

Novel BRICHOS-Related Antimicrobial Peptides from the Marine Worm Heteromastus filiformis: Transcriptome Mining, Synthesis, Biological Activities, and Therapeutic Potential
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. The Marine Polychaete H. filiformis Is a Rich Natural Source of BRICHOS-Related Peptides
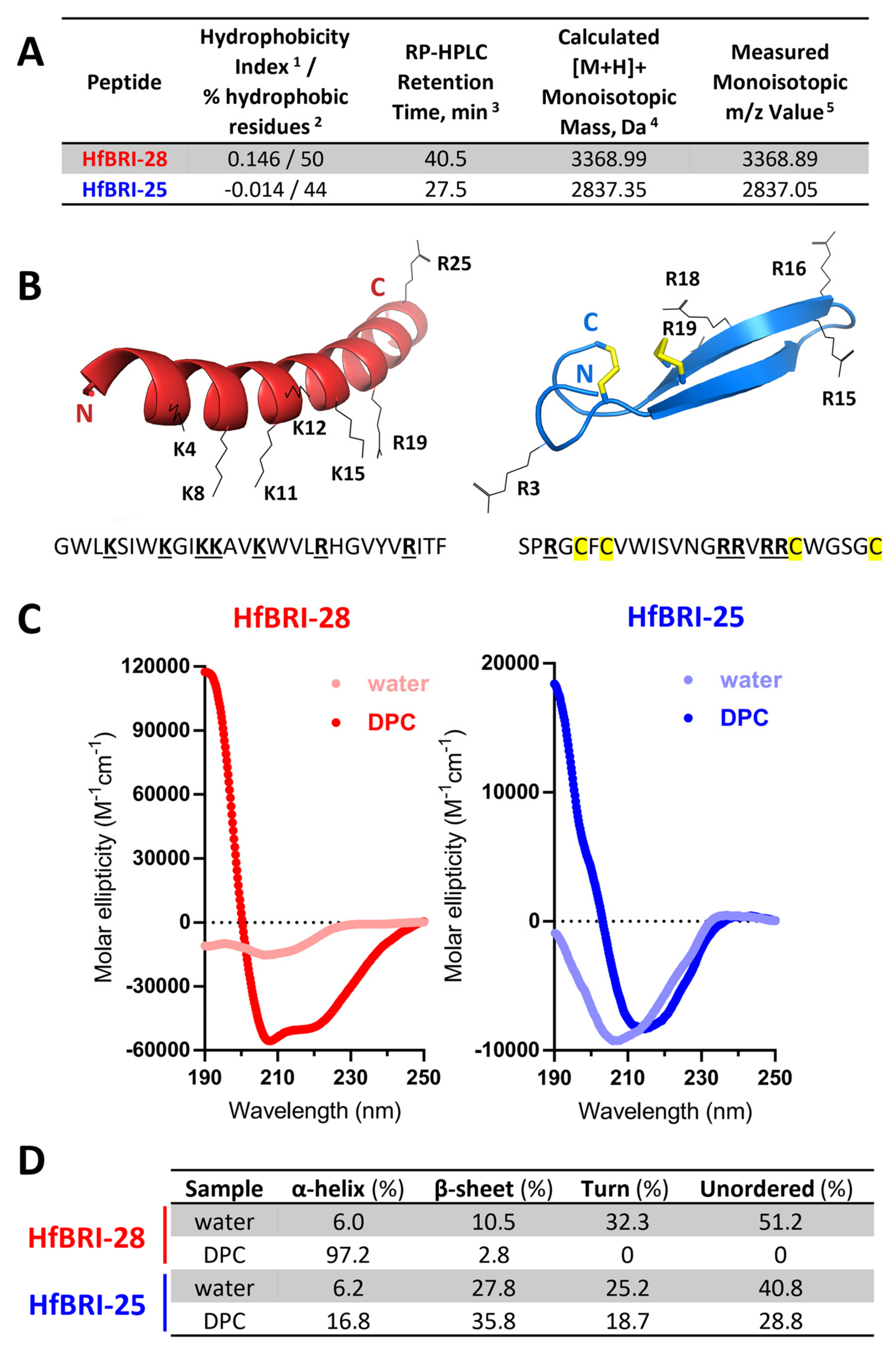
2.2. Production and Structural Analysis of Novel Peptides HfBRI-25 and HfBRI-28

2.3. HfBRI-25 Acts Selectively against Bacterial Cells
2.4. HfBRI-25 and HfBRI-28 Kill Bacterial Cells via Membrane Damage
2.5. In Vivo Efficiency and Stability of HfBRI-25 in Serum
2.6. HfBRI-25 Effectively Kills Uropathogenic E. coli Strains and Embedded Cells inside Bacterial Biofilms
3. Materials and Methods
3.1. Transcriptome Assembly and Prepropeptides Identification
3.2. Recombinant Production of HfBRI-25
3.3. Chemical Synthesis of HfBRI-28
3.4. Analytical Reversed-Phase HPLC
3.5. Identification of Disulfide Pairing
3.6. Circular Dichroism Spectroscopy
3.7. Antimicrobial Assay
3.8. Hemolysis and Cytotoxicity Assay
3.9. Assessment of Bacterial Membrane Permeabilization
3.10. Resistance Induction Assay
3.11. Stability in Serum
3.12. The Effect of Peptides on Established Biofilm
3.13. Animal Studies
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bruno, R.; Maresca, M.; Canaan, S.; Cavalier, J.-F.; Mabrouk, K.; Boidin-Wichlacz, C.; Olleik, H.; Zeppilli, D.; Brodin, P.; Massol, F.; et al. Worms’ Antimicrobial Peptides. Mar. Drugs 2019, 17, 512. [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikova, T.V.; Aleshina, G.M.; Balandin, S.V.; Krasnosdembskaya, A.D.; Markelov, M.L.; Frolova, E.I.; Leonova, Y.F.; Tagaev, A.A.; Krasnodembsky, E.G.; Kokryakov, V.N. Purification and Primary Structure of Two Isoforms of Arenicin, a Novel Antimicrobial Peptide from Marine Polychaeta Arenicola Marina. FEBS Lett. 2004, 577, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Presto, J.; Johansson, J. The BRICHOS Domain: Its Proproteins and Functions. In The BRICHOS Domain; Briefs in Molecular Science; Springer International Publishing: Cham, Switzerland, 2015; pp. 1–28. ISBN 978-3-319-16563-9. [Google Scholar]
- Ovchinnikova, T.V.; Shenkarev, Z.O.; Nadezhdin, K.D.; Balandin, S.V.; Zhmak, M.N.; Kudelina, I.A.; Finkina, E.I.; Kokryakov, V.N.; Arseniev, A.S. Recombinant Expression, Synthesis, Purification, and Solution Structure of Arenicin. Biochem. Biophys. Res. Commun. 2007, 360, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Safronova, V.N.; Bolosov, I.A.; Kruglikov, R.N.; Korobova, O.V.; Pereskokova, E.S.; Borzilov, A.I.; Panteleev, P.V.; Ovchinnikova, T.V. Novel β-Hairpin Peptide from Marine Polychaeta with a High Efficacy against Gram-Negative Pathogens. Mar. Drugs 2022, 20, 517. [Google Scholar] [CrossRef] [PubMed]
- Tasiemski, A.; Jung, S.; Boidin-Wichlacz, C.; Jollivet, D.; Cuvillier-Hot, V.; Pradillon, F.; Vetriani, C.; Hecht, O.; Sönnichsen, F.D.; Gelhaus, C.; et al. Characterization and Function of the First Antibiotic Isolated from a Vent Organism: The Extremophile Metazoan Alvinella Pompejana. PLoS ONE 2014, 9, e95737. [Google Scholar] [CrossRef] [PubMed]
- Papot, C.; Massol, F.; Jollivet, D.; Tasiemski, A. Antagonistic Evolution of an Antibiotic and Its Molecular Chaperone: How to Maintain a Vital Ectosymbiosis in a Highly Fluctuating Habitat. Sci. Rep. 2017, 7, 1454. [Google Scholar] [CrossRef]
- Panteleev, P.V.; Tsarev, A.V.; Safronova, V.N.; Reznikova, O.V.; Bolosov, I.A.; Sychev, S.V.; Shenkarev, Z.O.; Ovchinnikova, T.V. Structure Elucidation and Functional Studies of a Novel β-Hairpin Antimicrobial Peptide from the Marine Polychaeta Capitella Teleta. Mar. Drugs 2020, 18, 620. [Google Scholar] [CrossRef]
- Panteleev, P.V.; Tsarev, A.V.; Bolosov, I.A.; Paramonov, A.S.; Marggraf, M.B.; Sychev, S.V.; Shenkarev, Z.O.; Ovchinnikova, T.V. Novel Antimicrobial Peptides from the Arctic Polychaeta Nicomache Minor Provide New Molecular Insight into Biological Role of the BRICHOS Domain. Mar. Drugs 2018, 16, 401. [Google Scholar] [CrossRef]
- Bruno, R.; Boidin-Wichlacz, C.; Melnyk, O.; Zeppilli, D.; Landon, C.; Thomas, F.; Cambon, M.-A.; Lafond, M.; Mabrouk, K.; Massol, F.; et al. The Diversification of the Antimicrobial Peptides from Marine Worms Is Driven by Environmental Conditions. Sci. Total Environ. 2023, 879, 162875. [Google Scholar] [CrossRef]
- Stanovova, M.V.; Gazizova, G.R.; Gorbushin, A.M. Transcriptomic Profiling of Immune-associated Molecules in the Coelomocytes of Lugworm Arenicola Marina (Linnaeus, 1758). J. Exp. Zool. Part B Mol. Dev. Evol. 2023, 340, 34–55. [Google Scholar] [CrossRef]
- Jeong, M.-K.; Soh, H.Y.; Suh, H.-L. Three New Species of Heteromastus (Annelida, Capitellidae) from Korean Waters, with Genetic Evidence Based on Two Gene Markers. ZooKeys 2019, 869, 1–18. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD3: The Antimicrobial Peptide Database as a Tool for Research and Education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Sun, J.; Zhou, M.; Zhou, J.; Lao, X.; Zheng, H.; Xu, H. DRAMP: A Comprehensive Data Repository of Antimicrobial Peptides. Sci. Rep. 2016, 6, 24482. [Google Scholar] [CrossRef] [PubMed]
- Dong, N.; Ma, Q.; Shan, A.; Lv, Y.; Hu, W.; Gu, Y.; Li, Y. Strand Length-Dependent Antimicrobial Activity and Membrane-Active Mechanism of Arginine- and Valine-Rich β-Hairpin-Like Antimicrobial Peptides. Antimicrob. Agents Chemother. 2012, 56, 2994–3003. [Google Scholar] [CrossRef]
- Franchi, N.; Ballarin, L.; Cima, F. Botryllin, a Novel Antimicrobial Peptide from the Colonial Ascidian Botryllus Schlosseri. Mar. Drugs 2023, 21, 74. [Google Scholar] [CrossRef] [PubMed]
- Safronova, V.N.; Panteleev, P.V.; Sukhanov, S.V.; Toropygin, I.Y.; Bolosov, I.A.; Ovchinnikova, T.V. Mechanism of Action and Therapeutic Potential of the β-Hairpin Antimicrobial Peptide Capitellacin from the Marine Polychaeta Capitella Teleta. Mar. Drugs 2022, 20, 167. [Google Scholar] [CrossRef]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent Updates, New Developments and Status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. [Google Scholar] [CrossRef]
- Panteleev, P.V.; Ovchinnikova, T.V. Improved Strategy for Recombinant Production and Purification of Antimicrobial Peptide Tachyplesin I and Its Analogs with High Cell Selectivity. Biotechnol. Appl. Biochem. 2017, 64, 35–42. [Google Scholar] [CrossRef]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making Protein Folding Accessible to All. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef] [PubMed]
- Provencher, S.W.; Glöckner, J. Estimation of Globular Protein Secondary Structure from Circular Dichroism. Biochemistry 1981, 20, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, G.; Kumar, T.K.; Arunkumar, A.I.; Yu, C. 2,2,2-Trifluoroethanol Induces Helical Conformation in an All Beta-Sheet Protein. Biochem. Biophys. Res. Commun. 1996, 222, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Woody, R.W. Circular Dichroism. Methods Enzym. 1995, 246, 34–71. [Google Scholar] [CrossRef]
- Perczel, A.; Hollosi, M.; Foxman, B.M.; Fasman, G.D. Conformational Analysis of Pseudocyclic Hexapeptides Based on Quantitative Circular Dichroism (CD), NOE, and x-Ray Data. The Pure CD Spectra of Type I and Type II.Beta.-Turns. J. Am. Chem. Soc. 1991, 113, 9772–9784. [Google Scholar] [CrossRef]
- Bolosov, I.A.; Panteleev, P.V.; Sychev, S.V.; Khokhlova, V.A.; Safronova, V.N.; Toropygin, I.Y.; Kombarova, T.I.; Korobova, O.V.; Pereskokova, E.S.; Borzilov, A.I.; et al. Design of Protegrin-1 Analogs with Improved Antibacterial Selectivity. Pharmaceutics 2023, 15, 2047. [Google Scholar] [CrossRef] [PubMed]
- Kuzmin, D.V.; Emelianova, A.A.; Kalashnikova, M.B.; Panteleev, P.V.; Balandin, S.V.; Serebrovskaya, E.O.; Belogurova-Ovchinnikova, O.Y.; Ovchinnikova, T.V. Comparative in Vitro Study on Cytotoxicity of Recombinant β-Hairpin Peptides. Chem. Biol. Drug Des. 2018, 91, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Panteleev, P.V.; Bolosov, I.A.; Ovchinnikova, T.V. Bioengineering and Functional Characterization of Arenicin Shortened Analogs with Enhanced Antibacterial Activity and Cell Selectivity: Bioengineering of Arenicin Shortened Analogs with Enhanced Selectivity. J. Pept. Sci. 2016, 22, 82–91. [Google Scholar] [CrossRef]
- Zhang, X.-H.; He, X.; Austin, B. Vibrio Harveyi: A Serious Pathogen of Fish and Invertebrates in Mariculture. Mar. Life Sci. Technol. 2020, 2, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, G.S.; Costa, R.P.; Gomes, P.; Gomes, M.S.; Silva, T.; Teixeira, C. Antimicrobial Peptides as Potential Anti-Tubercular Leads: A Concise Review. Pharmaceuticals 2021, 14, 323. [Google Scholar] [CrossRef] [PubMed]
- Panteleev, P.V.; Safronova, V.N.; Kruglikov, R.N.; Bolosov, I.A.; Bogdanov, I.V.; Ovchinnikova, T.V. A Novel Proline-Rich Cathelicidin from the Alpaca Vicugna Pacos with Potency to Combat Antibiotic-Resistant Bacteria: Mechanism of Action and the Functional Role of the C-Terminal Region. Membranes 2022, 12, 515. [Google Scholar] [CrossRef] [PubMed]
- Gajdács, M.; Ábrók, M.; Lázár, A.; Burián, K. Comparative Epidemiology and Resistance Trends of Common Urinary Pathogens in a Tertiary-Care Hospital: A 10-Year Surveillance Study. Medicina 2019, 55, 356. [Google Scholar] [CrossRef]
- Slukin, P.V.; Svetoch, E.A.; Aslanyan, E.M.; Astashkin, E.I.; Ershova, M.G.; Poletaeva, E.D.; Shepelin, A.P.; Fursova, N.K. Phenotypic and molecular genetic properties of Escherichia coli clinical strains isolated from patients with urological diseases. Urologiia 2020, 2, 23–30. [Google Scholar] [CrossRef]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of Plasmid-Mediated Colistin Resistance Mechanism MCR-1 in Animals and Human Beings in China: A Microbiological and Molecular Biological Study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Gastaldi Guerrieri, C.; Teixeira Gonçalves, M.; Ferreira Da Silva, A.; Souza Dos Santos, A.L.; Dos Santos, K.V.; Cruz Spano, L. Remarkable Antibiofilm Activity of Ciprofloxacin, Cefoxitin, and Tobramycin, by Themselves or in Combination, against Enteroaggregative Escherichia Coli in Vitro. Diagn. Microbiol. Infect. Dis. 2023, 107, 116048. [Google Scholar] [CrossRef] [PubMed]
- de Sena Brandine, G.; Smith, A.D. Falco: High-Speed FastQC Emulation for Quality Control of Sequencing Data. F1000Res 2021, 8, 1874. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and Memory-Efficient Alignment of Short DNA Sequences to the Human Genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [PubMed]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER Web Server: 2018 Update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef]
- Panteleev, P.V.; Bolosov, I.A.; Kalashnikov, A.À.; Kokryakov, V.N.; Shamova, O.V.; Emelianova, A.A.; Balandin, S.V.; Ovchinnikova, T.V. Combined Antibacterial Effects of Goat Cathelicidins With Different Mechanisms of Action. Front. Microbiol. 2018, 9, 2983. [Google Scholar] [CrossRef]
- Andrushchenko, V.V.; Vogel, H.J.; Prenner, E.J. Optimization of the Hydrochloric Acid Concentration Used for Trifluoroacetate Removal from Synthetic Peptides. J. Pept. Sci. 2007, 13, 37–43. [Google Scholar] [CrossRef]
- Barlos, K.; Chatzi, O.; Gatos, D.; Stavropoulos, G. 2-Chlorotrityl Chloride Resin. Studies on Anchoring of Fmoc-Amino Acids and Peptide Cleavage. Int. J. Pept. Protein Res. 1991, 37, 513–520. [Google Scholar]
- Knorr, R.; Trzeciak, A.; Bannwarth, W.; Gillessen, D. New Coupling Reagents in Peptide Chemistry. Tetrahedron Lett. 1989, 30, 1927–1930. [Google Scholar] [CrossRef]
- Subirós-Funosas, R.; Prohens, R.; Barbas, R.; El-Faham, A.; Albericio, F. Oxyma: An Efficient Additive for Peptide Synthesis to Replace the Benzotriazole-Based HOBt and HOAt with a Lower Risk of Explosion. Chem. A Eur. J. 2009, 15, 9394–9403. [Google Scholar] [CrossRef]
- Carpino, L.A.; Han, G.Y. 9-Fluorenylmethoxycarbonyl Function, a New Base-Sensitive Amino-Protecting Group. J. Am. Chem. Soc. 1970, 92, 5748–5749. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, A.; De La Torre, B.G.; Albericio, F. In Situ Fmoc Removal—A Sustainable Solid-Phase Peptide Synthesis Approach. Green. Chem. 2022, 24, 4887–4896. [Google Scholar] [CrossRef]
- King, D.S.; Fields, C.G.; Fields, G.B. A Cleavage Method Which Minimizes Side Reactions Following Fmoc Solid Phase Peptide Synthesis. Int. J. Pept. Protein Res. 2009, 36, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Panteleev, P.V.; Bolosov, I.A.; Balandin, S.V.; Ovchinnikova, T.V. Design of Antimicrobial Peptide Arenicin Analogs with Improved Therapeutic Indices. J. Pept. Sci. 2015, 21, 105–113. [Google Scholar] [CrossRef]
- Zanetti, M.; Gennaro, R.; Romeo, D. Cathelicidins: A Novel Protein Family with a Common Proregion and a Variable C-Terminal Antimicrobial Domain. FEBS Lett. 1995, 374, 1–5. [Google Scholar] [CrossRef]
- Zanetti, M.; Gennaro, R.; Scocchi, M.; Skerlavaj, B. Structure and Biology of Cathelicidins. Adv. Exp. Med. Biol. 2000, 479, 203–218. [Google Scholar] [CrossRef]
- Muratspahić, E.; Aslanoglou, D.; White, A.M.; Draxler, C.; Kozisek, X.; Farooq, Z.; Craik, D.J.; McCormick, P.J.; Durek, T.; Gruber, C.W. Development of Melanocortin 4 Receptor Agonists by Exploiting Animal-Derived Macrocyclic, Disulfide-Rich Peptide Scaffolds. ACS Pharmacol. Transl. Sci. 2023, 6, 1373–1381. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | Minimum Inhibitory Concentration (μM) | |||
|---|---|---|---|---|
| HfBRI-25 | HfBRI-28 | |||
| MHB | MHB + 0.9% NaCl | MHB | MHB + 0.9% NaCl | |
| Gram-negative | ||||
| Escherichia coli ML-35p | 0.25 | 1 | 2 | 2 |
| Escherichia coli ATCC 25922 | 0.5 | 1 | 2 | 1 |
| Acinetobacter baumannii XDR CI 2675 | 0.125 | 1 | 2 | 2 |
| Enterobacter cloacae XDR CI 4172 | 1 | 16 | 0.5 | 2 |
| Klebsiella pneumonia ATCC 700603 | 2 | 32 | 16 | 16 |
| Pseudomonas aeruginosa ATCC 27853 | 2 | >32 | 32 | >32 |
| Vibrio harveyi BB120 | 1 * | 0.06 * | ||
| Gram-positive | ||||
| Staphylococcus aureus ATCC 6538P | 2 | >16 | 8 | 8 |
| Staphylococcus aureus ATCC 29213 | n.d. | >16 | n.d. | >32 |
| Bacillus mycoides VKM B-814 | 0.5 | 32 | 0.5 | 2 |
| Bacillus subtilis VKM B-886 | 1 | 4 | 1 | 0.5 |
| Bacillus licheniformis VKM B-511 | 1 | 16 | 8 | 4 |
| Mycobacterium phlei Ac-1291 | 2 | 16 | 2 | 1 |
| Mycobacterium smegmatis mc(2)155 | 1 ** | 1 ** | ||
| Bacteria | Minimum Inhibitory Concentration (μM) | ||
|---|---|---|---|
| MHB | MHB + 0.9% NaCl | MHB + 0.9% NaCl + 5% Serum | |
| E. coli ML-35p | 0.25 | 1 | 1 |
| E. coli ATCC 25922 | 0.5 | 1 | 2 |
| E. coli ATCC 23724 | 0.5 | 1 | 2 |
| E. coli K12 BW25113 | 0.125 | 1 | 2 |
| E. coli K12 SBS 1936 | 0.5 | 2 | 2 |
| E. coli CI 214 | 0.25 | 1 | 1 |
| E. coli XDR CI 3600 | 0.5 | 2 | 2 |
| E. coli MDR 1057 | 0.5 | 1 | 1 |
| E. coli MDR 1057 (PmxB-res) * | 1 | 2 | 2 |
| E. coli XDR U10 (mcr-1+) | 0.125 | 1 | 1 |
| E. coli XDR U22 (mcr-1+) | 0.125 | 1 | 1 |
| Bacterial Strain | Characteristics (Source, Antibiotic Resistance) |
|---|---|
| Escherichia coli DH10B | Cloning strain (Invitrogen) |
| Escherichia coli BL21 (DE3) | Expression strain (Novagen) |
| Escherichia coli ClearColi BL21(DE3) | Expression strain (Research Corporation Technologies) |
| Escherichia coli ML-35p | Laboratory strain (ATCC collection) |
| Escherichia coli ATCC 25922 | Laboratory strain (ATCC collection) |
| Escherichia coli ATCC 23724 | Laboratory strain (ATCC collection) |
| Escherichia coli K12 BW25113 | Laboratory strain (Keio collection) |
| Escherichia coli K12 SBS 1936 | Laboratory strain (IMG RAS collection) |
| Escherichia coli U10 SRCAMB B-8551 | XDR clinical isolate (urine, kidney stone disease; ESBL+; mcr-1+) |
| Escherichia coli U22 SRCAMB B-8553 | XDR clinical isolate (urine, UTI; ESBL+; mcr-1+) |
| Escherichia coli 1057 SRCAMB B-10910 | MDR clinical isolate (urine, UTI; ESBL+) |
| Escherichia coli (MDR CI 3600) | MDR clinical isolate (urine, UTI; ESBL+) |
| Escherichia coli (CI 214) | Clinical isolate (urine, acute pyelonephritis) |
| Enterobacter cloacae (XDR CI 4172) | XDR clinical isolate (MBL+) |
| Acinetobacter baumanii (XDR CI 2675) | XDR clinical isolate (MBL+) |
| Klebsiella pneumoniae ATCC 700603 | Laboratory strain (ATCC collection) |
| Pseudomonas aeruginosa ATCC 27853 | Laboratory strain (ATCC collection) |
| Vibrio harveyi BB120 | Laboratory strain (ATCC collection) |
| Bacillus subtilis B-886 | Laboratory strain (VKM collection) |
| Bacillus licheniformis VKM B511 | Laboratory strain (VKM collection) |
| Bacillus mycoides VKM | Laboratory strain (VKM collection) |
| Staphylococcus aureus ATCC 29213 | Laboratory strain (ATCC collection) |
| Staphylococcus aureus ATCC 6538P | Laboratory strain (ATCC collection) |
| Mycobacterium smegmatis mc(2)155 | Laboratory strain (ATCC collection) |
| Mycobacterium phlei Ac-1291 | Laboratory strain (VKM collection) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panteleev, P.V.; Safronova, V.N.; Duan, S.; Komlev, A.S.; Bolosov, I.A.; Kruglikov, R.N.; Kombarova, T.I.; Korobova, O.V.; Pereskokova, E.S.; Borzilov, A.I.; et al. Novel BRICHOS-Related Antimicrobial Peptides from the Marine Worm Heteromastus filiformis: Transcriptome Mining, Synthesis, Biological Activities, and Therapeutic Potential. Mar. Drugs 2023, 21, 639. https://doi.org/10.3390/md21120639
Panteleev PV, Safronova VN, Duan S, Komlev AS, Bolosov IA, Kruglikov RN, Kombarova TI, Korobova OV, Pereskokova ES, Borzilov AI, et al. Novel BRICHOS-Related Antimicrobial Peptides from the Marine Worm Heteromastus filiformis: Transcriptome Mining, Synthesis, Biological Activities, and Therapeutic Potential. Marine Drugs. 2023; 21(12):639. https://doi.org/10.3390/md21120639
Chicago/Turabian StylePanteleev, Pavel V., Victoria N. Safronova, Shuting Duan, Alexey S. Komlev, Ilia A. Bolosov, Roman N. Kruglikov, Tatiana I. Kombarova, Olga V. Korobova, Eugenia S. Pereskokova, Alexander I. Borzilov, and et al. 2023. "Novel BRICHOS-Related Antimicrobial Peptides from the Marine Worm Heteromastus filiformis: Transcriptome Mining, Synthesis, Biological Activities, and Therapeutic Potential" Marine Drugs 21, no. 12: 639. https://doi.org/10.3390/md21120639