

A Comparative Study of Oncolytic Vaccinia Viruses Harboring Different Marine Lectins in Breast Cancer Cells
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
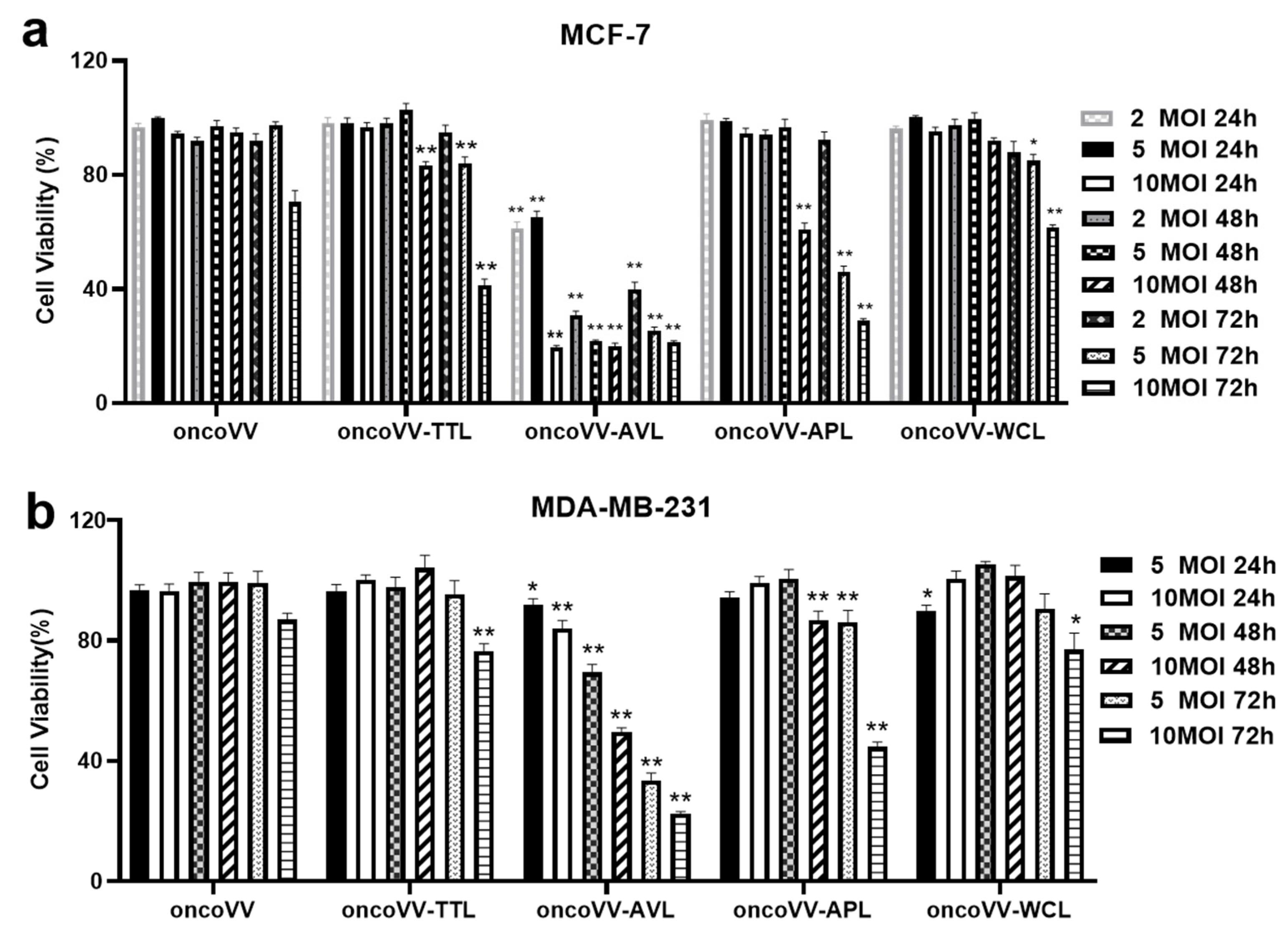
2.1. Cytotoxicity of oncoVV-AVL/APL/TTL/WCL in BC
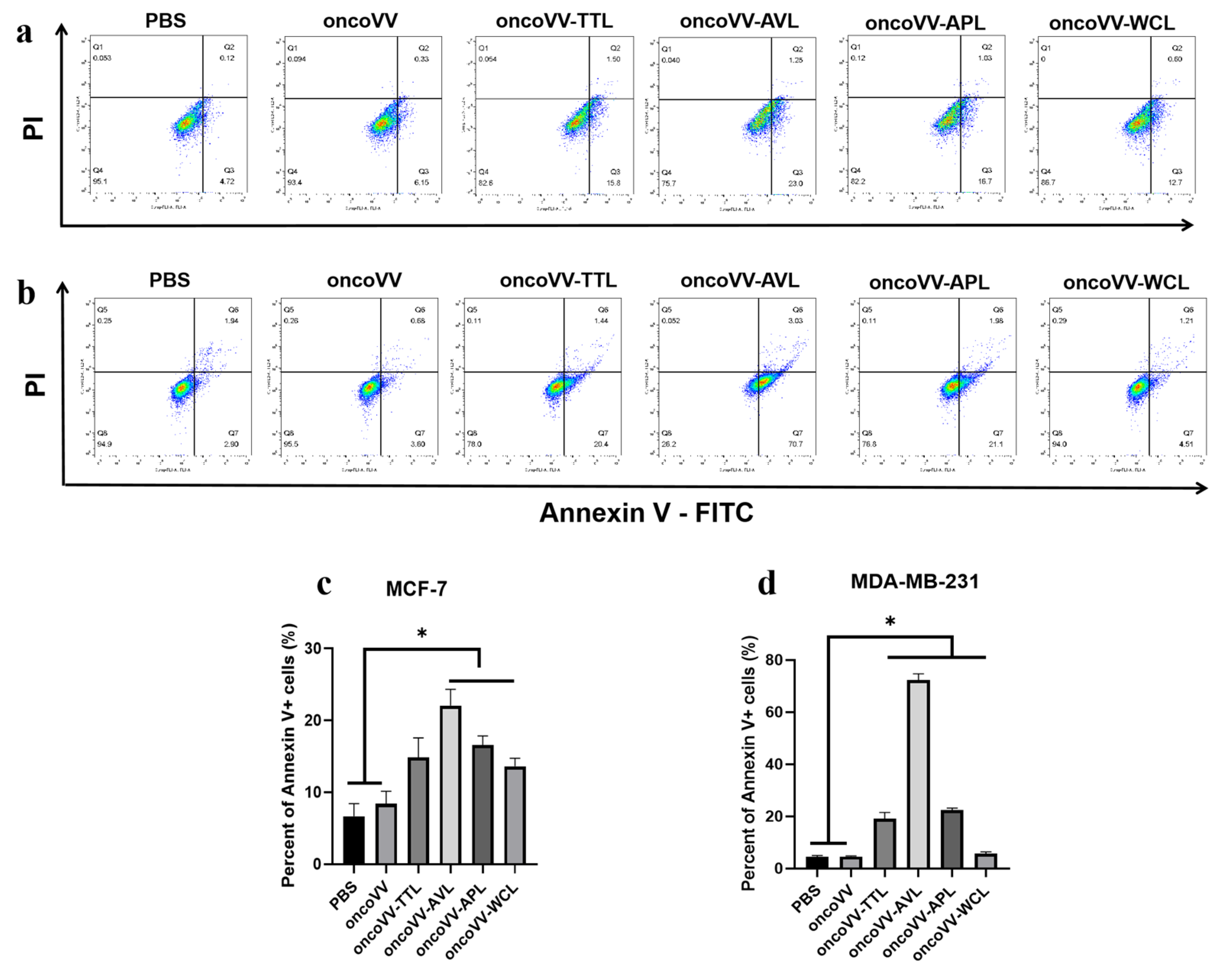
2.2. Apoptotic Effect of oncoVV-lectins in BC
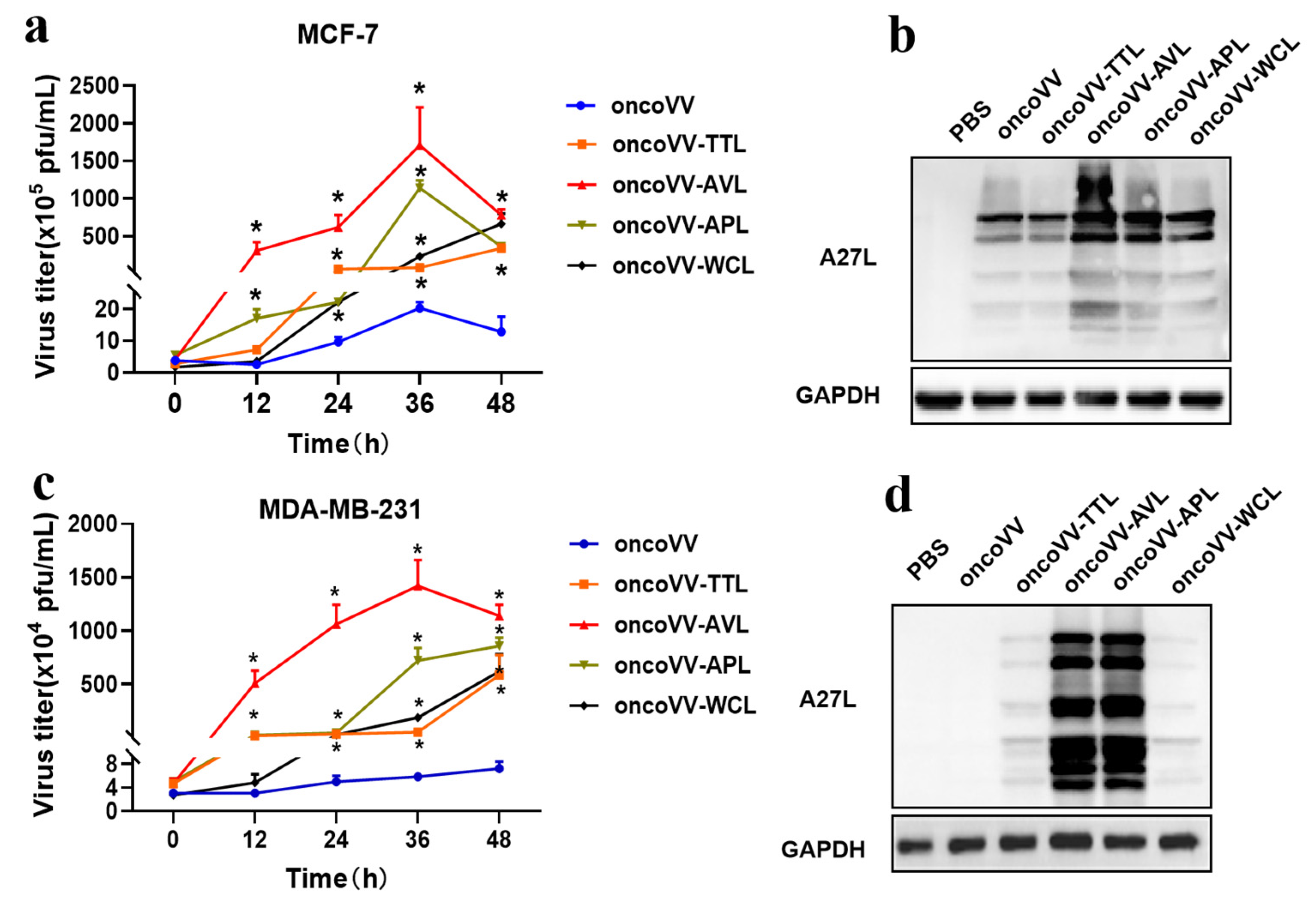
2.3. Lectins Harboring Improved the Replication of Oncolytic Vaccinia Viruses in BC
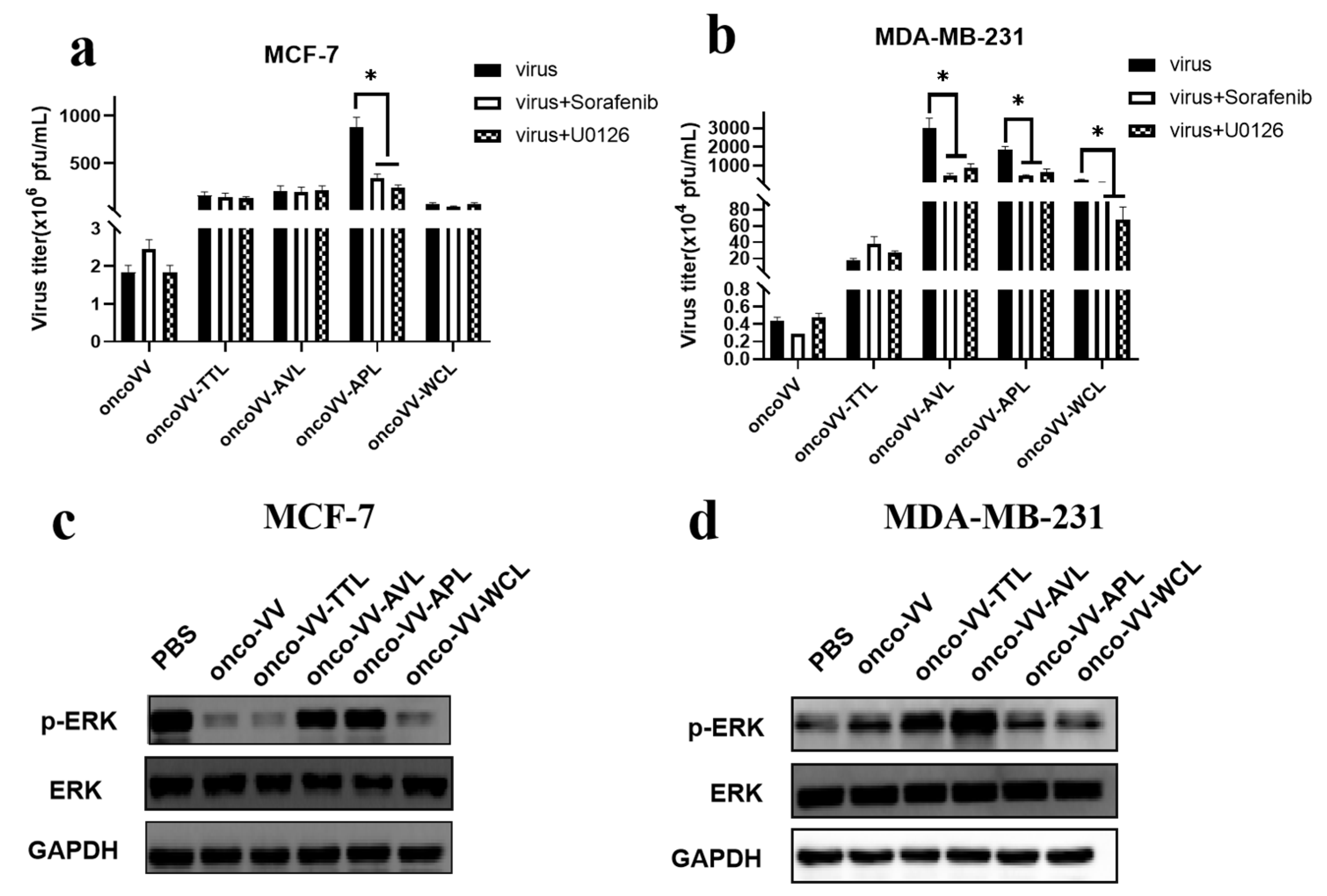
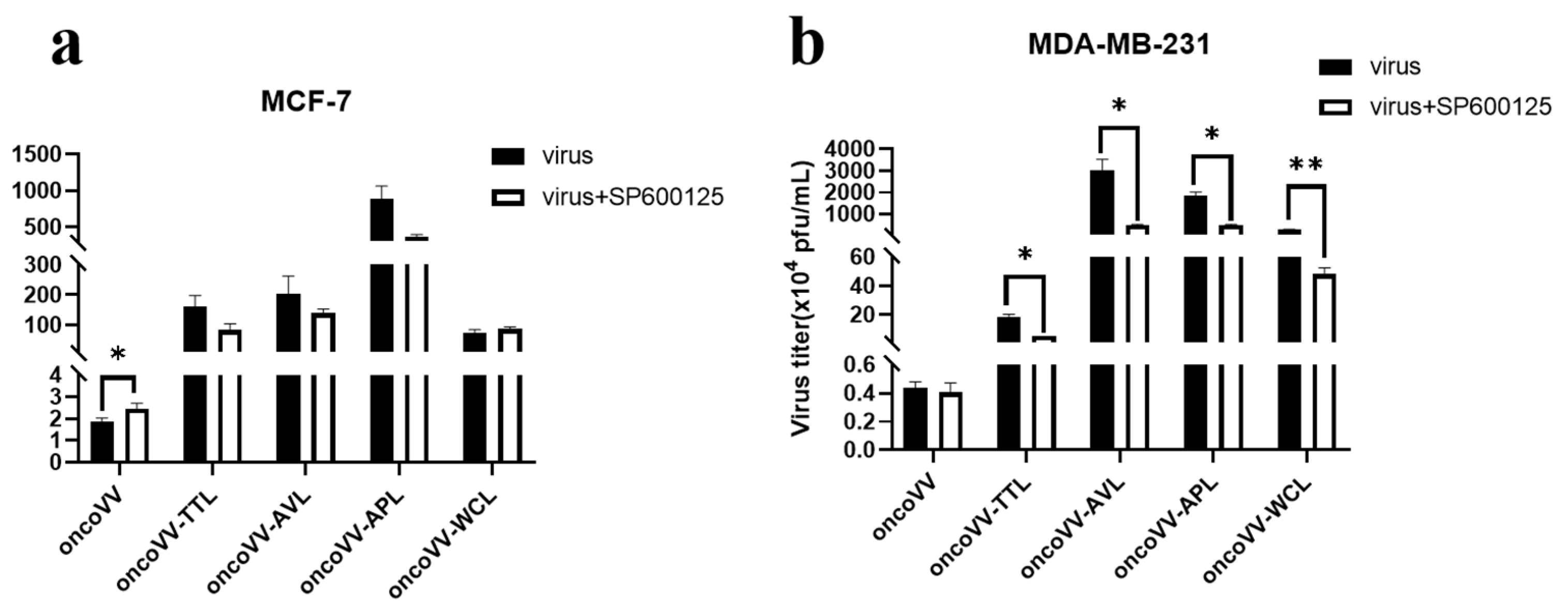
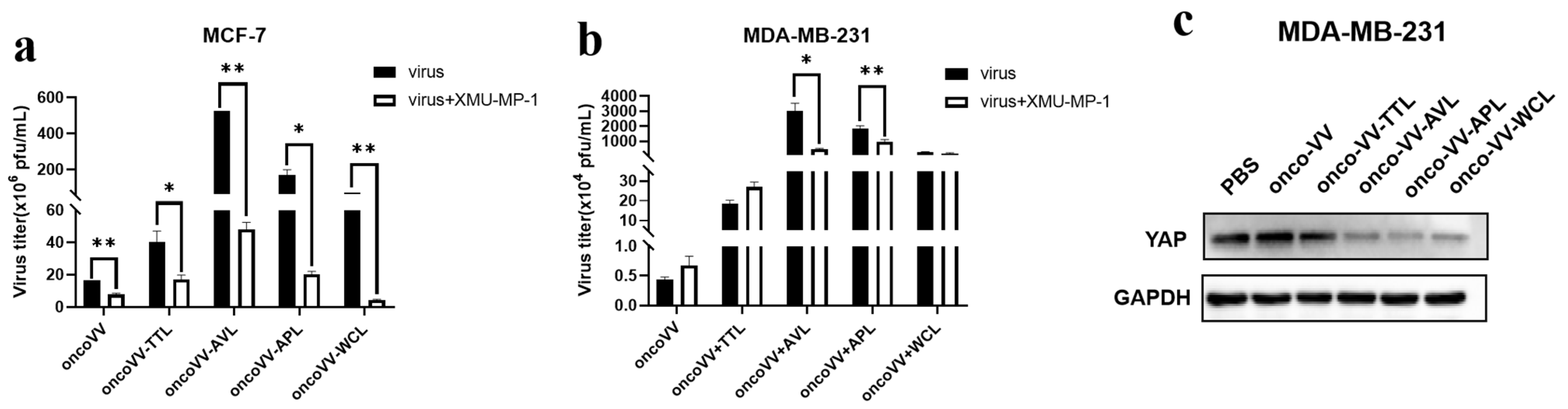
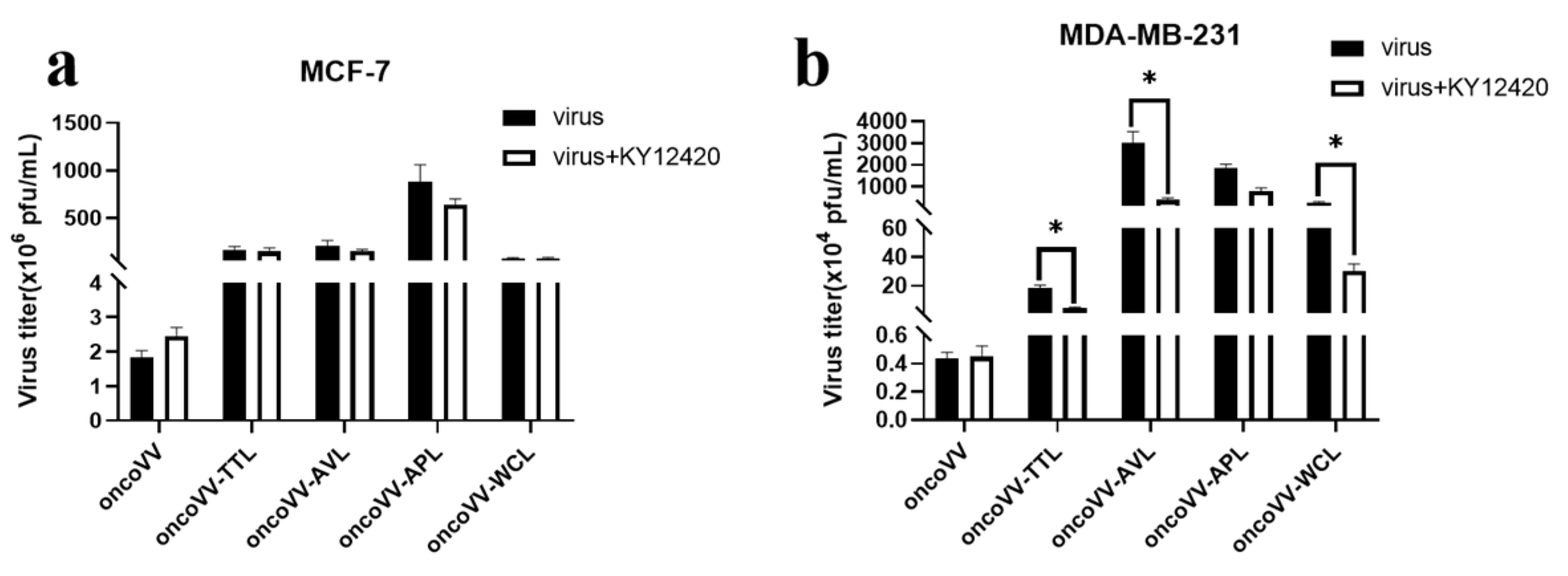
2.4. Pathways Associated with the Replication Ability of OncoVV-lectins
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Cell Culture
4.2. Virus Amplification and Crude Purification
4.3. Cell Viability Assay
4.4. Cell Apoptosis Detection
4.5. Detection of Viral Replication Ability
4.6. Western Blot Analysis
4.7. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer. 2021, 149, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Li, T.; Bai, Z.; Yang, Y.; Liu, X.; Zhan, J.; Shi, B. Breast cancer intrinsic subtype classification, clinical use and future trends. Am. J. Cancer Res. 2015, 5, 2929–2943. [Google Scholar] [PubMed]
- Yersal, O.; Barutca, S. Biological subtypes of breast cancer: Prognostic and therapeutic implications. World J. Clin. Oncol. 2014, 5, 412–424. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.J.; McGowan, P.M.; Crown, J. Targeted therapy for triple-negative breast cancer: Where are we? Int. J. Cancer 2012, 131, 2471–2477. [Google Scholar] [CrossRef] [Green Version]
- Kirn, D.H.; Thorne, S.H. Targeted and armed oncolytic poxviruses: A novel multi-mechanistic therapeutic class for cancer. Nat. Rev. Cancer 2009, 9, 64–71. [Google Scholar] [CrossRef]
- Kim, M. Replicating poxviruses for human cancer therapy. J. Microbiol. 2015, 53, 209–218. [Google Scholar] [CrossRef]
- Guse, K.; Cerullo, V.; Hemminki, A. Oncolytic vaccinia virus for the treatment of cancer. Expert. Opin. Biol. Ther. 2011, 11, 595–608. [Google Scholar] [CrossRef]
- Guo, Z.S.; Lu, B.; Guo, Z.; Giehl, E.; Feist, M.; Dai, E.; Liu, W.; Storkus, W.J.; He, Y.; Liu, Z.; et al. Vaccinia virus-mediated cancer immunotherapy: Cancer vaccines and oncolytics. J. Immunother. Cancer 2019, 7, 6. [Google Scholar] [CrossRef]
- Torres-Domínguez, L.E.; McFadden, G. Poxvirus oncolytic virotherapy. Expert. Opin. Biol. Ther. 2019, 19, 561–573. [Google Scholar] [CrossRef]
- Yang, X.; Huang, B.; Deng, L.L.; Hu, Z.G. Progress in gene therapy using oncolytic vaccinia virus as vectors. J. Cancer Res. Clin. 2018, 144, 2433–2440. [Google Scholar] [CrossRef]
- Sharon, N. Lectins: Carbohydrate-specific reagents and biological recognition molecules. J. Biol. Chem. 2007, 282, 2753–2764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues Mantuano, N.; Natoli, M.; Zippelius, A.; Läubli, H. Tumor-associated carbohydrates and immunomodulatory lectins as targets for cancer immunotherapy. J. Immunother. Cancer 2020, 8, e001222. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, S.B.; Gupta, N.; Bhagyawant, S.S.; Gaikwad, S.M. Anticancer Activity of Lectins from Bauhinia purpurea and Wisteria floribunda on Breast Cancer MCF-7 Cell Lines. Protein. Pept. Lett. 2020, 27, 870–877. [Google Scholar] [CrossRef] [PubMed]
- Tatsuta, T.; Sato, S.; Sato., T.; Sugawara, S.; Suzuki, T.; Hara, A.; Hosono, M. Sialic Acid-Binding Lectin from Bullfrog Eggs Exhibits an Anti-Tumor Effect Against Breast Cancer Cells Including Triple-Negative Phenotype Cells. Molecules 2018, 23, 2714. [Google Scholar] [CrossRef] [Green Version]
- Hasan, I.; Ozeki, Y. Histochemical localization of N-acetylhexosamine-binding lectin HOL-18 in Halichondria okadai (Japanese black sponge), and its antimicrobial and cytotoxic anticancer effects. Int. J. Biol. Macromol. 2019, 124, 819–827. [Google Scholar] [CrossRef]
- Zhu, Z.; McGray, A.J.R.; Jiang, W.; Lu, B.; Kalinski, P.; Guo, Z.S. Improving cancer immunotherapy by rationally combining oncolytic virus with modulators targeting key signaling pathways. Mol. Cancer. 2022, 21, 196. [Google Scholar] [CrossRef]
- Li, G.; Cheng, J.; Mei, S.; Wu, T.; Ye, T. Tachypleus tridentatus Lectin Enhances Oncolytic Vaccinia Virus Replication to Suppress In Vivo Hepatocellular Carcinoma Growth. Mar. Drugs. 2018, 16, 200. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Xiang, Y.; Liu, T. Oncolytic Vaccinia Virus Expressing Aphrocallistes vastus Lectin as a Cancer Therapeutic Agent. Mar. Drugs. 2019, 17, 363. [Google Scholar] [CrossRef] [Green Version]
- Ni, J.; Feng, H.; Xu, X. Oncolytic Vaccinia Virus Harboring Aphrocallistes vastus Lectin Inhibits the Growth of Cervical Cancer Cells Hela S3. Mar. Drugs. 2021, 19, 532. [Google Scholar] [CrossRef]
- Jiang, R.; Qiu, Y.; Zhang, X. Oncolytic Vaccinia Virus Harboring Aphrocallistes vastus Lectin Inhibits the Growth of Hepatocellular Carcinoma Cells. Mar. Drugs. 2022, 20, 378. [Google Scholar] [CrossRef]
- Zheng, X.; Xu, W.; Ying, Q.; Ni, J.; Jia, X.; Zhou, Y.; Ye, T.; Li, G.; Chen, K. Oncolytic Vaccinia Virus Carrying Aphrocallistes vastus Lectin (oncoVV-AVL) Enhances Inflammatory Response in Hepatocellular Carcinoma Cells. Mar. Drugs. 2022, 20, 667. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhou, N.; Liu, T. Oncolytic Vaccinia Virus Expressing White-Spotted Charr Lectin Regulates Antiviral Response in Tumor Cells and Inhibits Tumor Growth In Vitro and In Vivo. Mar. Drugs. 2021, 19, 292. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Walde, P.; Miyazaki, T.; Sakayama, K.; Nakamura, A.; Kameda, K.; Masuda, S.; Umakoshi, H.; Kato, K. Active Targeting to Osteosarcoma Cells and Apoptotic Cell Death Induction by the Novel Lectin Eucheuma serra Agglutinin Isolated from a Marine Red Alga. J. Drug Deliv. 2012, 2012, 842785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabelo, L.; Monteiro, N.; Serquiz, R.; Santos, P.; Oliveira, R.; Oliveira, A.; Rocha, H.; Morais, A.H.; Uchoa, A.; Santos, E. A lactose-binding lectin from the marine sponge Cinachyrella apion (Cal) induces cell death in human cervical adenocarcinoma cells. Mar. Drugs. 2012, 10, 727–743. [Google Scholar] [CrossRef] [Green Version]
- Vázquez, M.I.; Esteban, M. Identification of functional domains in the 14-kilodalton envelope protein (A27L) of vaccinia virus. J. Virol. 1999, 73, 9098–9109. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.L.; Vanderplasschen, A.; Law, M. The formation and function of extracellular enveloped vaccinia virus. J. Gen. Virol. 2002, 83, 2915–2931. [Google Scholar] [CrossRef]
- Vázquez, M.I.; Rivas, G.; Cregut, D.; Serrano, L.; Esteban, M. The vaccinia virus 14-kilodalton (A27L) fusion protein forms a triple coiled-coil structure and interacts with the 21-kilodalton (A17L) virus membrane protein through a C-terminal alpha-helix. J. Virol. 1998, 72, 10126–10137. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Q.L.; Zhang, S.; Tian, M.; Zhang, S.Y.; Xie, T.; Chen, D.Y.; Chen, Y.J.; He, J.; Liu, J.; Ouyang, L.; et al. Plant lectins, from ancient sugar-binding proteins to emerging anti-cancer drugs in apoptosis and autophagy. Cell. Prolif. 2015, 48, 17–28. [Google Scholar] [CrossRef]
- Kumar, R.; Khandelwal, N.; Thachamvally, R. Role of MAPK/MNK1 signaling in virus replication. Virus Res. 2018, 253, 48–61. [Google Scholar] [CrossRef]
- Wang, S.; Li, H.; Weng, S.; Li, C.; He, J. White Spot Syndrome Virus Establishes a Novel IE1/JNK/c-Jun Positive Feedback Loop to Drive Replication. Science. 2020, 23, 100752. [Google Scholar] [CrossRef]
- Garcia, G., Jr.; Paul, S.; Beshara, S. Hippo Signaling Pathway Has a Critical Role in Zika Virus Replication and in the Pathogenesis of Neuroinflammation. Am. J. Pathol. 2020, 190, 844–861. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhuo, S.; Zhuang, T.; Cho, Y.S.; Wu, G.; Liu, Y.; Mu, K.; Zhang, K.; Su, P.; Yang, Y.; et al. YAP inhibits ERα and ER+ breast cancer growth by disrupting a TEAD-ERα signaling axis. Nat. Commun. 2022, 13, 3075. [Google Scholar] [CrossRef]
- Ma, S.; Meng, Z.; Chen, R.; Guan, K.L. The Hippo Pathway: Biology and Pathophysiology. Ann. Rev. Biochem. 2019, 88, 577–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, J.A.P.; Leite, F.G.G.; Andrade, L.G.; Torres, A.A.; De Sousa, L.P.; Barcelos, L.S.; Teixeira, M.M.; Ferreira, P.C.P.; Kroon, E.G.; Souto-Padron, T. Activation of the PI3K/Akt pathway early during vaccinia and cowpox virus infections is required for both host survival and viral replication. J. Virol. 2009, 83, 6883–6899. [Google Scholar] [CrossRef] [Green Version]
- Cejalvo, J.M.; Falato, C.; Villanueva, L.; Tolosa, P.; González, X.; Pascal, M.; Canes, J.; Gavilá, J.; Manso, L.; Pascual, T.; et al. Oncolytic viruses: A new immunotherapeutic approach for breast cancer treatment? Cancer Treat. Rev. 2022, 106, 102392. [Google Scholar] [CrossRef]
- Carter, M.E.; Koch, A.; Lauer, U.M.; Hartkopf, A.D. Clinical Trials of Oncolytic Viruses in Breast Cancer. Front Oncol. 2021, 11, 803050. [Google Scholar] [CrossRef]
- Hung, L.D.; Ly, B.M.; Hao, V.T.; Trung, D.T.; Trang, V.T.D.; Trinh, P.T.H.; Ngoc, N.T.D.; Quang, T.M. Purification, characterization and biological effect of lectin from the marine sponge Stylissa flexibilis (Levi, 1961). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2018, 216, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Marques, D.N.; Almeida, A.S.; Sousa, A.R.D.; Pereira, R.; Andrade, A.L.; Chaves, R.P.; Carneiro, R.F.; Vasconcelos, M.A.; Nascimento-Neto, L.G.; Pinheiro, U. Antibacterial activity of a new lectin isolated from the marine sponge Chondrilla caribensis. Int. J. Biol. Macromol. 2018, 109, 1292–1301. [Google Scholar] [CrossRef]
- Song, R.X.; McPherson, R.A.; Adam, L.; Bao, Y.; Shupnik, M.; Kumar, R.; Santen, R.J. Linkage of rapid estrogen action to MAPK activation by ERalpha-Shc association and Shc pathway activation. Mol. Endocrinol. 2002, 16, 116–127. [Google Scholar]
- Razandi, M.; Pedram, A.; Greene, G.L.; Levin, E.R. Cell membrane and nuclear estrogen receptors (ERs) originate from a single transcript: Studies of ERalpha and ERbeta expressed in Chinese hamster ovary cells. Mol. Endocrinol. 1999, 13, 307–319. [Google Scholar] [CrossRef]
- Castoria, G.; Migliaccio, A.; Bilancio, A.; Di Domenico, M.; de Falco, A.; Lombardi, M.; Fiorentino, R.; Varricchio, L.; Barone, M.V.; Auricchio, F. PI3-kinase in concert with Src promotes the S-phase entry of oestradiol-stimulated MCF-7 cells. EMBO J. 2001, 20, 6050–6059. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Muñoz, E.; Hernández-Hernández, O.T.; Camacho-Arroyo, I. Role of estradiol and progesterone in HIV susceptibility and disease progression. Mini Rev. Med. Chem. 2012, 12, 1049–1054. [Google Scholar] [CrossRef] [PubMed]
- Lasso, G.; Mayer, S.V.; Winkelmann, E.R.; Chu, T.; Elliot, O.; Patino-Galindo, J.A.; Park, K.; Rabadan, R.; Honig, B.; Shapira, S.D. A Structure-Informed Atlas of Human-Virus Interactions. Cell 2019, 178, 1526–1541. [Google Scholar] [CrossRef]
- LaBarre, D.D.; Lowy, R.J. Improvements in methods for calculating virus titer estimates from TCID50 and plaque assays. J. Virol. Methods. 2001, 96, 107–126. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Wang, Q.; Ying, Q.; Zhang, X.; Ye, T.; Chen, K.; Li, G. A Comparative Study of Oncolytic Vaccinia Viruses Harboring Different Marine Lectins in Breast Cancer Cells. Mar. Drugs 2023, 21, 77. https://doi.org/10.3390/md21020077
Zhou Y, Wang Q, Ying Q, Zhang X, Ye T, Chen K, Li G. A Comparative Study of Oncolytic Vaccinia Viruses Harboring Different Marine Lectins in Breast Cancer Cells. Marine Drugs. 2023; 21(2):77. https://doi.org/10.3390/md21020077
Chicago/Turabian StyleZhou, Yanrong, Qianpeng Wang, Qi Ying, Xiaomei Zhang, Ting Ye, Kan Chen, and Gongchu Li. 2023. "A Comparative Study of Oncolytic Vaccinia Viruses Harboring Different Marine Lectins in Breast Cancer Cells" Marine Drugs 21, no. 2: 77. https://doi.org/10.3390/md21020077