

Biochemical Properties of a Cold-Active Chitinase from Marine Trichoderma gamsii R1 and Its Application to Preparation of Chitin Oligosaccharides
Abstract
:1. Introduction
2. Results and Discussion
2.1. Purification and Identification of Chitinase from T. gamsii R1
2.2. Substrate Specificity and Kinetic Parameters of ChiTg
2.3. Effects of Temperature and pH on Activity and Stability of ChiTg
2.4. Effects of Different Metal Ions on Stability of ChiTg
2.5. Hydrolytic Pattern of ChiTg
2.6. Preparation of COSs from Colloidal Chitin
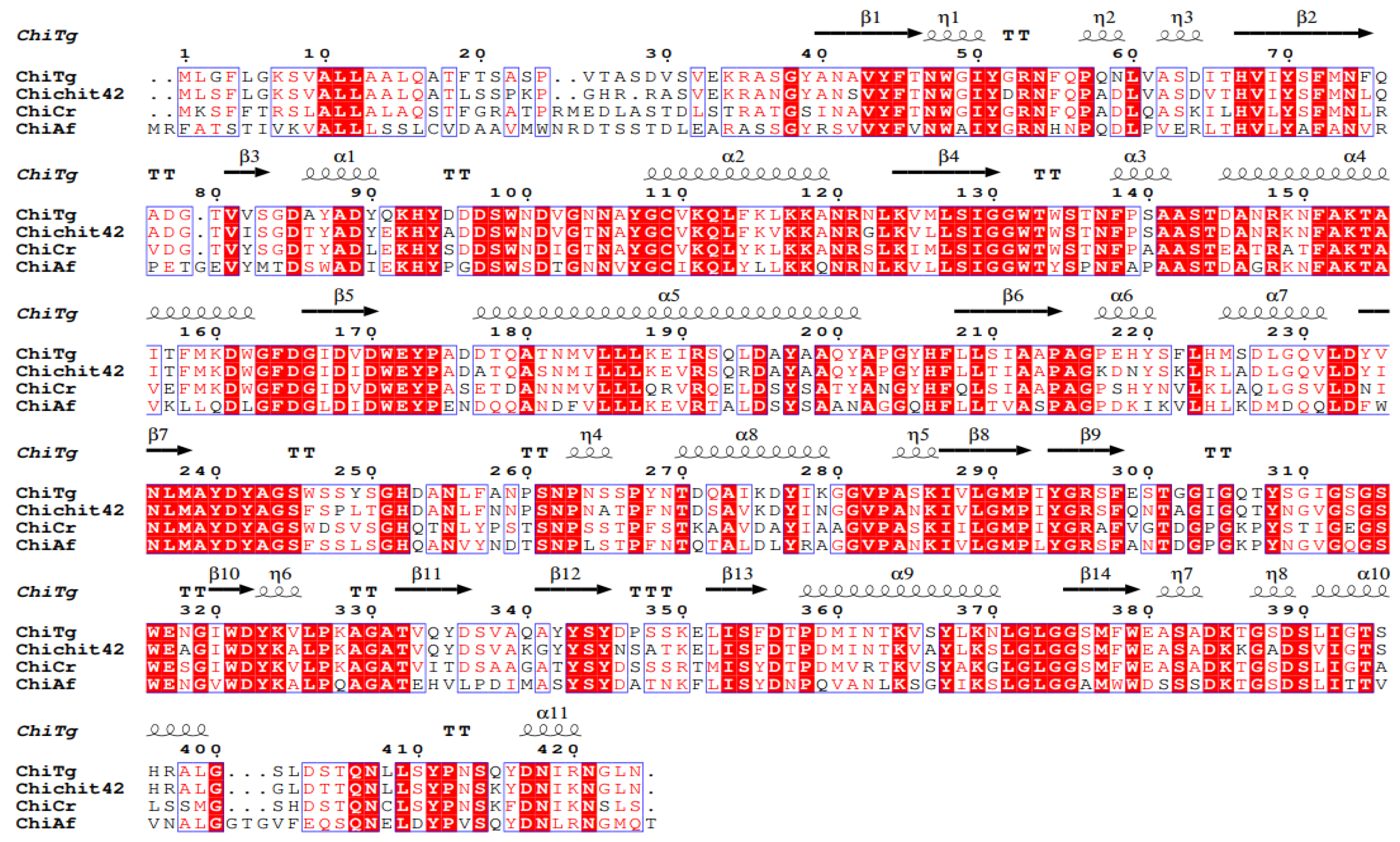
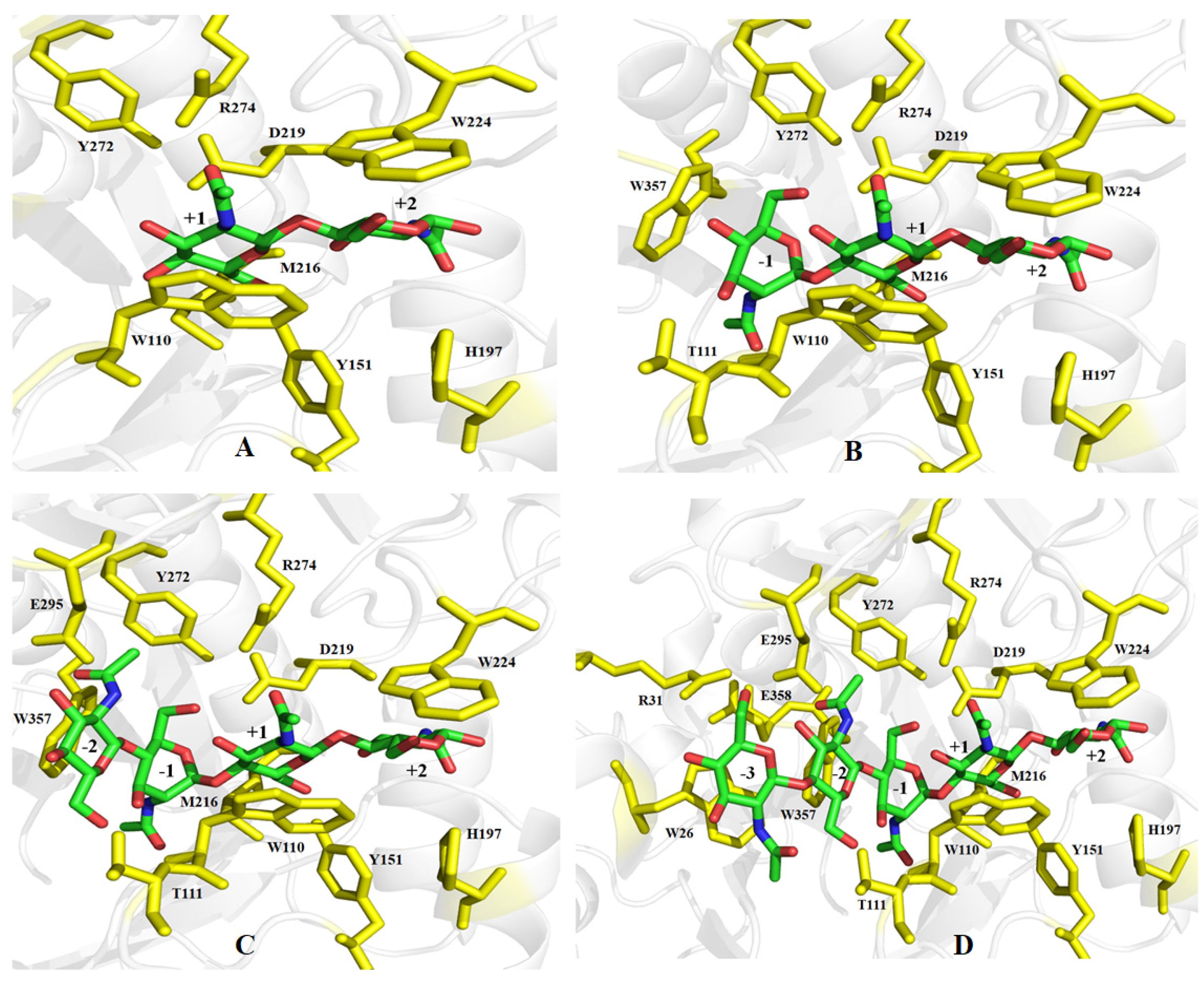
2.7. Gene Cloning and Bioinformatics Analysis of ChiTg
3. Materials and Methods
3.1. Strains and Materials
3.2. Production of Chitinase from T. gamsii R1 in Shake Flasks
3.3. Enzyme Activity Assay
3.4. Enzyme Purification and Peptide Sequence Analysis
3.5. Kinetic Parameters of Purified ChiTg
3.6. Effects of pH and Temperature on Activity and Stability of ChiTg
3.7. Effects of Different Metal Ions on Stability of ChiTg
3.8. Hydrolysis Properties of ChiTg
3.9. Substrate Specificity of ChiTg
3.10. Preparation of COSs from Colloidal Chitin
3.11. Gene Cloning and Bioinformatics Analysis of ChiTg
3.12. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Amiri, H.; Aghbashlo, M.; Sharma, M.; Gaffey, J.; Manning, L.; Moosavi Basri, S.M.; Kennedy, J.F.; Gupta, V.K.; Tabatabaei, M. Chitin and chitosan derived from crustacean waste valorization streams can support food systems and the UN Sustainable Development Goals. Nat. Food 2022, 3, 822–828. [Google Scholar] [CrossRef]
- Arnold, N.D.; Bruck, W.M.; Garbe, D.; Bruck, T.B. Enzymatic Modification of Native Chitin and Conversion to Specialty Chemical Products. Mar. Drugs 2020, 18, 93. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, C.; Auza, L.G.; Koberidze, D.; Rasche, S.; Fischer, R.; Bortesi, L. Conversion of Chitin to Defined Chitosan Oligomers: Current Status and Future Prospects. Mar. Drugs 2019, 17, 452. [Google Scholar] [CrossRef]
- Giraldo, J.D.; Garrido-Miranda, K.A.; Schoebitz, M. Chitin and its derivatives: Functional biopolymers for developing bioproducts for sustainable agriculture—A reality? Carbohydr. Polym. 2023, 299, 120196. [Google Scholar] [CrossRef]
- Li, K.; Xing, R.; Liu, S.; Li, P. Chitin and Chitosan Fragments Responsible for Plant Elicitor and Growth Stimulator. J. Agric. Food Chem. 2020, 68, 12203–12211. [Google Scholar] [CrossRef]
- Tabassum, N.; Ahmed, S.; Ali, M.A. Chitooligosaccharides and their structural-functional effect on hydrogels: A review. Carbohydr. Polym. 2021, 261, 117882. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Zhou, S.; Tan, Y.; Feng, J.; Bai, Y.; He, J.; Cao, H.; Che, Q.; Guo, J.; Su, Z. Biodegradation and Prospect of Polysaccharide from Crustaceans. Mar. Drugs 2022, 20, 310. [Google Scholar] [CrossRef] [PubMed]
- Therien, J.P.D.; Hammerer, F.; Friscic, T.; Auclair, K. Mechanoenzymatic Breakdown of Chitinous Material to N-Acetylglucosamine: The Benefits of a Solventless Environment. ChemSusChem 2019, 12, 3481–3490. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Sun, J.; Secundo, F.; Gao, X.; Xue, C.; Mao, X. Cloning, characterization and substrate degradation mode of a novel chitinase from Streptomyces albolongus ATCC 27414. Food Chem. 2018, 261, 329–336. [Google Scholar] [CrossRef]
- Mukherjee, S.; Behera, P.K.; Madhuprakash, J. Efficient conversion of crystalline chitin to N-acetylglucosamine and N,N′-diacetylchitobiose by the enzyme cocktail produced by Paenibacillus sp. LS1. Carbohydr. Polym. 2020, 250, 116889. [Google Scholar] [CrossRef]
- Tamadoni Jahromi, S.; Barzkar, N. Marine bacterial chitinase as sources of energy, eco-friendly agent, and industrial biocatalyst. Int. J. Biol. Macromol. 2018, 120, 2147–2154. [Google Scholar] [CrossRef]
- Doan, C.T.; Tran, T.N.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.L. Production of a Thermostable Chitosanase from Shrimp Heads via Paenibacillus mucilaginosus TKU032 Conversion and its Application in the Preparation of Bioactive Chitosan Oligosaccharides. Mar. Drugs 2019, 17, 217. [Google Scholar] [CrossRef]
- Mathew, G.M.; Madhavan, A.; Arun, K.B.; Sindhu, R.; Binod, P.; Singhania, R.R.; Sukumaran, R.K.; Pandey, A. Thermophilic Chitinases: Structural, Functional and Engineering Attributes for Industrial Applications. Appl. Biochem. Biotechnol. 2021, 193, 142–164. [Google Scholar] [CrossRef] [PubMed]
- Bruno, S.; Coppola, D.; di Prisco, G.; Giordano, D.; Verde, C. Enzymes from Marine Polar Regions and Their Biotechnological Applications. Mar. Drugs 2019, 17, 544. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Guo, Y.; Jin, Y.; Ma, M. Bioconversion of chitin waste using a cold-adapted chitinase to produce chitin oligosaccharides. Lwt 2020, 133, 109863. [Google Scholar] [CrossRef]
- Yang, G.; Mozzicafreddo, M.; Ballarini, P.; Pucciarelli, S.; Miceli, C. An In-Silico Comparative Study of Lipases from the Antarctic Psychrophilic Ciliate Euplotes focardii and the Mesophilic Congeneric Species Euplotes crassus: Insight into Molecular Cold-Adaptation. Mar. Drugs 2021, 19, 67. [Google Scholar] [CrossRef]
- Park, H.J.; Kim, D.; Kim, I.H.; Lee, C.-E.; Kim, I.-C.; Kim, J.Y.; Kim, S.J.; Lee, H.K.; Yim, J.H. Characteristics of cold-adaptive endochitinase from Antarctic bacterium Sanguibacter antarcticus KOPRI 21702. Enzym. Microb. Technol. 2009, 45, 391–396. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, Y.; Tan, H.; Chi, N.; Zhang, Q.; Du, Y.; Yin, H. Characterisation of a chitinase from Pseudoalteromonas sp. DL-6, a marine psychrophilic bacterium. Int. J. Biol. Macromol. 2014, 70, 455–462. [Google Scholar] [CrossRef]
- Goni, O.; Sanchez-Ballesta, M.T.; Merodio, C.; Escribano, M.I. Two cold-induced family 19 glycosyl hydrolases from cherimoya (Annona cherimola) fruit: An antifungal chitinase and a cold-adapted chitinase. Phytochemistry 2013, 95, 94–104. [Google Scholar] [CrossRef]
- Deng, J.J.; Shi, D.; Mao, H.H.; Li, Z.W.; Liang, S.; Ke, Y.; Luo, X.C. Heterologous expression and characterization of an antifungal chitinase (Chit46) from Trichoderma harzianum GIM 3.442 and its application in colloidal chitin conversion. Int. J. Biol. Macromol. 2019, 134, 113–121. [Google Scholar] [CrossRef]
- Abdel Wahab, W.A.; Abd El Aty, A.A.; Mostafa, F.A. Improvement of catalytic, thermodynamics and antifungal activity of constitutive Trichoderma longibrachiatum KT693225 exochitinase by covalent coupling to oxidized polysaccharides. Int. J. Biol. Macromol. 2018, 112, 179–187. [Google Scholar] [CrossRef]
- Kidibule, P.E.; Santos-Moriano, P.; Jimenez-Ortega, E.; Ramirez-Escudero, M.; Limon, M.C.; Remacha, M.; Plou, F.J.; Sanz-Aparicio, J.; Fernandez-Lobato, M. Use of chitin and chitosan to produce new chitooligosaccharides by chitinase Chit42: Enzymatic activity and structural basis of protein specificity. Microb. Cell Fact. 2018, 17, 47. [Google Scholar] [CrossRef]
- Luong, N.N.; Tien, N.Q.D.; Huy, N.X.; Tue, N.H.; Man, L.Q.; Sinh, D.D.H.; Van Thanh, D.; Chi, D.T.K.; Hoa, P.T.B.; Loc, N.H. Expression of 42 kDa chitinase of Trichoderma asperellum (Ta-CHI42) from a synthetic gene in Escherichia coli. FEMS Microbiol. Lett. 2021, 368, fnab110. [Google Scholar] [CrossRef] [PubMed]
- Jinzhu, S.; Qian, Y.; Beidong, L.; Dianfu, C. Expression of the chitinase gene from Trichoderma aureoviride in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2005, 69, 39–43. [Google Scholar] [CrossRef] [PubMed]
- de la Cruz, J.; Hidalgo-Gallego, A.; Lora, J.M.; Benitez, T.; Pintor-Toro, J.A.; Llobell, A. Isolation and characterization of three chitinases from Trichoderma harzianum. Eur. J. Biochem. 1992, 206, 859–867. [Google Scholar] [CrossRef]
- Jimenez-Ortega, E.; Kidibule, P.E.; Fernandez-Lobato, M.; Sanz-Aparicio, J. Structure-Function Insights into the Fungal Endo-Chitinase Chit33 Depict its Mechanism on Chitinous Material. Int. J. Mol. Sci. 2022, 23, 7599. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.L.; Wang, S.; Yang, D.F.; Yang, L.Y.; Wang, Q.Y.; Yu, J.; Li, N.; Pan, L.X. The Discovery, Enzymatic Characterization and Functional Analysis of a Newly Isolated Chitinase from Marine-Derived Fungus Aspergillus fumigatus df347. Mar. Drugs 2022, 20, 520. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, Y.; Ma, J.; Yan, Q.; Jiang, Z.; Yang, S. Biochemical characterization of a bifunctional chitinase/lysozyme from Streptomyces sampsonii suitable for N-acetyl chitobiose production. Biotechnol. Lett. 2020, 42, 1489–1499. [Google Scholar] [CrossRef]
- Zhang, W.; Ma, J.; Yan, Q.; Jiang, Z.; Yang, S. Biochemical characterization of a novel acidic chitinase with antifungal activity from Paenibacillus xylanexedens Z2-4. Int. J. Biol. Macromol. 2021, 182, 1528–1536. [Google Scholar] [CrossRef]
- Perez-Martinez, A.S.; De Leon-Rodriguez, A.; Harris, L.J.; Herrera-Estrella, A.; Barba de la Rosa, A.P. Overexpression, purification and characterization of the Trichoderma atroviride endochitinase, Ech42, in Pichia pastoris. Protein. Expr. Purif. 2007, 55, 183–188. [Google Scholar] [CrossRef]
- Loc, N.H.; Huy, N.D.; Quang, H.T.; Lan, T.T.; Thu Ha, T.T. Characterisation and antifungal activity of extracellular chitinase from a biocontrol fungus, Trichoderma asperellum PQ34. Mycology 2020, 11, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Han, H.; Wang, D.; Guo, X.; Zhou, Y.; Fukamizo, T.; Yang, Q. Potent Fungal Chitinase for the Bioconversion of Mycelial Waste. J. Agric. Food Chem. 2020, 68, 5384–5390. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Salwan, R.; Sharma, P.N.; Kanwar, S.S. Molecular cloning and characterization of ech46 endochitinase from Trichoderma harzianum. Int. J. Biol. Macromol. 2016, 92, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Krolicka, M.; Hinz, S.W.A.; Koetsier, M.J.; Joosten, R.; Eggink, G.; van den Broek, L.A.M.; Boeriu, C.G. Chitinase Chi1 from Myceliophthora thermophila C1, a Thermostable Enzyme for Chitin and Chitosan Depolymerization. J. Agric. Food Chem. 2018, 66, 1658–1669. [Google Scholar] [CrossRef]
- Kumar, M.; Brar, A.; Vivekanand, V.; Pareek, N. Process optimization, purification and characterization of a novel acidic, thermostable chitinase from Humicola grisea. Int. J. Biol. Macromol. 2018, 116, 931–938. [Google Scholar] [CrossRef]
- Xie, X.H.; Fu, X.; Yan, X.Y.; Peng, W.F.; Kang, L.X. A Broad-Specificity Chitinase from Penicillium oxalicum k10 Exhibits Antifungal Activity and Biodegradation Properties of Chitin. Mar. Drugs 2021, 19, 356. [Google Scholar] [CrossRef]
- Zhang, M.; Puri, A.K.; Govender, A.; Wang, Z.; Singh, S.; Permaul, K. The multi-chitinolytic enzyme system of the compost-dwelling thermophilic fungus Thermomyces lanuginosus. Process Biochem. 2015, 50, 237–244. [Google Scholar] [CrossRef]
- Duarte, A.W.F.; Dos Santos, J.A.; Vianna, M.V.; Vieira, J.M.F.; Mallagutti, V.H.; Inforsato, F.J.; Wentzel, L.C.P.; Lario, L.D.; Rodrigues, A.; Pagnocca, F.C.; et al. Cold-adapted enzymes produced by fungi from terrestrial and marine Antarctic environments. Crit. Rev. Biotechnol. 2018, 38, 600–619. [Google Scholar] [CrossRef]
- Wang, X.; Chi, N.; Bai, F.; Du, Y.; Zhao, Y.; Yin, H. Characterization of a cold-adapted and salt-tolerant exo-chitinase (ChiC) from Pseudoalteromonas sp. DL-6. Extremophiles 2016, 20, 167–176. [Google Scholar] [CrossRef]
- Lv, C.; Gu, T.; Ma, R.; Yao, W.; Huang, Y.; Gu, J.; Zhao, G. Biochemical characterization of a GH19 chitinase from Streptomyces alfalfae and its applications in crystalline chitin conversion and biocontrol. Int. J. Biol. Macromol. 2021, 167, 193–201. [Google Scholar] [CrossRef]
- Rani, T.S.; Madhuprakash, J.; Podile, A.R. Chitinase-E from Chitiniphilus shinanonensis generates chitobiose from chitin flakes. Int. J. Biol. Macromol. 2020, 163, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Cheng, G.; Li, Q.; Jiao, S.; Feng, C.; Zhao, X.; Yin, H.; Du, Y.; Liu, H. Chitin Oligosaccharide Modulates Gut Microbiota and Attenuates High-Fat-Diet-Induced Metabolic Syndrome in Mice. Mar. Drugs 2018, 16, 66. [Google Scholar] [CrossRef]
- Baharlouei, P.; Rahman, A. Chitin and Chitosan: Prospective Biomedical Applications in Drug Delivery, Cancer Treatment, and Wound Healing. Mar. Drugs 2022, 20, 460. [Google Scholar] [CrossRef]
- Song, W.; Zhang, N.; Yang, M.; Zhou, Y.; He, N.; Zhang, G. Multiple strategies to improve the yield of chitinase a from Bacillus licheniformis in Pichia pastoris to obtain plant growth enhancer and GlcNAc. Microb. Cell Fact. 2020, 19, 181. [Google Scholar] [CrossRef] [PubMed]
- Noby, N.; Auhim, H.S.; Winter, S.; Worthy, H.L.; Embaby, A.M.; Saeed, H.; Hussein, A.; Pudney, C.R.; Rizkallah, P.J.; Wells, S.A.; et al. Structure and in silico simulations of a cold-active esterase reveals its prime cold-adaptation mechanism. Open Biol. 2021, 11, 210182. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, K.S.; Cavicchioli, R. Cold-adapted enzymes. Annu. Rev. Biochem. 2006, 75, 403–433. [Google Scholar] [CrossRef]
- Yang, G.; Yao, H.; Mozzicafreddo, M.; Ballarini, P.; Pucciarelli, S.; Miceli, C. Rational Engineering of a Cold-Adapted Amylase from the Antarctic Ciliate Euplotes focardii for Simultaneous Improvement of Thermostability and Catalytic Activity. Appl. Environ. Microbiol. 2017, 83, e00449-17. [Google Scholar] [CrossRef]
- Yang, G.; De Santi, C.; de Pascale, D.; Pucciarelli, S.; Pucciarelli, S.; Miceli, C. Characterization of the first eukaryotic cold-adapted patatin-like phospholipase from the psychrophilic Euplotes focardii: Identification of putative determinants of thermal-adaptation by comparison with the homologous protein from the mesophilic Euplotes crassus. Biochimie 2013, 95, 1795–1806. [Google Scholar]
- Yang, G.; Yang, G.; Aprile, L.; Turturo, V.; Pucciarelli, S.; Pucciarelli, S.; Miceli, C. Characterization and comparative analysis of psychrophilic and mesophilic alpha-amylases from Euplotes species: A contribution to the understanding of enzyme thermal adaptation. Biochem. Biophys. Res. Commun. 2013, 438, 715–720. [Google Scholar] [CrossRef]
- van der Ent, F.; Lund, B.A.; Svalberg, L.; Purg, M.; Chukwu, G.; Widersten, M.; Isaksen, G.V.; Brandsdal, B.O.; Aqvist, J. Structure and Mechanism of a Cold-Adapted Bacterial Lipase. Biochemistry 2022, 61, 933–942. [Google Scholar] [CrossRef]
- Lv, J.; Zhang, Y.; Ma, M.; Oh, D.H.; Fu, X. Characterization of chitinase from Exiguobacterium antarcticum and its bioconversion of crayfish shell into chitin oligosaccharides. Food Res. Int. 2022, 158, 111517. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, P.; Zhu, M.; Chen, W.; Yu, S.; Zhong, B. Overexpression and Biochemical Properties of a GH46 Chitosanase from Marine Streptomyces hygroscopicus R1 Suitable for Chitosan Oligosaccharides Preparation. Front. Microbiol. 2021, 12, 816845. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, X.; Chen, H.; Lin, B.; Zhao, L. Heterologous Expression and Characterization of a High-Efficiency Chitosanase from Bacillus mojavensis SY1 Suitable for Production of Chitosan Oligosaccharides. Front. Microbiol. 2021, 12, 781138. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Lin, B.; Zhang, R.; Gong, Z.; Wen, M.; Su, W.; Zhou, J.; Zhao, L.; Wang, J. Controllable preparation of chitosan oligosaccharides via a recombinant chitosanase from marine Streptomyces lydicus S1 and its potential application on preservation of pre-packaged tofu. Front. Microbiol. 2022, 13, 1007201. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Step | Total Activity (U) | Protein (mg) | Specific Activity (U/mg) | Purification Factor (fold) | Recovery (%) |
|---|---|---|---|---|---|
| Crude culture | 1076.8 ± 42.5 | 468.2 ± 22.1 | 2.3 ± 0.1 | 1 | 100 |
| Ultrafiltration | 764.2 ± 35.2 | 201.1 ± 9.5 | 3.8 ± 0.2 | 1.65 | 71 |
| QSFF chromatography | 204.6 ± 12.3 | 5.6 ± 0.3 | 36.6 ± 1.6 | 16.8 | 19 |
| Substrate | Activity (U/mg) |
|---|---|
| Powdery chitin | 5.7 ± 0.2 |
| Colloidal chitin | 36.6 ± 1.8 |
| ball milled chitin | 27.9 ± 1.6 |
| xylan | ND * |
| microcrystalline cellulose | ND |
| Colloidal chitosan with 85% DDA | 1.1 ± 0.1 |
| Colloidal chitosan with 90% DDA | 1.3 ± 0.1 |
| Colloidal chitosan with 95% DDA | 1.3 ± 0.1 |
| pretreated shrimp shell powder | 4.5 ± 0.3 |
| Microorganism | Optimum Temperature (°C) | Relative Activity at 5 °C (%) | Relative Activity at 10 °C (%) | Relative Activity at 30 °C (%) | Reference |
|---|---|---|---|---|---|
| T. gamsii R1 | 40 | 40.2 | 50.1 | 82.1 | This study |
| T. harzianum GIM 3.442 | 45 | NR * | NR | about 48% | [20] |
| T. harzianum CECT2413 | 35 | NR | NR | about 88% | [22] |
| T. asperellum SH16 | 45 | NR | NR | about 62% | [23] |
| T. asperellum PQ34 | 45 | NR | NR | about 60% | [33] |
| A. niger CBS 513.88 | 40 | NR | NR | about 83% | [34] |
| T. harzianum ThHP3 | 60 | about 20% | about 23% | about 62% | [31] |
| Myceliophthora thermophila C1 | 55 | NR | NR | about 31% | [32] |
| H. grisea ITCC 10,360.16 | 70 | NR | NR | NR | [35] |
| Penicillium oxalicum k10 | 40 | NR | NR | about 62% | [36] |
| Aspergillus fumigatus df347 | 45 | NR | NR | about 47% | [27] |
| Thermomyces lanuginosus SSBP | 50 | NR | NR | NR | [37] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Zhu, M.; Wang, P.; Chen, W. Biochemical Properties of a Cold-Active Chitinase from Marine Trichoderma gamsii R1 and Its Application to Preparation of Chitin Oligosaccharides. Mar. Drugs 2023, 21, 332. https://doi.org/10.3390/md21060332
Wang J, Zhu M, Wang P, Chen W. Biochemical Properties of a Cold-Active Chitinase from Marine Trichoderma gamsii R1 and Its Application to Preparation of Chitin Oligosaccharides. Marine Drugs. 2023; 21(6):332. https://doi.org/10.3390/md21060332
Chicago/Turabian StyleWang, Jianrong, Mujin Zhu, Ping Wang, and Wei Chen. 2023. "Biochemical Properties of a Cold-Active Chitinase from Marine Trichoderma gamsii R1 and Its Application to Preparation of Chitin Oligosaccharides" Marine Drugs 21, no. 6: 332. https://doi.org/10.3390/md21060332