
Impact of Enzymatically Extracted High Molecular Weight Fucoidan on Lipopolysaccharide-Induced Endothelial Activation and Leukocyte Adhesion
 , ,
, , 
Abstract
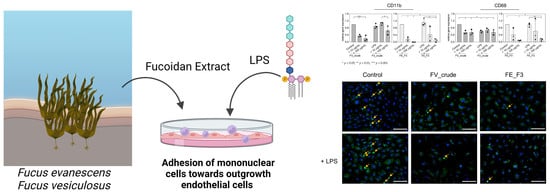
:
1. Introduction
2. Results
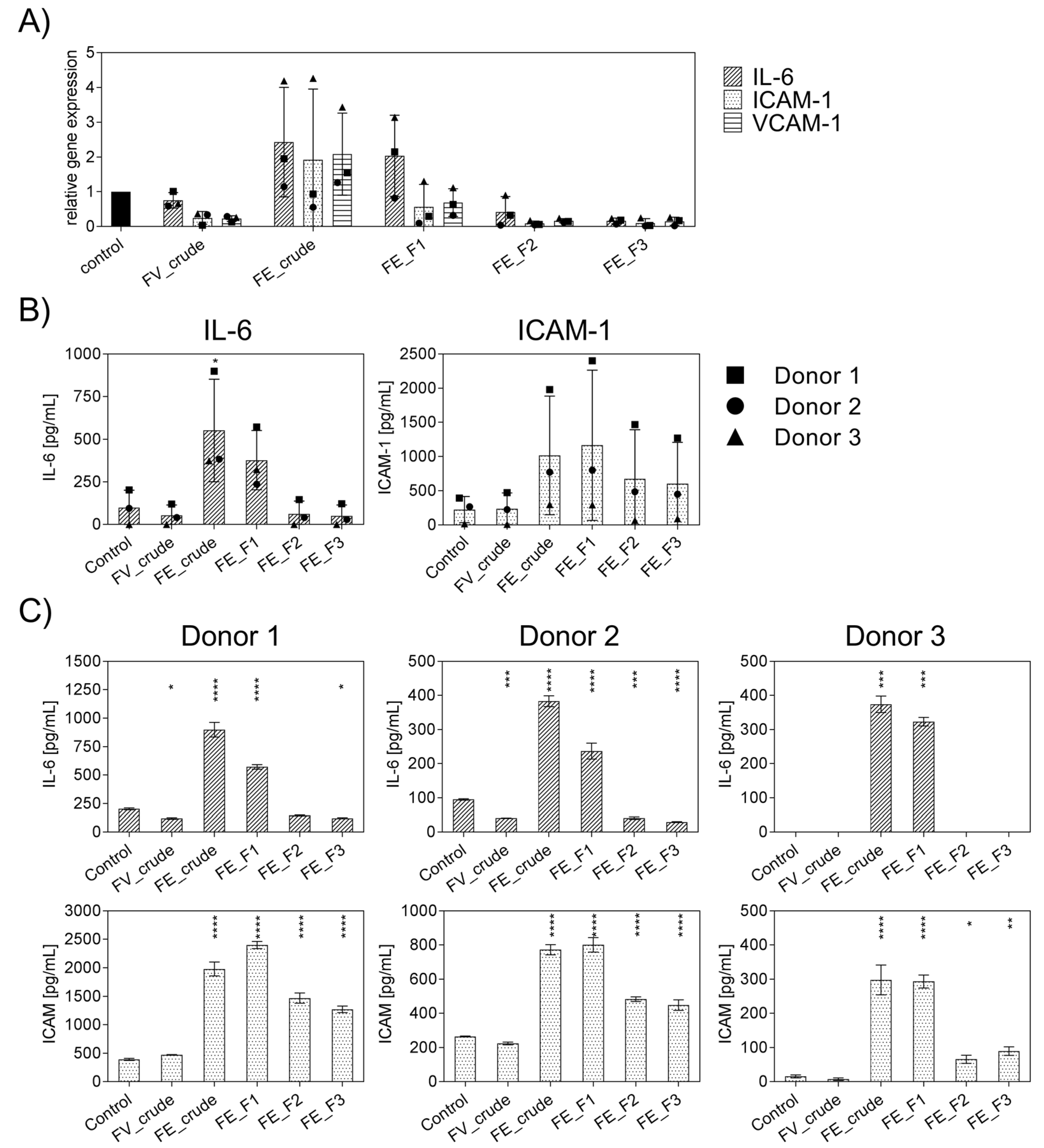
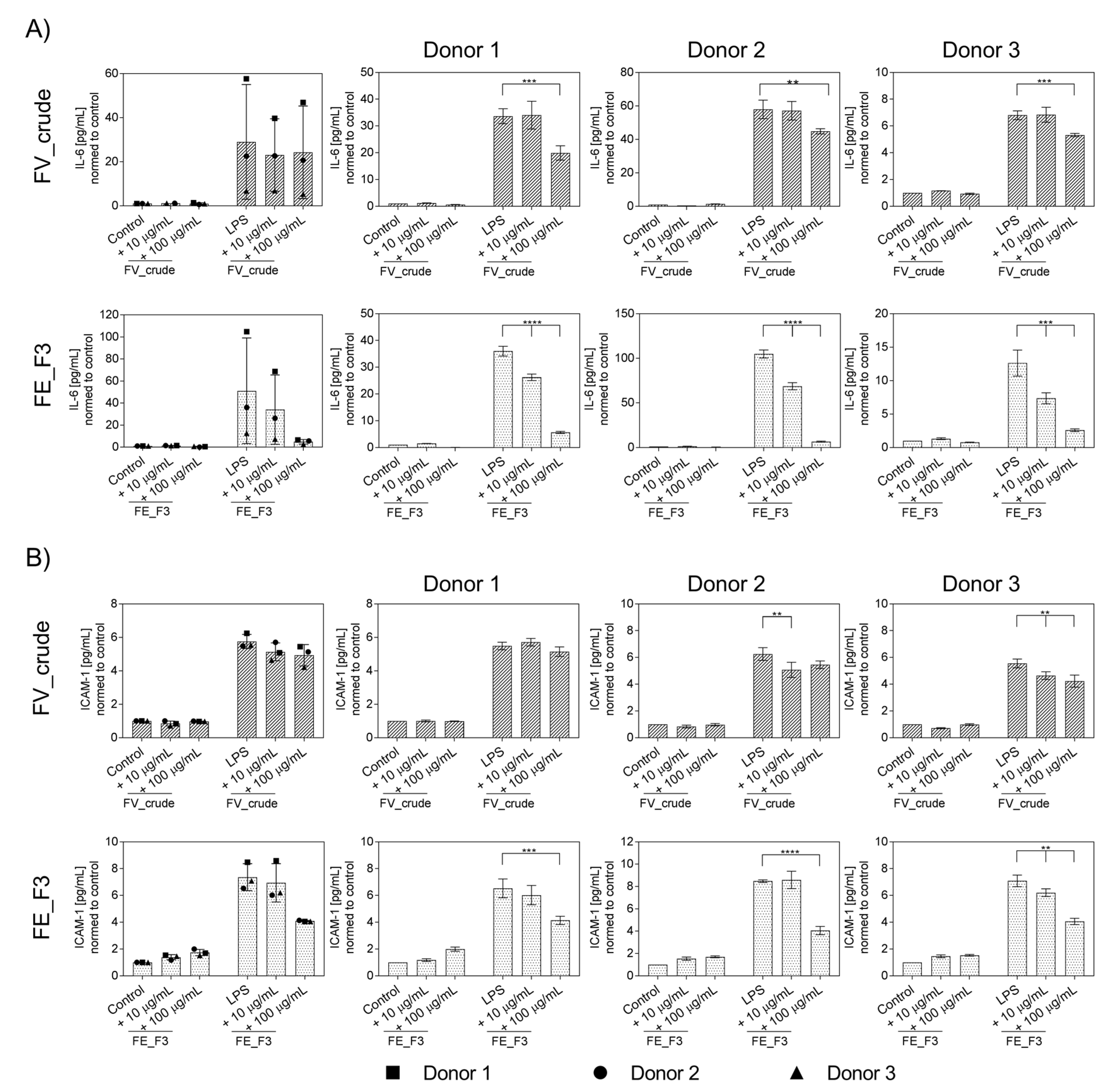
2.1. Influence of Fucoidan Extracts on Inflammatory Mediators in OECs
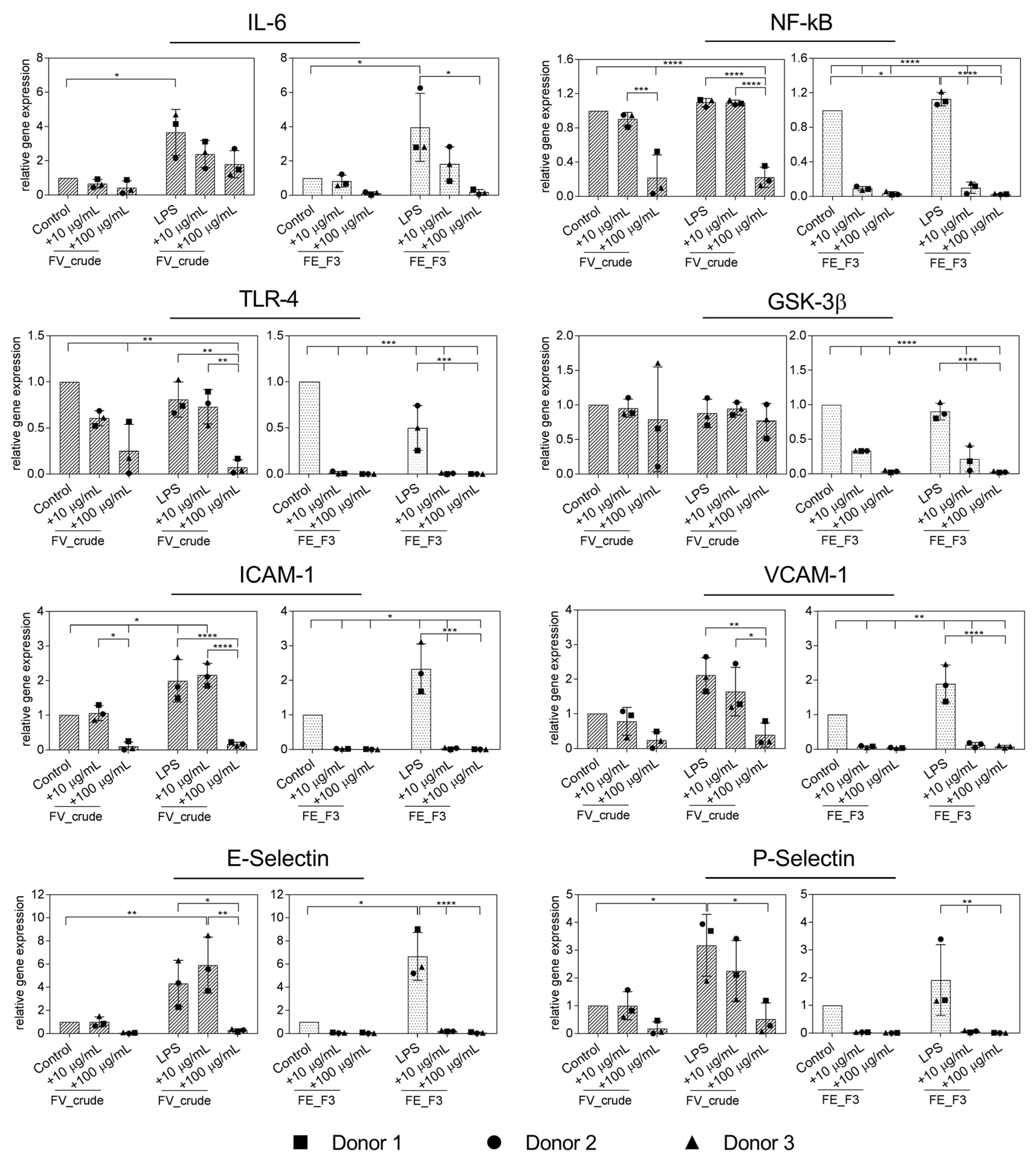
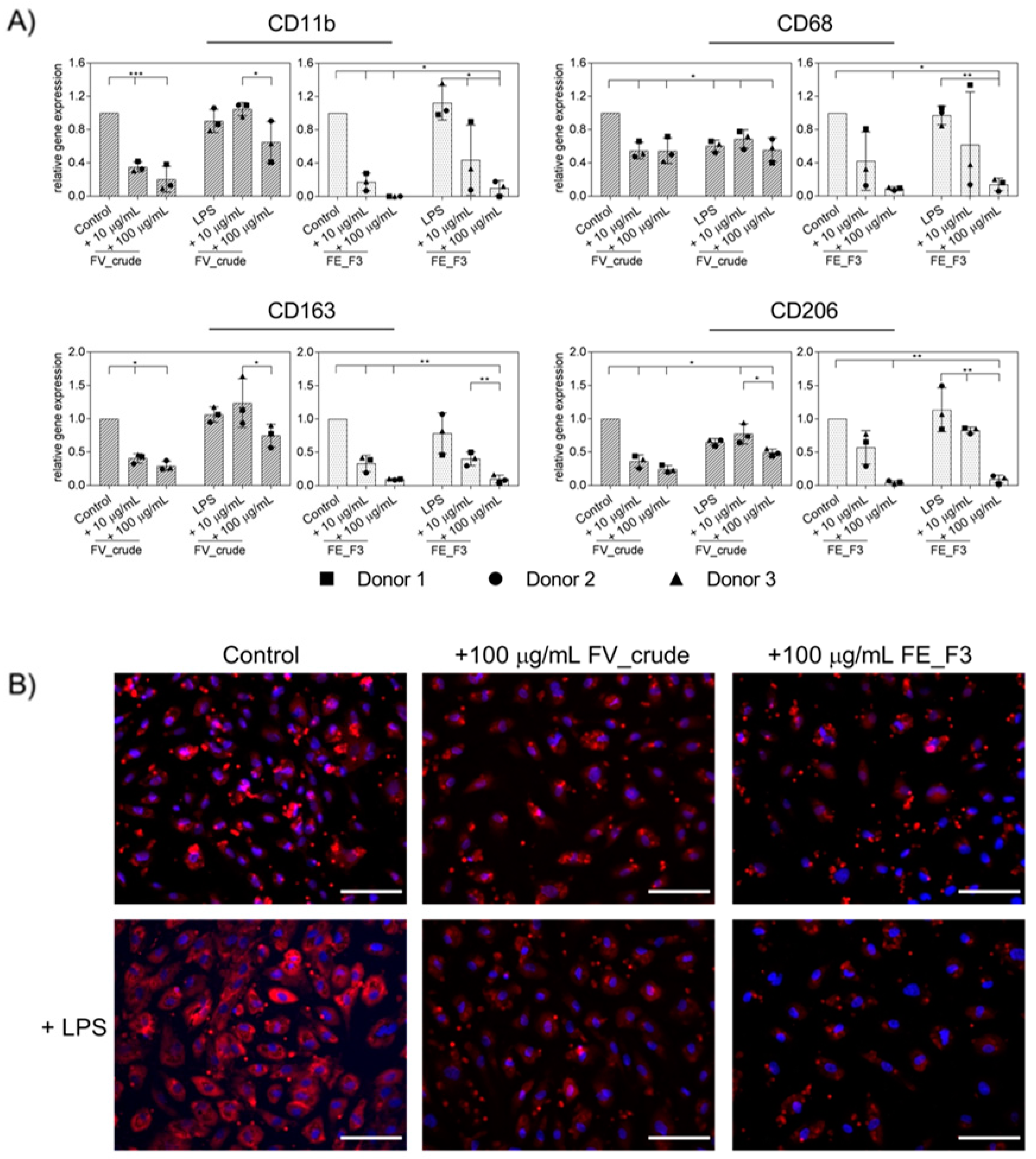
2.2. Effect of Fucoidan Extracts on Rna Expression of Markers for Endothelial Activation in LPS-Stimulated OECs
2.3. Effect of Fucoidan Extracts on Endothelial Activation in LPS-Stimulated OEC
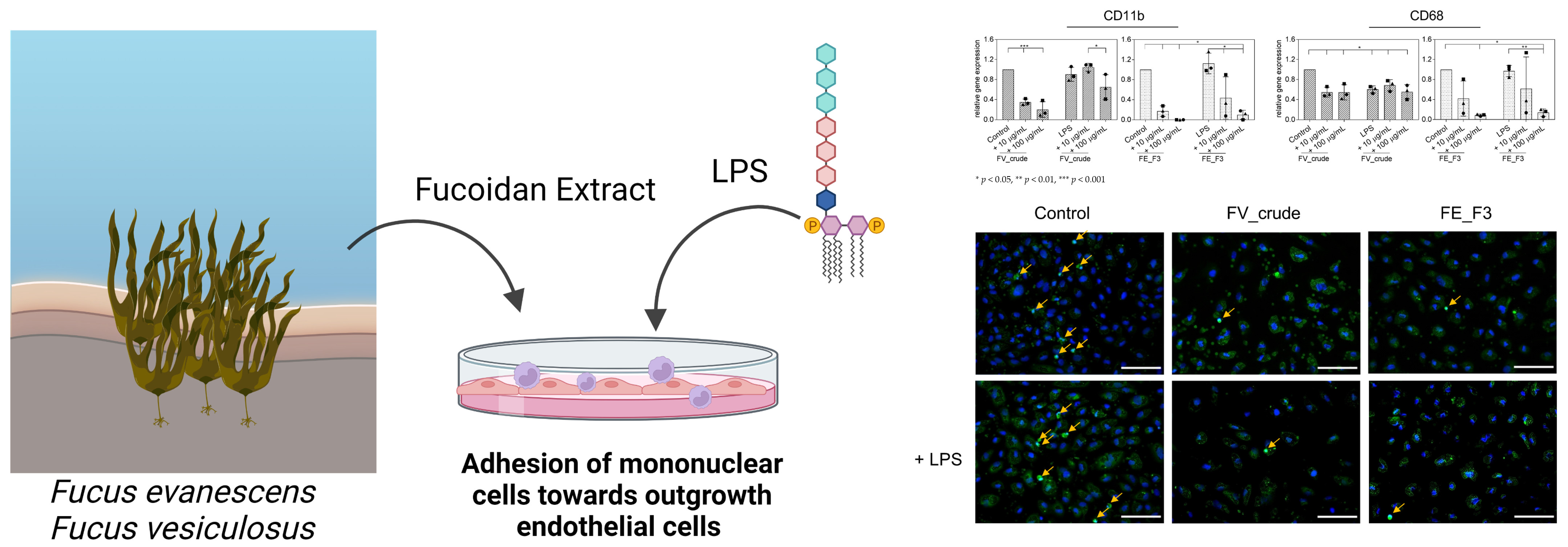
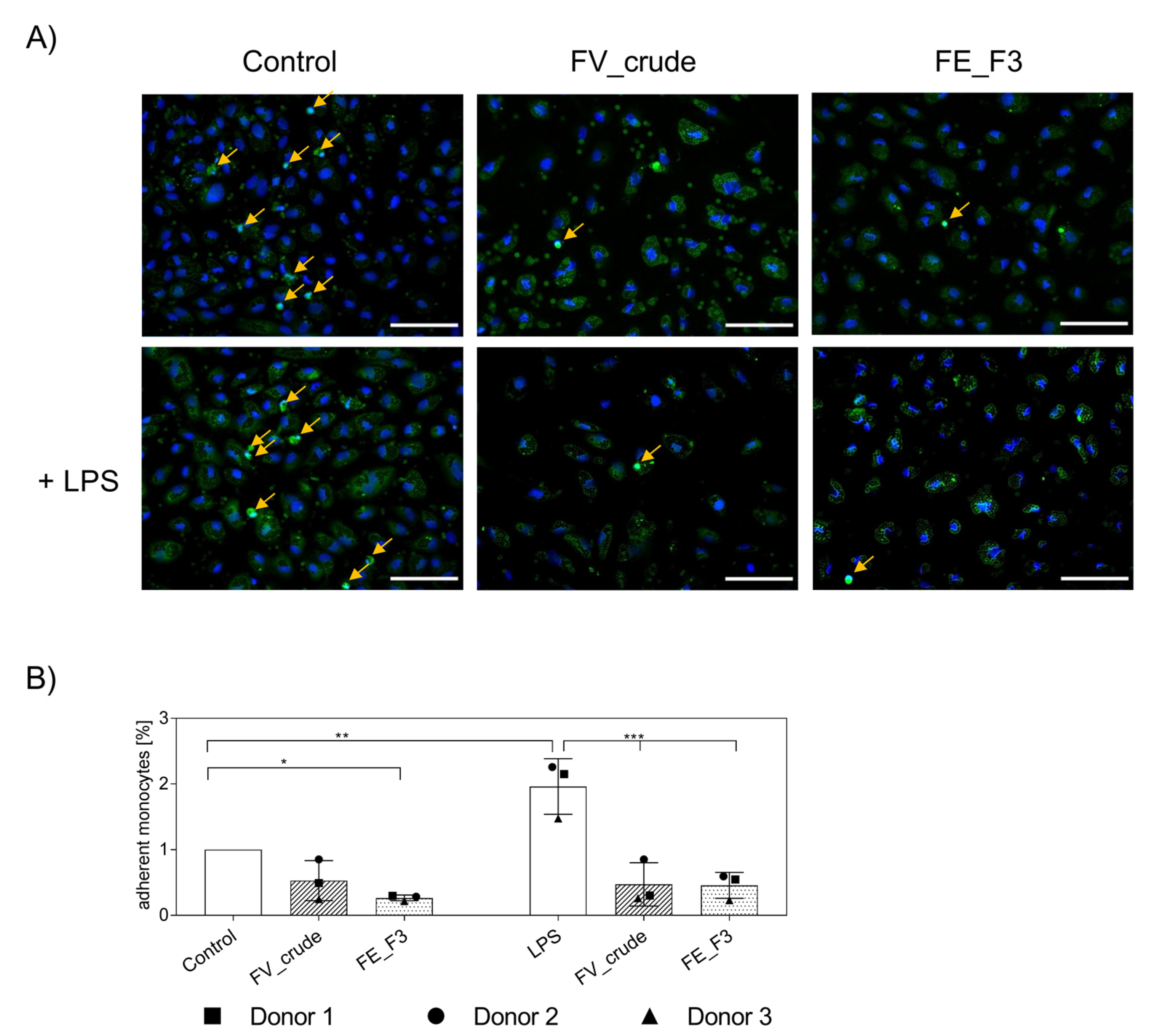
2.4. Impact of Fucoidan Extracts on the Adherence of Monocytes to the Endothelial Cell Layer
2.5. Influence of Fucoidan Extracts on the Adherence of Native Monocytes to the Endothelial Cell Layer
3. Discussion
4. Materials and Methods
4.1. Extraction of Fucoidan from Algae
4.2. Isolation of Endothelial Progenitor Cells and Outgrowth Endothelial Cells (OECs) from the Peripheral Blood
4.3. Magnetic Cell Sorting to Isolate Cd14 Positive Monocytes from Human Peripheral Blood
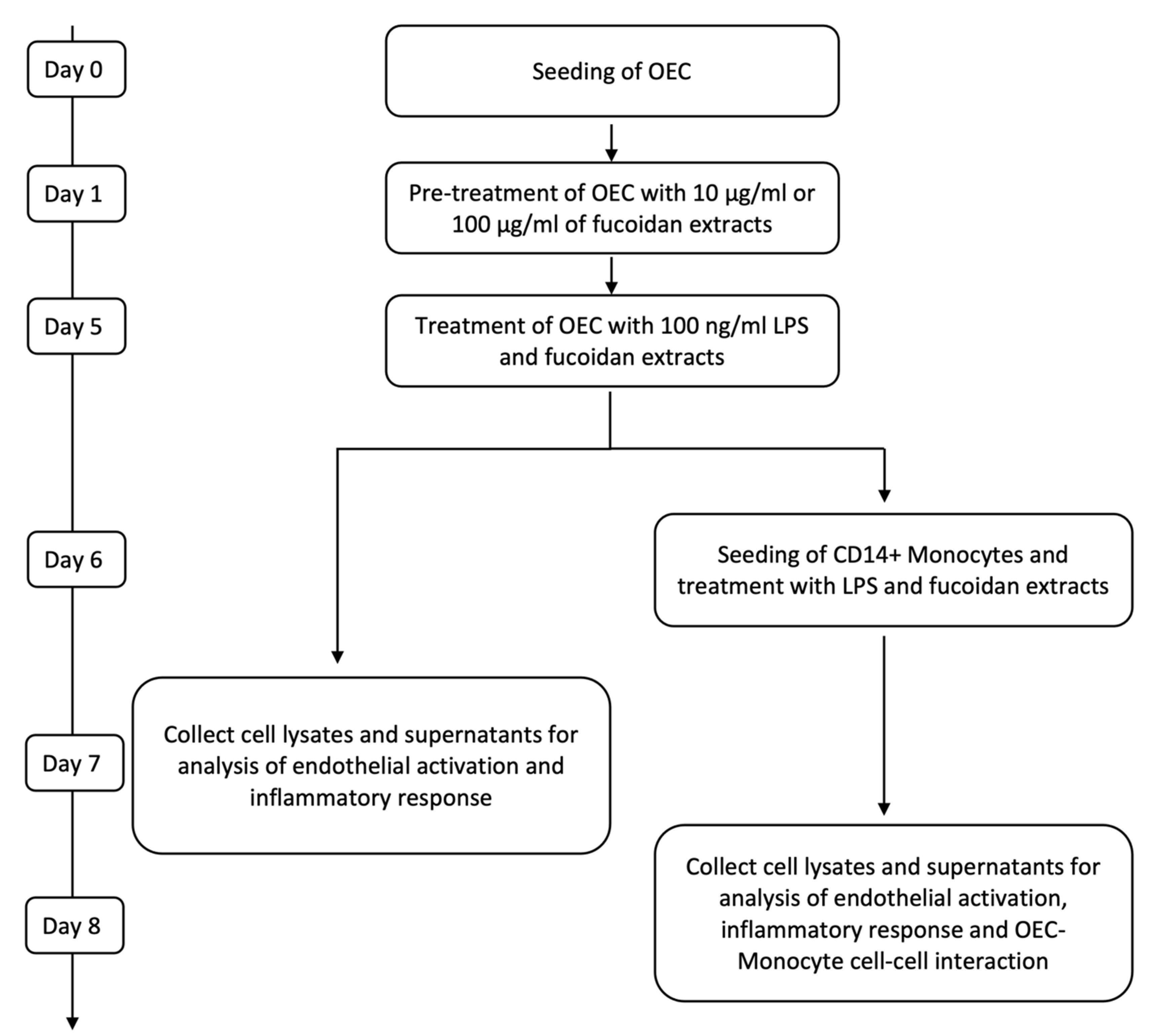
4.4. Fucoidan and LPS Treatment of OEC Mono-Cultures and OECMonocyte Co-Cultures
4.5. Analysis of Endothelial Cell Activation and Inflammatory Response in Mono- and Co-Cultures by Semiquantative Real-Time PCR
4.6. Enzyme-Linked Immunosorbent Assay (ELISA) for Inflammatory Markers
4.7. Immunocytochemistry of Endothelial Cell Activation and Interaction with Mononuclear Cells
4.8. Quantification of Mononuclear Cell Adhesion on Endothelial Cells in Response to Fucoidan
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Trepels, T.; Zeiher, A.M.; Fichtlscherer, S. The Endothelium and Inflammation. Endothelium 2006, 13, 423–429. [Google Scholar] [CrossRef]
- Pober, J.S.; Cotran, R.S. The Role of Endothelial Cells in Inflammation. Transplantation 1990, 50, 537–544. [Google Scholar] [CrossRef]
- Bonetti, P.O.; Lerman, L.O.; Lerman, A. Endothelial Dysfunction: A Marker of Atherosclerotic Risk. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 168–175. [Google Scholar] [CrossRef]
- Peters, K. Molecular Basis of Endothelial Dysfunction in Sepsis. Cardiovasc. Res. 2003, 60, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Boos, C.; Goon, P.; Lip, G. The Endothelium, Inflammation, and Coagulation in Sepsis. Clin. Pharmacol. Ther. 2006, 79, 20–22. [Google Scholar] [CrossRef] [PubMed]
- Joffre, J.; Hellman, J.; Ince, C.; Ait-Oufella, H. Endothelial Responses in Sepsis. Am. J. Respir. Crit. Care Med. 2020, 202, 361–370. [Google Scholar] [CrossRef]
- Pober, J.S.; Sessa, W.C. Evolving Functions of Endothelial Cells in Inflammation. Nat. Rev. Immunol. 2007, 7, 803–815. [Google Scholar] [CrossRef]
- Poltorak, A. Defective LPS Signaling in C3H/HeJ and C57BL/10ScCr Mice: Mutations in Tlr4 Gene. Science 1998, 282, 2085–2088. [Google Scholar] [CrossRef] [PubMed]
- Faure, E.; Equils, O.; Sieling, P.A.; Thomas, L.; Zhang, F.X.; Kirschning, C.J.; Polentarutti, N.; Muzio, M.; Arditi, M. Bacterial Lipopolysaccharide Activates NF-ΚB through Toll-like Receptor 4 (TLR-4) in Cultured Human Dermal Endothelial Cells. J. Biol. Chem. 2000, 275, 11058–11063. [Google Scholar] [CrossRef]
- Takeda, K.; Akira, S. TLR Signaling Pathways. Semin. Immunol. 2004, 16, 3–9. [Google Scholar] [CrossRef]
- Martin, M.; Rehani, K.; Jope, R.S.; Michalek, S.M. Toll-like Receptor–Mediated Cytokine Production Is Differentially Regulated by Glycogen Synthase Kinase 3. Nat. Immunol. 2005, 6, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Rehani, K.; Wang, H.; Garcia, C.A.; Kinane, D.F.; Martin, M. Toll-Like Receptor-Mediated Production of IL-1Ra Is Negatively Regulated by GSK3 via the MAPK ERK1/2. J. Immunol. 2009, 182, 547–553. [Google Scholar] [CrossRef]
- Bierhaus, A.; Chen, J.; Liliensiek, B.; Nawroth, P.P. LPS and Cytokine-Activated Endothelium. Semin. Thromb. Hemost. 2000, 26, 571–588. [Google Scholar] [CrossRef] [PubMed]
- Maruo, N.; Morita, I.; Shirao, M.; Murota, S. IL-6 Increases Endothelial Permeability in Vitro. Endocrinology 1992, 131, 710–714. [Google Scholar] [CrossRef] [PubMed]
- Romano, M.; Sironi, M.; Toniatti, C.; Polentarutti, N.; Fruscella, P.; Ghezzi, P.; Faggioni, R.; Luini, W.; van Hinsbergh, V.; Sozzani, S.; et al. Role of IL-6 and Its Soluble Receptor in Induction of Chemokines and Leukocyte Recruitment. Immunity 1997, 6, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Nishino, T.; Nagumo, T. Sugar constituents and blood-anticoagulant activities of fucose-containing sulfated polysaccharides in nine brown seaweed species. J. Agric. Chem. Soc. Jpn. 1987, 61, 361–363. [Google Scholar] [CrossRef]
- Matsubara, K.; Xue, C.; Zhao, X.; Mori, M.; Sugawara, T.; Hirata, T. Effects of Middle Molecular Weight Fucoidans on in Vitro and Ex Vivo Angiogenesis of Endothelial Cells. Int. J. Mol. Med. 2005, 15, 695–699. [Google Scholar] [CrossRef]
- Ohmes, J.; Xiao, Y.; Wang, F.; Mikkelsen, M.D.; Nguyen, T.T.; Schmidt, H.; Seekamp, A.; Meyer, A.S.; Fuchs, S. Effect of Enzymatically Extracted Fucoidans on Angiogenesis and Osteogenesis in Primary Cell Culture Systems Mimicking Bone Tissue Environment. Mar. Drugs 2020, 18, 481. [Google Scholar] [CrossRef]
- Wang, F.; Xiao, Y.; Neupane, S.; Ptak, S.H.; Römer, R.; Xiong, J.; Ohmes, J.; Seekamp, A.; Fretté, X.; Alban, S.; et al. Influence of Fucoidan Extracts from Different Fucus Species on Adult Stem Cells and Molecular Mediators in In Vitro Models for Bone Formation and Vascularization. Mar. Drugs 2021, 19, 194. [Google Scholar] [CrossRef]
- Park, H.Y.; Han, M.H.; Park, C.; Jin, C.-Y.; Kim, G.-Y.; Choi, I.-W.; Kim, N.D.; Nam, T.-J.; Kwon, T.K.; Choi, Y.H. Anti-Inflammatory Effects of Fucoidan through Inhibition of NF-ΚB, MAPK and Akt Activation in Lipopolysaccharide-Induced BV2 Microglia Cells. Food Chem. Toxicol. 2011, 49, 1745–1752. [Google Scholar] [CrossRef]
- Lee, S.-H.; Ko, C.-I.; Jee, Y.; Jeong, Y.; Kim, M.; Kim, J.-S.; Jeon, Y.-J. Anti-Inflammatory Effect of Fucoidan Extracted from Ecklonia Cava in Zebrafish Model. Carbohydr. Polym. 2013, 92, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Obluchinskaya, E.D.; Pozharitskaya, O.N.; Shikov, A.N. In Vitro Anti-Inflammatory Activities of Fucoidans from Five Species of Brown Seaweeds. Mar. Drugs 2022, 20, 606. [Google Scholar] [CrossRef] [PubMed]
- Ale, M.T.; Mikkelsen, J.D.; Meyer, A.S. Important Determinants for Fucoidan Bioactivity: A Critical Review of Structure-Function Relations and Extraction Methods for Fucose-Containing Sulfated Polysaccharides from Brown Seaweeds. Mar. Drugs 2011, 9, 2106–2130. [Google Scholar] [CrossRef] [PubMed]
- Alboofetileh, M.; Rezaei, M.; Tabarsa, M.; Rittà, M.; Donalisio, M.; Mariatti, F.; You, S.; Lembo, D.; Cravotto, G. Effect of Different Non-Conventional Extraction Methods on the Antibacterial and Antiviral Activity of Fucoidans Extracted from Nizamuddinia Zanardinii. Int. J. Biol. Macromol. 2019, 124, 131–137. [Google Scholar] [CrossRef]
- Fletcher, H.R.; Biller, P.; Ross, A.B.; Adams, J.M.M. The Seasonal Variation of Fucoidan within Three Species of Brown Macroalgae. Algal Res. 2017, 22, 79–86. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Mikkelsen, M.D.; Tran, V.H.N.; Trang, V.T.D.; Rhein-Knudsen, N.; Holck, J.; Rasin, A.B.; Cao, H.T.T.; Van, T.T.T.; Meyer, A.S. Enzyme-Assisted Fucoidan Extraction from Brown Macroalgae Fucus Distichus Subsp. Evanescens and Saccharina Latissima. Mar. Drugs 2020, 18, 296. [Google Scholar] [CrossRef]
- Kylin, H. Biochemistry of Sea Algae. HZ Physiol. Chem. 1913, 83, 3. [Google Scholar]
- Ale, M.T.; Meyer, A.S. Fucoidans from Brown Seaweeds: An Update on Structures, Extraction Techniques and Use of Enzymes as Tools for Structural Elucidation. RSC Adv. 2013, 3, 8131–8141. [Google Scholar] [CrossRef]
- Ehrig, K.; Alban, S. Sulfated Galactofucan from the Brown Alga Saccharina Latissima—Variability of Yield, Structural Composition and Bioactivity. Mar. Drugs 2014, 13, 76–101. [Google Scholar] [CrossRef]
- Zayed, A.; Ulber, R. Fucoidans: Downstream Processes and Recent Applications. Mar. Drugs 2020, 18, 170. [Google Scholar] [CrossRef]
- Fiedler, U.; Augustin, H.G. Angiopoietins: A Link between Angiogenesis and Inflammation. Trends Immunol. 2006, 27, 552–558. [Google Scholar] [CrossRef]
- Wei, X.; Cai, L.; Liu, H.; Tu, H.; Xu, X.; Zhou, F.; Zhang, L. Chain Conformation and Biological Activities of Hyperbranched Fucoidan Derived from Brown Algae and Its Desulfated Derivative. Carbohydr. Polym. 2019, 208, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Ohmes, J.; Mikkelsen, M.D.; Nguyen, T.T.; Tran, V.H.N.; Meier, S.; Nielsen, M.S.; Ding, M.; Seekamp, A.; Meyer, A.S.; Fuchs, S. Depolymerization of Fucoidan with Endo-Fucoidanase Changes Bioactivity in Processes Relevant for Bone Regeneration. Carbohydr. Polym. 2022, 286, 119286. [Google Scholar] [CrossRef] [PubMed]
- Apostolova, E.; Lukova, P.; Baldzhieva, A.; Katsarov, P.; Nikolova, M.; Iliev, I.; Peychev, L.; Trica, B.; Oancea, F.; Delattre, C.; et al. Immunomodulatory and Anti-Inflammatory Effects of Fucoidan: A Review. Polymers 2020, 12, 2338. [Google Scholar] [CrossRef]
- Matsumoto, S.; Nagaoka, M.; Hara, T.; Kimura-Takagi, I.; Mistuyama, K.; Ueyama, S. Fucoidan Derived from Cladosiphon Okamuranus Tokida Ameliorates Murine Chronic Colitis through the down-Regulation of Interleukin-6 Production on Colonic Epithelial Cells. Clin. Exp. Immunol. 2004, 136, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Jayawardena, T.U.; Sanjeewa, K.K.A.; Nagahawatta, D.P.; Lee, H.-G.; Lu, Y.-A.; Vaas, A.P.J.P.; Abeytunga, D.T.U.; Nanayakkara, C.M.; Lee, D.-S.; Jeon, Y.-J. Anti-Inflammatory Effects of Sulfated Polysaccharide from Sargassum Swartzii in Macrophages via Blocking TLR/NF-Κb Signal Transduction. Mar. Drugs 2020, 18, 601. [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Obluchinskaya, E.D.; Shikov, A.N. Mechanisms of Bioactivities of Fucoidan from the Brown Seaweed Fucus Vesiculosus L. of the Barents Sea. Mar. Drugs 2020, 18, 275. [Google Scholar] [CrossRef]
- Jin, J.-O.; Yu, Q. Fucoidan Delays Apoptosis and Induces Pro-Inflammatory Cytokine Production in Human Neutrophils. Int. J. Biol. Macromol. 2015, 73, 65–71. [Google Scholar] [CrossRef]
- Kim, K.-J.; Yoon, K.-Y.; Lee, B.-Y. Low Molecular Weight Fucoidan from the Sporophyll of Undaria Pinnatifida Suppresses Inflammation by Promoting the Inhibition of Mitogen-Activated Protein Kinases and Oxidative Stress in RAW264.7 Cells. Fitoterapia 2012, 83, 1628–1635. [Google Scholar] [CrossRef]
- Morya, V.K.; Kim, J.; Kim, E.-K. Algal Fucoidan: Structural and Size-Dependent Bioactivities and Their Perspectives. Appl. Microbiol. Biotechnol. 2012, 93, 71–82. [Google Scholar] [CrossRef]
- Nomura, F.; Akashi, S.; Sakao, Y.; Sato, S.; Kawai, T.; Matsumoto, M.; Nakanishi, K.; Kimoto, M.; Miyake, K.; Takeda, K.; et al. Cutting Edge: Endotoxin Tolerance in Mouse Peritoneal Macrophages Correlates with Down-Regulation of Surface Toll-Like Receptor 4 Expression. J. Immunol. 2000, 164, 3476–3479. [Google Scholar] [CrossRef]
- Asanka Sanjeewa, K.K.; Jayawardena, T.U.; Kim, H.-S.; Kim, S.-Y.; Shanura Fernando, I.P.; Wang, L.; Abetunga, D.T.U.; Kim, W.-S.; Lee, D.-S.; Jeon, Y.-J. Fucoidan Isolated from Padina Commersonii Inhibit LPS-Induced Inflammation in Macrophages Blocking TLR/NF-ΚB Signal Pathway. Carbohydr. Polym. 2019, 224, 115195. [Google Scholar] [CrossRef]
- Pei, Z.; Pang, H.; Qian, L.; Yang, S.; Wang, T.; Zhang, W.; Wu, X.; Dallas, S.; Wilson, B.; Reece, J.M.; et al. MAC1 Mediates LPS-Induced Production of Superoxide by Microglia: The Role of Pattern Recognition Receptors in Dopaminergic Neurotoxicity. Glia 2007, 55, 1362–1373. [Google Scholar] [CrossRef] [PubMed]
- Kourtzelis, I.; Mitroulis, I.; von Renesse, J.; Hajishengallis, G.; Chavakis, T. From Leukocyte Recruitment to Resolution of Inflammation: The Cardinal Role of Integrins. J. Leukoc. Biol. 2017, 102, 677–683. [Google Scholar] [CrossRef]
- Zhang, X.W.; Liu, Q.; Thorlacius, H. Inhibition of Selectin Function and Leukocyte Rolling Protects Against Dextran Sodium Sulfate-Induced Murine Colitis. Scand. J. Gastroenterol. 2001, 36, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Bachelet, L.; Bertholon, I.; Lavigne, D.; Vassy, R.; Jandrot-Perrus, M.; Chaubet, F.; Letourneur, D. Affinity of Low Molecular Weight Fucoidan for P-Selectin Triggers Its Binding to Activated Human Platelets. Biochim. Biophys. Acta BBA-Gen. Subj. 2009, 1790, 141–146. [Google Scholar] [CrossRef]
- Rouzet, F.; Bachelet-Violette, L.; Alsac, J.-M.; Suzuki, M.; Meulemans, A.; Louedec, L.; Petiet, A.; Jandrot-Perrus, M.; Chaubet, F.; Michel, J.-B.; et al. Radiolabeled Fucoidan as a P-Selectin Targeting Agent for In Vivo Imaging of Platelet-Rich Thrombus and Endothelial Activation. J. Nucl. Med. 2011, 52, 1433–1440. [Google Scholar] [CrossRef]
- Zhou, M.; Ding, Y.; Cai, L.; Wang, Y.; Lin, C.; Shi, Z. Low Molecular Weight Fucoidan Attenuates Experimental Abdominal Aortic Aneurysm through Interfering the Leukocyte-Endothelial Cells Interaction. Mol. Med. Rep. 2018, 17, 7089–7096. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, S.; Hermanns, M.I.; Kirkpatrick, C.J. Retention of a Differentiated Endothelial Phenotype by Outgrowth Endothelial Cells Isolated from Human Peripheral Blood and Expanded in Long-Term Cultures. Cell Tissue Res. 2006, 326, 79–92. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Catalog Number |
|---|---|
| Interleukin-6 (IL-6) | QT00083720 |
| Nuclear Factor NF-kappa-B p105 subunit (NF-kB) | QT00063791 |
| Toll-like receptor 4 (TLR4) | QT01670123 |
| Glycogen synthase kinase 3 (GSK3β) | QT00057134 |
| Intercellular Adhesion Molecule 1 (ICAM-1) | QT00074900 |
| Vascular Cell Adhesion Protein 1 (VCAM-1) | QT00018347 |
| E-Selectin | QT00015358 |
| P-Selectin | QT00012516 |
| Integrin α 1 (CD11b) | QT00031500 |
| Macrosialin (CD68) | QT00037184 |
| CD163 | QT00074641 |
| Mannose receptor C type 1 (CD206) | QT00012810 |
| 60S Ribosomal Protein L13a (RPL13A) | QT00089915 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirsten, N.; Ohmes, J.; Mikkelsen, M.D.; Nguyen, T.T.; Blümel, M.; Wang, F.; Tasdemir, D.; Seekamp, A.; Meyer, A.S.; Fuchs, S. Impact of Enzymatically Extracted High Molecular Weight Fucoidan on Lipopolysaccharide-Induced Endothelial Activation and Leukocyte Adhesion. Mar. Drugs 2023, 21, 339. https://doi.org/10.3390/md21060339
Kirsten N, Ohmes J, Mikkelsen MD, Nguyen TT, Blümel M, Wang F, Tasdemir D, Seekamp A, Meyer AS, Fuchs S. Impact of Enzymatically Extracted High Molecular Weight Fucoidan on Lipopolysaccharide-Induced Endothelial Activation and Leukocyte Adhesion. Marine Drugs. 2023; 21(6):339. https://doi.org/10.3390/md21060339
Chicago/Turabian StyleKirsten, Nora, Julia Ohmes, Maria Dalgaard Mikkelsen, Thuan Thi Nguyen, Martina Blümel, Fanlu Wang, Deniz Tasdemir, Andreas Seekamp, Anne S. Meyer, and Sabine Fuchs. 2023. "Impact of Enzymatically Extracted High Molecular Weight Fucoidan on Lipopolysaccharide-Induced Endothelial Activation and Leukocyte Adhesion" Marine Drugs 21, no. 6: 339. https://doi.org/10.3390/md21060339