
Protective Properties of Marine Alkyl Glycerol Ethers in Chronic Stress
Abstract
:1. Introduction
2. Results
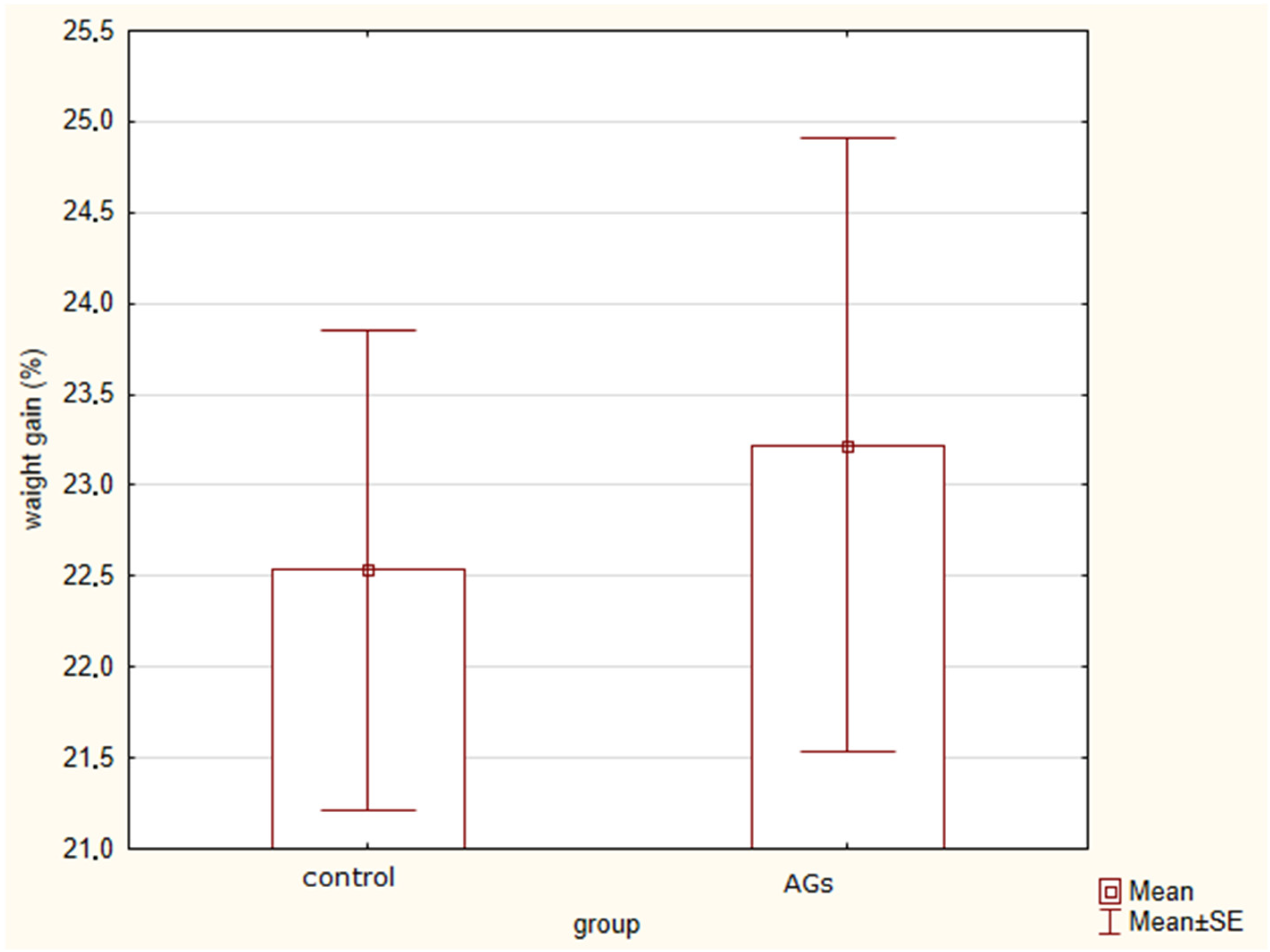
2.1. Changes in Body Weight in Rats Treated with AGs
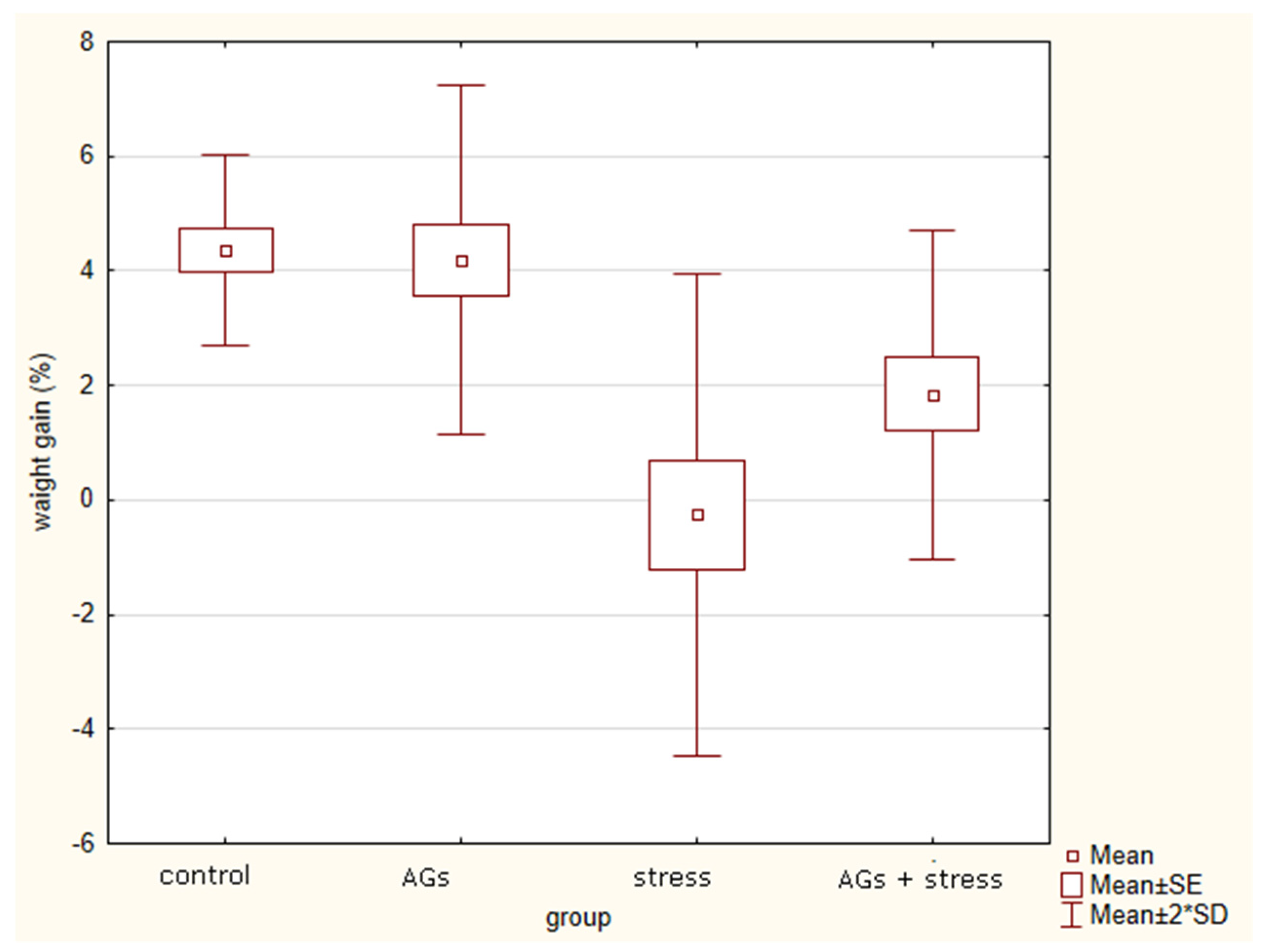
2.2. Body Weight Changes under Stress
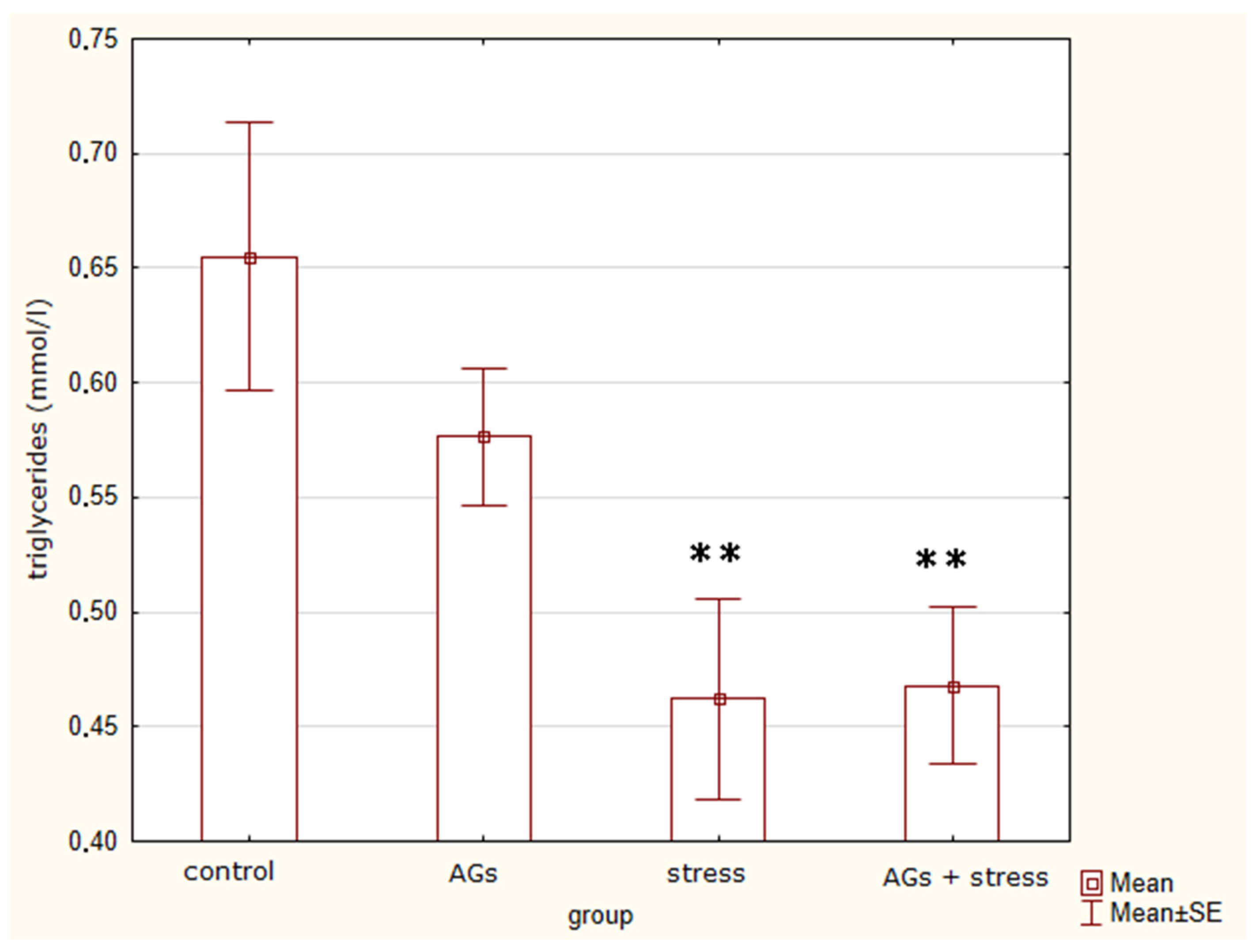
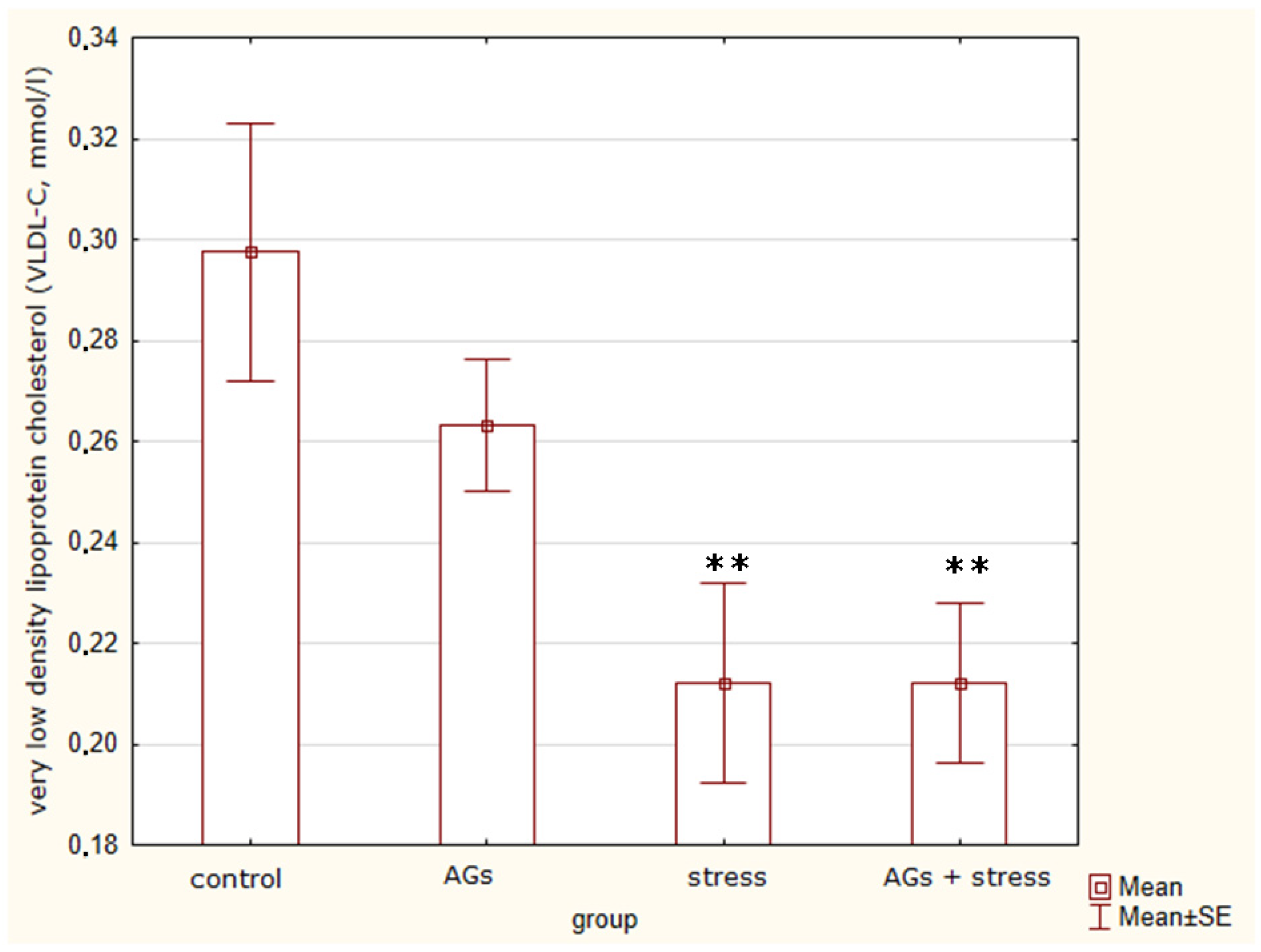
2.3. Content of Lipoproteins in Blood Serum
2.4. Effect of AGs on Content and Composition of Fatty Acids and Plasmalogen Levels in Plasma Lipids
2.5. Blood Test Results
2.6. Morphometry
2.7. Hormone Levels
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Preparations
4.2.1. Preparation of the AGs
4.2.2. Determination of AGs Composition
4.3. Biological Experiment
4.3.1. Animal Treatment
4.3.2. Stress Procedure
4.3.3. Measurement of Body Weight
4.3.4. Collection of Samples
4.3.5. The Serum Lipid Spectrum of Rats
4.3.6. The Content of Fatty Acids and Dimethyl Acetals (DMAs) in the Total Plasma Lipids of Rats
4.3.7. Hormone Research
4.3.8. Hematological Analysis
4.4. Statistical Analysis
5. Conclusions
- AGs prevent many of the negative effects of acute and chronic stress. In particular, they improve the immune status of animals under chronic stress.
- Preparations with the participation of AGs can be useful in the prevention and treatment of socially significant diseases of today’s world.
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weight Gain (%) | AGs | Stress | AGs + Stress | |
|---|---|---|---|---|
| 1st week | 4.25 3.5–5.4 | 0.7 −2.0–1.2 | 1.8 1.2–2.3 | H = 9.0 p = 0.01 * |
| 2nd week | 3.6 0.3–4.7 | 1.7 0.3–2.4 | 1.0 −5.8–2 | H = 3.0 p = 0.22 |
| 3d period (about week) | 1.5 −2.1–2.6 | −2 −0.9–−2.4 | 0.4 0.0–0.4 | H = 4 p = 0.13 |
| general weight gain (about 3 weeks) | 7.6 6.3–7.6 | 0.3 −4–2.5 | 1.0 −4–1.8 | H = 9.3 p = 0.01 * |
Appendix B

Appendix C
| FAME/DMA | Control | AG | Stress | AG + Stress |
|---|---|---|---|---|
| 14:0 | 0.35 ± 0.05 | 0.29 ± 0.03 | 0.26 ± 0.05 * | 0.36 ± 0.08 |
| 15:0 | 0.34 ± 0.11 | 0.30 ± 0.02 | 0.30 ± 0.04 | 0.35 ± 0.08 |
| 16:0DMA | 0.08 ± 0.03 | 0.14 ± 0.02 * | 0.08 ± 0.02 | 0.16 ± 0.02 * |
| 16:0 | 19.01 ± 1.02 | 18.21 ± 0.76 | 18.14 ± 1.13 | 18.61 ± 2.64 |
| 16:1n-9 | 0.07 ± 0.13 | 0.09 ± 0.11 | 0.12 ± 0.08 | 0.04 ± 0.09 |
| 16:1n-7 | 1.09 ± 0.16 | 0.80 ± 0.26 | 0.69 ± 0.14 * | 0.69 ± 0.23 * |
| 17:0 | 0.34 ± 0.07 | 0.36 ± 0.03 | 0.32 ± 0.02 | 0.42 ± 0.08 |
| 18:0DMA | 0.08 ± 0.02 | 0.10 ± 0.03 | 0.07 ± 0.03 | 0.08 ± 0.04 |
| 18:0 | 11.30 ± 0.91 | 10.72 ± 0.66 | 10.95 ± 0.63 | 12.27 ± 1.22 |
| 18:1n-9 | 11.82 ± 1.72 | 10.23 ± 1.7 | 9.93 ± 0.97 | 9.14 ± 0.79 * |
| 18:1n-7 | 1.83 ± 0.18 | 1.22 ± 0.82 | 1.65 ± 0.11 | 1.71 ± 0.14 |
| 18:2n-6 | 24.96 ± 1.04 | 23.56 ± 1.46 | 23.63 ± 0.45 | 23.76 ± 3.01 |
| 18:3n-6 | 0.17 ± 0.02 | 0.27 ± 0.03 * | 0.25 ± 0.04 * | 0.17 ± 0.12 |
| 18:3n-3 | 0.91 ± 0.08 | 0.84 ± 0.13 | 0.76 ± 0.03 * | 0.63 ± 0.04 |
| 20:1n-11 | 0.06 ± 0.08 | 0.03 ± 0.06 | 0.02 ± 0.05 | 0.04 ± 0.08 |
| 20:1n-9 | 0.25 ± 0.06 | 0.19 ± 0.01 | 0.19 ± 0.03 | 0.24 ± 0.05 |
| 20:1n-7 | 0.33 ± 0.12 | 0.38 ± 0.12 | 0.37 ± 0.04 | 0.41 ± 0.14 |
| 20:2n-6 | 0.26 ± 0.05 | 0.23 ± 0.03 | 0.20 ± 0.04 | 0.32 ± 0.05 |
| 20:3n-6 | 0.40 ± 0.07 | 0.32 ± 0.03 | 0.33 ± 0.04 | 0.41 ± 0.12 |
| 20:4n-6 | 21.33 ± 4.34 | 26.03 ± 2.91 | 26.09 ± 1.89 | 24.64 ± 2.74 |
| 20:5n-3 | 1.10 ± 0.09 | 1.07 ± 0.17 | 1.02 ± 0.14 | 1.01 ± 0.21 |
| 22:4n-6 | 0.37 ± 0.08 | 0.39 ± 0.1 | 0.44 ± 0.09 | 0.54 ± 0.14 |
| 22:5n-3 | 0.73 ± 0.05 | 0.81 ± 0.22 | 0.69 ± 0.13 | 0.71 ± 0.1 |
| 22:6n-3 | 2.82 ± 0.58 | 3.40 ± 0.18 | 3.49 ± 0.44 | 3.33 ± 0.6 |
| 16:0DMA/16:0 × 00 | 0.44 ± 0.15 | 0.76 ± 0.11 * | 0.46 ± 0.12 | 0.85 ± 0.07 * |
| 18:0DMA/18:0 × 100 | 0.73 ± 0.25 | 0.93 ± 0.2 | 0.62 ± 0.27 | 0.61 ± 0.32 |
| SFA | 31.35 ± 1.99 | 29.89 ± 0.68 | 29.97 ± 1.64 | 32.01 ± 3.99 |
| MUFA | 15.44 ± 2.07 | 12.95 ± 1.31 | 12.98 ± 1.21 | 12.26 ± 0.86 * |
| PUFA | 49.05 ± 11.47 | 56.58 ± 1.93 | 56.9 ± 2.28 | 55.24 ± 3.62 |
| n-6/n-3 | 7.76 ± 1.47 | 8.28 ± 0.79 | 8.60 ± 0.61 | 8.77 ± 0.55 |
Appendix D
| Indicator | Control | AGs | Stress | AGs + Stress | |
|---|---|---|---|---|---|
| Hematocrit, % | 46.0 44.2–46.0 | 43.5 42.2–44.2 | 42.6 38.6–44.4 | 41.4 40.1–43.6 | H = 4.3′ p = 0.22 |
| Red blood cells, 1012/L | 8.1 ± 0.2 | 7.9 ± 0.2 | 7.6 ± 0.48 | 7.62 ± 0.5 | F = 1.95″ p = 0.17 |
| Mean corpuscular volume, fl | 55.4 ± 0.8 | 55.1 ± 1.4 | 55.4 ± 1.2 | 55.2 ± 0.7 | F = 0.11 p = 0.95 |
| Hemoglobin, g/L | 147.3 ± 3 | 140.4 ± 6.8 | 133.3 ± 10 | 135.8 ± 6.7 | F = 2.5 p = 0.11 |
| Mean corpuscular hemoglobin concentration, g/L | 325 321–326 | 321.5 319–324 | 319 316.5–325.5 | 326 325–326 | H = 2.87 p = 0.41 |
| Mean corpuscular hemoglobin, pg | 17.8 ± 0.13 | 17.67 ± 0.45 | 17.78 ± 0.43 | 17.7 ± 0.44 | F = 0.27 p = 0.84 |
| Red cell distribution width, coefficient of variation, % | 15.1 ± 0.6 | 14.2 ± 0.7 | 14.5 ± 1.1 | 13.8 ± 0.8 | F = 2.16 p = 0.14 |
| Red cell distribution width, standard deviation, fl | 29.9 ± 1.5 | 28.5 ± 1.5 | 29.4 ± 1.9 | 28.5 ± 1.6 | F = 0.86 p = 0.48 |
| Platelets, 109/L | 248 197–665 | 667.5 397–711 | 577.5 381–651.5 | 570 611–688 | H = 1.58 p = 0.66 |
| Mean platelet volume, fl | 6.7 6.4–7 | 6.25 6.0–6.6 | 6.15 5.9–6.25 | 5.9 5.8–6.1 | H = 5.25 p = 0.15 |
| Platelet distribution width, % | 15.6 15.6–15.6 | 16.1 15.5–16.3 | 15.7 15.6–16.5 | 15.5 15.4–15.8 | H = 1.59 p = 0.66 |
| Platelet crit, % | 0.19 0.13–0.47 | 0.44 0.25–0.47 | 0.35 0.24–0.38 | 0.35 0.35–0.39 | H = 1.79 p = 0.62 |
| White blood cells, 109/L | 4.95 3.9–5.8 | 4.2 2.5–4.8 | 4.55 3.6–5.9 | 5.7 4–6 | H = 2.3 p = 0.5 |
| Lymphocytes, 109/L | 2.5 2.3–3.1 | 2.5 1.1–2.9 | 2.15 2.15–3.1 | 3.6 2.5–4.2 | H = 2.75 p = 0.43 |
| Lymphocytes, % | 58.3 45.2–61.3 | 55.9 36.1–63 | 47.7 41.2–53.55 | 66.7 62.8–67 | H = 8.49 p = 0.037 * |
| Neutrophils, 109/L | 1.4 1.1–1.75 | 1.15 0.9–1.5 | 1.85 1.45–2.05 | 1.3 1.2–1.6 | H = 4.05 p = 0.26 |
| Neutrophils, % | 28 ± 3.8 | 27.9 ± 3 | 34.5 ± 10.2 | 26.2 ± 3.2 | F = 0.15 p = 0.47 |
| Neutrophil–lymphocyte ratio | 0.5 0.42–0.56 | 0.6 0.43–0.81 | 0.67 0.6–0.99 | 0.41 0.37–0.44 | H = 9.15 p = 0.0273 * |
| MID (mixed), 109/L | 0.4 0.3–0.5 | 0.2 0.3–0.4 | 0.45 0.4–0.6 | 0.5 0.3–0.5 | H = 3.88 p = 0.27 |
| MID (mixed), % | 9.7 9.4–10.5 | 7.45 6.5–9.5 | 9.9 8.5–11.9 | 8.4 8.0–8.4 | H = 7.16 p = 0.067 |
References
- Selye, H. The Physiology and Pathology of Exposure to Stress; ACTA, Inc.: Montreal, QC, Canada, 1950. [Google Scholar]
- Dorninger, F.; Forss-Petter, S.; Wimmerb, I.; Berger, J. Plasmalogens, platelet-activating factor and beyond—Ether lipids in signaling and neurodegeneration. Neurobiol. Dis. 2020, 145, 105061. [Google Scholar] [CrossRef]
- Braverman, N.E.; Moser, A.B. Functions of plasmalogen lipids in health and disease. Biochim. Biophys. Acta 2012, 1822, 1442–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, S.; Lancaster, G.I.; Meiklea, P.J. Plasmalogens: A potential therapeutic target for neurodegenerative and cardiometabolic disease. Prog. Lipid Res. 2019, 74, 186–195. [Google Scholar] [CrossRef]
- Poleschuk, T.S.; Sultanov, R.M.; Ermolenko, E.V.; Shulgina, L.V.; Kasyanov, S.P. Protective action of alkylglycerols under stress. Stress 2019, 23, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, N.; Maiti, P.P.; Choudhary, A.; Tuli, A.; Masih, D.; Khan, M.M.U.; Ara, T. Animal models in the study of stress: A review. NSHM J. Pharm. Healthc. Manag. 2011, 2, 42–50. [Google Scholar]
- Lee, H.-J.; Park, H.-J.; Starkweather, A.; An, K.; Shim, I. Decreased Interleukin-4 release from the neurons of the locus coeruleus in response to immobilization stress. Mediat. Inflamm. 2016, 2016, 3501905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khuzhakhmetova, L.K.; Teply, D.L. Pharmacological correction of lipid peroxidation and peroxide hemolysis of red blood cells in mature rats during immobilization stress. Nat. Sci. 2016, 2, 66–71. [Google Scholar] [CrossRef]
- Tian, W.; Akanda, M.R.; Islam, A.; Yang, H.-D.; Lee, S.-C.; Lee, J.-H.; Kim, S.-K.; Choi, Y.-J.; Im, S.-Y.; Park, B.-Y. The anti-stress effect of mentha arvensis in immobilized rats. Int. J. Mol. Sci. 2018, 19, 355. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Sun, S.; Tang, N.; Cai, W.; Qian, L. Oral administration of alkylglycerols differentially modulates high-fat diet-induced obesity and insulin resistance in mice. Evid.-Based Complement. Altern. Med. 2013, 2013, e11. [Google Scholar] [CrossRef] [Green Version]
- Perevoznikov, I.E.; Poleschuk, T.S.; Velansky, P.V. Effect of chronic immobilization stress and 1-0-alkylglycerols on fatty acid composition of rat’s spleen. In Anniversary International Scientific Conference “90 Years—From a Plant to a Drug: Achievements and Prospects” Collection of Scientific Papers; VILAR: Moscow, Russia, 2021. [Google Scholar]
- Swan, M.P.; Hickman, D.L. Evaluation of the neutrophil-lymphocyte ratio as a measure of distress in rats. Lab Anim. 2014, 43, 276–282. [Google Scholar] [CrossRef]
- Qian, L.; Zhang, M.; Wu, S.; Zhong, Y.; Van Tol, E.; Cai, W. Alkylglycerols modulate the proliferation and differentiation of non-specific agonist and specific antigen-stimulated splenic lymphocytes. PLoS ONE 2014, 9, e96207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Facciotti, F.; Ramanjaneyulu, G.S.; Lepore, M.; Sansano, S.; Cavallari, M.; Kistowska, M.; Forss-Petter, S.; Ni, G.; Colone, A.; Singhal, A.; et al. Peroxisome-derived lipids are self antigens that stimulate invariant natural killer T cells in the thymus. Nat. Immunol. 2012, 13, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Vitorino, D.C.; Buzzachera, C.F.; Curi, R.; Fernandes, L.C. Effect of chronic supplementation with shark liver oil on immune responses of exercise-trained rats. Eur. J. Appl. Physiol. 2010, 108, 1225–1232. [Google Scholar] [CrossRef] [PubMed]
- Mitre, R.; Etienne, M.; Martinais, S.; Salmon, H.; Allaume, P.; Legrand, P.; Legrand, A.B. Humoral defence improvement and haematopoiesis stimulation in sows and offspring by oral supply of shark-liver oil to mothers during gestation and lactation. Br. J. Nutr. 2005, 94, 753–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmieri, B.; Pennelli, A.; Di Cerbo, A. Jurassic surgery and immunity enhancement by alkyglycerols of shark liver oil. Lipids Health Dis. 2014, 13, 178. [Google Scholar] [CrossRef] [Green Version]
- Iannitti, T.; Palmieri, B. An update on the therapeutic role of alkylglycerols. Mar. Drugs 2010, 8, 2267–2300. [Google Scholar] [CrossRef] [Green Version]
- Bozelli, J.C.; Azher, S.; Epand, R.M. Plasmalogens and Chronic Inflammatory Diseases. Front. Physiol. 2021, 12, 730829. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1957, 37, 911–917. [Google Scholar] [CrossRef]
- Christie, W.W. Lipid Analysis: Isolation, Separation, Identification and Structural Analysis of Lipids; The Oily Press: Bridgwater, UK, 2003. [Google Scholar]
- Ermolenko, E.V.; Latyshev, N.A.; Sultanov, R.M.; Kasyanov, S.P. Technological approach of 1-O-alkyl-sn-glycerols separation from Berryteuthis magister squid liver oil. J. Food Sci. Technol. 2016, 53, 1722–1726. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.P.; Dinger, N.; Levine, B.S. Stress produced by gavage administration in the rat. Contemp. Top. Lab. Anim. Sci. 2000, 39, 17–21. [Google Scholar]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimationof the concentrationof low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Muniz, F.J.; Bastida, S. Do not use the Friedewald formula to calculate LDL-cholestero in hypercholesterolaemic rats. Eur. J. Lipid Sci. Technol. 2008, 110, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.-P.; Li, R.; Dai, W.; Yu, B.-L.; Chen, L.-Z.; Huang, X.-S. Xuezhikang contributes to greater triglyceride reduction than simvastatin in hypertriglyceridemia rats by up-regulating apolipoprotein A5 via the PPARα signaling pathway. PLoS ONE 2017, 12, e0184949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhardwaj, S.; Bhattacharjee, J.; Bhatnagar, M.K.; Tyagi, S. Atherogenic index of plasma, Castelli risk index and atherogenic coefficient—New parameters in assessing cardiovascular risk. Int. J. Pharm. Biol. Sci. 2013, 3, 359–364. Available online: www.ijpbs.com (accessed on 15 December 2022).
- Carreau, J.P.; Dubacq, J.P. Adaptation of a macro-scale method to the micro-scale for fatty acid methyl transesterification of biological lipid extracts. J. Chromatogr. A 1978, 151, 384–390. [Google Scholar] [CrossRef]
- Bjorkhem, I.; Sisfontes, L.; Bostrom, B.; Kase, B.F.; Blomstrand, R. Simple diagnosis of the Zellweger syndrome by gas-liquid chromatography of dimethyl acetals. J. Lipid Res. 1986, 27, 786–791. [Google Scholar] [CrossRef]
- Scheff, S.W. Fundamental Statistical Principles for the Neurobiologist; Academic Press: London, UK, 2016. [Google Scholar]







| Indicator | Control | AGs | Stress | AGs + Stress | |
|---|---|---|---|---|---|
| Total cholesterol, mmol/L | 1.8 1.4–2.05 | 1.6 1.2–1.6 | 1.4 1.2–1.5 | 1.1 1.1–1.4 | H = 6.64′ p = 0.08 |
| HDL-C, mmol/L | 0.88 ± 0.16 | 0.98 ± 0,16 | 0.93 ± 0.16 | 0.9 + 0.19 | F = 0.34″ p = 0.79 |
| LDL-C, mmol/L | 0.55 0.24–0.85 | 0.18 0.1–0.45 | 0.15 0.06–0.26 | 0.1 0.05–0.11 | H = 3.84 p = 0.28 |
| VLDL-C, mmol/L | 0.3 ± 0.05 | 0.26 ± 0.03 | 0.21 ± 0.04 ** | 0.21 ± 0.04 ** | F = 4.9 p = 0.013 * |
| Atherogenic coefficient | 0.89 0.65–1.34 | 0.43 0.36–0.7 | 0.34 0.33–0.56 | 0.45 0.22–0.47 | H = 4.45 p = 0.22 |
| Triglycerides, mmol/L | 0.66 ± 0.12 | 0.58 ± 0.07 | 0.46 ± 0.1 ** | 0.47 ± 0.08 ** | F = 4.8 p = 0.014 * |
| Indicator | Control | AGs | Stress | AGs + Stress | |
|---|---|---|---|---|---|
| Spleen index | 0.32 0.29–0.35 | 0.29 0.24–0.32 | 0.29 0.28–0.39 | 0.28 0.23–0.34 | H = 1.65′ p = 0.65 |
| Glandula adrenalis index | 0.026 0.024–0.027 | 0.028 0.026–0.028 | 0.027 0.027–0.028 | 0.027 0.027–0.03 | H = 3.11 p = 0.38 |
| Thymus index | 0.098 ± 0.02 | 0.1 ± 0.014 | 0.093 ± 0.037 | 0.089 ± 0.008 | F = 0.26″ p = 0.85 |
| Indicator | Control | AGs | Stress | AGs + Stress | |
|---|---|---|---|---|---|
| Testosterone, nmol/L | 12.6 11.8–18.3 | 11.7 8.2–16.9 | 24.4 7.65–31.6 | 16.5 11.8–24.7 | H = 1.26′ p = 0.74 |
| Cortisol, nmol/L | 13.7 10.7–18.2 | 56.9 17.5–58.3 | 32.1 13.7–38.4 | 28.3 17.6–51 | F = 1.4″ p = 0.28 |
| Alkyl Chain | AGs Composition | Trivial Name |
|---|---|---|
| C14:0 a | 1.52 ± 0.15 b | |
| C16:0 | 93.77 ± 0.16 | Chimyl alcohol |
| C18:0 | 4.71 ± 0.25 | Batyl alcohol |
| Σsat | 100 | |
| Σunsat | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sultanov, R.M.; Poleshchuk, T.S.; Ermolenko, E.V.; Kasyanov, S.P. Protective Properties of Marine Alkyl Glycerol Ethers in Chronic Stress. Mar. Drugs 2023, 21, 202. https://doi.org/10.3390/md21040202
Sultanov RM, Poleshchuk TS, Ermolenko EV, Kasyanov SP. Protective Properties of Marine Alkyl Glycerol Ethers in Chronic Stress. Marine Drugs. 2023; 21(4):202. https://doi.org/10.3390/md21040202
Chicago/Turabian StyleSultanov, Ruslan M., Tatiana S. Poleshchuk, Ekaterina V. Ermolenko, and Sergey P. Kasyanov. 2023. "Protective Properties of Marine Alkyl Glycerol Ethers in Chronic Stress" Marine Drugs 21, no. 4: 202. https://doi.org/10.3390/md21040202