat. Prod. 2009, 72, 2043–2045. [Google Scholar] [CrossRef] [Green Version]
- Moore, R.E.; Bornemann, V.; Niemczura, W.P.; Gregson, J.M.; Chen, J.; Norton, T.R.; Patterson, G.M.L.; Helms, G.L. Puwainaphycin C, a cardioactive cyclic peptide from the blue-green alga Anabaena BQ-16-1. Use of two-dimensional carbon-13-carbon-13 and carbon-13-nitrogen-15 correlation spectroscopy in sequencing the amino acid units. J. Am. Chem. Soc. 1989, 9, 6128–6132. [Google Scholar] [CrossRef]
- Halary, S.; Duperron, S.; Kim Tiam, S.; Duval, C.; Dhenaim, E.; Bernard, C.; Marie, B. Unexpected Micro-Spatial Scale Genomic Diversity of the Bloom-Forming Cyanobacterium Aphanizomenon Gracile and Its Phycosphere. PrePrint 2021, 1–31. [Google Scholar]
- Shishido, T.K.; Popin, R.V.; Jokela, J.; Wahlsten, M.; Fiore, M.F.; Fewer, D.P.; Herfindal, L.; Sivonen, K. Dereplication of Natural Products with Antimicrobial and Anticancer Activity from Brazilian Cyanobacteria. Toxins 2019, 12, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urajová, P.; Hájek, J.; Wahlsten, M.; Jokela, J.; Galica, T.; Fewer, D.P.; Kust, A.; Zapomělová-Kozlíková, E.; Delawská, K.; Sivonen, K.; et al. A Liquid Chromatography-Mass Spectrometric Method for the Detection of Cyclic β-Amino Fatty Acid Lipopeptides. J. Chromatogr. A 2016, 1438, 76–83. [Google Scholar] [CrossRef]
- Cheel, J.; Urajová, P.; Hájek, J.; Hrouzek, P.; Kuzma, M.; Bouju, E.; Faure, K.; Kopecký, J. Separation of Cyclic Lipopeptide Puwainaphycins from Cyanobacteria by Countercurrent Chromatography Combined with Polymeric Resins and HPLC. Anal. Bioanal. Chem. 2017, 409, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.T. Marine Cyanobacteria: A Treasure Trove of Bioactive Secondary Metabolites for Drug Discovery. In Studies in Natural Products Chemistry; Elsevier, B.V.: Amsterdam, The Netherlands, 2012; Volume 232, pp. 67–110. ISBN 9780444538369. [Google Scholar]
- Frankmölle, W.P.; Larsen, L.K.; Caplan, F.R.; Patterson, G.M.L.; Knubel, G.; Levine, I.A.; Moore, R.E. Blue-Green Alga Anabaena laxa I. Isolation and Biological Properties. J. Antibiot. 1992, 45, 1451–1457. [Google Scholar] [CrossRef]
- Pennings, S.C.; Pablo, S.R.; Paul, V.J. Chemical Defenses of the Tropical, Benthic Marine Cyanobacterium Hormothamnion enteromorphoides: Diverse Consumers and Synergisms. Limnol. Oceanogr. 1997, 42, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Dussault, D.; Dang, K.; Vansach, T.; Horgen, F.D.; Lacroix, M. Antimicrobial Effects of Marine Algal Extracts and Cyanobacterial Pure Compounds against Five Foodborne Pathogens. Food Chem. 2016, 199, 114–118. [Google Scholar] [CrossRef]
- Bornancin, L.; Boyaud, F.; Mahiout, Z.; Bonnard, I.; Mills, S.C.; Banaigs, B.; Inguimbert, N. Isolation and Synthesis of Laxaphycin B—Type Peptides: A Case Study and Clues to Their Biosynthesis. Mar. Drugs 2015, 13, 7285–7300. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.; Kang, H.; Krunic, A.; Chen, W.; Yang, J.; Woodard, J.L.; Fuchs, J.R.; Hyun, S.; Franzblau, S.G.; Swanson, S.M.; et al. Bioorganic & Medicinal Chemistry Trichormamides C and D, Antiproliferative Cyclic Lipopeptides from the Cultured Freshwater Cyanobacterium Cf. Oscillatoria Sp. UIC 10045. Bioorg. Med. Chem. 2015, 23, 3153–3162. [Google Scholar] [CrossRef] [Green Version]
- Bornancin, L.; Alonso, E.; Alvariño, R.; Inguimbert, N.; Bonnard, I.; Botana, L.M.; Banaigs, B. Bioorganic & Medicinal Chemistry Structure and Biological Evaluation of New Cyclic and Acyclic Laxaphycin-A Type Peptides. Bioorg. Med. Chem. 2019, 27, 1966–1980. [Google Scholar] [CrossRef]
- Alvariño, R.; Alonso, E.; Bornancin, L.; Bonnard, I.; Inguimbert, N.; Banaigs, B.; Botana, L.M. Biological Activities of Cyclic and Acyclic B-Type Laxaphycins in SH-SY5Y Human Neuroblastoma Cells. Mar. Drugs 2020, 18, 364. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, P.; Krunic, A.; Burdette, J.E.; Orjala, J. Laxaphycins B5 and B6 from the Cultured Cyanobacterium UIC 10484. J. Antibiot. 2020, 73, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Gerwick, W.H.; Tan, L.I.K.T. Nitrogen-Containing Metabolites from Marine Cyanobacteria. Alkaloids Chem. Biol. 2001, 57, 75–184. [Google Scholar] [CrossRef]
- Guerrero-Pepinosa, N.Y.; Cardona-Trujillo, M.C.; Garzón-Castaño, S.C.; Veloza, L.A.; Sepúlveda-Arias, J.C. Antiproliferative Activity of Thiazole and Oxazole Derivatives: A Systematic Review of in Vitro and in Vivo Studies. Biomed. Pharmacother. 2021, 138, 111495. [Google Scholar] [CrossRef]
- Mhlongo, J.T.; Brasil, E.; de la Torre, B.G.; Albericio, F. Naturally Occurring Oxazole-Containing Peptides. Mar. Drugs 2020, 18, 203. [Google Scholar] [CrossRef] [Green Version]
- Urda, C.; Fern, R.; Rodr, J.; Marta, P.; Jim, C.; Cuevas, C. Bistratamides M and N, Oxazole-Thiazole Containing Cyclic Hexapeptides Isolated from Lissoclinum bistratum Interaction of Zinc (II) with Bistratamide K. Mar. Drugs 2017, 15, 209. [Google Scholar] [CrossRef] [Green Version]
- Todorova, A.; Jüttner, F. Ecotoxicological Analysis of Nostocyclamide, a Modified Cyclic Hexapeptide from Nostoc. Phycologia 1996, 35, 183–188. [Google Scholar] [CrossRef]
- Vishwakarma, R.; Rai, A.K. Separation of Bioactive Metabolites from Aphanothece halophytica Through HPLC and Characterization of the Analytes Through ESI-MS and NMR. Nat. Prod. J. 2013, 3, 151–157. [Google Scholar] [CrossRef]
- Cetusic, J.R.P.; Green, F.R.; Graupner, P.R.; Oliver, M.P. Total Synthesis of Hectochlorin. Org. Lett. 2002, 4, 1307–1310. [Google Scholar] [CrossRef]
- Hai, Y.; Wei, M.Y.; Wang, C.Y.; Gu, Y.C.; Shao, C.L. The Intriguing Chemistry and Biology of Sulfur-Containing Natural Products from Marine Microorganisms (1987–2020). Mar. Life Sci. Technol. 2021, 3, 488–518. [Google Scholar] [CrossRef]
- Petitbois, J.G.; Casalme, L.O.; Lopez, J.A.V.; Alarif, W.M.; Abdel-Lateff, A.; Al-Lihaibi, S.S.; Yoshimura, E.; Nogata, Y.; Umezawa, T.; Matsuda, F.; et al. Serinolamides and Lyngbyabellins from an Okeania sp. Cyanobacterium Collected from the Red Sea. J. Nat. Prod. 2017, 80, 2708–2715. [Google Scholar] [CrossRef]
- Sweeney-Jones, A.M.; Gagaring, K.; Antonova-Koch, J.; Zhou, H.; Mojib, N.; Soapi, K.; Skolnick, J.; McNamara, C.W.; Kubanek, J. Antimalarial Peptide and Polyketide Natural Products from the Fijian Marine Cyanobacterium Moorea producens. Mar. Drugs 2020, 18, 167. [Google Scholar] [CrossRef]
- Fathoni, I.; Petitbois, J.G.; Alarif, W.M.; Abdel-Lateff, A.; Al-Lihaibi, S.S.; Yoshimura, E.; Nogata, Y.; Vairappan, C.S.; Sholikhah, E.N.; Okino, T. Bioactivities of Lyngbyabellins from Cyanobacteria of Moorea and Okeania Genera. Molecules 2020, 25, 3986. [Google Scholar] [CrossRef] [PubMed]
- Zimba, P.V.; Shalygin, S.; Huang, I.S.; Momčilović, M.; Abdulla, H. A New Boring Toxin Producer–Perforafilum tunnelli Gen. & sp. Nov. (Oscillatoriales, Cyanobacteria) Isolated from Laguna Madre, Texas, USA. Phycologia 2021, 60, 10–24. [Google Scholar] [CrossRef]
- Han, B.; McPhail, K.L.; Gross, H.; Goeger, D.E.; Mooberry, S.L.; Gerwick, W.H. Isolation and Structure of Five Lyngbyabellin Derivatives from a Papua New Guinea Collection of the Marine Cyanobacterium Lyngbya majuscula. Tetrahedron 2005, 61, 11723–11729. [Google Scholar] [CrossRef]
- Fuentes-Valdés, J.J.; Soto-Liebe, K.; Pérez-Pantoja, D.; Tamames, J.; Belmar, L.; Pedrós-Alió, C.; Garrido, D.; Vásquez, M. Draft Genome Sequences of Cylindrospermopsis raciborskii strains CS-508 and MVCC14, Isolated from Freshwater Bloom Events in Australia and Uruguay. Stand. Genom. Sci. 2018, 13, 26. [Google Scholar] [CrossRef] [PubMed]
- Österholm, J.; Popin, R.V.; Fewer, D.P.; Sivonen, K. Phylogenomic Analysis of Secondary Metabolism in the Toxic Cyanobacterial Genera Anabaena, Dolichospermum and Aphanizomenon. Toxins 2020, 12, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pancrace, C.; Barny, M.A.; Ueoka, R.; Calteau, A.; Scalvenzi, T.; Pédron, J.; Barbe, V.; Piel, J.; Humbert, J.F.; Gugger, M. Insights into the Planktothrix Genus: Genomic and Metabolic Comparison of Benthic and Planktic Strains. Sci. Rep. 2017, 7, 41181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Ye, X.; Chen, L.; Zhao, H.; Shi, Q.; Xiao, Y.; Ma, L.; Hou, X.; Chen, Y.; Yang, F. Functional Role of Bloom-Forming Cyanobacterium Planktothrix in Ecologically Shaping Aquatic Environments. Sci. Total Environ. 2020, 710, 136314. [Google Scholar] [CrossRef]
- Popin, R.V.; Alvarenga, D.O.; Castelo-Branco, R.; Fewer, D.P.; Sivonen, K. Mining of Cyanobacterial Genomes Indicates Natural Product Biosynthetic Gene Clusters Located in Conjugative Plasmids. Front. Microbiol. 2021, 12, 684565. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Sivonen, K.; Rouhiainen, L.; Fewer, D.P.; Lyra, C.; Rantala-Ylinen, A.; Vestola, J.; Jokela, J.; Rantasärkkä, K.; Li, Z.; et al. Genome-Derived Insights into the Biology of the Hepatotoxic Bloom-Forming Cyanobacterium Anabaena sp. strain 90. BMC Genom. 2012, 13, 613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crnkovic, C.M.; May, D.S.; Orjala, J. The Impact of Culture Conditions on Growth and Metabolomic Profiles of Freshwater Cyanobacteria. J. Appl. Phycol. 2018, 30, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Abreu, V.A.C.; Popin, R.V.; Alvarenga, D.O.; Schaker, P.D.C.; Hoff-Risseti, C.; Varani, A.M.; Fiore, M.F. Genomic and Genotypic Characterization of Cylindrospermopsis raciborskii: Toward an Intraspecific Phylogenetic Evaluation by Comparative Genomics. Front. Microbiol. 2018, 9, 306. [Google Scholar] [CrossRef] [PubMed]
- Freitas, S.; Castelo-Branco, R.; Wenzel-Storjohann, A.; Vasconcelos, V.M.; Tasdemir, D.; Leão, P.N. Structure and Biosynthesis of Desmamides A–C, Lipoglycopeptides from the Endophytic Cyanobacterium Desmonostoc muscorum LEGE 12446. J. Nat. Prod. 2022, 85, 1704–1714. [Google Scholar] [CrossRef]
- Leão, P.N.; Vasconcelos, M.T.S.D.; Vasconcelos, V.M. Allelopathy in Freshwater Cyanobacteria Allelopathy in Freshwater Cyanobacteria. Crit. Rev. Microbiol. 2009, 35, 271–282. [Google Scholar] [CrossRef]
- Hirsch, C.F.; Liesch, J.M.; Salvatore, M.J.; Schwartz, R.E.; Sesin, D.F. Antifungal Fermentation Product and Method. US Patent 4946835A, 15 July 1988. [Google Scholar]
- Trimurtulu, G.; Ohtani, I.; Patterson, G.M.L.; Moore, R.E.; Corbett, T.H.; Valeriote, F.A.; Demchik, L. Total Structures of Cryptophycins, Potent Antitumor Depsipeptides from the Blue-Green Alga Nostoc sp. strain GSV 224. J. Am. Chem. Soc. 1994, 116, 4729–4737. [Google Scholar] [CrossRef]
- Schwartz, R.E.; Hirsch, C.F.; Sesin, D.F.; Flor, J.E.; Chartrain, M.; Fromtling, R.E.; Harris, G.H.; Salvatore, M.J.; Liesch, J.M.; Yudin, K. Pharmaceuticals from Cultured Algae. J. Ind. Microbiol. 1990, 5, 113–123. [Google Scholar] [CrossRef]
- Golakoti, T.; Ogino, J.; Heltzel, C.E.; Husebo, T.L.; Jensen, C.M.; Larsen, L.K.; Patterson, G.M.L.; Moore, R.E.; Mooberry, S.L.; Corbett, T.H.; et al. Structure Determination, Conformational Analysis, Chemical Stability Studies, and Antitumor Evaluation of the Cryptophycins. Isolation of 18 New Analogs from Nostoc sp. strain GSV 224. J. Am. Chem. Soc. 1995, 117, 12030–12049. [Google Scholar] [CrossRef]
- Martinelli, M.J.; Vaidyanathan, R.; Khau, V.V.; Staszak, M.A. Reaction of Cryptophycin 52 with Thiols. Tetrahedron Lett. 2002, 43, 3365–3367. [Google Scholar] [CrossRef]
- Chaganty, S.; Golakoti, T.; Heltzel, C.; Moore, R.E.; Yoshida, W.Y. Isolation and Structure Determination of Cryptophycins 38, 326, and 327 from the Terrestrial Cyanobacterium Nostoc sp. GSV 224. J. Nat. Prod. 2004, 67, 1403–1406. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Mevers, E.; Byrumb, T.; Valeriote, F.A.; Gerwick, W.H. Lyngbyabellins K-N from Two Palmyra Atoll Collections of the Marine Cyanobacterium Moorea bouillonii. Eur. J. Org. Chem. 2012, 27, 5141–5150. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.K.; Bischoff, A. Asymmetric Syntheses of Potent Antitumor Macrolides Cryptophycin B and Arenastatin A. Eur. J. Org. Chem. 2004, 2004, 2131–2141. [Google Scholar] [CrossRef] [PubMed]
- Nowruzi, B.; Khavari-Nejad, R.-A.; Sivonen, K.; Kazemi, B.; Najafi, F.; Nejadsattari, T. Identification and Toxigenic Potential of a Nostoc sp. ALGAE 2012, 27, 303–313. [Google Scholar] [CrossRef]
- Pagels, F.; Guedes, A.C.; Amaro, H.M.; Kijjoa, A.; Vasconcelos, V. Phycobiliproteins from Cyanobacteria: Chemistry and Biotechnological Applications. Biotechnol. Adv. 2019, 37, 422–443. [Google Scholar] [CrossRef]
- Pagels, F.; Pereira, R.N.; Vicente, A.A.; Guedes, A.C. Extraction of Pigments from Microalgae and Cyanobacteria-a Review on Current Methodologies. Appl. Sci. 2021, 11, 5187. [Google Scholar] [CrossRef]
- Nowruzi, B.; Fahimi, H.; Lorenzi, A.S. Recovery of Pure C-Phycoerythrin from a Limestone Drought Tolerant Cyanobacterium Nostoc sp. and Evaluation of Its Biological Activity. An. Biol. 2020, 42, 115–128. [Google Scholar] [CrossRef]
- Murugan, T.; Radhamadhavan, T. Screening for Antifungal and Antiviral Activity of C-Phycocyanin from Spirulina platensis. J. Pharm. Res. 2011, 4, 4161–4163. [Google Scholar]
- Najdenski, H.M.; Gigova, L.G.; Iliev, I.I.; Pilarski, P.S.; Lukavský, J.; Tsvetkova, I.V.; Ninova, M.S.; Kussovski, V.K. Antibacterial and Antifungal Activities of Selected Microalgae and Cyanobacteria. Int. J. Food Sci. Technol. 2013, 48, 1533–1540. [Google Scholar] [CrossRef]
- Digamber Rao, B.; Shamitha, G.; Renuka, G.; Ramesh Babu, M. Action of C-Phycocyanin Pigment and Cell Extracts of Tolypothrix sp. on the Biochemical Activity of Eri Silkworm and Their Antifungal Activity. Nat. Environ. Pollut. Technol. 2011, 10, 351–356. [Google Scholar]
- Hemlata; Afreen, S.; Fatma, T. Extraction, Purification and Characterization of Phycoerythrin from Michrochaete and Its Biological Activities. Biocatal. Agric. Biotechnol. 2018, 13, 84–89. [Google Scholar] [CrossRef]
- Righini, H.; Francioso, O.; Di Foggia, M.; Quintana, A.M.; Roberti, R. Preliminary Study on the Activity of Phycobiliproteins against Botrytis cinerea. Mar. Drugs 2020, 18, 600. [Google Scholar] [CrossRef] [PubMed]
- Schlumbaum, A.; Mauch, F.; Vögeli, U.; Boller, T. Plant Chitinases Are Potent Inhibitors of Fungal Growth. Nature 1986, 324, 365–367. [Google Scholar] [CrossRef]
- Veliz, E.A.; Martínez-Hidalgo, P.; Hirsch, A.M. Chitinase-Producing Bacteria and Their Role in Biocontrol. AIMS Microbiol. 2017, 3, 689–705. [Google Scholar] [CrossRef]
- Xie, X.H.; Fu, X.; Yan, X.Y.; Peng, W.F.; Kang, L.X. A Broad-Specificity Chitinase from Penicillium Oxalicum K10 Exhibits Antifungal Activity and Biodegradation Properties of Chitin. Mar. Drugs 2021, 19, 356. [Google Scholar] [CrossRef]
- Urrutia-Cordero, P.; Agha, R.; Cirés, S.; Lezcano, M.á.; Sánchez-Contreras, M.; Waara, K.O.; Utkilen, H.; Quesada, A. Effects of Harmful Cyanobacteria on the Freshwater Pathogenic Free-Living Amoeba Acanthamoeba castellanii. Aquat. Toxicol. 2013, 130–131, 9–17. [Google Scholar] [CrossRef]
- Agha, R.; Quesada, A. Oligopeptides as Biomarkers of Cyanobacterial Subpopulations. Toward an Understanding of Their Biological Role. Toxins 2014, 6, 1929–1950. [Google Scholar] [CrossRef] [Green Version]
- Lima, S.L.; Colombo, A.L.; de Almeida Junior, J.N. Fungal Cell Wall: Emerging Antifungals and Drug Resistance. Front. Microbiol. 2019, 10, 2573. [Google Scholar] [CrossRef] [Green Version]
- Ibe, C.; Munro, C.A. Fungal Cell Wall: An Underexploited Target for Antifungal Therapies. PLoS Pathog. 2021, 17, e1009470. [Google Scholar] [CrossRef]
- Gonzalez, M.; de Groot, P.W.J.; Klis, F.M.; Lipke, P.N. Glycoconjugate Structure and Function in Fungal Cell Walls. In Microbial Glycobiology; Elsevier: Amsterdam, The Netherlands, 2010; pp. 169–183. ISBN 9780123745460. [Google Scholar]
- Bowman, S.M.; Free, S.J. The Structure and Synthesis of the Fungal Cell Wall. BioEssays 2006, 28, 799–808. [Google Scholar] [CrossRef]
- Ait-Lahsen, H.; Soler, A.; Rey, M.; De La Cruz, J.; Monte, E.; Llobell, A. An Antifungal Exo-α-1,3-Glucanase (AGN13.1) from the Biocontrol Fungus Trichoderma harzianum. Appl. Environ. Microbiol. 2001, 67, 5833–5839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baiyee, B.; Ito, S.i.; Sunpapao, A. Trichoderma asperellum T1 Mediated Antifungal Activity and Induced Defense Response against Leaf Spot Fungi in Lettuce (Lactuca Sativa L.). Physiol. Mol. Plant. Pathol. 2019, 106, 96–101. [Google Scholar] [CrossRef]
- Chen, H.; Ju, H.; Wang, Y.; Du, G.; Yan, X.; Cui, Y.; Yuan, Y.; Yue, T. Antifungal Activity and Mode of Action of Lactic Acid Bacteria Isolated from Kefir against Penicillium expansum. Food Control. 2021, 130, 108274. [Google Scholar] [CrossRef]
- Natarajan, C.; Gupta, V.; Kumar, K.; Prasanna, R. Molecular Characterization of a Fungicidal Endoglucanase from the Cyanobacterium Calothrix elenkinii. Biochem. Genet. 2013, 51, 766–779. [Google Scholar] [CrossRef]
- Gupta, V.; Natarajan, C.; Kumar, K.; Prasanna, R. Identification and Characterization of Endoglucanases for Fungicidal Activity in Anabaena laxa (Cyanobacteria). J. Appl. Phycol. 2011, 23, 73–81. [Google Scholar] [CrossRef]
- Gupta, V.; Prasanna, R.; Chaudhary, V.; Nain, L. Biochemical, Structural and Functional Characterization of Two Novel Antifungal Endoglucanases from Anabaena laxa. Biocatal. Agric. Biotechnol. 2012, 1, 338–347. [Google Scholar] [CrossRef]
- Zhang, X.; Yuan, J.; Li, F.; Xiang, J. Chitin Synthesis and Degradation in Crustaceans: A Genomic View and Application. Mar. Drugs 2021, 19, 153. [Google Scholar] [CrossRef]
- Ali, M.A.; Ren, H.; Ahmed, T.; Luo, J.; An, Q.; Qi, X.; Li, B. Antifungal Effects of Rhizospheric Bacillus Species Against Bayberry Twig Blight Pathogen Pestalotiopsis Versicolor. Agronomy 2020, 10, 1811. [Google Scholar] [CrossRef]
- Poveda, J.; Francisco, M.; Cartea, M.E.; Velasco, P. Development of Transgenic Brassica Crops against Biotic Stresses Caused |by Pathogens and Arthropod Pests. Plants 2020, 9, 1664. [Google Scholar] [CrossRef]
- Filyushin, M.A.; Anisimova, O.K.; Kochieva, E.Z.; Shchennikova, A.V. Genome-wide Identification and Expression of Chitinase Class I Genes in Garlic (Allium sativum l.) Cultivars Resistant and Susceptible to Fusarium proliferatum. Plants 2021, 10, 720. [Google Scholar] [CrossRef]
- Jiang, X.; Chen, D.; Chen, L.; Yang, G.; Zou, S. Purification, Characterization, and Action Mode of a Chitosanase from Streptomyces roseolus Induced by Chitin. Carbohydr. Res. 2012, 355, 40–44. [Google Scholar] [CrossRef]
- Prasanna, R.; Nain, L.; Tripathi, R.; Gupta, V.; Chaudhary, V.; Middha, S.; Joshi, M.; Ancha, R.; Kaushik, B.D. Evaluation of Fungicidal Activity of Extracellular Filtrates of Cyanobacteria—Possible Role of Hydrolytic Enzymes. J. Basic. Microbiol. 2008, 48, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Prasanna, R.; Natarajan, C.; Srivastava, A.K.; Sharma, J. Identification, Characterization, and Regulation of a Novel Antifungal Chitosanase Gene (Cho) in Anabaena sp. Appl. Environ. Microbiol. 2010, 76, 2769–2777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, V.; Prasanna, R.; Srivastava, A.K.; Sharma, J. Purification and Characterization of a Novel Antifungal Endo-Type Chitosanase from Anabaena Fertilissima. Ann. Microbiol. 2012, 62, 1089–1098. [Google Scholar] [CrossRef]
- Ahmad, N.H.; Mustafa, S.; Man, Y.B.C. Microbial Polysaccharides and Their Modification Approaches: A Review. Int. J. Food Prop. 2015, 18, 332–347. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Guo, H.; Yi, W.; Song, J. Current Understanding on Biosynthesis of Microbial Polysaccharides. Curr. Top. Med. Chem. 2008, 8, 141–151. [Google Scholar] [CrossRef]
- Cano, M.; Holland, S.C.; Artier, J.; Burnap, R.L.; Ghirardi, M.; Morgan, J.A.; Yu, J. Glycogen Synthesis and Metabolite Overflow Contribute to Energy Balancing in Cyanobacteria. Cell. Rep. 2018, 23, 667–672. [Google Scholar] [CrossRef]
- Bhatnagar, M.; Bhatnagar, A. Diversity of Polysaccharides in Cyanobacteria. In Microbial Diversity in Ecosystem Sustainability and Biotechnological Applications; Springer: Singapore, 2019; pp. 447–496. ISBN 1557527318. [Google Scholar]
- Maeda, K.; Okuda, Y.; Enomoto, G.; Watanabe, S.; Ikeuchi, M. Biosynthesis of a Sulfated Exopolysaccharide, Synechan, and Bloom Formation in the Model Cyanobacterium Synechocystis Sp. Strain Pcc 6803. Elife 2021, 10, e66538. [Google Scholar] [CrossRef]
- Parwani, L.; Bhatt, M.; Singh, J. Potential Biotechnological Applications of Cyanobacterial Exopolysaccharides. Braz. Arch. Biol. Technol. 2021, 64, 1–13. [Google Scholar] [CrossRef]
- Cruz, D.; Vasconcelos, V.; Pierre, G.; Michaud, P.; Delattre, C. Exopolysaccharides from Cyanobacteria: Strategies for Bioprocess Development. Appl. Sci. 2020, 10, 3763. [Google Scholar] [CrossRef]
- Pierre, G.; Sopena, V.; Juin, C.; Mastouri, A.; Graber, M.; Maugard, T. Antibacterial Activity of a Sulfated Galactan Extracted from the Marine Alga Chaetomorpha aerea against Staphylococcus aureus. Biotechnol. Bioprocess. Eng. 2011, 16, 937–945. [Google Scholar] [CrossRef]
- Klarzynski, O.; Plesse, B.; Joubert, J.M.; Yvin, J.C.; Kopp, M.; Kloareg, B.; Fritig, B. Linear β-1,3 Glucans are Elicitors of Defense Responses in Tobacco. Plant. Physiol. 2000, 124, 1027–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapp, J.; Rath, P.; Kilian, J.; Brilisauer, K.; Grond, S.; Forchhammer, K. A Bioactive Molecule Made by Unusual Salvage of Radical SAM Enzyme Byproduct 5-Deoxyadenosine Blurs the Boundary of Primary and Secondary Metabolism. J. Biol. Chem. 2021, 296, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, S.A.C.; Preto, M.; Moreira, G.; Martins, T.P.; Abt, K.; Melo, A.; Vasconcelos, V.M.; Leão, P.N. Discovery of Cyanobacterial Natural Products Containing Fatty Acid Residues. Angew. Chem. Int. Ed. 2021, 60, 10064–10072. [Google Scholar] [CrossRef]
- Zheng, C.J.; Yoo, J.S.; Lee, T.G.; Cho, H.Y.; Kim, Y.H.; Kim, W.G. Fatty Acid Synthesis is a Target for Antibacterial Activity of Unsaturated Fatty Acids. FEBS Lett. 2005, 579, 5157–5162. [Google Scholar] [CrossRef] [Green Version]
- Kwan, J.C.; Meickle, T.; Ladwa, D.; Teplitski, M.; Paul, V.; Luesch, H. Lyngbyoic Acid, a “Tagged” Fatty Acid from a Marine Cyanobacterium, Disrupts Quorum Sensing in P. aeruginosa. Mol. Biosyst. 2011, 7, 1205–1216. [Google Scholar] [CrossRef]
- Abt, K.; Castelo-Branco, R.; Leão, P.N. Biosynthesis of Chlorinated Lactylates in Sphaerospermopsis sp. LEGE 00249. J. Nat. Prod. 2021, 84, 278–286. [Google Scholar] [CrossRef]
- Taylor, S.L.; Hefle, S.L. Naturally Occurring Toxicants in Foods. In Foodborne Diseases, 3rd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. 327–344. ISBN 9780123850072. [Google Scholar]
- Qiu, S.; Sun, H.; Zhang, A.H.; Xu, H.Y.; Yan, G.L.; Han, Y.; Wang, X.J. Natural Alkaloids: Basic Aspects, Biological Roles, and Future Perspectives. Chin. J. Nat. Med. 2014, 12, 401–406. [Google Scholar] [CrossRef]
- Walton, K.; Berry, J.P. Indole Alkaloids of the Stigonematales (Cyanophyta): Chemical Diversity, Biosynthesis, and Biological Activity. Mar. Drugs 2016, 14, 73. [Google Scholar] [CrossRef] [Green Version]
- Hohlman, R.M.; Sherman, D.H. Recent Advances in Hapalindole-Type Cyanobacterial Alkaloids: Biosynthesis, Synthesis, and Biological Activity. Nat. Prod. Rep. 2021, 38, 1567–1588. [Google Scholar] [CrossRef]
- Awakawa, T.; Abe, I. Molecular Basis for the Plasticity of Aromatic Prenyltransferases in Hapalindole Biosynthesis. Beilstein J. Org. Chem. 2019, 15, 1545–1551. [Google Scholar] [CrossRef]
- Wang, J.; Chen, C.C.; Yang, Y.; Liu, W.; Ko, T.P.; Shang, N.; Hu, X.; Xie, Y.; Huang, J.W.; Zhang, Y.; et al. Structural Insight into a Novel Indole Prenyltransferase in Hapalindole-Type Alkaloid Biosynthesis. Biochem. Biophys. Res. Commun. 2018, 495, 1782–1788. [Google Scholar] [CrossRef]
- Hillwig, M.L.; Zhu, Q.; Liu, X. Biosynthesis of Ambiguine Indole Alkaloids in Cyanobacterium Fischerella ambigua. ACS Chem. Biol. 2014, 9, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Bhat, V.; Dave, A.; MacKay, J.A.; Rawal, V.H. The Chemistry of Hapalindoles, Fischerindoles, Ambiguines, and Welwitindolinones. In Alkaloids: Chemistry and Biology; Elsevier Inc.: Amsterdam, The Netherlands, 2014; Volume 73, pp. 65–160. ISBN 9780124115651. [Google Scholar]
- Acuña, U.M.; Zi, J.; Orjala, J.; Carcache de Blanco, E.J. Ambiguine I Isonitrile from Fischerella ambigua Induces Caspase-Independent Cell Death in MCF-7 Hormone Dependent Breast Cancer Cells. Int. J. Cancer Res. 2015, 49, 1655–1662. [Google Scholar]
- Moore, R.E.; Cheuk, C.; Patterson, G.M.L. Hapalindoles: New Alkaloids from the Blue-Green Alga Hapalosiphon fontinalis. J. Am. Chem. Soc. 1984, 106, 6456–6457. [Google Scholar] [CrossRef]
- Huber, U.; Moore, R.E.; Patterson, G.M.L. Isolation of a Nitrile-Containing Indole Alkaloid from the Terrestrial Blue-Green Alga Hapalosiphon delicatulus. J. Nat. Prod. 1998, 61, 1304–1306. [Google Scholar] [CrossRef] [PubMed]
- Koodkaew, I.; Sunohara, Y.; Matsuyama, S.; Matsumoto, H. Isolation of Ambiguine D Isonitrile from Hapalosiphon sp. and Characterization of Its Phytotoxic Activity. Plant. Growth Regul. 2012, 68, 141–150. [Google Scholar] [CrossRef]
- Walton, K.; Gantar, M.; Gibbs, P.D.L.; Schmale, M.C.; Berry, J.P. Indole Alkaloids from Fischerella Inhibit Vertebrate Development in the Zebrafish (Danio rerio) Embryo Model. Toxins 2014, 6, 3568–3581. [Google Scholar] [CrossRef] [Green Version]
- Bonjouklian, R.; Moore, R.E.; Patterson, G.M.L.; Smitka, T.A. Hapalindole Alkaloids as Antifungal and Antitumor Agents. European Patent Office EP0543516A1, 22 November 1992. [Google Scholar]
- Volk, R.B. Screening of Microalgae for Species Excreting Norharmane, a Manifold Biologically Active Indole Alkaloid. Microbiol. Res. 2008, 163, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, Z.A. Allelopathic Activity of the Norharmane-Producing Cyanobacterium Synechocystis aquatilis against Cyanobacteria and Microalgae. Oceanol. Hydrobiol. Stud. 2013, 42, 1–7. [Google Scholar] [CrossRef]
- Avendano, C.; Menendez, J. Synthetic Studies on N-Methylwelwitindolinone C Isothiocyanate (Welwistatin) and Related Substructures. Curr. Org. Synth. 2005, 1, 65–82. [Google Scholar] [CrossRef]
- Brailsford, J.A.; Lauchli, R.; Shea, K.J. Synthesis of the Bicyclic Welwitindolinone Core via an Alkylation/Cyclization Cascade Reaction. Org. Lett. 2009, 11, 5330–5333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chilczuk, T.; Schäberle, T.F.; Vahdati, S.; Mettal, U.; El Omari, M.; Enke, H.; Wiese, M.; König, G.M.; Niedermeyer, T.H.J. Halogenation-Guided Chemical Screening Provides Insight into Tjipanazole Biosynthesis by the Cyanobacterium Fischerella ambigua. ChemBioChem 2020, 21, 2170–2177. [Google Scholar] [CrossRef] [PubMed]
- Voldoire, A.; Moreau, P.; Sancelme, M.; Matulova, M.; Léonce, S.; Pierré, A.; Hickman, J.; Pfeiffer, B.; Renard, P.; Dias, N.; et al. Analogues of Antifungal Tjipanazoles from Rebeccamycin. Bioorg. Med. Chem. 2004, 12, 1955–1962. [Google Scholar] [CrossRef] [Green Version]
- Jung, P.; D’agostino, P.M.; Büdel, B.; Lakatos, M. Symphyonema bifilamentata Sp. Nov., the Right Fischerella ambigua 108b: Half a Decade of Research on Taxonomy and Bioactive Compounds in New Light. Microorganisms 2021, 9, 745. [Google Scholar] [CrossRef]
- Bourke, C.A.; Stevens, G.R.; Carrigan, M.J. Locomotor Effects in Sheep of Alkaloids Identified in Australian Tribulus terrestris. Aust. Vet. J. 1992, 69, 163–165. [Google Scholar] [CrossRef]
- Kodani, S.; Imoto, A.; Mitsutani, A.; Murakami, M. Isolation and Identification of the Antialgal Compound from Algicidal Bacterium Pseudomonas sp. K44-1. Fish. Sci. 2002, 68, 623–624. [Google Scholar] [CrossRef] [Green Version]
- Inoue, S.; Okada, K.; Tanino, H.; Kakoi, H.; Goto, T. Trace Characterization of the Fluorescent Substances of a Dinoflagellate, Noctiluca miliaris. Chem. Lett. 1980, 9, 297–298. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Hafez, S.I.I.; Abo-Elyousr, K.A.M.; Abdel-Rahim, I.R. Fungicidal Activity of Extracellular Products of Cyanobacteria against Alternaria porri. Eur. J. Phycol. 2015, 50, 239–245. [Google Scholar] [CrossRef]
- Becher, P.G.; Beuchat, J.; Gademann, K.; Jüttner, F. Nostocarboline: Isolation and Synthesis of a New Cholinesterase Inhibitor from Nostoc 78-12A. J. Nat. Prod. 2005, 68, 1793–1795. [Google Scholar] [CrossRef]
- Becher, P.G.; Baumann, H.I.; Gademann, K.; Jüttner, F. The Cyanobacterial Alkaloid Nostocarboline: An Inhibitor of Acetylcholinesterase and Trypsin. J. Appl. Phycol. 2009, 21, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Blom, J.F.; Brütsch, T.; Barbaras, D.; Bethuel, Y.; Locher, H.H.; Hubschwerlen, C.; Gademann, K. Potent Algicides Based on the Cyanobacterial Alkaloid Nostocarboline. Org. Lett. 2006, 8, 737–740. [Google Scholar] [CrossRef]
- Gademann, K. Cyanobacterial Natural Products for the Inhibition of Biofilm Formation and Biofouling. Chimia (Aarau) 2007, 61, 373–377. [Google Scholar] [CrossRef] [Green Version]
- Gademann, K. Natural Product Hybrids. Chimia (Aarau) 2006, 60, 841–845. [Google Scholar] [CrossRef] [Green Version]
- Lim, L.; McFadden, G.I. The Evolution, Metabolism and Functions of the Apicoplast. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 749–763. [Google Scholar] [CrossRef] [Green Version]
- Barbaras, D.; Kaiser, M.; Brun, R.; Gademann, K. Potent and Selective Antiplasmodial Activity of the Cyanobacterial Alkaloid Nostocarboline and Its Dimers. Bioorg. Med. Chem. Lett. 2008, 18, 4413–4415. [Google Scholar] [CrossRef] [Green Version]
- Graber, M.A.; Gerwick, W.H. Graber, Gerwick, 1998, Kalkipyrone, a Toxic γ -Pyrone from an Assemblage of the Marine Cyanobacteria Lyngbya majuscula and Tolypothrix sp. J. Nat. Prod. 1998, 16, 677–680. [Google Scholar] [CrossRef]
- Koyama, T.; Kawazoe, Y.; Iwasaki, A.; Ohno, O.; Suenaga, K.; Uemura, D. Anti-Obesity Activities of the Yoshinone A and the Related Marine γ-Pyrone Compounds. J. Antibiot. 2016, 69, 348–351. [Google Scholar] [CrossRef]
- Shibata, T. Asymmetric Cycloaddition Reactions. In Comprehensive Inorganic Chemistry II.; Elsevier: Amsterdam, The Netherlands, 2013; Volume 6, pp. 249–269. ISBN 9780080965291. [Google Scholar]
- Bui, H.T.N.; Jansen, R.; Pham, H.T.L.; Mundt, S. Carbamidocyclophanes A−E, Chlorinated Paracyclophanes with Cytotoxic and Antibiotic Activity from the Vietnamese Cyanobacterium Nostoc sp. J. Nat. Prod. 2007, 70, 499–503. [Google Scholar] [CrossRef]
- Preisitsch, M.; Harmrolfs, K.; Pham, H.T.L.; Heiden, S.E.; Füssel, A.; Wiesner, C.; Pretsch, A.; Swiatecka-Hagenbruch, M.; Niedermeyer, T.H.J.; Müller, R.; et al. Anti-MRSA-Acting Carbamidocyclophanes H-L from the Vietnamese Cyanobacterium Nostoc sp. CAVN2. J. Antibiot. 2015, 68, 165–177. [Google Scholar] [CrossRef]
- Preisitsch, M.; Bui, H.T.N.; Bäcker, C.; Mundt, S. Impact of Temperature on the Biosynthesis of Cytotoxically Active Carbamidocyclophanes A–E in Nostoc sp. CAVN10. J. Appl. Phycol. 2016, 28, 951–963. [Google Scholar] [CrossRef]
- Pérez, V.T.; Ticona, L.A.; Cabanillas, A.H.; Corral, S.M.; Perles, J.; Valencia, D.F.R.; Quintana, A.M.; Domenech, M.O.; Sánchez, Á.R. Antitumoral Potential of Carbamidocyclophanes and Carbamidocylindrofridin A Isolated from the Cyanobacterium Cylindrospermum stagnale BEA 0605B. Phytochemistry 2020, 180, 112529. [Google Scholar] [CrossRef]
- Preisitsch, M.; Heiden, S.E.; Beerbaum, M.; Niedermeyer, T.H.J.; Schneefeld, M.; Herrmann, J.; Kumpfmüller, J.; Thürmer, A.; Neidhardt, I.; Wiesner, C.; et al. Effects of Halide Ions on the Carbamidocyclophane Biosynthesis in Nostoc sp. CAVN2. Mar. Drugs 2016, 14, 21. [Google Scholar] [CrossRef] [Green Version]
- Moore, R.E.; Patterson, G.M.L.; Mynderse, J.S.; Barchi, J.; Norton, T.R.; Furusawa, E.; Furusawa, S. Toxins from Cyanophytes Belonging to the Scytonemataceae. Pure Appl. Chem. 1986, 58, 263–271. [Google Scholar] [CrossRef]
- Carmeli, S.; Moore, R.E.; Patterson, G.M.L. Tolytoxin and New Scytophycins from Three Species of Scytonema. J. Nat. Prod. 1990, 53, 1533–1542. [Google Scholar] [CrossRef]
- Moore, R.E.; Furusawa, E.; Norton, T.R.; Patterson, G.M.L.; Mynderse, J.S. Scytophycins. United States Patent US 4996229A, 31 August 1989. [Google Scholar]
- Jung, J.H.; Moore, R.E.; Patterson, G.M.L. Scytophycins from a Blue-Green Alga Belonging to the Nostocaceae. Phytochemistry 1991, 30, 3615–3616. [Google Scholar] [CrossRef]
- Tomsickova, J.; Ondrej, M.; Cerny, J.; Hrouzek, P.; Kopecky, J. Analysis and Detection of Scytophycin Variants by HPLC-ESI-MS. Chem. Nat. Compd. 2014, 49, 1170–1171. [Google Scholar] [CrossRef]
- Andrianasolo, E.H.; Gross, H.; Goeger, D.; Musafija-Girt, M.; McPhail, K.; Leal, R.M.; Mooberry, S.L.; Gerwick, W.H. Isolation of Swinholide A and Related Glycosylated Derivatives from Two Field Collections of Marine Cyanobacteria. Org. Lett. 2005, 7, 1375–1378. [Google Scholar] [CrossRef]
- Kobayashi, M.; Tanaka, J.i.; Katori, T.; Matsuura, M.; Kitagawa, I. Structure of Swinholide A, a Potent Cytotoxic Macrolide from the Okinawan Marine Sponge Theonella swinhoei. Tetrahedron Lett. 1989, 30, 2963–2966. [Google Scholar] [CrossRef]
- Tao, Y.; Li, P.; Zhang, D.; Glukhov, E.; Gerwick, L.; Zhang, C.; Murray, T.F.; Gerwick, W.H. Samholides, Swinholide-Related Metabolites from a Marine Cyanobacterium Cf. Phormidium sp. J. Org. Chem. 2018, 83, 3034–3046. [Google Scholar] [CrossRef] [Green Version]
- Elsadek, L.A.; Matthews, J.H.; Nishimura, S.; Nakatani, T.; Ito, A.; Gu, T.; Luo, D.; Salvador-Reyes, L.A.; Paul, V.J.; Kakeya, H.; et al. Genomic and Targeted Approaches Unveil the Cell Membrane as a Major Target of the Antifungal Cytotoxin Amantelide A. ChemBioChem 2021, 22, 1790–1799. [Google Scholar] [CrossRef] [PubMed]
- Crnkovic, C.M.; Braesel, J.; Krunic, A.; Eustáquio, A.S.; Orjala, J. Scytodecamide from the Cultured Scytonema Sp. UIC 10036 Expands the Chemical and Genetic Diversity of Cyanobactins. ChemBioChem 2020, 21, 845–852. [Google Scholar] [CrossRef]
- Cano, M.M.S.D.; De Mul, M.C.Z.; De Caire, G.Z.; De Halperin, D.R. Inhibition of Candida albicans and Staphylococcus aureus by Phenolic Compounds from the Terrestrial Cyanobacterium Nostoc muscorum. J. Appl. Phycol. 1990, 2, 79–81. [Google Scholar] [CrossRef]
- Abdel-Raouf, N.; Ibraheem, I.B.M.; Abdel-Tawab, S.; Naser, Y.A.G. Antimicrobial and Antihyperlipidemic Activities of Isolated Quercetin from Anabaena aequalis. J. Phycol. 2011, 47, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Martelli, F.; Cirlini, M.; Lazzi, C.; Neviani, E.; Bernini, V. Edible Seaweeds and Spirulina Extracts for Food Application: In Vitro and in Situ Evaluation of Antimicrobial Activity towards Foodborne Pathogenic Bacteria. Foods 2020, 9, 1442. [Google Scholar] [CrossRef]
- Boudet, A.M. Evolution and Current Status of Research in Phenolic Compounds. Phytochemistry 2007, 68, 2722–2735. [Google Scholar] [CrossRef]
- El-Sheekh, M.M.; Osman, M.E.H.; Dyab, M.A.; Amer, M.S. Production and Characterization of Antimicrobial Active Substance from the Cyanobacterium Nostoc muscorum. Environ. Toxicol. Pharmacol. 2006, 21, 42–50. [Google Scholar] [CrossRef]
- Falch, B.S.; König, G.M.; Wright, A.D.; Sticher, O.; Röegger, H.; Bernardinelli, G. Ambigol A and B: New Biologically Active Polychlorinated Aromatic Compounds from the Terrestrial Blue-Green Alga Fischerella ambigua. J. Org. Chem. 1993, 58, 6570–6575. [Google Scholar] [CrossRef]
- Milzarek, T.M.; Gulder, T.A.M. Total Synthesis of the Ambigols: A Cyanobacterial Class of Polyhalogenated Natural Products. Org. Lett. 2021, 23, 102–106. [Google Scholar] [CrossRef]
- Wright, A.D.; Papendorf, O.; König, G.M.; Oberemm, A. Effects of Cyanobacterium Fischerella ambigua Isolates and Cell Free Culture Media on Zebrafish (Danio Rerio) Embryo Development. Chemosphere 2006, 65, 604–608. [Google Scholar] [CrossRef]
- Duell, E.R.; Milzarek, T.M.; El Omari, M.; Linares-Otoya, L.J.; Schäberle, T.F.; König, G.M.; Gulder, T.A.M. Identification, Cloning, Expression and Functional Interrogation of the Biosynthetic Pathway of the Polychlorinated Triphenyls Ambigol A-C from Fischerella ambigua 108b. Org. Chem. Front. 2020, 7, 3193–3201. [Google Scholar] [CrossRef]
- Kresna, I.D.M.; Linares-Otoya, L.; Milzarek, T.; Duell, E.R.; Mir Mohseni, M.; Mettal, U.; König, G.M.; Gulder, T.A.M.; Schäberle, T.F. In Vitrocharacterization of 3-Chloro-4-Hydroxybenzoic Acid Building Block Formation in Ambigol Biosynthesis. Org. Biomol. Chem. 2021, 19, 2302–2311. [Google Scholar] [CrossRef] [PubMed]
- Morales-Jiménez, M.; Gouveia, L.; Yañez-Fernandez, J.; Castro-Muñoz, J.; Barragan-Huerta, B.E. Microalgae-Based Biopolymer as a Potential Bioactive Film. Coatings 2020, 10, 120. [Google Scholar] [CrossRef] [Green Version]
- Pattanaik, B.; Lindberg, P. Terpenoids and Their Biosynthesis in Cyanobacteria. Life 2015, 5, 269–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabanillas, A.H.; Tena Pérez, V.; Maderuelo Corral, S.; Rosero Valencia, D.F.; Martel Quintana, A.; Ortega Doménech, M.; Rumbero Sánchez, Á. Cybastacines A and B: Antibiotic Sesterterpenes from a Nostoc sp. Cyanobacterium. J. Nat. Prod. 2018, 81, 410–413. [Google Scholar] [CrossRef]
- Mouritsen, O.G.; Zuckermann, M.J. What’s so Special about Cholesterol? Lipids 2004, 39, 1101–1113. [Google Scholar] [CrossRef] [PubMed]
- Dupont, S.; Lemetais, G.; Ferreira, T.; Cayot, P.; Gervais, P.; Beney, L. Ergosterol Biosynthesis: A Fungal Pathway for Life on Land? Evolution 2012, 66, 2961–2968. [Google Scholar] [CrossRef]
- Li, Y.; Li, Y.; Mengist, H.M.; Shi, C.; Zhang, C.; Wang, B.; Li, T.; Huang, Y.; Xu, Y.; Jin, T. Structural Basis of the Pore-Forming Toxin/Membrane Interaction. Toxins 2021, 13, 128. [Google Scholar] [CrossRef]
- Oftedal, L.; Myhren, L.; Jokela, J.; Gausdal, G.; Sivonen, K.; Doskeland, S.O.; Herfindal, L. The Lipopeptide Toxins Anabaenolysin A and B Target Biological Membranes in a Cholesterol-Dependent Manner. Biochim. Biophys. Acta Biomembr. 2012, 1818, 3000–3009. [Google Scholar] [CrossRef] [Green Version]
- Humisto, A.; Jokela, J.; Teigen, K.; Wahlsten, M.; Permi, P.; Sivonen, K.; Herfindal, L. Characterization of the Interaction of the Antifungal and Cytotoxic Cyclic Glycolipopeptide Hassallidin with Sterol-Containing Lipid Membranes. Biochim. Biophys. Acta Biomembr. 2019, 1861, 1510–1521. [Google Scholar] [CrossRef]
- Hrouzek, P.; Kuzma, M.; Černyý, J.; Novák, P.; Fisšer, R.; Sšimek, P.; Lukesšová, A.; Kopeckyý, J. The Cyanobacterial Cyclic Lipopeptides Puwainaphycins F/G Are Inducing Necrosis via Cell Membrane Permeabilization and Subsequent Unusual Actin Relocalization. Chem. Res. Toxicol. 2012, 25, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Vašíček, O.; Hájek, J.; Bláhová, L.; Hrouzek, P.; Babica, P.; Kubala, L.; Šindlerová, L. Cyanobacterial Lipopeptides Puwainaphycins and Minutissamides Induce Disruptive and Pro-Inflammatory Processes in Caco-2 Human Intestinal Barrier Model. Harmful Algae 2020, 96, 101849. [Google Scholar] [CrossRef] [PubMed]
- Patterson, G.M.L.; Smith, C.D.; Kimura, L.H.; Britton, B.A.; Carmeli, S. Action of Tolytoxin on Cell Morphology, Cytoskeletal Organization, and Actin Polymerization. Cell. Motil. Cytoskelet. 1993, 24, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Koiso, Y.; Morita, K.; Kobayashi, M.; Wang, W.; Ohyabu, N.; Iwasaki, S. Effects of Arenastatin A and Its Synthetic Analogs on Microtubule Assembly. Chem. Biol. Interact. 1996, 102, 183–191. [Google Scholar] [CrossRef]
- Parker, A.L.; Kavallaris, M.; McCarroll, J.A. Microtubules and Their Role in Cellular Stress in Cancer. Front. Oncol. 2014, 4, 153. [Google Scholar] [CrossRef] [Green Version]
- Brouhard, G.J.; Rice, L.M. Microtubule Dynamics: An Interplay of Biochemistry and Mechanics. Nat. Rev. Mol. Cell. Biol. 2018, 19, 451–463. [Google Scholar] [CrossRef]
- De Forges, H.; Bouissou, A.; Perez, F. Interplay between Microtubule Dynamics and Intracellular Organization. Int. J. Biochem. Cell. Biol. 2012, 44, 266–274. [Google Scholar] [CrossRef]
- Kaul, R.; Risinger, A.L.; Mooberry, S.L. Microtubule-Targeting Drugs: More than Antimitotics. J. Nat. Prod. 2019, 82, 680–685. [Google Scholar] [CrossRef]
- Steinmetz, M.O.; Prota, A.E. Microtubule-Targeting Agents: Strategies To Hijack the Cytoskeleton. Trends Cell. Biol. 2018, 28, 776–792. [Google Scholar] [CrossRef]
- Wordeman, L.; Vicente, J.J. Microtubule Targeting Agents in Disease: Classic Drugs, Novel Roles. Cancers 2021, 13, 5650. [Google Scholar] [CrossRef]
- Smith, C.D.; Zhang, X. Mechanism of Action of Cryptophycin: Interaction with the Vinca Alkaloid Domain of Tubulin. J. Biol. Chem. 1996, 271, 6192–6198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mooberry, S.L.; Taoka, C.R.; Busquets, L. Cryptophycin 1 Binds to Tubulin at a Site Distinct from the Colchicine Binding Site and at a Site That May Overlap the Vinca Binding Site. Cancer Lett. 1996, 107, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Liebman, C.; McColloch, A.; Rabiei, M.; Bowling, A.; Cho, M. Mechanics of the Cell: Interaction Mechanisms and Mechanobiological Models. In Current Topics in Membranes; Elsevier Inc.: Amsterdam, The Netherlands, 2020; Volume 86, pp. 143–184. ISBN 9780128210215. [Google Scholar]
- Klenchin, V.A.; Allingham, J.S.; King, R.; Tanaka, J.; Marriott, G.; Rayment, I. Trisoxazole Macrolide Toxins Mimic the Binding of Actin-Capping Proteins to Actin. Nat. Struct. Biol. 2003, 10, 1058–1063. [Google Scholar] [CrossRef]
- Klenchin, V.A.; King, R.; Tanaka, J.; Marriott, G.; Rayment, I. Structural Basis of Swinholide A Binding to Actin. Chem. Biol. 2005, 12, 287–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gbankoto, A.; Vigo, J.; Dramane, K.; Banaigs, B.; Aina, E.; Salmon, J. Cytotoxic Effect of Laxaphycins A and B on Human Lymphoblastic Cells (CCRF-CEM) Using Digitised Videomicrofluorometry. Vivo 2005, 582, 577–582. [Google Scholar]
- Schaller, M.; Borelli, C.; Korting, H.C.; Hube, B. Hydrolytic Enzymes as Virulence Factors of Candida albicans. Mycoses 2005, 48, 365–377. [Google Scholar] [CrossRef]
- Manivannan, P.; Muralitharan, G. Molecular Modeling of Abc Transporter System—Permease Proteins from Microcoleus chthonoplastes PCC 7420 for Effective Binding against Secreted Aspartyl Proteinases in Candida albicans—A Therapeutic Intervention. Interdiscip. Sci. 2014, 6, 63–70. [Google Scholar] [CrossRef]
- Madhumathi, V.; Vijayakumar, S. Identification of Novel Cyanobacterial Compounds for Oral Disease through in Vitro and Insilico Approach. Biomed. Aging Pathol. 2014, 4, 223–228. [Google Scholar] [CrossRef]
- Pagnussatt, F.A.; Kupski, L.; Darley, F.T.; Filoda, P.F.; Ponte, É.M.D.; Garda-Buffon, J.; Badiale-Furlong, E. Fusarium graminearum Growth Inhibition Mechanism Using Phenolic Compounds from Spirulina sp. Food Sci. Technol. 2013, 33, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Niedermeyer, T.H.J.; Chilczuk, T.; Monson, R.; Schmieder, P.; Christov, V.; Enke, H.; Salmond, G. Ambigols from the Cyanobacterium Fischerella ambigua Increase Prodigiosin Production in Serratia sp. ACS Chem. Biol. 2020, 15, 2929–2936. [Google Scholar] [CrossRef]
- Hazarika, D.J.; Gautom, T.; Parveen, A.; Goswami, G.; Barooah, M.; Modi, M.K.; Boro, R.C. Mechanism of Interaction of an Endofungal Bacterium Serratia marcescens D1 with Its Host and Non-Host Fungi. PLoS ONE 2020, 15, e0224051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habash, S.S.; Brass, H.U.C.; Klein, A.S.; Klebl, D.P.; Weber, T.M.; Classen, T.; Pietruszka, J.; Grundler, F.M.W.; Schleker, A.S.S. Novel Prodiginine Derivatives Demonstrate Bioactivities on Plants, Nematodes, and Fungi. Front. Plant. Sci. 2020, 11, 579807. [Google Scholar] [CrossRef] [PubMed]
- Wiltbank, L.B.; Kehoe, D.M. Diverse Light Responses of Cyanobacteria Mediated by Phytochrome Superfamily Photoreceptors. Nat. Rev. Microbiol. 2019, 17, 37–50. [Google Scholar] [CrossRef]
- Muhetaer, G.; Asaeda, T.; Jayasanka, S.M.D.H.; Baniya, M.B.; Abeynayaka, H.D.L.; Rashid, M.H.; Yan, H.Y. Effects of Light Intensity and Exposure Period on the Growth and Stress Responses of Two Cyanobacteria Species: Pseudanabaena galeata and Microcystis aeruginosa. Water 2020, 12, 407. [Google Scholar] [CrossRef] [Green Version]
- Vonshak, A.; Chanawongse, L.; Bunnag, B.; Tanticharoen, M. Light Acclimation and Photoinhibition in Three Spirulina platensis (Cyanobacteria) Isolates. J. Appl. Phycol. 1996, 8, 35–40. [Google Scholar] [CrossRef]
- Zheng, T.; Zhou, M.; Yang, L.; Wang, Y.; Wang, Y.; Meng, Y.; Liu, J.; Zuo, Z. Effects of High Light and Temperature on Microcystis aeruginosa Cell Growth and β-Cyclocitral Emission. Ecotoxicol. Environ. Saf. 2020, 192, 110313. [Google Scholar] [CrossRef]
- Klepacz-Smółka, A.; Pietrzyk, D.; Szeląg, R.; Głuszcz, P.; Daroch, M.; Tang, J.; Ledakowicz, S. Effect of Light Colour and Photoperiod on Biomass Growth and Phycocyanin Production by Synechococcus PCC 6715. Bioresour. Technol. 2020, 313, 123700. [Google Scholar] [CrossRef]
- Patterson, G.M.L.; Bolis, C.M. Regulation of Scytophycin Accumulation in Cultures of Scytonema ocellatum. I. Physical Factors. Appl. Microbiol. Biotechnol. 1993, 40, 375–381. [Google Scholar] [CrossRef]
- Polyzois, A.; Kirilovsky, D.; Dufat, T.H.; Michel, S. Effects of Modification of Light Parameters on the Production of Cryptophycin, Cyanotoxin with Potent Anticancer Activity, in Nostoc sp. Toxins 2020, 12, 809. [Google Scholar] [CrossRef]
- Gupta, V.; Prasanna, R.; Singh, S.; Dureja, P.; Nageena, R.; Sharma, J. Enhancing the Production of an Antifungal Compound from Anabaena laxa through Modulation of Environmental Conditions and Its Characterization. Process Biochem. 2013, 48, 768–774. [Google Scholar] [CrossRef]
- Khajepour, F.; Hosseini, S.A.; Ghorbani Nasrabadi, R.; Markou, G. Effect of Light Intensity and Photoperiod on Growth and Biochemical Composition of a Local Isolate of Nostoc calcicola. Appl. Biochem. Biotechnol. 2015, 176, 2279–2289. [Google Scholar] [CrossRef] [PubMed]
- Ibraheem, I.; Abdel-Raouf, N.; Hammouda, O.; Abdel-Wahab, N. The Potential for Using Culture Filtrate of Chroococcus minutus As Fungicidal Agent Against Phytopathogenic Pythium sp. Egypt. J. Phycol. 2008, 9, 99–114. [Google Scholar] [CrossRef]
- Cramer, W.A.; Singh, S.K. A Structure Perspective on Organelle Bioenergetics. In Encyclopedia of Cell Biology; Elsevier: Amsterdam, The Netherlands, 2016; Volume 3, pp. 298–308. ISBN 9780123944474. [Google Scholar]
- Luimstra, V.M.; Schuurmans, J.M.; Verschoor, A.M.; Hellingwerf, K.J.; Huisman, J.; Matthijs, H.C.P. Blue Light Reduces Photosynthetic Efficiency of Cyanobacteria through an Imbalance between Photosystems I and II. Photosynth. Res. 2018, 138, 177–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karan, T.; Kayir, O.; Altuner, Z.; Erenler, R. Growth and Norharmane Production of Chroococcus minutus under Various Stress Conditions. Int. J. Chem. Technol. 2018, 2, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Bloor, S.; England, R.R. Antibiotic Production by the Cyanobacterium Nostoc muscorum. J. Appl. Phycol. 1989, 1, 367–372. [Google Scholar] [CrossRef]
- Chetsumon, A.; Fujieda, K.; Hirata, K.; Yagi, K.; Miura, Y. Optimization of Antibiotic Production by the Cyanobacterium Scytonema sp. TISTR 8208 Immobilized on Polyurethane Foam. J. Appl. Phycol. 1993, 5, 615–622. [Google Scholar] [CrossRef]
- Codd, G.A.; Fish, S.A. Analysis of Culture Conditions Controlling the Yield of Bioactive Material Produced by the Thermotolerant Cyanobacterium (Blue-Green Alga) Phormidium. Eur. J. Phycol. 1994, 29, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Abdel Hameed, M.S.; Hassan, S.H.; Mohammed, R.; Gamal, R. Isolation and Characterization of Antimicrobial Active Compounds from the Cyanobacterium Nostoc commune Vauch. J. Pure Appl. Microbiol. 2013, 7, 109–116. [Google Scholar] [CrossRef]
- Gradíssimo, D.G.; Oliveira da Silva, V.C.; Xavier, L.P.; do Nascimento, S.V.; Valadares, R.B.D.S.; Faustino, S.M.M.; Schneider, M.P.C.; Santos, A.V. Glucosidase Inhibitors Screening in Microalgae and Cyanobacteria Isolated from the Amazon and Proteomic Analysis of Inhibitor Producing Synechococcus sp. GFB01. Microorganisms 2021, 9, 1593. [Google Scholar] [CrossRef]
- Patterson, G.M.L.; Bolis, C.M. Regulation of Scytophycin Accumulation in Cultures of Scytonema ocellatum. II. Nutrient Requirements. Appl. Microbiol. Biotechnol. 1995, 43, 692–700. [Google Scholar] [CrossRef]
- Mahakhant, A.; Padungwong, P.; Arunpairojana, V.; Atthasampunna, P. Control of the Plant Pathogenic Fungus Macrophomina phaseolina in Mung Bean by a Microalgal Extract. Phycol. Res. 1998, 46, 3–7. [Google Scholar] [CrossRef]
- Årstøl, E.; Hohmann-Marriott, M.F. Cyanobacterial Siderophores—Physiology, Structure, Biosynthesis, and Applications. Mar. Drugs 2019, 17, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patterson, G.M.L.; Bolis, C.M. Fungal Cell-Wall Polysaccharides Elicit an Antifungal Secondary Metabolite (Phytoalexin) in the Cyanobacterium Scytonema ocellatum. J. Phycol. 1997, 33, 54–60. [Google Scholar] [CrossRef]
- Nowruzi, B. A Gene Expression Study on Strains of Nostoc (Cyanobacteria) Revealing Antimicrobial Activity under Mixotrophic Conditions. Afr. J. Biotechnol. 2012, 11, 11296–11308. [Google Scholar] [CrossRef]
- Chaudhary, V.; Prasanna, R.; Bhatnagar, A.K. Influence of Phosphorus and PH on the Fungicidal Potential of Anabaena Strains. J. Basic. Microbiol. 2013, 53, 201–213. [Google Scholar] [CrossRef]
- Battah, M.G.; Ibrahim, H.A.H.; El-Naggar, M.M.; Abdel_Gawad, F.K.; Amer, M.S. Antifungal Agent from Spirulina maxima: Extraction and Characterization. Glob. J. Pharmacol. 2014, 8, 228–236. [Google Scholar]
- Lürling, M.; Eshetu, F.; Faassen, E.J.; Kosten, S.; Huszar, V.L.M. Comparison of Cyanobacterial and Green Algal Growth Rates at Different Temperatures. Freshw. Biol. 2013, 58, 552–559. [Google Scholar] [CrossRef]
- Zhang, D.; Dechatiwongse, P.; del Rio-Chanona, E.A.; Maitland, G.C.; Hellgardt, K.; Vassiliadis, V.S. Modelling of Light and Temperature Influences on Cyanobacterial Growth and Biohydrogen Production. Algal Res. 2015, 9, 263–274. [Google Scholar] [CrossRef] [Green Version]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial Blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Mantzouki, E.; Lürling, M.; Fastner, J.; de Senerpont Domis, L.; Wilk-Woźniak, E.; Koreivienė, J.; Seelen, L.; Teurlincx, S.; Verstijnen, Y.; Krztoń, W.; et al. Temperature Effects Explain Continental Scale Distribution of Cyanobacterial Toxins. Toxins 2018, 10, 156. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, T.; Bryant, D.A. Growth at Low Temperature Causes Nitrogen Limitation in the Cyanobacterium Synechococcus sp. PCC 7002. Arch. Microbiol. 1997, 169, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Alghanmi, H.A.; Alkam, F.M.; Al-Taee, M.M. Effect of Light and Temperature on New Cyanobacteria Producers for Geosmin and 2-Methylisoborneol. J. Appl. Phycol. 2018, 30, 319–328. [Google Scholar] [CrossRef]
- Jacinavicius, F.R.; De Carvalho, L.R.; Carneiro, R.L.; Sant’Anna, C.L. The Influence of Temperature on Radiocystis Fernandoi Strain (Cyanobacteria) Growth and Microcystin Production. Rev. Bras. De Bot. 2018, 41, 675–680. [Google Scholar] [CrossRef]
- Gacheva, G.; Gigova, L.; Ivanova, N.; Iliev, I.; Toshkova, R.; Gardeva, E.; Kussovski, V.; Najdenski, H. Suboptimal Growth Temperatures Enhance the Biological Activity of Cultured Cyanobacterium Gloeocapsa Sp. J. Appl. Phycol. 2013, 25, 183–194. [Google Scholar] [CrossRef]
- Erenler, R.; Karan, T.; Altuner, Z. Growth and Metabolite Production of Chroococcus Minutus under Different Temperature and Light Conditions. J. New Results Sci. 2017, 6, 47–52. [Google Scholar]





























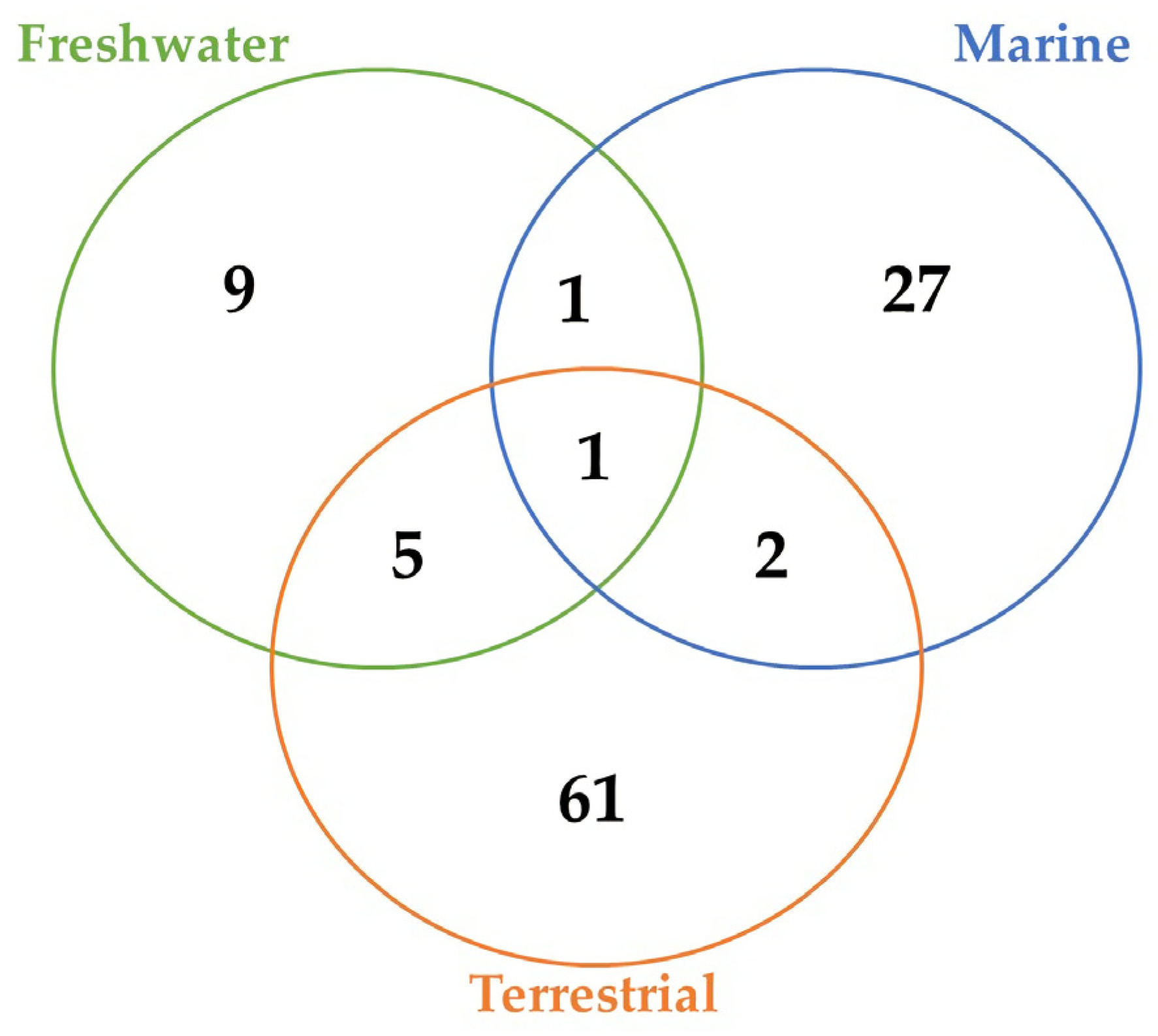{kind=link}
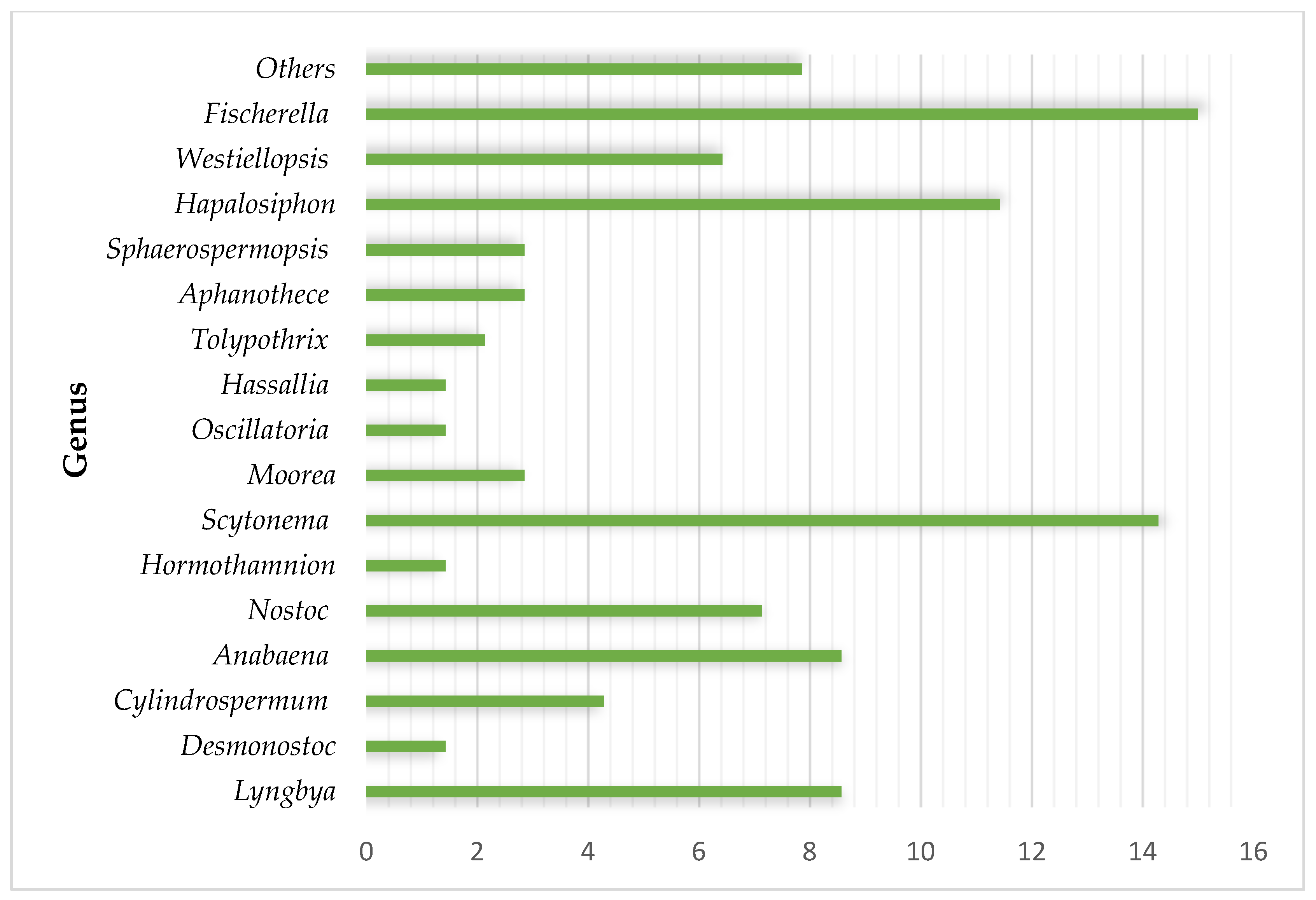{kind=link}
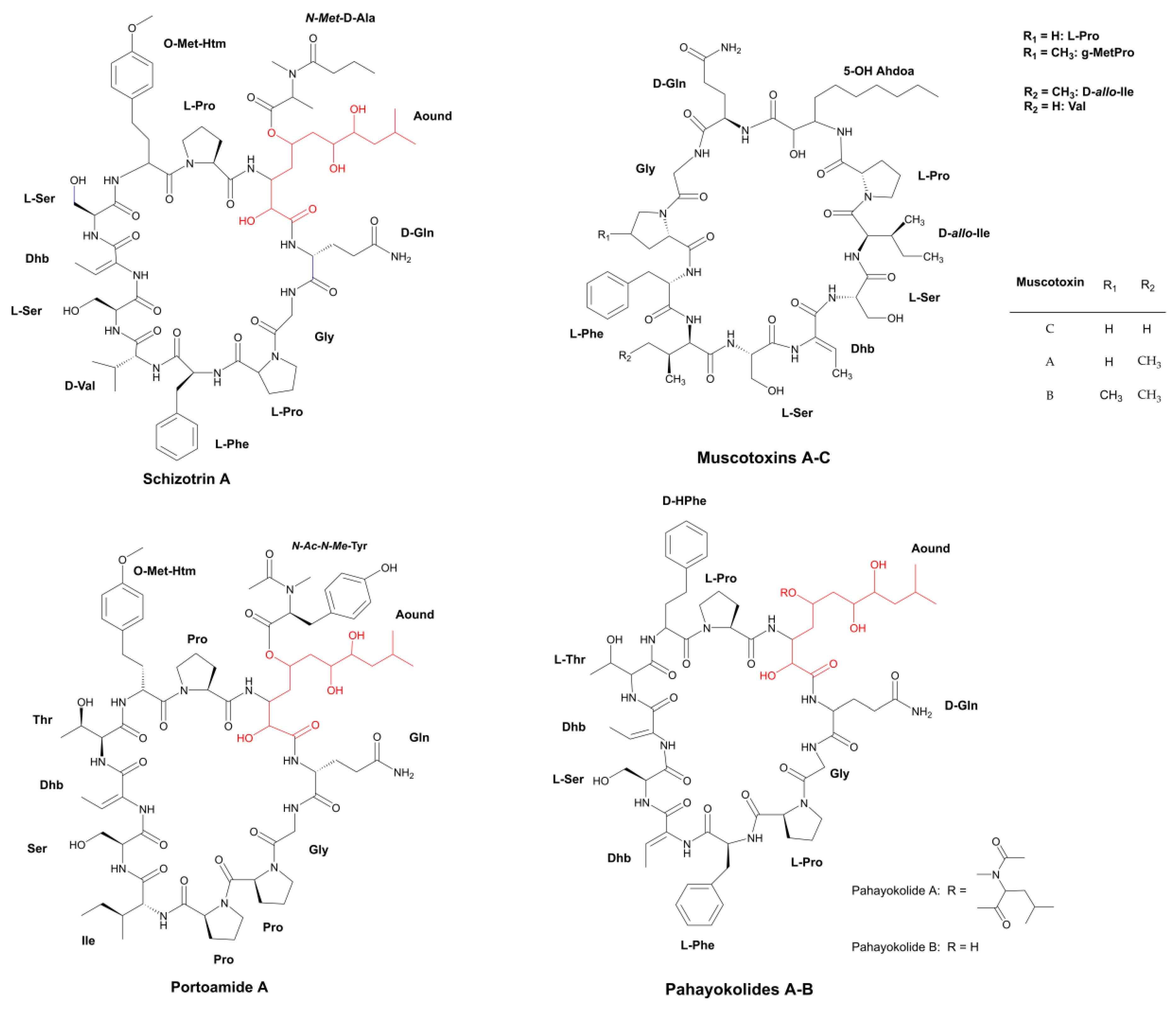{kind=link}
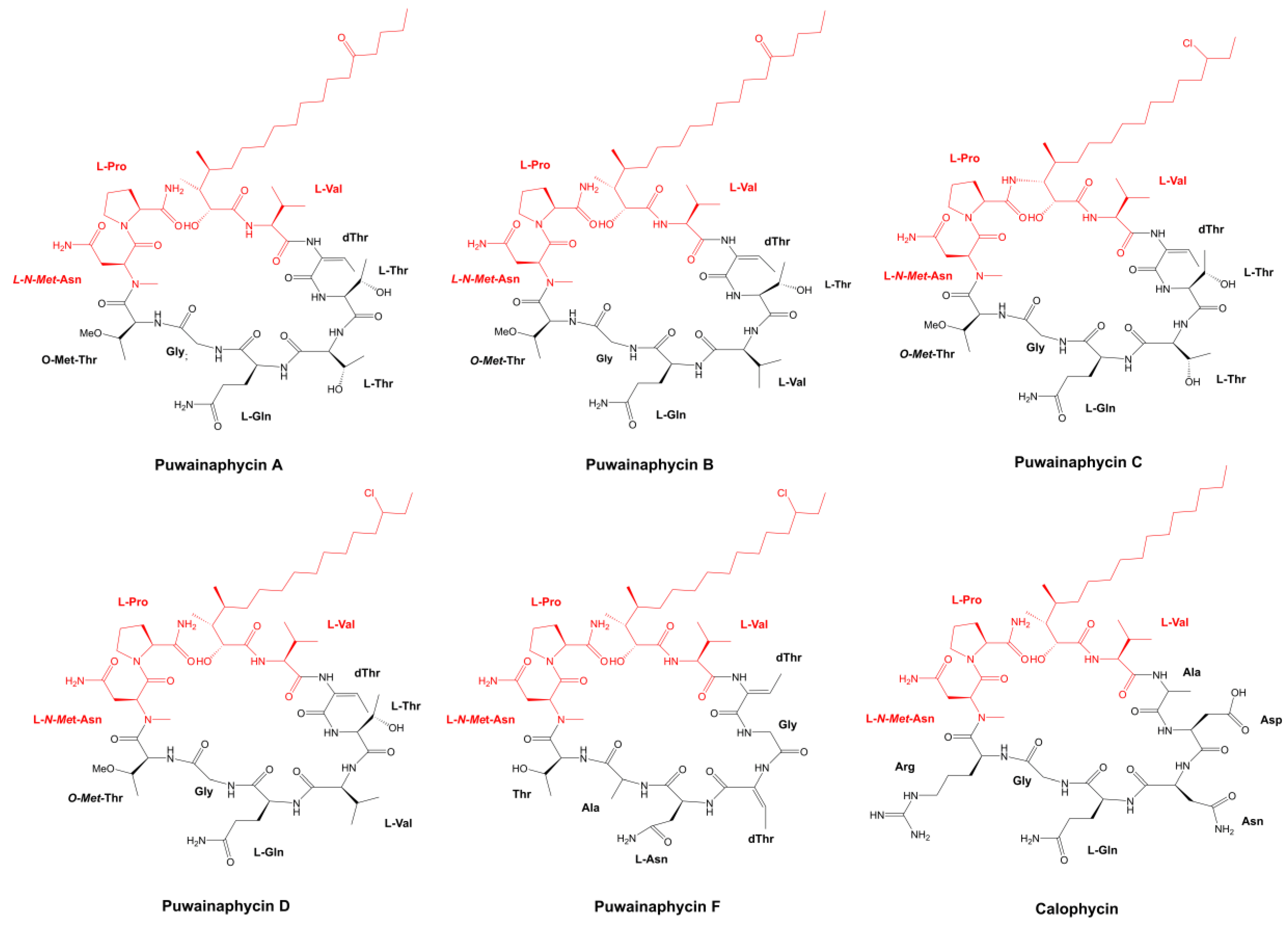{kind=link}
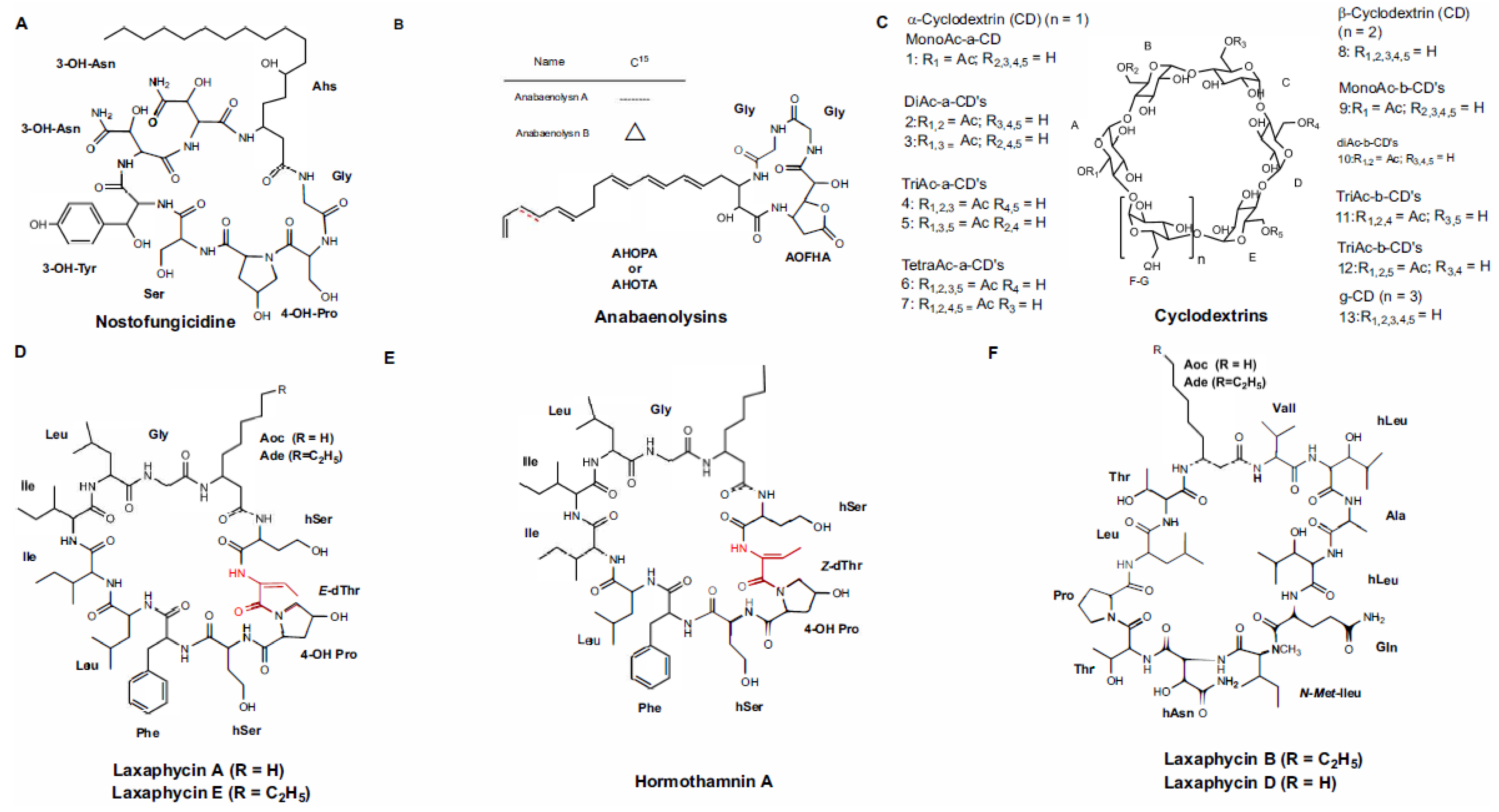{kind=link}
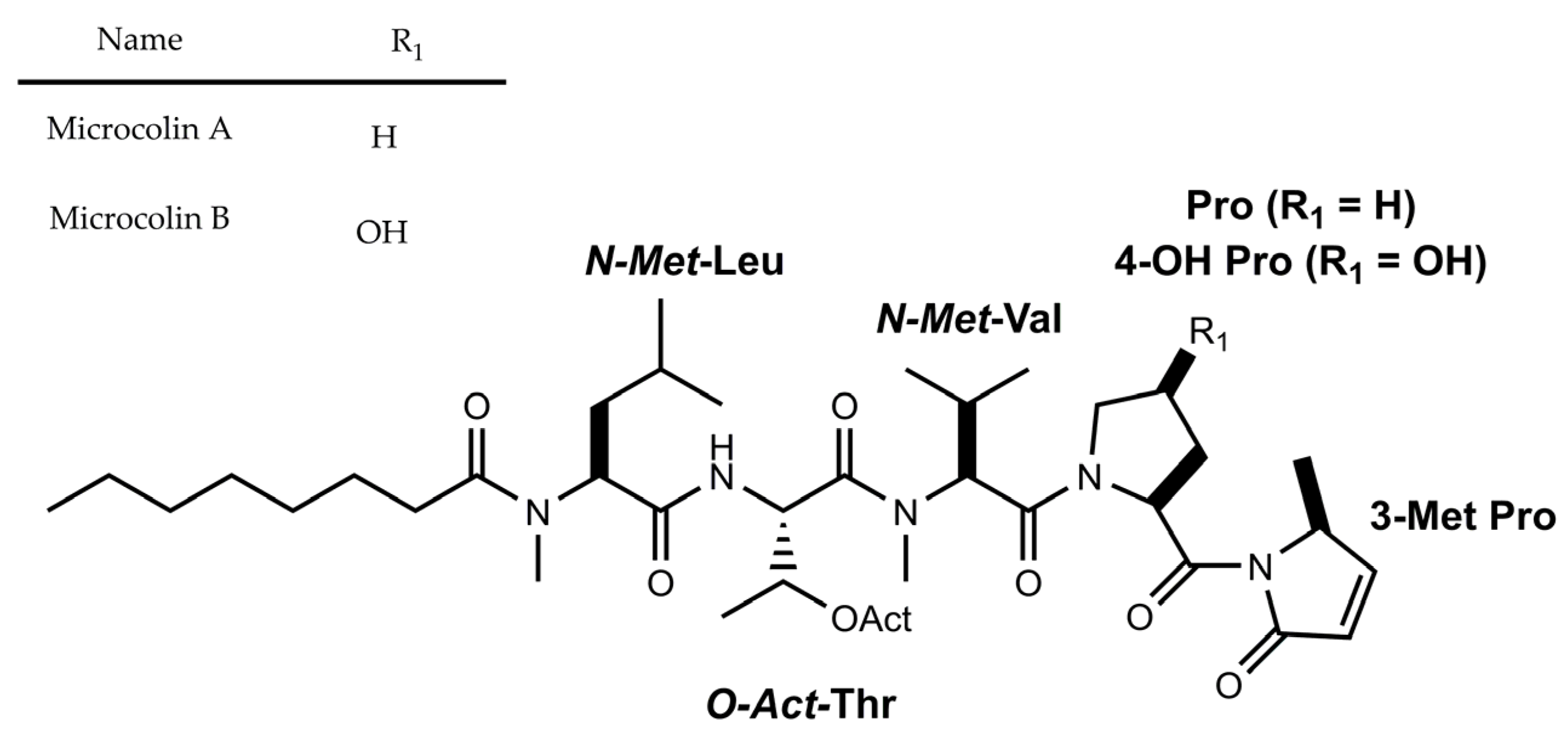{kind=link}
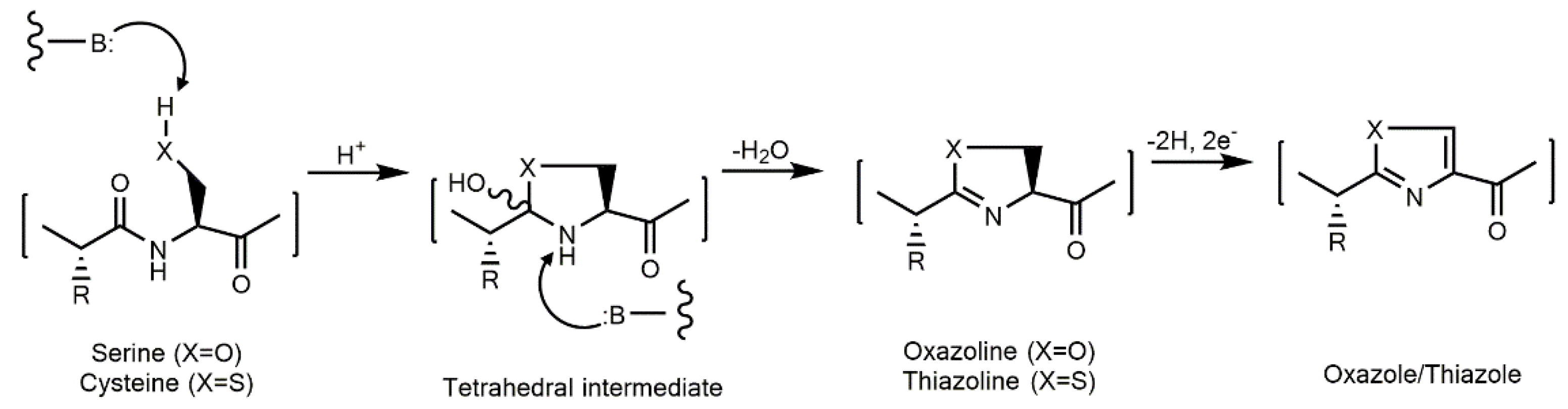{kind=link}
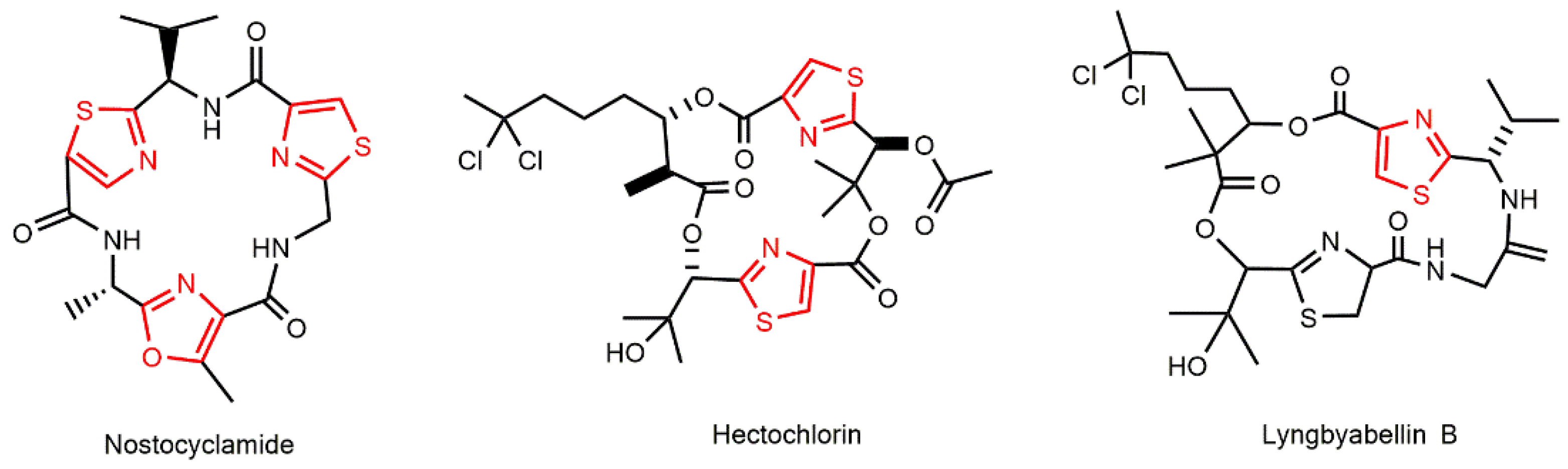{kind=link}
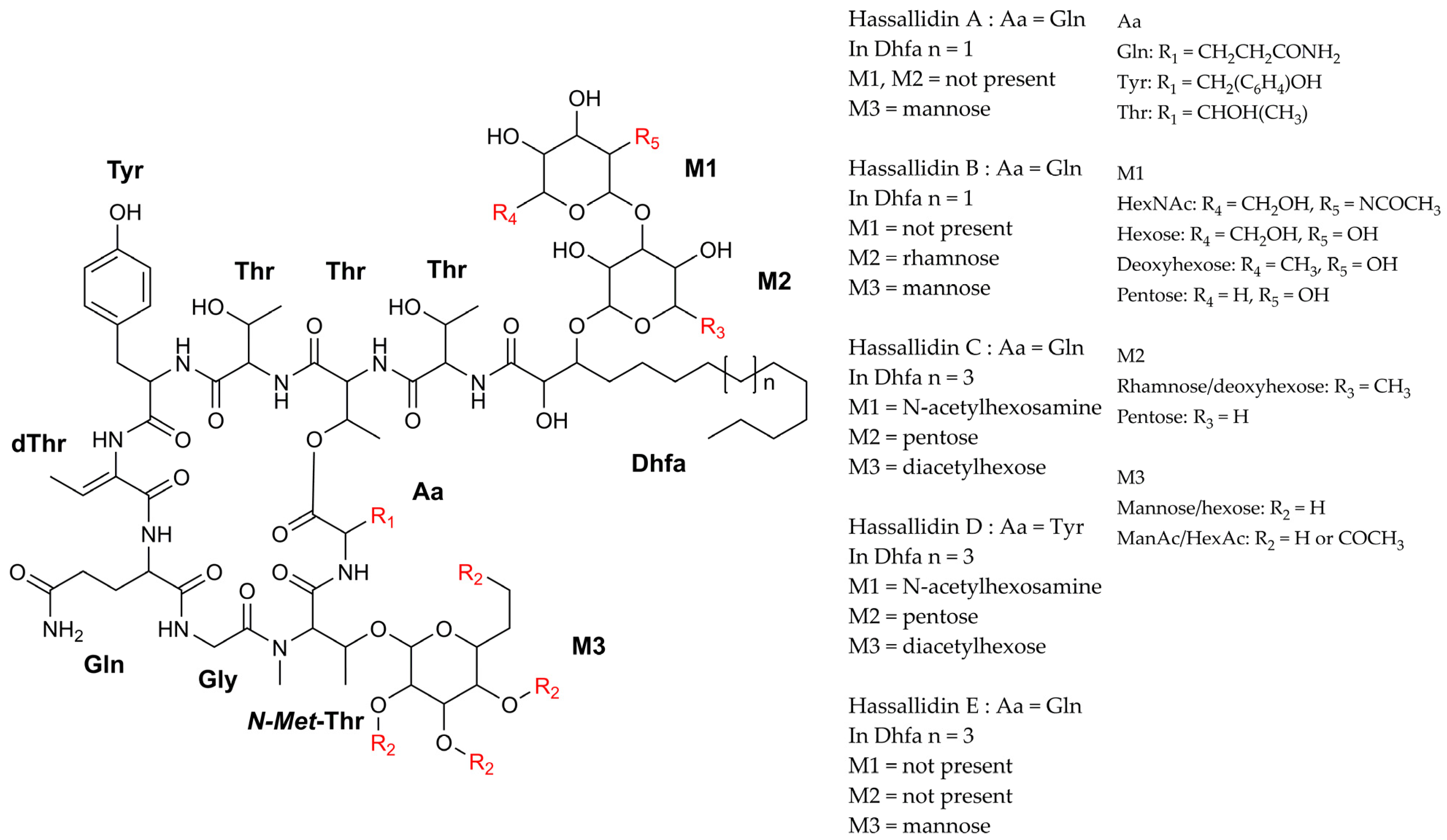{kind=link}
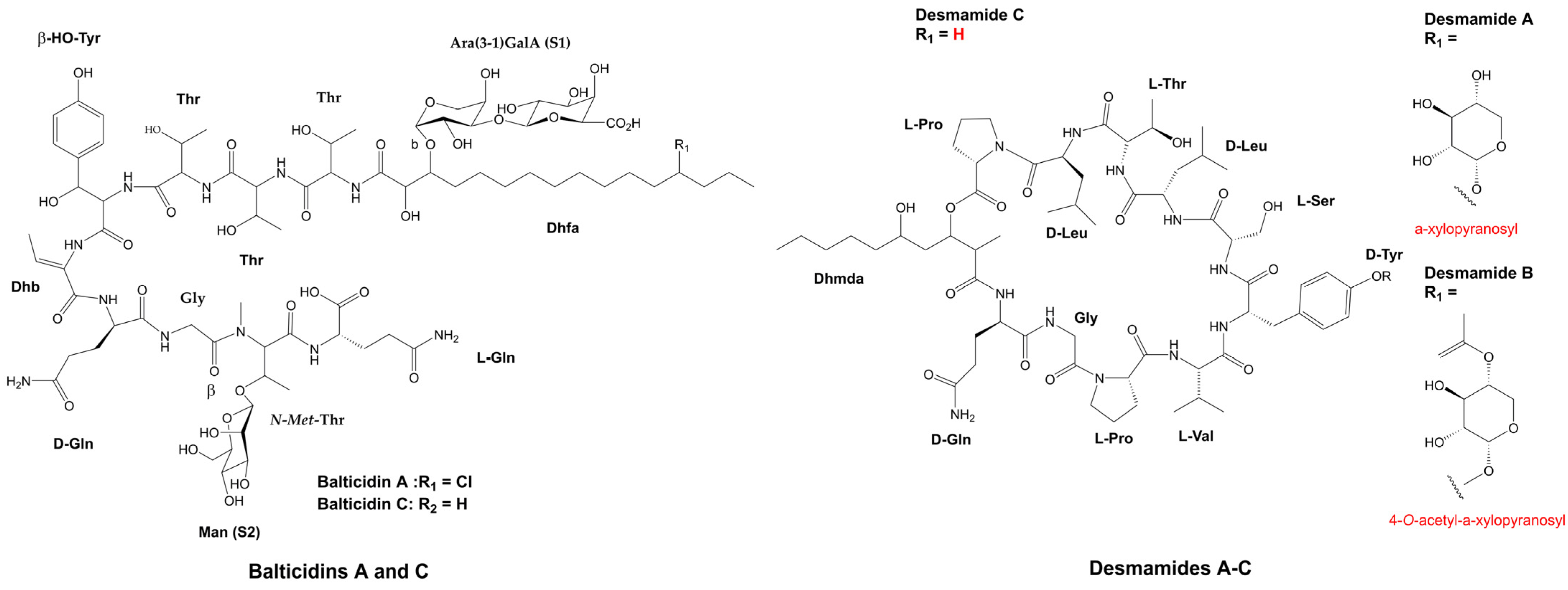{kind=link}
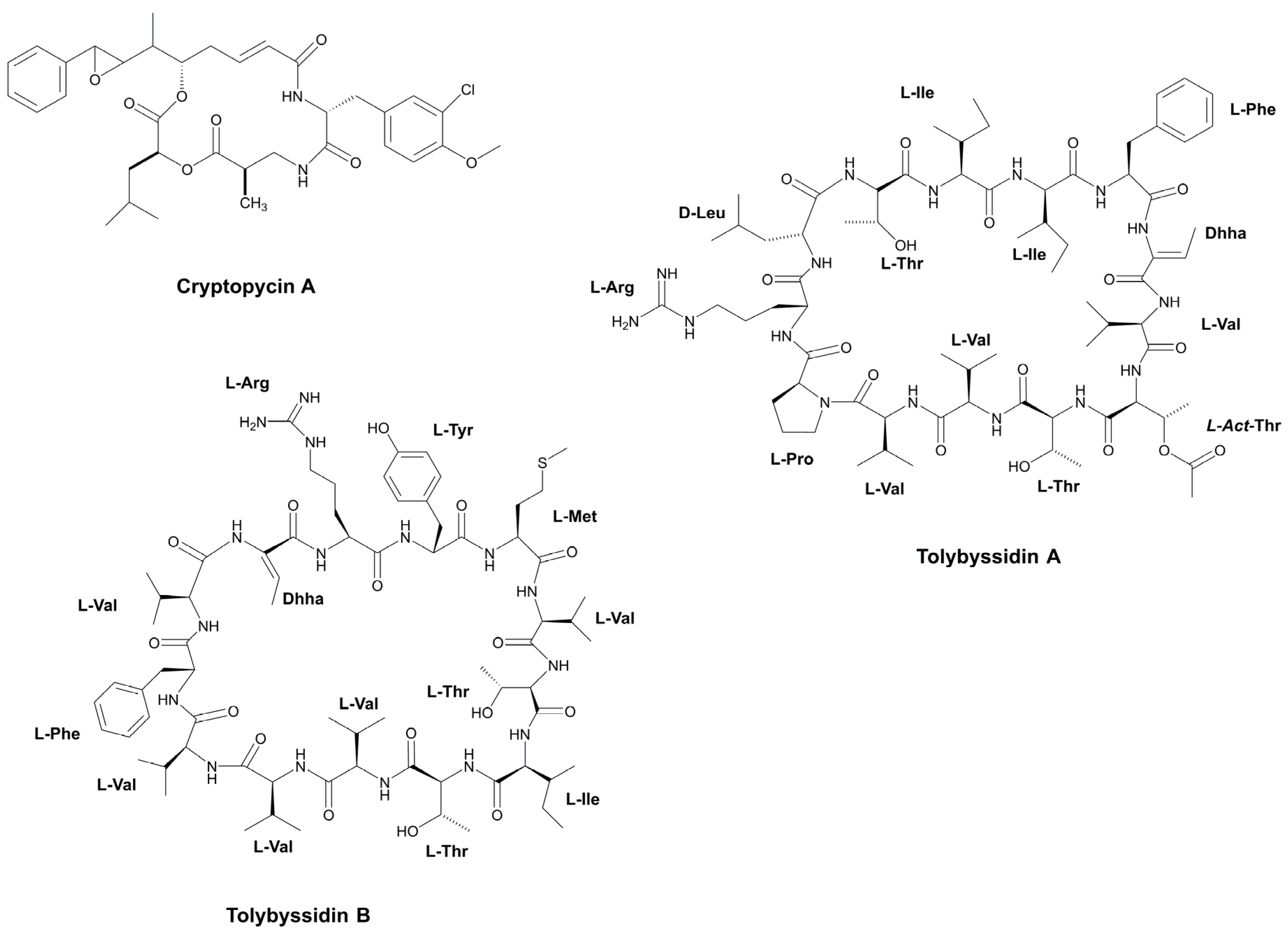{kind=link}
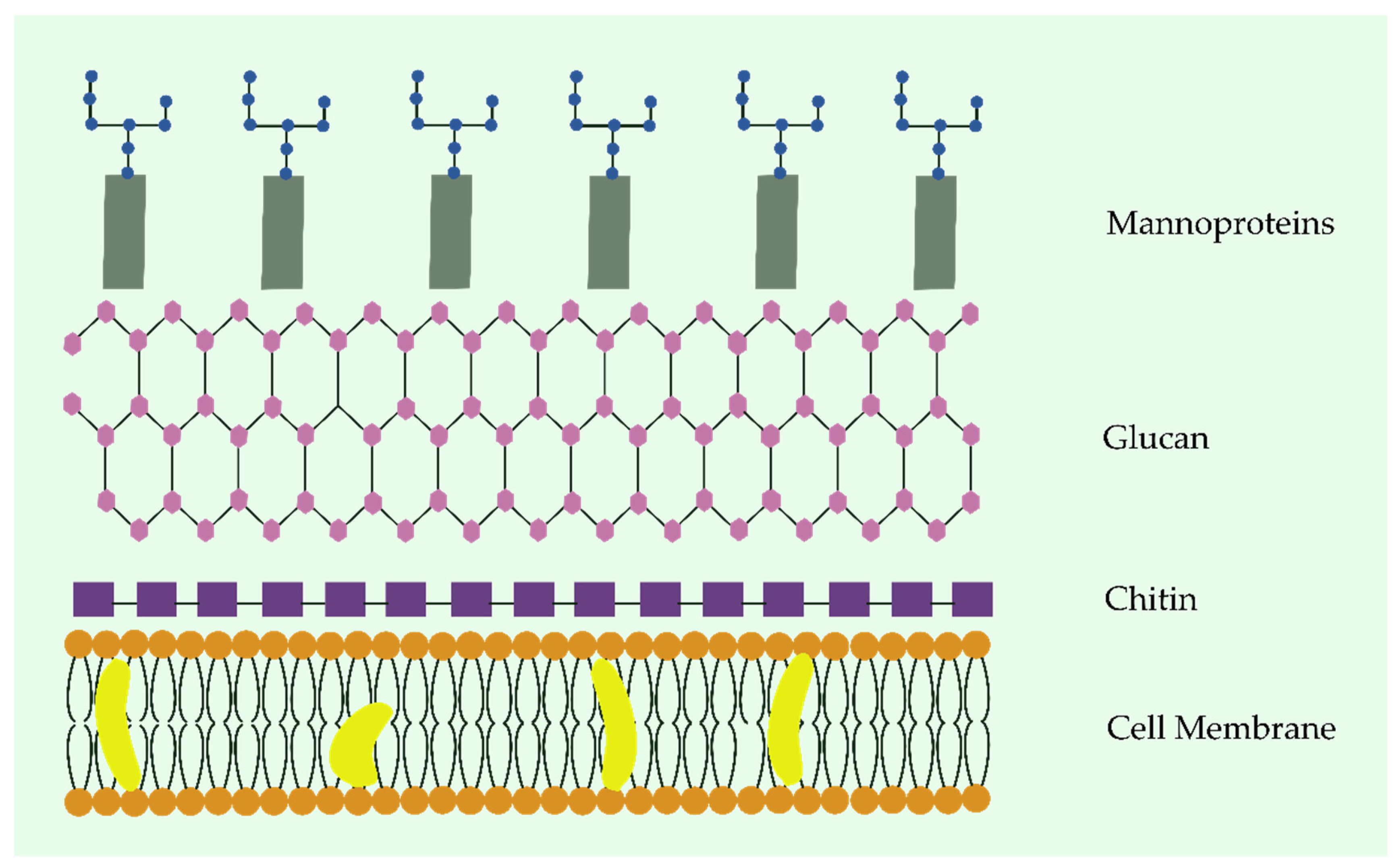{kind=link}
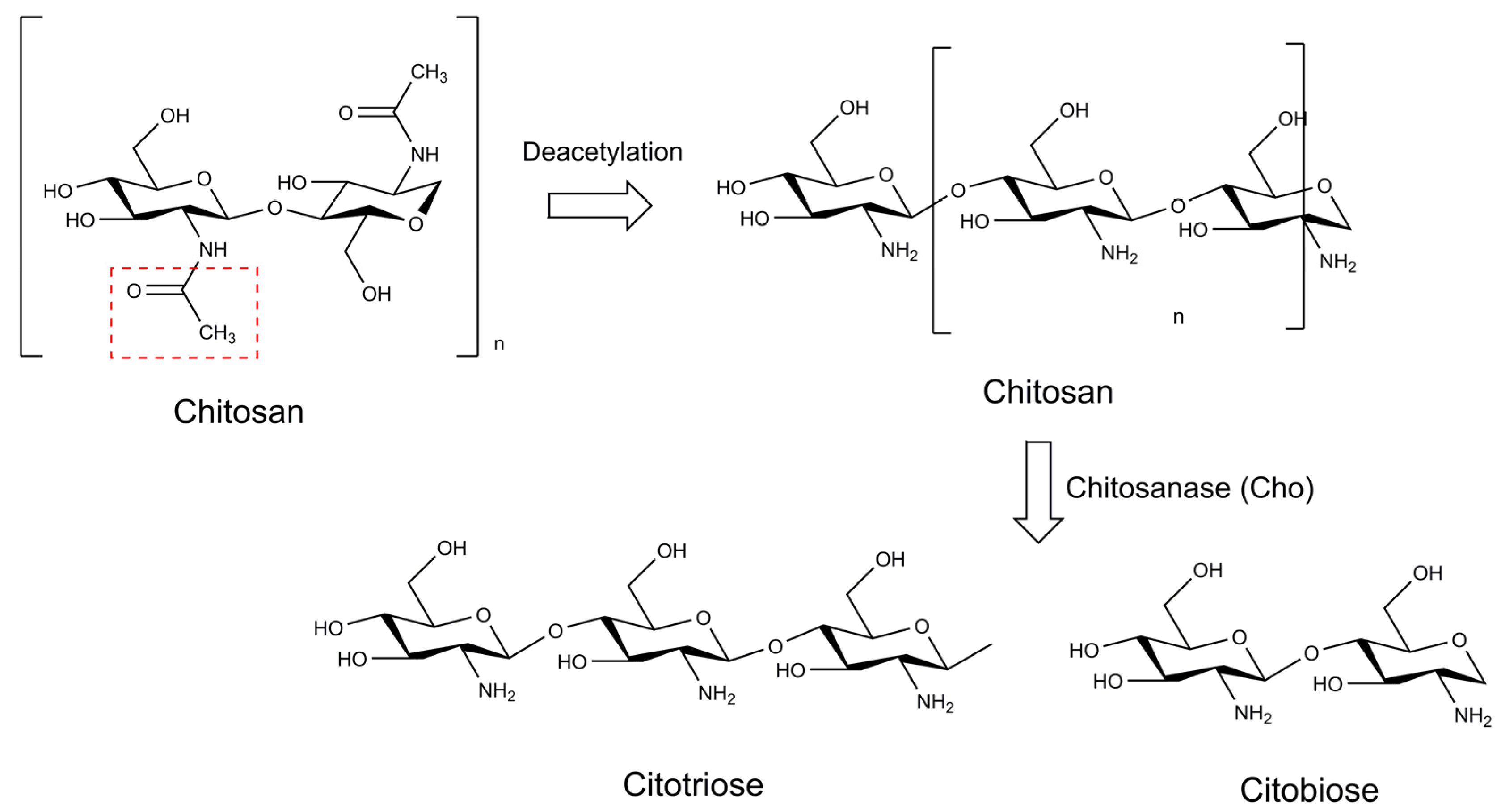{kind=link}
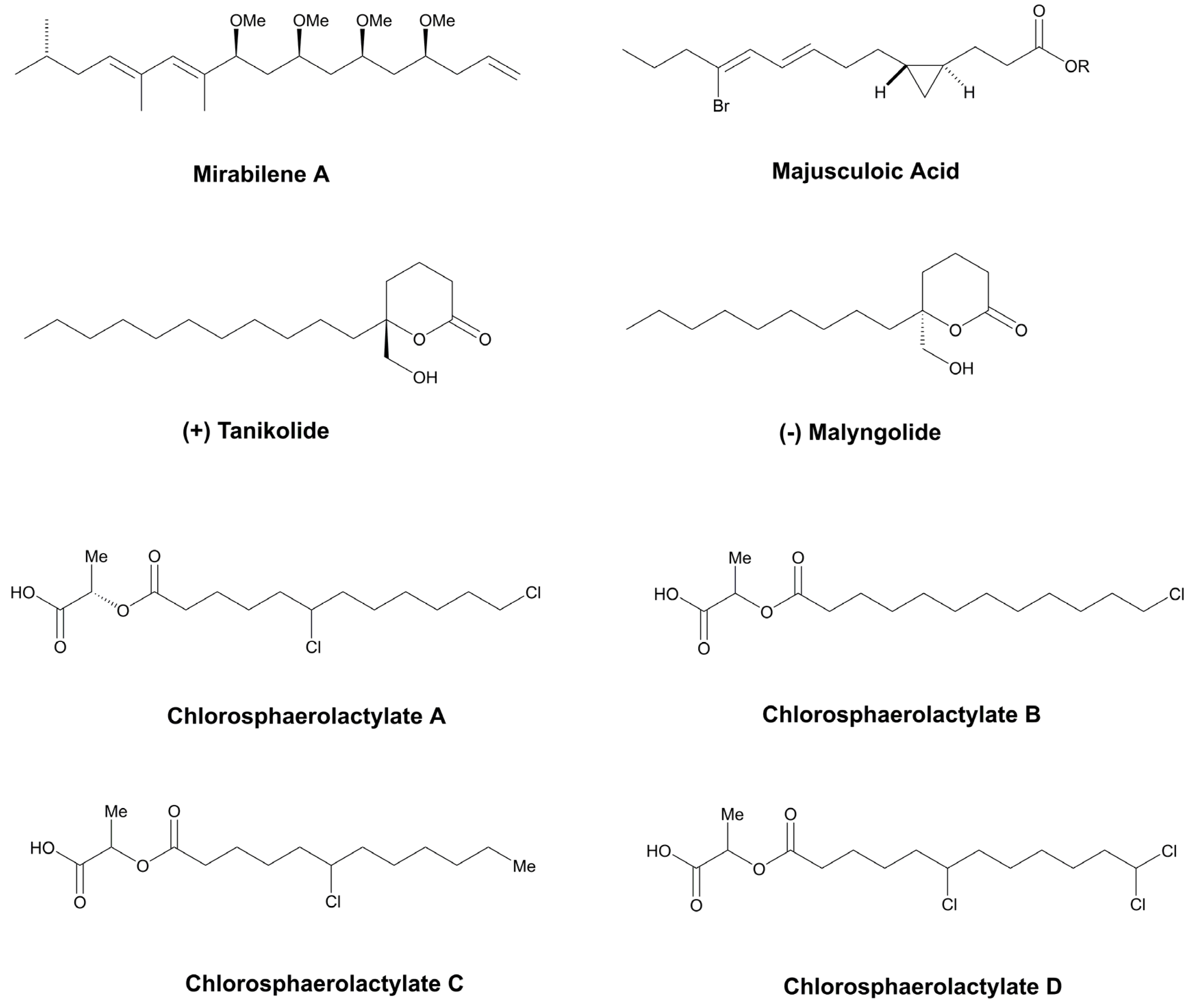{kind=link}
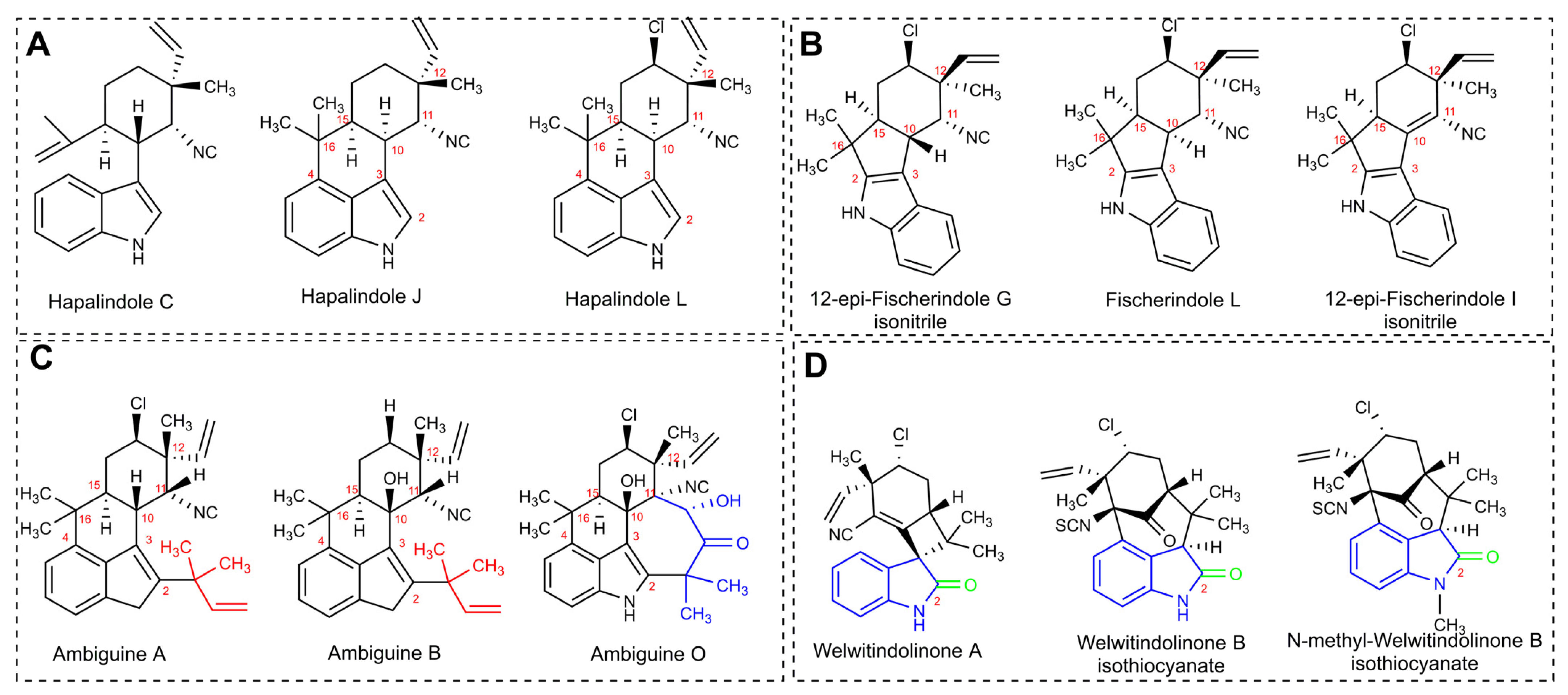{kind=link}
{kind=link}
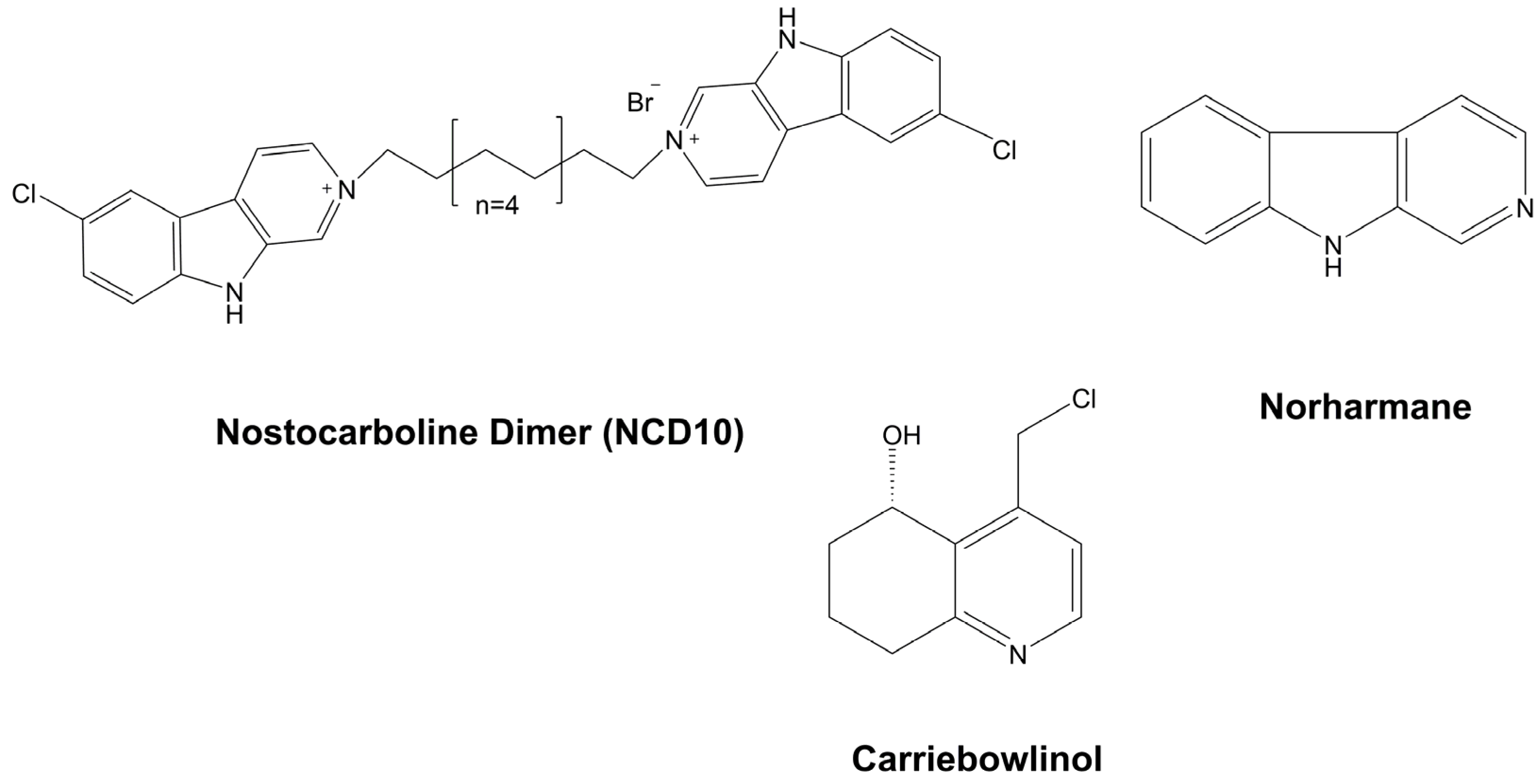{kind=link}
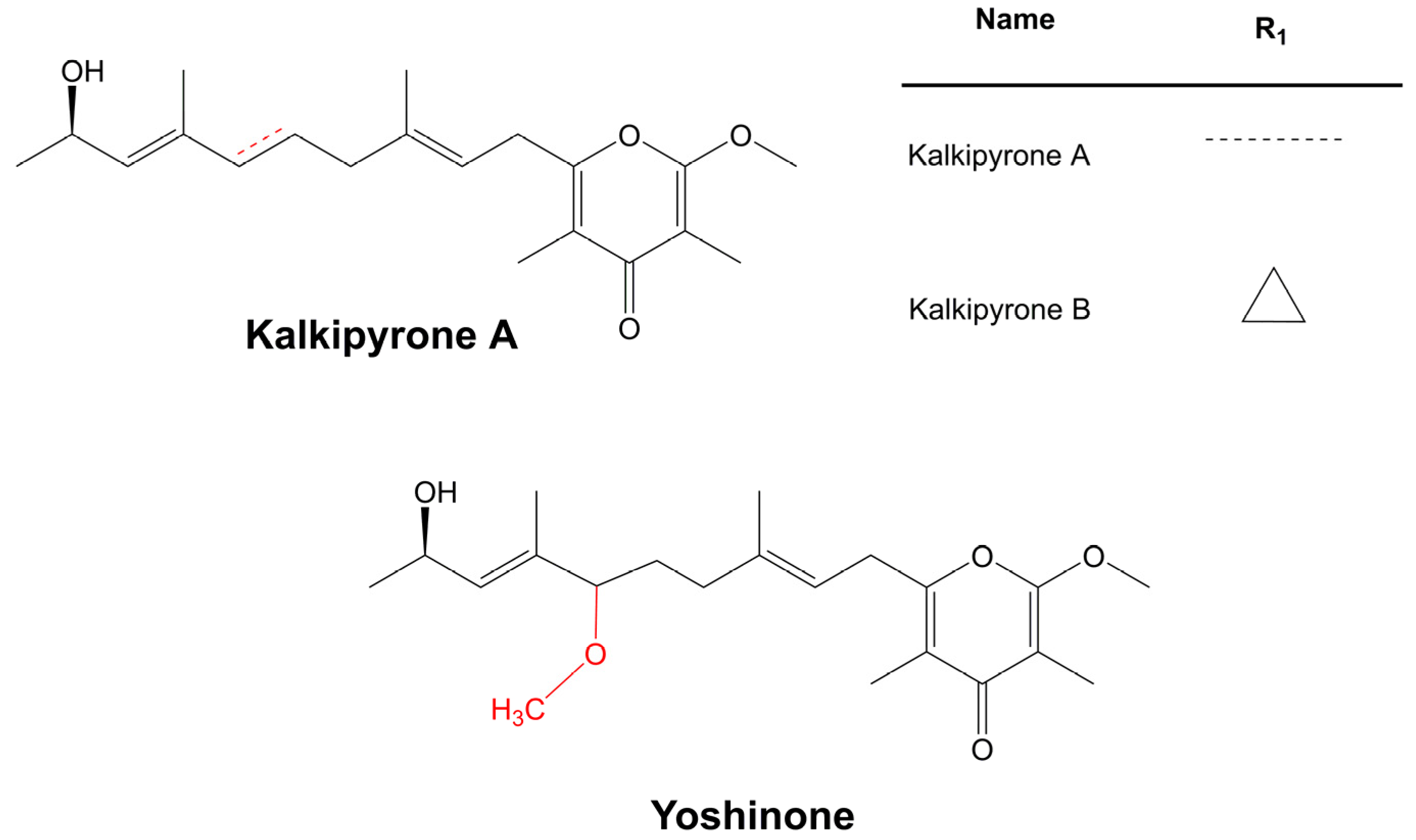{kind=link}
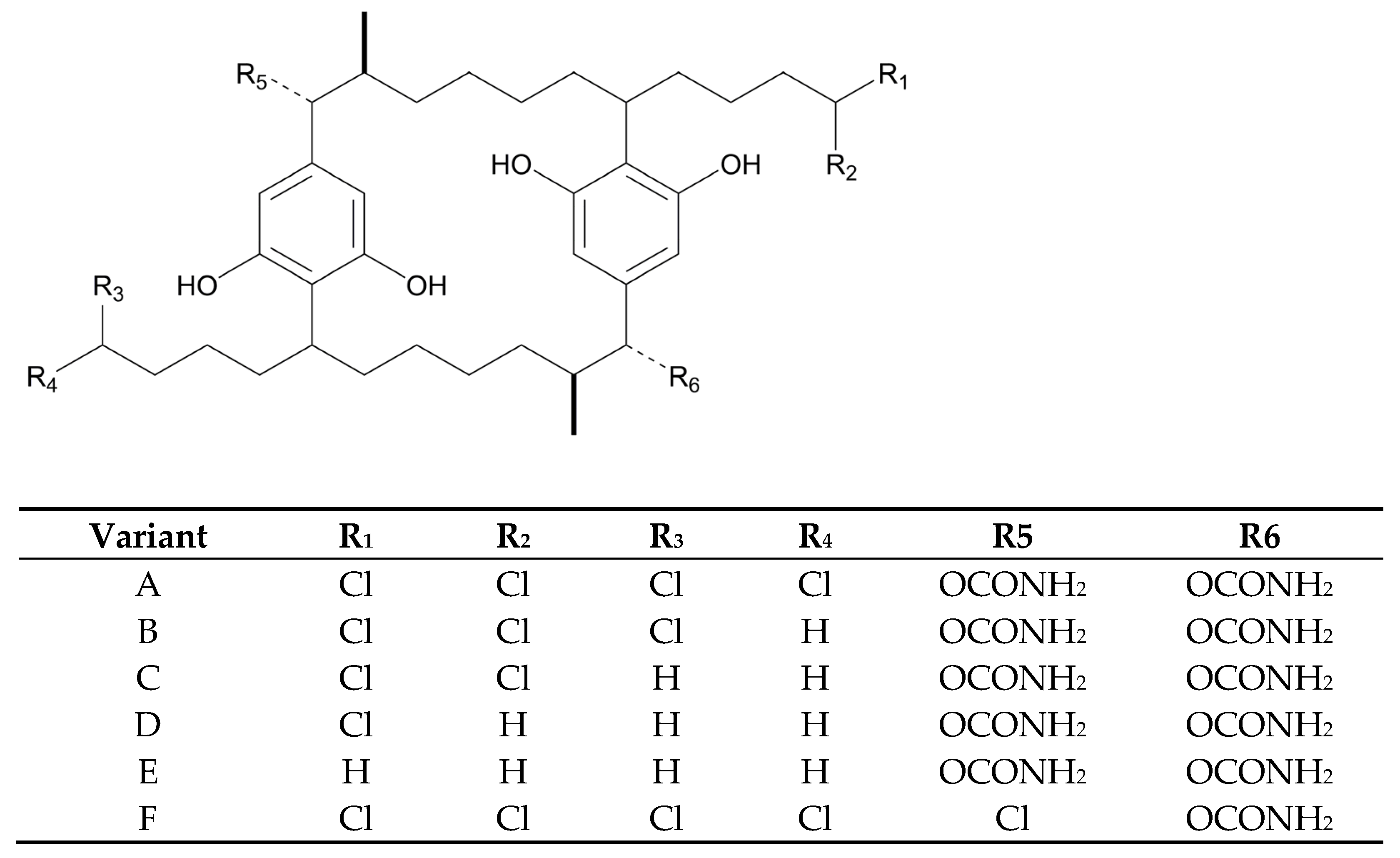{kind=link}
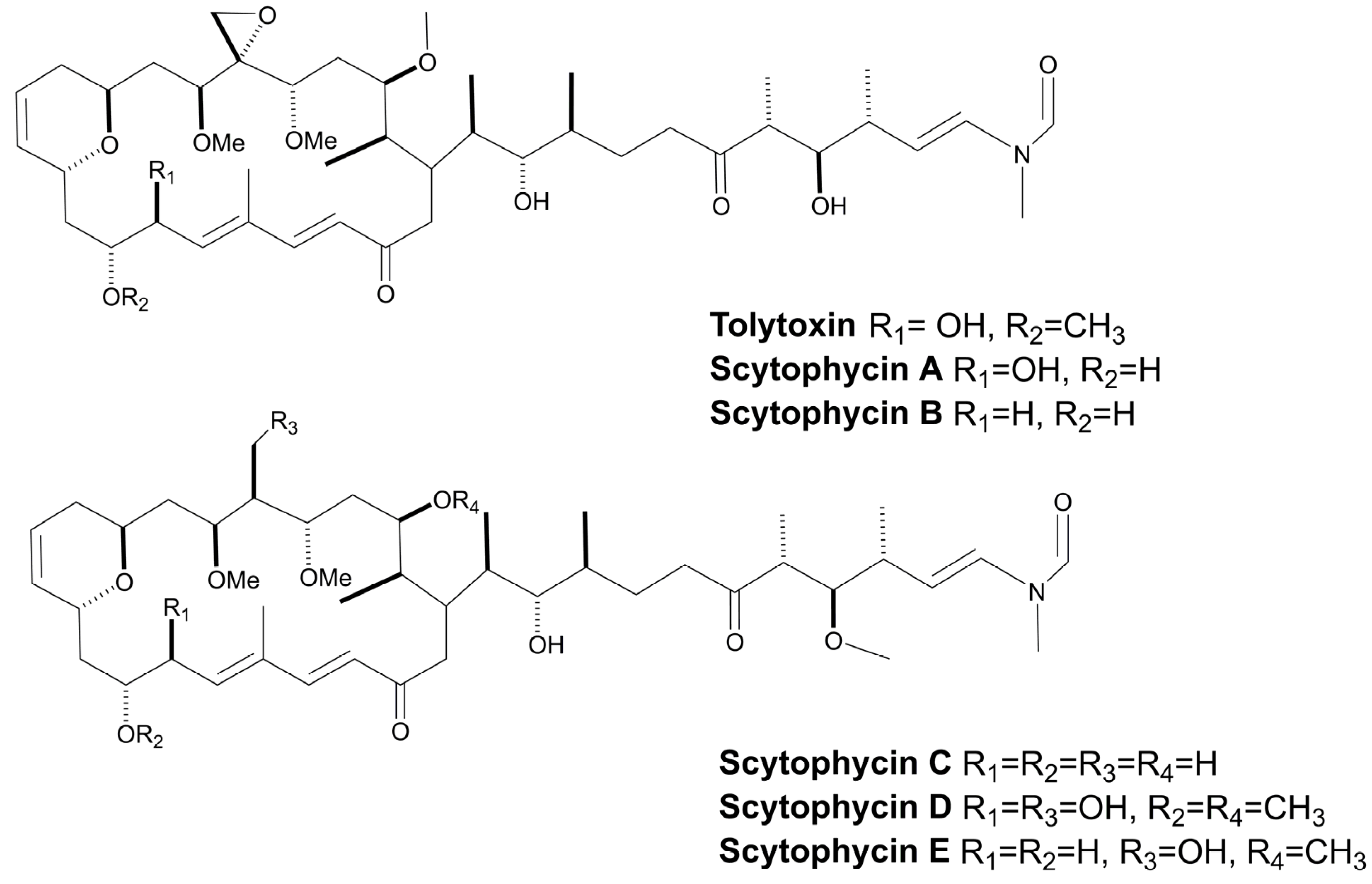{kind=link}
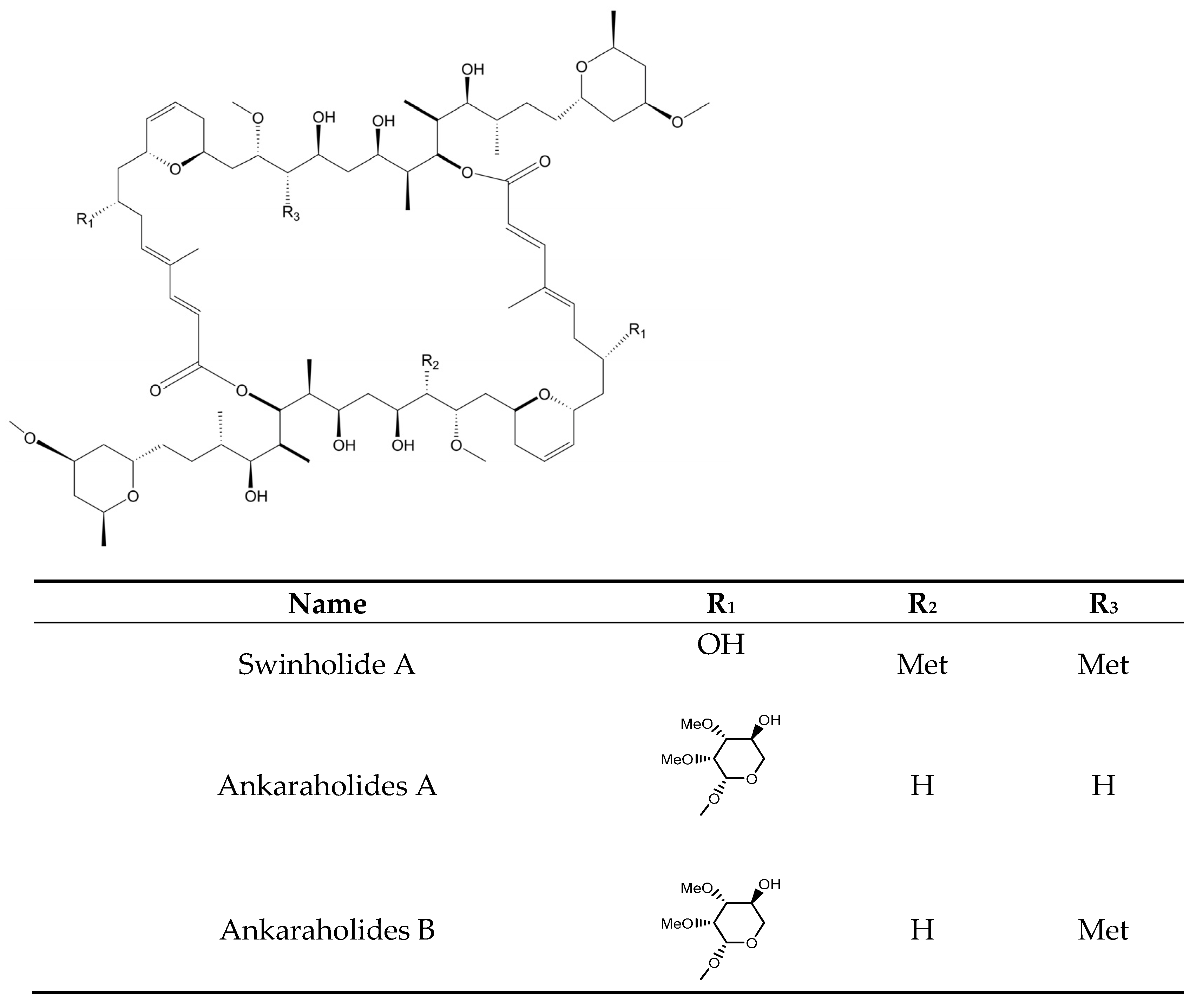{kind=link}
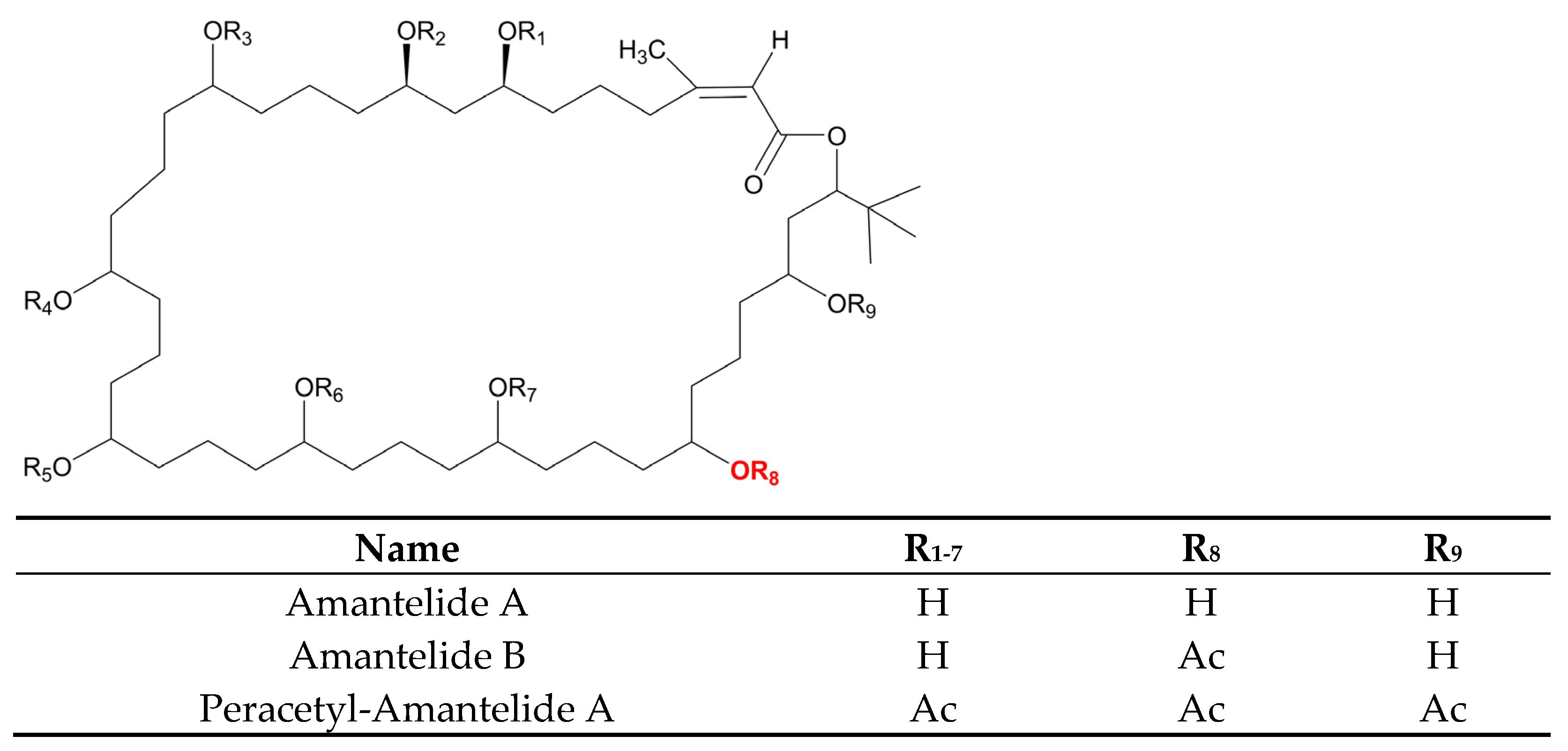{kind=link}
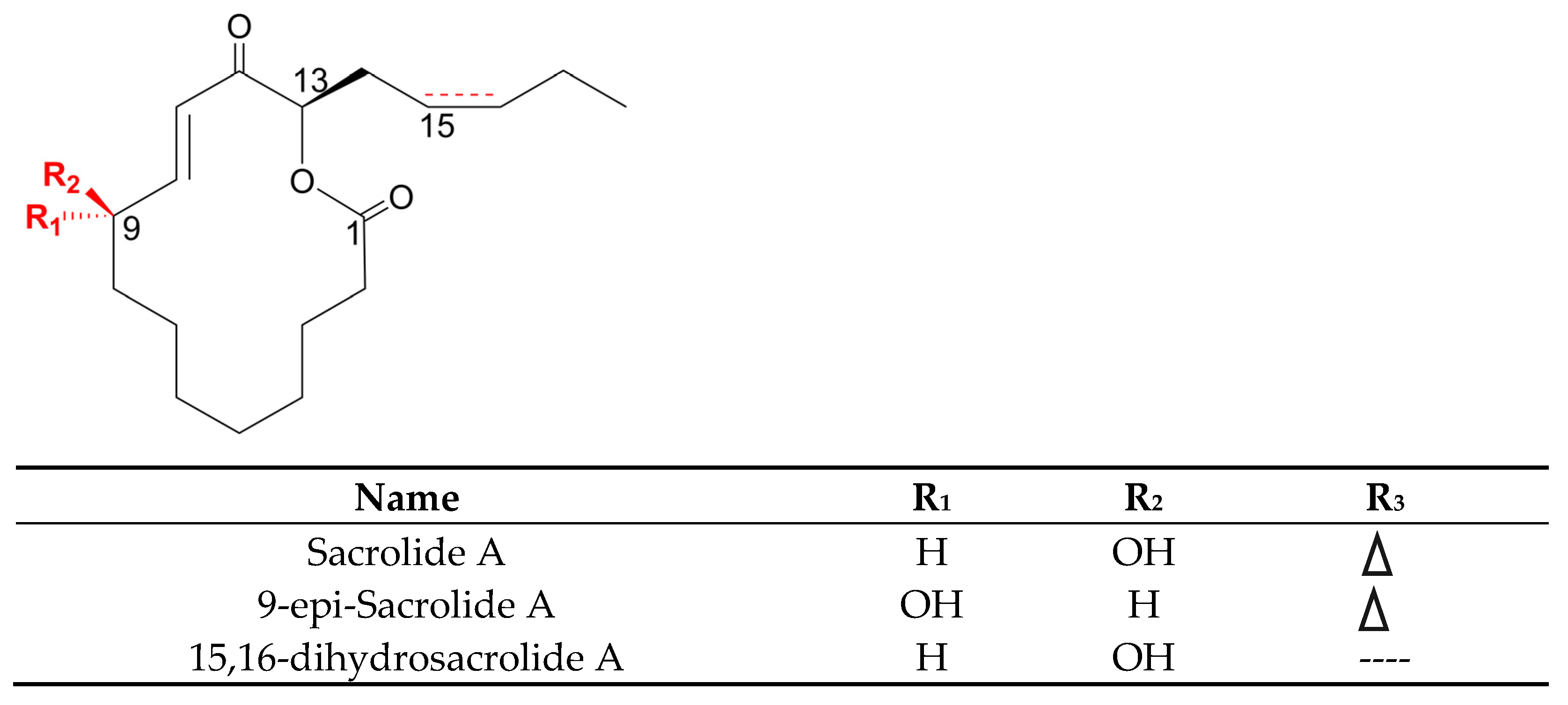{kind=link}
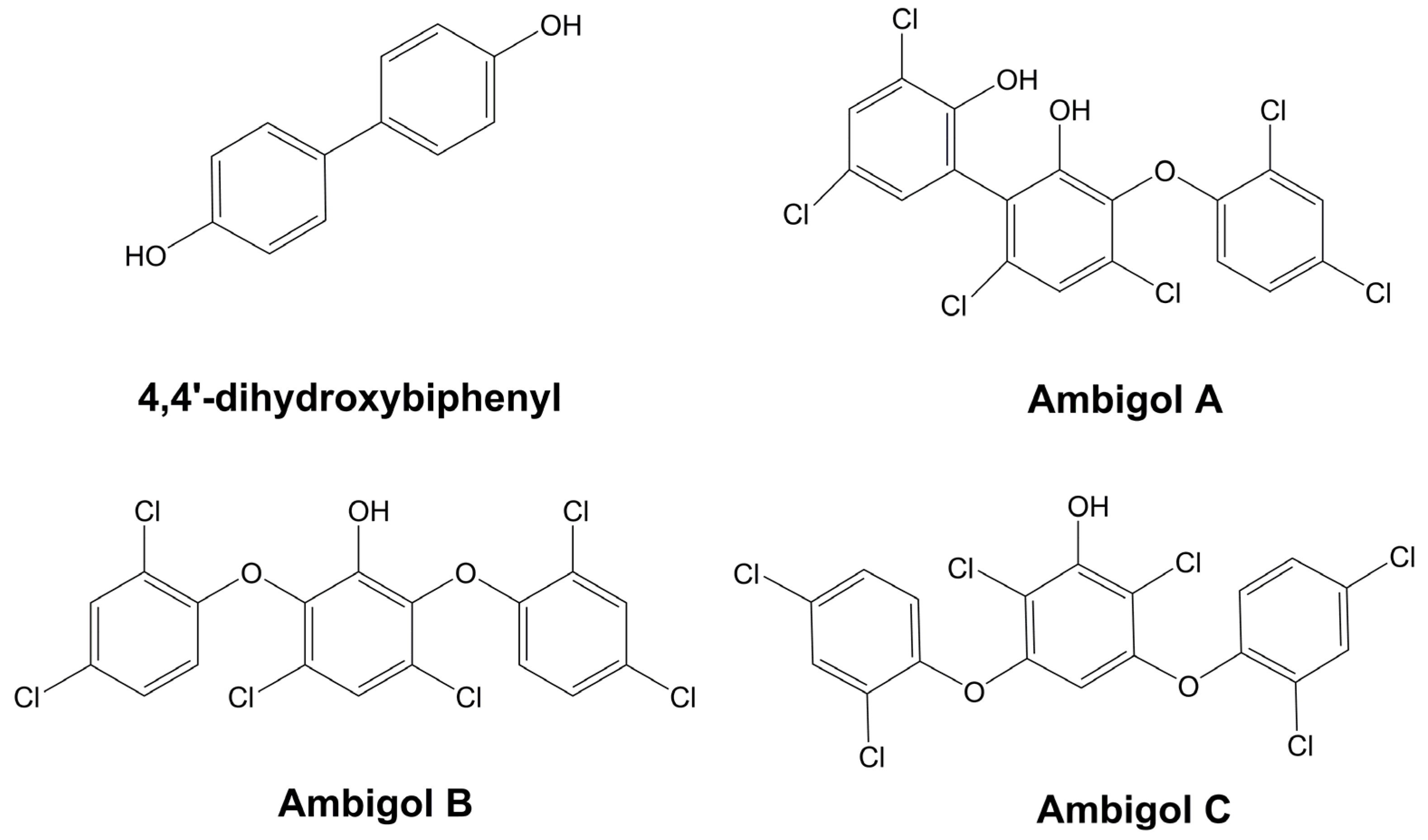{kind=link}
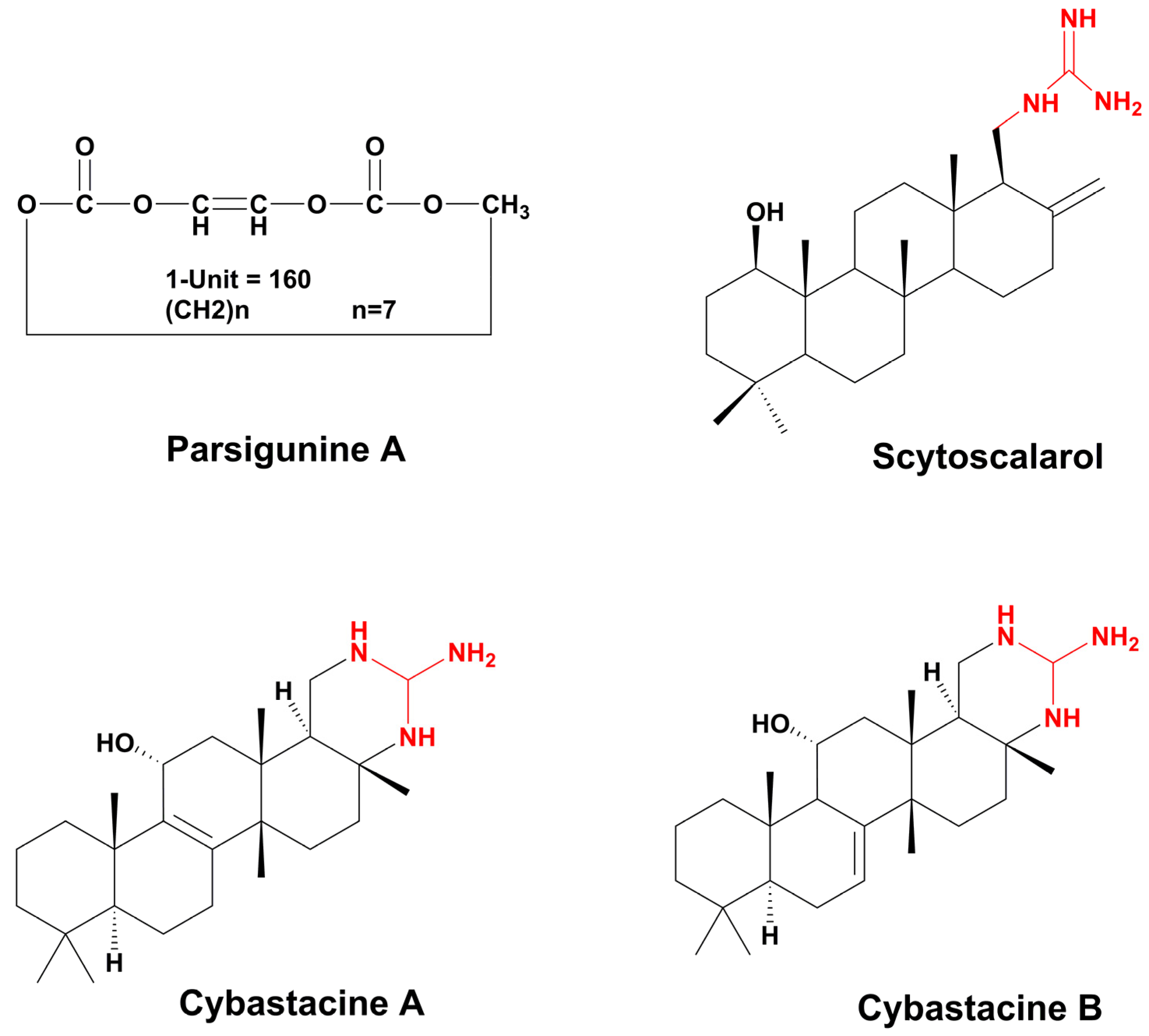{kind=link}
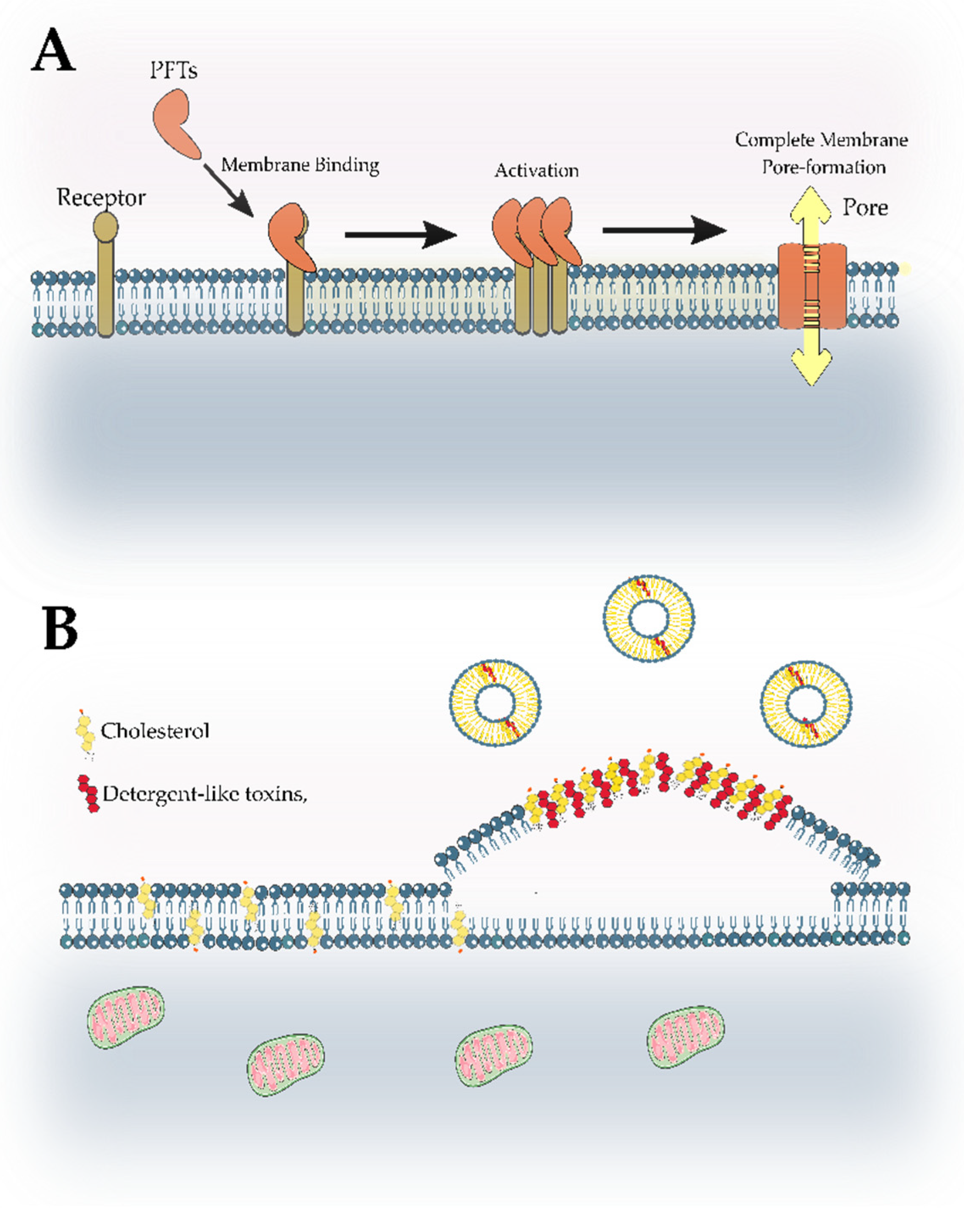{kind=link}
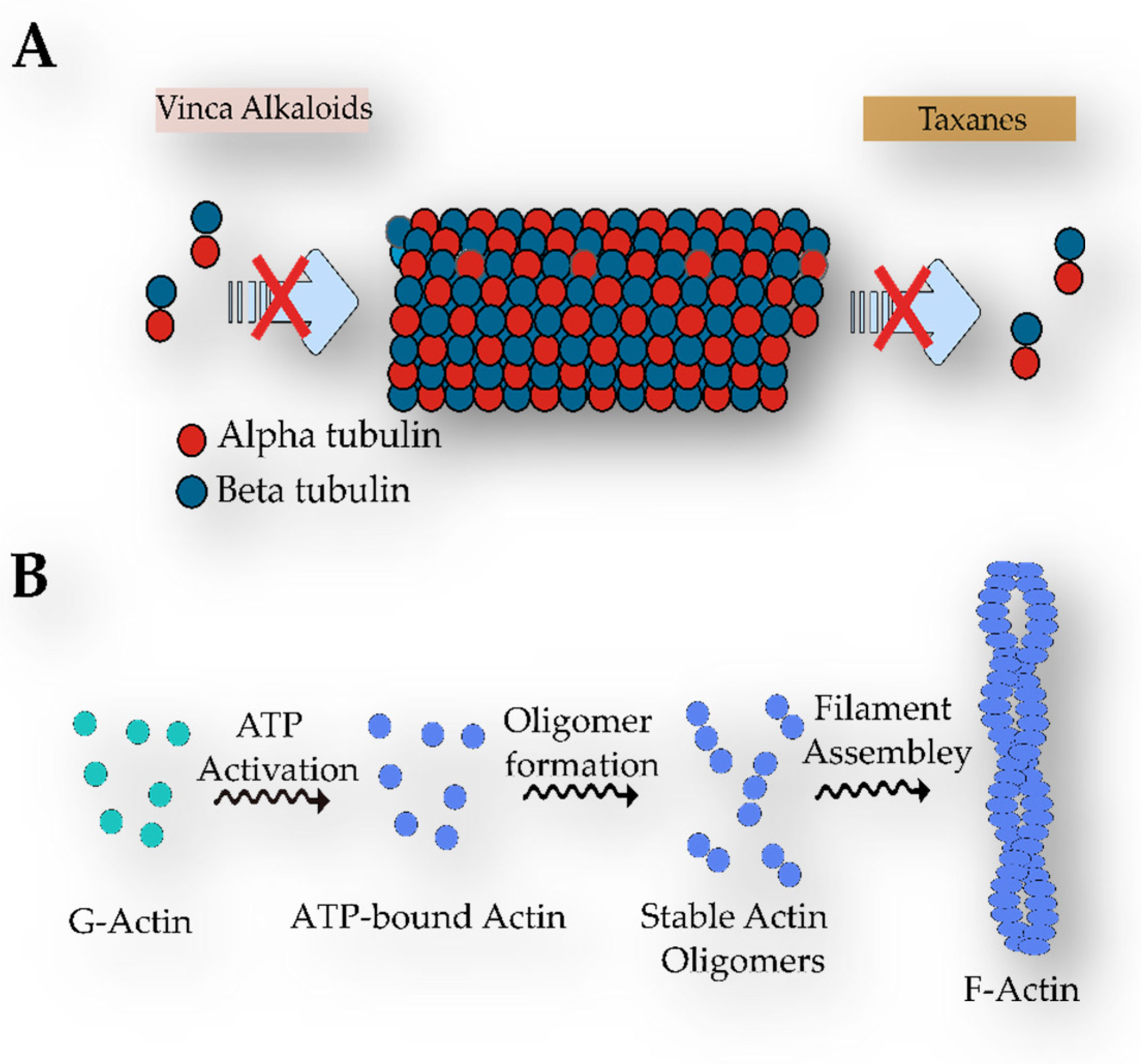{kind=link}
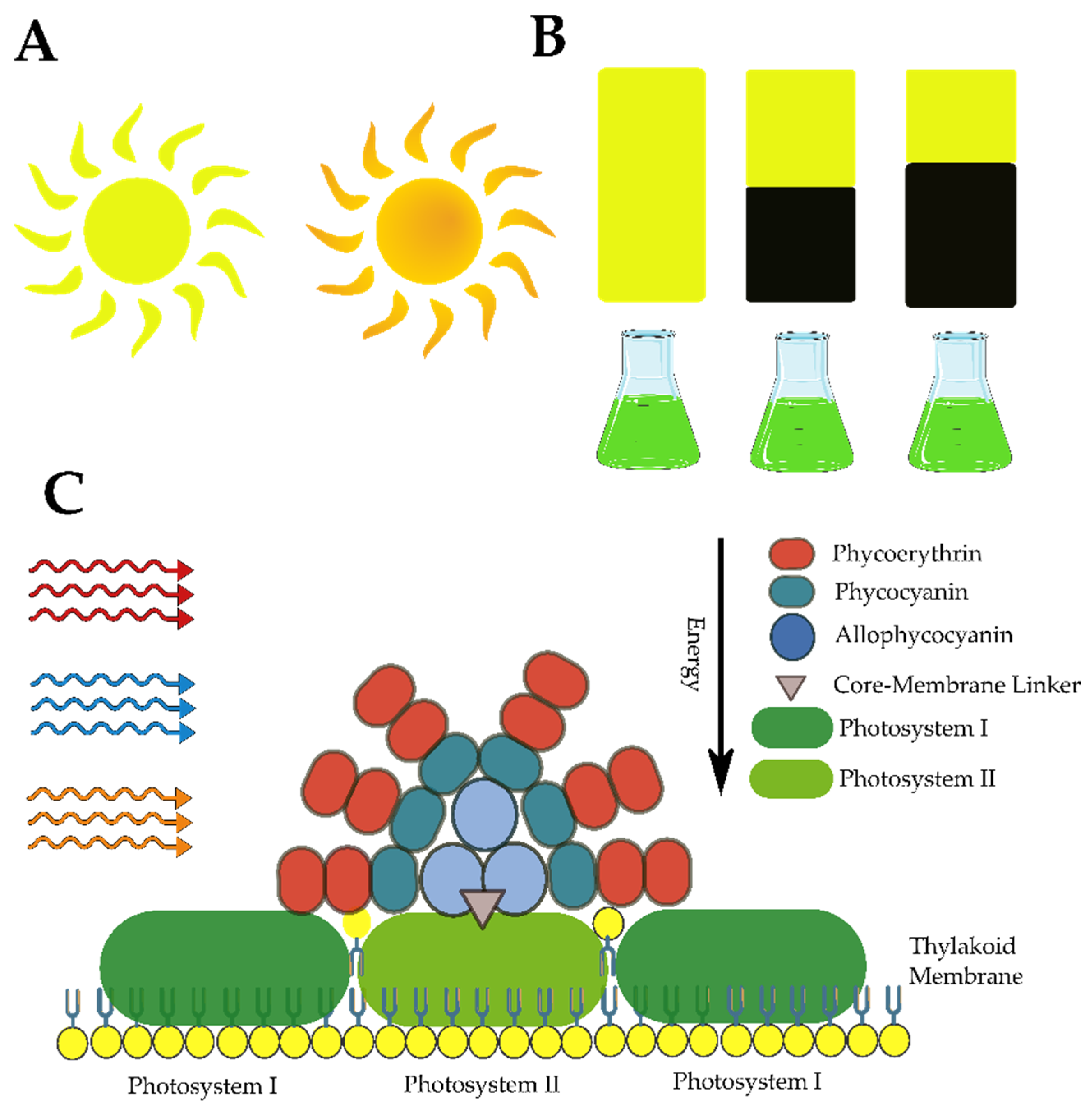{kind=link}
| Compound | Molecular Weight | Cyanobacterial Strain (Family) | Source | Location | Ref |
|---|---|---|---|---|---|
| Schizotrin A | 1490.71 | Schizothrix sp. IL-89-2 (Schizotrichaceae) | Terrestrial | Kfar Azar (Israel) | [33] |
| Pahayokolides A | 1472.68 | Lyngbya sp. 15-2 (Oscillatoriaceae) | Freshwater | Everglades (Florida, U.S) | [38] |
| Muscotoxins A | 1211.41 | Desmonostoc muscorum CCALA 125 (Nostocaceae) | Terrestrial | Dlouhá ves (Chelcice, Czech Republic) | [39,40] |
| Muscotoxins B | 1225.45 | ||||
| Calophycin | 1249.48 | Calothrix fusca EU-10-1 (Rivulariaceae) | Freshwater | Island of Oahu (Hawaii, U.S) | [36] |
| Puwainaphycin F | 1146.34 | Cylindrospermum alatosporum CCALA 988 (Aphanizomenonaceae) | Terrestrial | Riding Mountain National Park (Manitoba, Canada) | [41] |
| Minutissamide A | 1118.28 | Nostocales LEGE 17548 | Freshwater | Lagoa de Mira (Mira, Portugal) | [42] |
| Anabaena minutissima UTEX 1613 (Aphanizomenonaceae) | Terrestrial | South Texas, USA | [43,44] | ||
| Nostofungicidine | 885.57 | Nostoc commune (Nostocaceae) | Terrestrial | Gulf of Finland (Porkkala, Finland) | [45] |
| Anabaenolysin A | 558.63 | Benthic Anabaena strains (Aphanizomenonaceae) | Marine | Island of Oahu (Hawaii, U.S) | [46,47] |
| Anabaenolysin B | 560.64 | ||||
| Laxaphycin A | 1196.48 | Anabaena laxa FK-1-2 (Aphanizomenonaceae) | Terrestrial | Island of Oahu (Hawaii, U.S) | [48] |
| Hormothamnion Enteromorphoides (Nostocaceae) | Marine | Key West (Florida, U.S) | [49] | ||
| Lyngbya majuscula (Oscillatoriaceae) | Marine | Moorea atoll (French Polynesia) | [50] | ||
| Anabaena torulosa (Aphanizomenonaceae) | Marine | Moorea atoll (French Polynesia) | [51] | ||
| Laxaphycin B | 1395.70 | A. laxa FK-1-2 (Aphanizomenonaceae) | Terrestrial | Island of Oahu (Hawaii, U.S) | [48] |
| L. majuscula (Oscillatoriaceae) | Marine | Moorea atoll (French Polynesia) | [49] | ||
| A. torulosa (Aphanizomenonaceae) | Marine | Moorea atoll (French Polynesia) | [50] | ||
| Laxaphycin C | 1379.69 | A. laxa FK-1-2 (Aphanizomenonaceae) | Terrestrial | Island of Oahu (Hawaii, U.S) | [48] |
| Scytocyclamide A | 1223.52 | Scytonema hofmannii PCC 7110 (Scytonemataceae) | Terrestrial | Bermuda (United Kingdom) | [52] |
| Scytocyclamide A2 | 1208.51 | ||||
| Scytocyclamide B | 1367.65 | ||||
| Scytocyclamide B2 | 1352.64 | ||||
| Scytocyclamide B3 | 1336.64 | ||||
| Scytocyclamide C | 1351.65 | ||||
| Heinamides | 1186.65–1518.85 | Nostoc sp. UHCC 0702 (Nostocaceae) | Freshwater | Villähteen Kukkanen (Nastola, Finland) | [53] |
| Hormothamnin A | 1196.48 | H. Enteromorphoides (Nostocaceae) | Marine | La Parguera (Puerto Rico Caribbean) | [54] |
| Microcolin A | 747.96 | Lyngbya cf. polychroa (Oscillatoriaceae) | Marine | Coast of Hollywood (Florida, U.S) | [55] |
| Microcolin B | 731.96 | L. majuscula (Oscillatoriaceae) | Marine | La Blanquilla Island (Venezuela) | [56] |
| Moorea producens (Oscillatoriaceae) | Marine | Playa Kalki (Curaçao) | [57] | ||
| Oscillatoria sp. LP16 (Oscillatoriaceae) | Marine | Broward County (Florida, U.S) | [58] | ||
| Lobocyclamide A | 1198.45 | Lyngbya confervoides (Oscillatoriaceae) | Marine | Cay Lobos (Bahamas, Caribbean) | [59,60] |
| Lobocyclamide B | 1398.70 | ||||
| Lobocyclamide C | 1370.65 | ||||
| Nostocyclamide | 474.55 | Nostoc sp. 31 (Nostocaceae) | Freshwater | New Jersey (United States | [61,62] |
| Hectochlorin | 665.59 | L. majuscula UOG VP417 (Oscillatoriaceae) | Marine | Apra Harbor (Guam) | [63] |
| M. producens JHB (Oscillatoriaceae) | Hector Bay (Jamaica) | [64] | |||
| Lyngbyabellin B | 665.59 | L. majuscula JHB-22 (Oscillatoriaceae) | Marine | Hector Bay (Portland, Jamaica) | [65] |
| M. producens RS05 (Oscillatoriaceae) | Sharm el-Sheikh (Egypt) | [66] | |||
| Hassallidin A | 1382.52 | Hassallia sp. B02-07 (Tolypothrichaceae) | Marine | Orrido Clough (Bellano, Italy) | [67,68] |
| Hassallidin B | 1528.66 | ||||
| Hassallidin D | 1865.00 | Anabaena sp. SYKE748A (Aphanizomenonaceae) | Marine | Tuusulanjarvi Lake (Tuusulanjarvi, Finland) | [69] |
| Hassallidin E | 1410.58 | Planktothrix serta PCC 8927 (Microcoleaceae) | Marine | Berre le Clos (França) | [70] |
| Balticidin A | 1787.28 | Anabaena cylindrica Bio33 (Aphanizomenonaceae) | Marine | Baltic Sea (Rügen Island, Germany) | [71] |
| Balticidin B | 1769.26 | ||||
| Balticidin C | 1752.83 | ||||
| Balticidin D | 1734.81 | ||||
| Tolybyssidin A | 1465.80 | Tolypothrix byssoidea EAWAG 195 (Tolypothrichaceae) | Marine | Pokhara (Nepal) | [72] |
| Tolybyssidin B | 1491.86 | ||||
| Cryptophycin A | 655.19 | Nostoc sp. ATCC 53789 (Nostocaceae) | Terrestrial | Isle of Arran (Scotland) | [73,74] |
| Metabolite | Target | Activity | Ref | |
|---|---|---|---|---|
| Metabolite | Control | |||
| Anabaenolysin A | C. albicans HAMBI 261 | - | - | [47] |
| Anabaenolysin B | C. albicans HAMBI 261 | - | - | |
| Balticidin A | Candida maltosa SBUG700 | 12 mm | - | [71] |
| Balticidin B | 15 mm | - | ||
| Balticidin C | 9 mm | - | ||
| Balticidin D | 18 mm | - | ||
| Calophycin 1.2 µg | Aspergillus oryzae | 13 mm | No activity Amphotericin B | [36] |
| Penicillium notatum | 12 mm | No activity Amphotericin B | ||
| S. cerevisiae | 12 mm | 9 mm Amphotericin B | ||
| Calophycin | Trichophyton mentagrophytes | 2 µM | - | |
| Aspergillus fumigatus | 1 µM | 1.35 µM Amphotericin B | ||
| C. albicans | 1µM | 0.67 µM Amphotericin B | ||
| Cryptophycin A | Rhizonucor miehei, Scopulariopsis communes, Trichoderma lignorum, Verticilium serrae, Cryptococcus albidus, Cryptococcus laurentii, Ustilago zea, and Cercospora beticola. | 10–15 mm | - | [74] |
| A. fumigatus, Cephalosporium sp., F. oxysporum, and Ceratocystis ulmi | 16–20 mm | - | ||
| Alternaria solani, Aspergillus flavus, Aspergillus niger, Botrytis ali, Penicillium sp., Cochliobolus miyabeanus, Phona sp. | 20–25 mm | - | ||
| Cryptococcus neoformans MY1051 and MY1146 | ≤0.1 µM | <1.1 µM Amphotericin B | ||
| C. neoformans strains | 0.046 µM | |||
| Hassallidin A | C. albicans strains Candida guillermondii ATCC 90877 C. tropicalis ATCC 750 | 2.9 µM | 0.23–0.91 µM Caspofungin | [67,68,75] |
| Cryptococcus neoformans Trichosporon asahii Trichosporon mucoides | 7.3 -29.3 µM Caspofungin | |||
| Candida glabrata strains Candida parapsilosis strains C. tropicalis strains Candida krusei strains | 5.2 µM | 0.23–0.91 µM Caspofungin | ||
| Aspergillus niger Ustilago maydis Penicillium sp. Fusarium sambucium. | - | - | ||
| Hassallidin B | C. parapsilosis ATCC 22019 C. krusei ATCC 6258 C. albicans strains C. tropicalis ATCC 90874 | 5.2 µM | 0.23–0.91 µM Caspofungin | |
| C. neoformans strains | 7.3–29.3 µM Caspofungin | |||
| C. albicans ATCC 24433 C. glabrata strains C. parapsilosis ATCC 90018 C. tropicalis ATCC 750 C. krusei ATCC 90878 | 10.5 µM | 0.23–0.91 µM Caspofungin | ||
| Aspergillus niger Ustilago maydis Penicillium sp. Fusarium sambucium. | - | - | ||
| A. fumigatus and C. albicans | 3.1 µM | - | ||
| Hassallidin D | C. albicans ATCC 11006 C. albicans ATCC 10231 C. krusei ATCC 6258 | 1.5 µM | - | [69] |
| Hassallidin D 10 μg | Cryptococcus albidus ATCC 10666 | 17 mm | - | [69] |
| Filobasidiella neoformans ATCC 10226 | 11 mm | - | ||
| Hassallidin D Linear Form | C. albicans ATCC 11006 | 20 µM | - | |
| Hassallidin E | C. albicans CBS562 | 22.7 µM | - | [70] |
| C. neoformans H99 | - | |||
| C. parapsilosis ATCC22019 | - | |||
| C. krusei ATCC6258 | - | |||
| Hectochlorin 100 µg | C. albicans | 16 mm | - | [76] |
| Heinamides | Aspergillus flavus FBCC 2467 | - | - | [53] |
| Hormothamnins (10 µg) | C. albicans | 7–11 mm | - | [77] |
| Laxaphycins A + B (25 + 25 µg) | A. oryzae | 29 mm | - | [48] |
| C. albicans | 23 mm | - | ||
| P. notatum | 30 mm | - | ||
| S. cerevisiae | 22 mm | - | ||
| T. mentagrophytes | 30 mm | - | ||
| Laxaphycin B (50 µg) | A. oryzae | 19 mm 45.8 µM 4.6 µM (8.1 µM of Laxa A) | ||
| C. albicans | 8 mm | - | ||
| S. cerevisiae | 12 mm | - | ||
| T. mentagrophytes | 9 mm | - | ||
| Laxaphycin C (50 µg) | A. oryzae | 14 mm | - | |
| C. albicans | 9 mm | - | ||
| P. notatum | 11 mm | - | ||
| S. cerevisiae | 12 mm | - | ||
| Lobocyclamide A (150 µg) | C. albicans 96-489 | 7 mm | - | [59] |
| Lobocyclamide B (150 µg) | C. albicans 96-489 | 8 mm | - | |
| C. glabrata | 6 mm | - | ||
| Lobocyclamide C (150 µg) | C. albicans 96-489 | 10 mm | - | |
| C. glabrata | 8 mm | - | ||
| Lyngbyabellin B (100 µg) | C. albicans ATCC 14053 | 10.5 mm | - | [78] |
| Microcolin A | Dendryphiella salina SIO D. salina EBGJ | >250 µM | 3.4 µM Amphotericin B | [55] |
| Microcolin B | ||||
| Minutissamide A | A. fumigatus | 37.5 µM | - | [43] |
| Alternaria alternata | 75 µM | - | ||
| Muscotoxins A | Candida friedrichii | 61.9 µM | 26.1 µM Fluconazole | [40] |
| Trichoderma harzianum | 31.5 µM | 13.1 µM Fluconazole | ||
| Bipolaris sorokiniana | NA | NA Fluconazole | ||
| Alternaria alternata | 0.5 µM | 6.5 µM Fluconazole | ||
| Monographella cucumerina | 1.9 µM | 6.5 µM Fluconazole | ||
| A. fumigatus | 1.9 µM | 104.5 µM Fluconazole | ||
| Chaetomium globosum | 15.5 µM | 6.5 µM Fluconazole | ||
| Fusarium oxysporum | 61.9 µM | 26.1 µM Fluconazole | ||
| S. sclerotiorum | 41.3 µg * | - | ||
| Muscotoxins B | S. sclerotiorum | 20.4 µg * | - | |
| Nostocyclamide | S. cerevisiae | - | - | [62] |
| Nostofungicidine | Aspergillus candidus | 1.8 µM | - | [45] |
| Pahayokolides A | S. cerevisiae | 20 mm | - | [38] |
| Puwainaphycin F | C. albicans HAMBI 261 | 5.5 µM | - | [79] |
| S. cerevisiae HAMBI 1164 | - | |||
| Schizotrin A (13.4–16.7 nM) | S. cerevisiae C. albicans | 7 mm | - | [33] |
| C. tropicalis | 9 mm | - | ||
| R. rubra | 8 mm | - | ||
| S. rolfsii R. solani | 25–28% | - | ||
| C. gloeosporioides | 47% | - | ||
| Schizotrin A 33.5 nM | F. oxysporum | 37% | - | |
| Scytocyclamide A 200 µg | A. flavus FBCC 2467 | 10 mm | - | [52] |
| Scytocyclamide A2 200 µg | 7 mm | - | ||
| Scytocyclamide B 600 µg | 23 mm | - | ||
| Scytocyclamide B2 85 µg | 10 mm | - | ||
| Scytocyclamide B3 85 µg | 20 mm | - | ||
| Scytocyclamide C 160 µg | 22 mm | - | ||
| Scytocyclamide A + B 100 + 300 µg | 36 mm | - | ||
| Scytocyclamide A + C 100 + 80 µg | 33 mm | - | ||
| Scytocyclamide A2 + B2 100 + 43 µg | 24 mm | - | ||
| Scytocyclamide A2 + B3 100 + 43 µg | 25 mm | - | ||
| Scytocyclamide B + C 300 + 80 µg | 23 mm | - | ||
| Tolybyssidin A | Candida albicans | 21.8 µM | 19.2 µM Miconazole | [72] |
| Tolybyssidin B | 42.9 µM | |||
| 7-Deoxysedoheptulose | S. cerevisiae | 50 µM | 590 μM Glyphosate | [80] |
| Polysaccharide 10 µg | T. rhizoctonia | 15 mm | 23 mm Amphotericin | [81] |
| F. solani | 16 mm | 27 mm Amphotericin | ||
| F. oxysporum | 15 mm | 28 mm Amphotericin | ||
| A. niger | 17 mm | 25 mm Amphotericin | ||
| C. albicans ATCC 90028 | 15 mm | 20 mm Amphotericin | ||
| Polysaccharide | Botrytis cinerea | 1064 mg mL−1 (EC50) | - | [82] |
| Polysaccharide | A. niger | 707 mg mL−1 | - | [83] |
| Chlorosphaerolactylate A | C. parapsilosis SMI416 | 3 mM | [84] | |
| Chlorosphaerolactylates B–C | 3.3 mM | - | ||
| Chlorosphaerolactylate D | 5.5 mM | - | ||
| Majusculoic Acid | C. albicans ATCC 14503 | 8 µM | 1 µM Fluconazole | [85] |
| C. glabrata | 19.3 µM | - | ||
| Mirabilenes A–F 10 μg | P. notatum | 10–15 mm | - | [86] |
| A. oryzae | ||||
| Lyngbic Acid | Fusarium sp. | 2.1 µMIC50 | - | [87] |
| Lindra thalassiae | 2.7 µMIC50 | - | ||
| D. salina | 2.9 µMIC50 | - | ||
| Tanikolide 100 μg | C. albicans | 13 mm | - | [88] |
| Ambiguine A | C. albicans | 6.1 µM | 0.34 µM Amphotericin B | [89,90,91] |
| T. mentagrophytes | 24.6 µM | 0.06 µM Tolnaftate | ||
| A. fumigatus | 196.6 µM | 2.3 µM Amphotericin B | ||
| Ambiguine B | C. albicans | 5.9 µM | 0.34 µM Amphotericin B | |
| T. mentagrophytes | 5.9 µM | 0.06 µM Tolnaftate | ||
| A. fumigatus | 47.3 µM | 2.3 µM Amphotericin B | ||
| Ambiguine C | C. albicans | 3.2 µM | 0.34 µM Amphotericin B | |
| T. mentagrophytes | 1.6 µM | 0.06 µM Tolnaftate | ||
| A. fumigatus | >20.6 µM | 2.3 µM Amphotericin B | ||
| Ambiguine D | C. albicans | 2.8 µM | 0.34 µM Amphotericin B | |
| T. mentagrophytes | 1.4 µM | 0.06 µM Tolnaftate | ||
| A. fumigatus | 176.6 µM | 2.3 µM Amphotericin B | ||
| Ambiguine E | C. albicans | 5.7 µM | 0.34 µM Amphotericin B | |
| T. mentagrophytes | 5.7 µM | 0.06 µM Tolnaftate | ||
| A. fumigatus | >183.1 µM | 2.3 µM Amphotericin B | ||
| Ambiguine F | C. albicans | 2.7 µM | 0.34 µM Amphotericin B | |
| T. mentagrophytes | 2.7 µM | 0.06 µM Tolnaftate | ||
| A. fumigatus | >175.8 µM | 2.3 µM Amphotericin B | ||
| Ambiguine G | C. albicans | >100 µM | 0.03 µM Amphotericin B | |
| Ambiguine I | S. cerevisiae | 1.5 µM | 0.57 µM Puromycin | |
| C. albicans ATCC 90028 | 1.5 µM | 1.7 µM Amphotericin B | ||
| Ambiguine K | C. albicans | <0.9 µM | 0.03 µM Ketoconazole | [92] |
| Ambiguine L | <1.0 µM | |||
| Ambiguine M | 1.1 µM | |||
| Ambiguine N | <1.0 µM | |||
| Ambiguine O | <1.0 µM | |||
| Ambiguine P | 32.9 µM | |||
| Anhydrohapaloxidole A | C. albicans | 1.9 µM | 0.12 µM Amphotericin B | [93] |
| Carriebowlinol | Fusarium sp. | 0.2 µMIC50 | - | [87] |
| L. thalassiae | 0.4 µM IC50 | - | ||
| D. salina | 0.5 µM IC50 | - | ||
| Fischerindole L | C. albicans | 1.2 µM | 0.12 µM Amphotericin B | [93] |
| 12-epi-fischerindole U | C. albicans SC5314 | 1.2 µM | - | [94] |
| 12-epi-fischerindole G | 1.6 µM | - | ||
| 13R-Bromo 12-epi-fischerindole U | 2.5 µM | - | ||
| Fischambiguine A | C. albicans | 15.3 µM | 0.12 µM Amphotericin B | [92] |
| Hapalindole A | C. albicans | 3.7 µM | - | [95] |
| T. mentagrophytes | 3.7 µM | - | ||
| Hapalindole B | C. albicans | >53.9 µM | - | |
| T. mentagrophytes | >53.9 µM | - | ||
| Hapalindole C | C. albicans | 2.1 µM | - | |
| T. mentagrophytes | 2.1 µM | - | ||
| Hapalindole D | C. albicans | 59.4 µM | - | |
| T. mentagrophytes | 59.4 µM | - | ||
| Hapalindole E | C. albicans | 0.9 µM | ||
| T. mentagrophytes | 1.8 µM | - | ||
| Hapalindole F | C. albicans | 53.9 µM | - | |
| T. mentagrophytes | 53.9 µM | - | ||
| Hapalindole G | C. albicans | 29.5 µM | - | |
| T. mentagrophytes | 7.4 µM | - | ||
| Hapalindole H | C. albicans | 32.9 µM | - | |
| T. mentagrophytes | 4.1 µM | - | ||
| C. albicans | <0.6 µM | - | ||
| Hapalindole A | C. albicans | 1.2 µM | 0.12 µM Amphotericin B | [93] |
| Hapalindole J | 0.7 µM | 0.12 µM Amphotericin B | ||
| Hapalindole X | 2.5 µM | 0.12 µM Amphotericin B | ||
| Hapalonamide H | <0.6 µM | 0.12 µM Amphotericin B | ||
| Nostocarboline | S. cerevisiae A-136 | 4.6 µM | 7.9 µM Chlorhexidine | [96] |
| C. albicans T-3419 | 2.3 µM | |||
| Norharmane | C. albicans ATCC 10231 | 237 µM | - | [97] |
| Tjipanazole A | C. albicans | - | - | [98] |
| T. mentagrophytes | - | - | ||
| A. flavus | - | - | ||
| Welwitindolinone A isonitrile | A. oryzae | - | - | [99] |
| P. notatum | - | - | ||
| S. cerevisiae | - | - | ||
| Fischerellin A 611.9 µM | U. appendiculatus | 100% | - | [100] |
| Fischerellin A 2.44 µM | Erysiphe graminis | 100% | - | |
| Phytophthora infestans | 80% | - | ||
| Pyricularia oryzae | 80% | - | ||
| Monilinia fructigena | 80% | - | ||
| Pseudocercosporella herpotrichoides | 30% | - | ||
| Kalkipyrone A | S. cerevisiae ABC16-Monster | 14.6 µMIC50 | - | [101] |
| Kalkipyrone B | 13.4 µMIC50 | - | ||
| Yoshinone A | 63.8 µMIC50 | - | ||
| Carbamidocyclophane A | C. albicans | 5.5 µM | - | [102] |
| Carbamidocyclophane B | 1.3 µM | - | ||
| Carbamidocyclophane F | 2.9 µM | - | ||
| Scytophycin A | Saccharomyces pastorianus | 17 mm | - | [103] |
| Neurospora crassa | 25 mm | - | ||
| Candida albicans | 19 mm | - | ||
| Pythium ultimum | >30 mm | - | ||
| R. solani | 23 mm | - | ||
| Sclerotina homoeocarpa | 30 mm | - | ||
| Scytophycin B | S. pastorianus | 20 mm | - | |
| N. crassa | 27 mm | - | ||
| C. albicans | 22 mm | - | ||
| P. ultimum | 30 mm | - | ||
| R. solani | >30 mm | - | ||
| S. homoeocarpa | >30 mm | - | ||
| Scytophycin C | S. pastorianus | 17 mm | - | |
| N. crassa | 30 mm | - | ||
| C. albicans | 22 mm | - | ||
| P. ultimum | 12 mm | - | ||
| R. solani | 30 mm | - | ||
| S. homoeocarpa | 32 mm | - | ||
| Scytophycin D | S. pastorianus | 12 mm | - | |
| N. crassa | 25 mm | - | ||
| C. albicans | 19 mm | - | ||
| P. ultimum | 27 mm | - | ||
| R. solani | 30 mm | - | ||
| S. homoeocarpa | 26 mm | - | ||
| Scytophycin E | S. pastorianus | 23 mm | - | |
| N. crassa | 36 mm | - | ||
| C. albicans | 21 mm | - | ||
| P. ultimum | 40 mm | - | ||
| R. solani | 46 mm | - | ||
| S. homoeocarpa | 35 mm | - | ||
| Tolytoxin | C. albicans A26 T. mentagrophytes A23 | 8 nM | 0.12–1 nM Nystatin | [104] |
| S. cerevisiae Phytophtora nicotianae H729 A. alternata 1715 Colletotrichum eoecodes 1809 | 4 nM | |||
| Bipolaris incurvata 2118 Caloneetria critalarae 1809 | 2 nM | |||
| Sclerotium rolfsii 2133 Thielaviopsis paradoxa 1215 | 1 nM | |||
| A. oryzae Phyllosticta capitalensis | 0.5 nM | |||
| P. notatum R. solani 1165 | 0.25 nM | |||
| Swinholide A | - | - | - | [105] |
| Amantelide A 7.9 µM | D. salina | 0% | 100% Amphotericin B (67.6 µM) | [106] |
| L. thalassiae | 40% | 21% Amphotericin B (67.6 µM) | ||
| Fusarium sp. | 35% | 6% Amphotericin B (67.6 µM) | ||
| Sacrolide A | Penicillium chrysogenum | 3.2 µM | - | [107,108] |
| S. cerevisiae | 25.9 µM | - | ||
| C. albicans | 25.9 µM | - | ||
| 9-epi-sacrolide A | S. cerevisiae | ≦ 25.9 µM | - | |
| P. chrysogenum | ≦ 25.9 µM | - | ||
| 15,16-dihydrosacrolide A | S. cerevisiae | 25.8 µM | - | |
| P. chrysogenum | 25.8 µM | - | ||
| Ambigol A 50 µg | Microbotryum violaceum | 7 mm | 20 mm Miconazole | [109] |
| Eurotium repens | 7 mm | |||
| F. oxysporum | 8 mm | |||
| Mycotypha microspora | 4 mm | |||
| Ambigol C 50 µg | M. violaceum | 5 mm | ||
| 4,4′-dihydroxybiphenyl | C. albicans ATCC 10231 | 171.8 µM | - | [97] |
| Parsiguine | C. krusei ATCC 44507 | 20 µg mL−1 | - | [110] |
| Scytoscalarol | C. albicans | 4 µM | - | [111] |
| Compound | Molecular Weight | Cyanobacterial Strain (Family) | Source | Location | Ref |
|---|---|---|---|---|---|
| Polysaccharide | - | Nostoc Commune (Nostocaceae) | Terrestrial | Nanbu County (SiChuan, China) | [83] |
| - | Phormidium versicolor NCC 466 (Oscillatoriaceae) | Terrestrial | Sfax (Tunisia) | [81] | |
| - | Anabaena sp. BEA 0300B (Aphanizomenonaceae) | Terrestrial | Ajuy (Gran Canaria, Spain) | [82] | |
| 7-Deoxysedoheptulose | 194.18 | Synechococcus elongatus PCC 7942 (Synechococcaceae) | Freshwater | San Francisco Bay (California, U.S) | [80] |
| Compound | Molecular Weight | Cyanobacterial Strain (Family) | Source | Location | Ref |
|---|---|---|---|---|---|
| Mirabilene A isonitrile | 407.59 | Scytonema mirabile BY-8-1 (Scytonemataceae) | Terrestrial | Tantalus (Hawaii, U.S) | [86] |
| Mirabilene B isonitrile | 405.57 | ||||
| Mirabilene C isonitrile | |||||
| Mirabilene D isonitrile | |||||
| Mirabilene E isonitrile | |||||
| Mirabilene F isonitrile | 465.67 | ||||
| Tanikolide | 284.44 | L. majuscula MNT-6 (Oscillatoriaceae) | Marine | Tanikeli Island (Madagascar) | [88] |
| Majusculoic Acid | 315.25 | Environmental Sample | Marine | Sweetings Cay (Bahamas) | [16,85] |
| Aphanothece bullosa (Microcystaceae) | Freshwater | Banaras Hindu University (India) | |||
| Chlorosphaerolactylate A | 341.27 | Sphaerospermopsis sp. LEGE 00249 (Aphanizomenonaceae) | Freshwater | Maranhão Dam Reservoir (Montargil, Portugal) | [84] |
| Chlorosphaerolactylate B–C | 306.83 | Freshwater | |||
| Chlorosphaerolactylate D | 375.72 | Freshwater |
| Compound | Molecular Weight | Cyanobacterial Strain (Family) | Source | Location | Ref |
|---|---|---|---|---|---|
| Hapalindole A | 338.87 | Hapalosiphon fontinalis ATCC 39694 (Hapalosiphonaceae) | Terrestrial | Marshall Islands | [95] |
| Hapalindole B | 370.94 | ||||
| Hapalindole C | 304.43 | ||||
| Hapalindole D | 336.49 | ||||
| Hapalindole E | 338.87 | ||||
| Hapalindole F | 370.94 | ||||
| Hapalindole G | 338.87 | ||||
| Hapalindole H | 304.43 | ||||
| Hapalindole X | 304.43 | Westiellopsis sp. SAG 20.93 (Hapalosiphonaceae) | Terrestrial | Mae Hong Son (Thailand) | [93] |
| Hapalonamide H | 336.43 | ||||
| Hapalindole J | 304.43 | ||||
| Anhydrohapaloxindole A | 352.86 | Fischerella muscicola UTEX LB1829 (Hapalosiphonaceae) | Terrestrial | - | [93] |
| Fischerindole L | 338.87 | ||||
| 12-epi-fischerindole U | 304.43 | Chemical Synthesis | - | - | [94] |
| 12-epi-fischerindole G | 338.87 | ||||
| 13R-Bromo 12-epi-fischerindole U | - | ||||
| Fischambiguine A | 386.53 | F. ambigua UTEX 1903 (Hapalosiphonaceae) | Terrestrial | - | [91] |
| Ambiguine A isonitrile | 406.99 | F. ambigua UTEX 1903 Hapalosiphon hibernicus BZ-3-1 Westiellopsis prolifica EN-3-1 (Hapalosiphonaceae) | Terrestrial | - | [89,216] |
| Ambiguine B isonitrile | 422.99 | ||||
| Ambiguine C isonitrile | 388.55 | Maui Island (Hawaii, U.S) | |||
| Ambiguine D isonitrile | 452.97 | ||||
| Ambiguine E isonitrile | 436.98 | - | |||
| Ambiguine F isonitrile | 454.99 | ||||
| Ambiguine G isonitrile | 402.97 | H. delicatulus IC-13-1 (Hapalosiphonaceae) | Terrestrial | Australia | [213] |
| Ambiguine H isonitrile | 372.55 | Fischerella sp. TAU IL-199-3-1 (Hapalosiphonaceae) | Terrestrial | The Cactus Nursery (Herzliya, Israel) | [90] |
| Ambiguine I isonitrile | 402.54 | ||||
| Ambiguine K isonitrile | 420.98 | F. ambigua UTEX 1903 (Hapalosiphonaceae) | Terrestrial | - | [91,92] |
| Ambiguine L isonitrile | 386.54 | ||||
| Ambiguine M isonitrile | 438.99 | ||||
| Ambiguine N isonitrile | 404.55 | ||||
| Ambiguine O isonitrile | 452.98 | ||||
| Ambiguine P isonitrile | 359.51 | ||||
| Welwitindolinone A isonitrile | 588.98 | Hapalosiphon welwitschii UH IC-52-3 (Hapalosiphonaceae) | Terrestrial | Australia | [99] |
| Tjipanazole A | 471.33 | Tolypothrix tjipanasensis DB-1-1 (Tolypothrichaceae) | Terrestrial | Vero Beach (Florida, U.S) | [98] |
| Carriebowlinol | 197.66 | L. majuscula-Hormoscilla sp. Consortium | Marine | Coral reefs (Carrie Bow Cay, Belize) | [87] |
| Norharmane | 168.2 | N. harveyana 44.85 (Nodulariaceae) | Marine | United Kingdom | [97,217] |
| Synechocystis aquatilis (Microcystaceae) | Freshwater | Saudi Arabia | [218] | ||
| Anabaena cylindrica SAG 1403-2 (Aphanizomenonaceae) | Freshwater | United Kingdom | [97,217] | ||
| Anabaena inaequalis SAG 1403-10 (Aphanizomenonaceae) | Freshwater | Netherlands | |||
| Cylindrospermum Siamensis B 11.82 (Aphanizomenonaceae) | Terrestrial | Thailand | |||
| Chroococcus minutus SAG 41.79 (Chroococcaceae) | Freshwater | Romania | |||
| Nostoc carneum (Nostocaceae) | - | - | |||
| Phormidium foveolarum UTEX 427 (Oscillatoriaceae) | - | - | |||
| Nostoc commune SAG 1453-5 (Nostocaceae) | - | - |
| Compound | Molecular Weight | Cyanobacterial Strain (Family) | Source | Location | Ref |
|---|---|---|---|---|---|
| Kalkipyrone A | 332.44 | cf. Leptolyngbya sp. (Leptolyngbyaceae) | Marine | Fagasa Bay (American Samoa) | [101,235] |
| L. majuscula and Tolypothrix sp. assemblage | Curaçao | ||||
| Kalkipyrone B | 334.45 | cf. Leptolyngbya sp. (Leptolyngbyaceae) | Marine | Fagasa Bay (American Samoa) | |
| Yoshinone A | 364.48 | cf. Schizothrix sp. (Schizotrichaceae) | Marine | Panama | [101] |
| Leptolyngbya sp. (Leptolyngbyaceae) | Okinawa (Japan) | [236] |
| Compound | Molecular Weight | Cyanobacterial Strain | Source | Location | Ref |
|---|---|---|---|---|---|
| Carbamidocyclophane A | 808.66 | Nostoc sp. CAVN10 | Terrestrial | Vietnam | [240] |
| Nostoc sp. UIC 10274 | Freshwater | Des Plaines (Illinois, U.S) | [102] | ||
| Nostoc sp. CAVN02 | Freshwater | Vietnam | [239,242] | ||
| Cylindrospermum stagnale BEA 0605B | Freshwater | Canary Islands (Spain) | [241] | ||
| Carbamidocyclophane B | 774.21 | Nostoc sp. CAVN10 | Terrestrial | Vietnam | [240] |
| Nostoc sp. UIC 10274 | Freshwater | Des Plaines (Illinois, U.S) | [102] | ||
| Nostoc sp. CAVN2 | Freshwater | Vietnam | [242] | ||
| Carbamidocyclophane F | 765.63 | Nostoc sp. UIC 10274 | Freshwater | Des Plaines (Illinois, U.S) | [102] |
| Nostoc sp. CAVN2 | Freshwater | Vietnam | [242] | ||
| Cylindrospermum stagnale BEA 0605B | Freshwater | Canary Islands (Spain) | [241] |
| Compound | Molecular Weight | Cyanobacterial Strain (Family) | Source | Location | Ref |
|---|---|---|---|---|---|
| Scytophycin A | 822.08 | S. pseudohofmanni ATCC 53141 (Scytonemataceae) | Terrestrial | Island of Oahu (Hawaii, U.S) | [103] |
| Scytophycin B | 820.07 | Cylindrospermum muscicola GO-17-1 | Terrestrial | Island of Kauai (Hawaii, U.S) | |
| S. pseudohofmanni ATCC 53141 | Terrestrial | Island of Oahu (Hawaii, U.S) | |||
| Scytophycin C | 806.09 | S. pseudohofmanni ATCC 53141 | Terrestrial | Island of Oahu (Hawaii, U.S) | [103] |
| Scytonema sp. UIC 10036 | Freshwater | Homestead (Florida, U.S) | [252] | ||
| Scytophycin D | 822.09 | S. pseudohofmanni ATCC 53141 | Terrestrial | Island of Oahu (Hawaii, U.S) | [103] |
| Scytophycin E | 822.09 | C. muscicola GO-17-1 | Terrestrial | Island of Kauai (Hawaii, U.S) | [103] |
| S. pseudohofmanni ATCC 53141 | Terrestrial | Island of Oahu (Hawaii, U.S) | [103] | ||
| Tolytoxin | 850.09 | Scytonema ocellatum FF-66-3 | - | South Pasture Pond (Illinois, U.S) | [104] |
| S. ocellatum FF-65-1 | - | Columbia (Missouri, U.S) | |||
| S. ocellatum DD-8-1 | - | University of Guam (Guam) | |||
| S. mirabile BY-8-1 | Terrestrial | Island of Oahu (Hawaii, U.S) | |||
| S. burmanicum DO-4-1 | - | Moon Beach (Okinawa, Japan) | |||
| Scytonema sp. UIC 10036 | Freshwater | Homestead (Florida, U.S) | [252] | ||
| Swinholide A | 1389.89 | Symploca cf. sp. (Phormidiaceae) | Marine | Fiji Islands | [248] |
| Geitlerinema sp. (Coleofasciculaceae) | Nosy Mitsio (Madagascar) | ||||
| Amantelide A | 789.14 | A Member of Oscillatoriales family | Marine | Puntan dos Amates (Guam) | [106] |
| Sacrolide A | 308.41 | A. sacrum (Microcystaceae) | Freshwater | Kyushu District (Japan) | [107,108] |
| 9-epi-sacrolide A | 308.41 | ||||
| 15,16-dihydrosacrolide A | 310.43 |
| Compound | Molecular Weight | Cyanobacterial Strain (Family) | Source | Location | Ref |
|---|---|---|---|---|---|
| 4,4′-dihydroxybiphenyl | 186.21 | Nostoc insulare SAG 54.79 (Nostocaceae) | Terrestrial | - | [97] |
| Ambigol A | 484.97 | Symphyonema bifilamentata sp. nov. 97.28 (Symphyonemataceae) | Terrestrial | Mellingen (Switzerland) | [109,258] |
| Ambigol C | 484.97 | ||||
| Scytoscalarol | 415.66 | Scytonema sp. UTEX 1163 (Scytonemataceae) | Terrestrial | - | [111] |
| Parsiguine | 160.00 | F. ambigua (Hapalosiphonaceae) | Terrestrial | Noushahr (Iran) | [110] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
do Amaral, S.C.; Xavier, L.P.; Vasconcelos, V.; Santos, A.V. Cyanobacteria: A Promising Source of Antifungal Metabolites. Mar. Drugs 2023, 21, 359. https://doi.org/10.3390/md21060359
do Amaral SC, Xavier LP, Vasconcelos V, Santos AV. Cyanobacteria: A Promising Source of Antifungal Metabolites. Marine Drugs. 2023; 21(6):359. https://doi.org/10.3390/md21060359
Chicago/Turabian Styledo Amaral, Samuel Cavalcante, Luciana Pereira Xavier, Vítor Vasconcelos, and Agenor Valadares Santos. 2023. "Cyanobacteria: A Promising Source of Antifungal Metabolites" Marine Drugs 21, no. 6: 359. https://doi.org/10.3390/md21060359