

Characterization and Expression Analysis of Regeneration-Associated Protein (Aj-Orpin) during Intestinal Regeneration in the Sea Cucumber Apostichopus japonicus
Abstract
:1. Introduction
2. Results
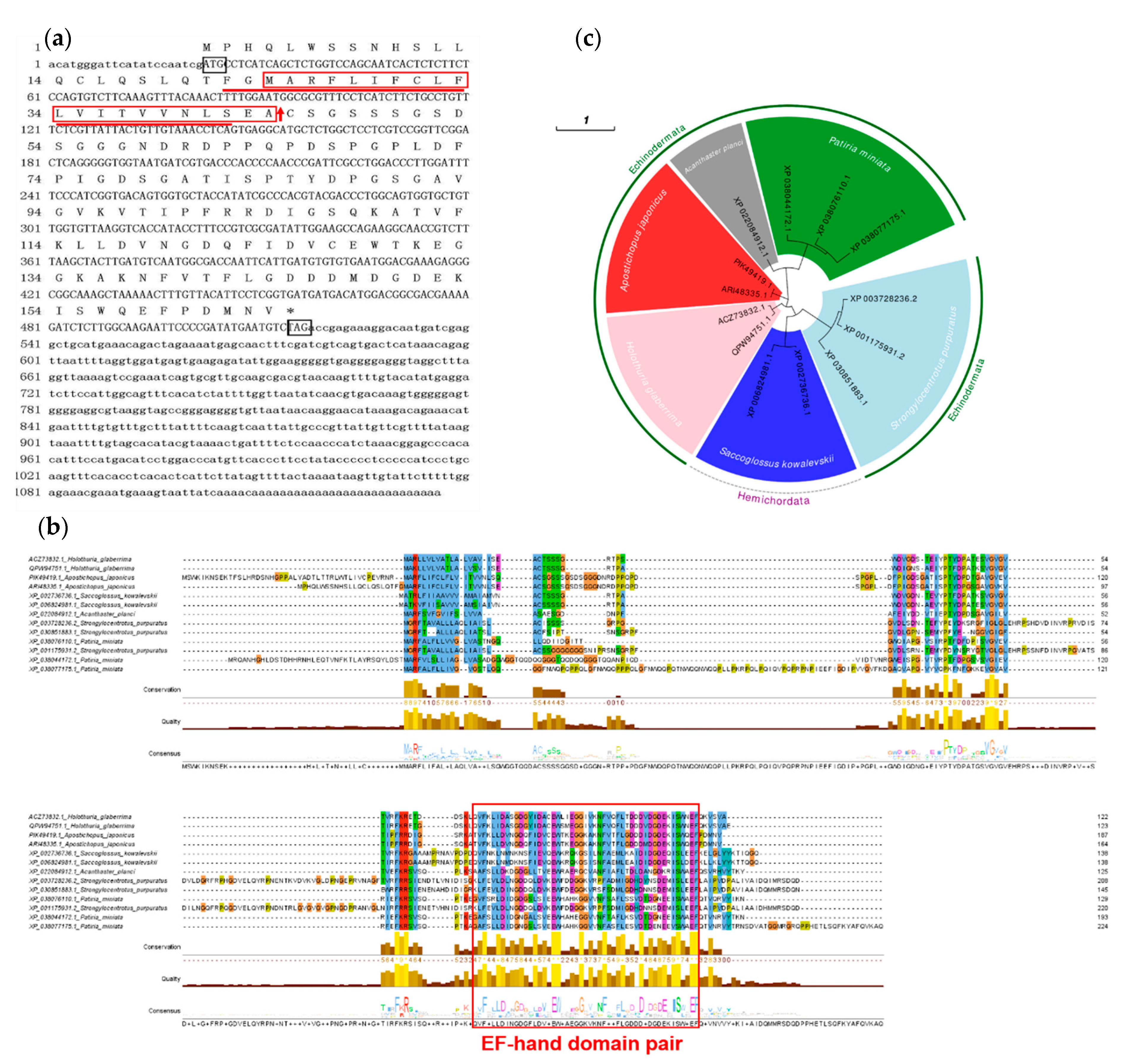
2.1. Cloning and Sequence Analysis of Aj-Orpin Full-Length cDNA in A. japonicus
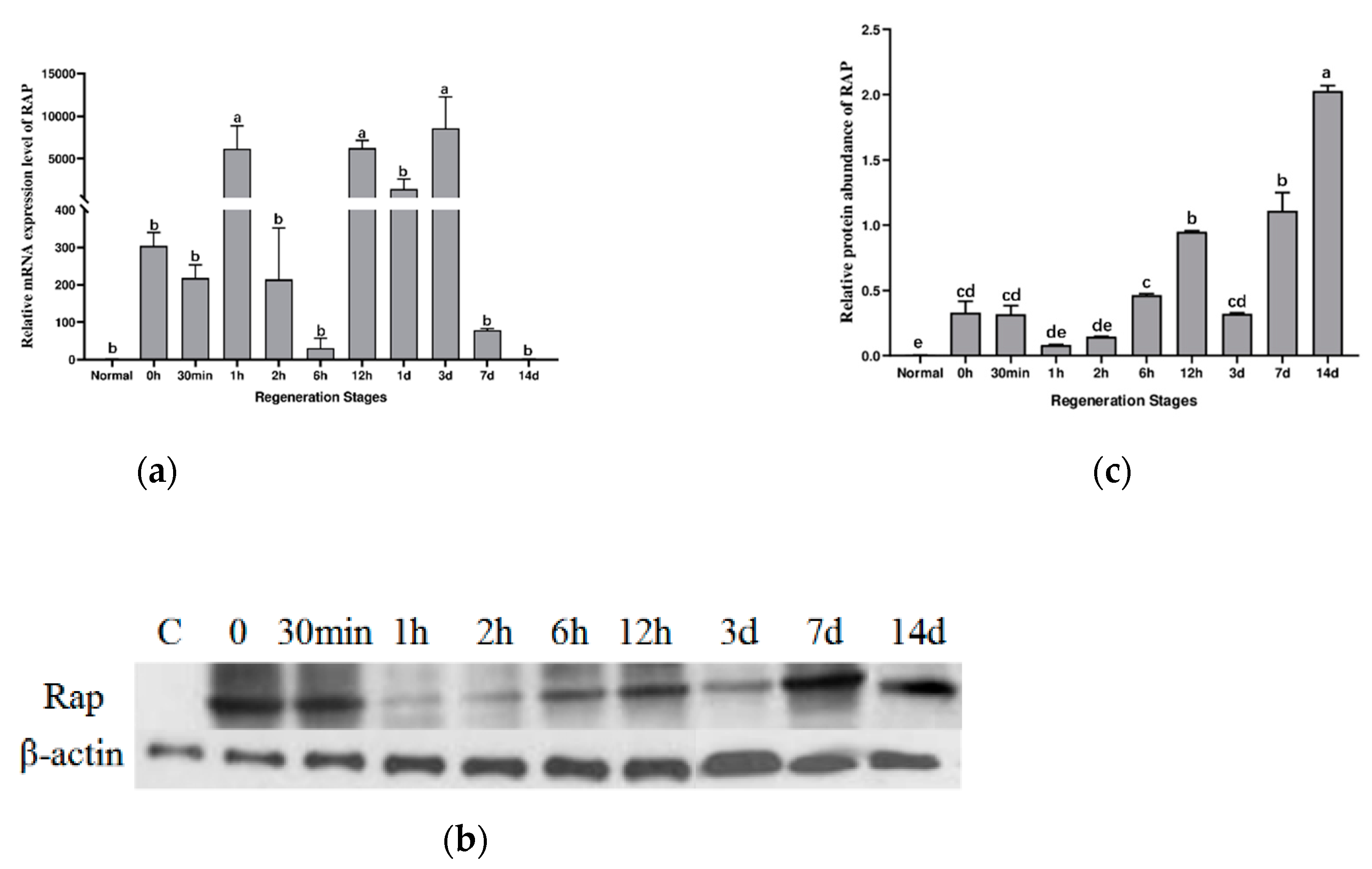
2.2. Aj-Orpin Gene Expression Pattern at the mRNA and Protein Level during Intestine Regeneration
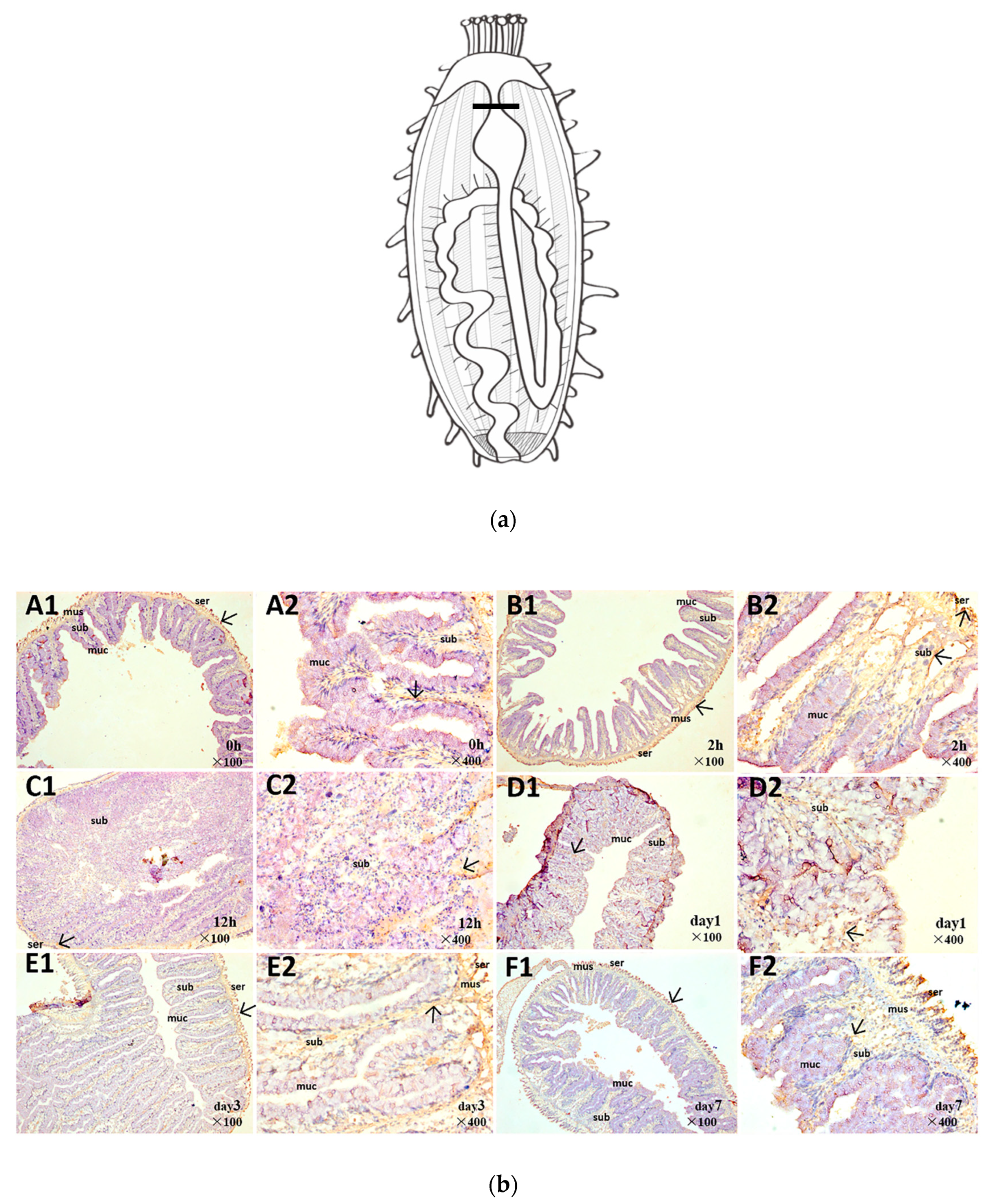
2.3. Spatiotemporal Pattern of Aj-Orpin Expression
3. Discussion
3.1. Aj-Orpin Is a Novel Secreted Calcium-Binding Protein (CaBP) Isolated from A. japonicus
3.2. RAP May Participate in Cell Proliferation during Intestinal Regeneration
4. Materials and Methods
4.1. Experimental Animals and Tissue Collection
4.2. RNA Extraction and Cloning of Full-Length Aj-Orpin Complementary DNA (cDNA)
4.3. Sequence Analysis
4.4. Gene Expression Analysis by Real-Time Quantitative PCR (qRT-PCR)
4.5. Western Blotting
4.6. Immunohistochemistry
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, L. Histocytological Events and Analysis of Key Genes during Intestine Regeneration in Sea Cucumber Apostichopus japonicus (Selenka). Ph.D. Thesis, The University of Chinese Academy of Sciences, Beijing, China, 2013. [Google Scholar]
- Dolmatov, I.Y. Molecular aspects of regeneration mechanisms in holothurians. Genes 2021, 12, 250. [Google Scholar] [CrossRef] [PubMed]
- Mashanov, V.; Frolova, L.; Dolmatov, I.Y. Structure of the digestive tube in the holothurian Eupentacta fraudatrix (Holothuroidea: Dendrochirota). Russ. J. Mar. Biol. 2004, 30, 314–322. [Google Scholar] [CrossRef]
- Shukalyuk, A.; Dolmatov, I.Y. Regeneration of the digestive tube in the holothurian Apostichopus japonicus after evisceration. Russ. J. Mar. Biol. 2001, 27, 168–173. [Google Scholar] [CrossRef]
- García-Arrarás, J.; Estrada-Rodgers, L.; Santiago, R.; Torres, I.I.; Díaz-Miranda, L.; Torres-Avillán, I. Cellular mechanisms of intestine regeneration in the sea cucumber, Holothuria glaberrima Selenka (Holothuroidea: Echinodermata). J. Exp. Zool. 1998, 281, 288–304. [Google Scholar] [CrossRef]
- Sun, L.N.; Yang, H.S.; Chen, M.Y.; Xu, D.X. Cloning and expression analysis of Wnt6 and Hox6 during intestinal regeneration in the sea cucumber Apostichopus japonicus. Genet. Mol. Res. 2013, 12, 5321–5334. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, X. The morphological and histological observation of regeneration of alimentary tract in sea cucumber Apostichopus japonicus. J. Dalian Fish. Univ. 2007, 22, 340–346. [Google Scholar]
- Candelaria, A.G.; Murray, G.; File, S.K.; García-Arrarás, J.E. Contribution of mesenterial muscle dedifferentiation to intestine regeneration in the sea cucumber Holothuria glaberrima. Cell Tissue Res. 2006, 325, 55–65. [Google Scholar] [CrossRef]
- Garcia-Arraras, J.E.; Bello, S.A.; Malavez, S. The Mesentery as the Epicenter for Intestinal Regeneration. In Seminars in Cell & Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2019; Volume 92, pp. 45–54. [Google Scholar]
- García-Arrarás, J.E.; Greenberg, M.J. Visceral regeneration in holothurians. Microsc. Res. Tech. 2001, 55, 438–451. [Google Scholar] [CrossRef]
- Mashanov, V.S.; Dolmatov, I.Y.; Heinzeller, T. Transdifferentiation in holothurian gut regeneration. Biol. Bull. 2005, 209, 184. [Google Scholar] [CrossRef] [PubMed]
- Nieves-Rios, C.; Alvarez-Falcon, S.; Malavez, S.; Rodriguez-Otero, J.; Garcia-Arraras, J.E. The nervous system component of the mesentery of the sea cucumber Holothuria glaberrima in normal and regenerating animals. Cell Tissue Res. 2019, 380, 67–77. [Google Scholar] [CrossRef]
- García-Arrarás, J.E.; Valentin-Tirado, G.; Flores, J.E.; Rosa, R.J.; Rivera-Cruz, A.; San Miguel-Ruiz, J.E.; Tossas, K. Cell dedifferentiation and epithelial to mesenchymal transitions during intestinal regeneration in H. glaberrima. BMC Dev. Biol. 2011, 11, 61–78. [Google Scholar] [CrossRef] [Green Version]
- Miao, T.; Wan, Z.X.; Sun, L.N.; Li, X.N.; Xing, L.L.; Bai, Y.C.; Wang, F.; Yang, H.S. Extracellular matrix remodeling and matrix metalloproteinases (ajMMP-2 like and ajMMP-16 like) characterization during intestine regeneration of sea cucumber Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2017, 212, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Quinones, J.L.; Rosa, R.; Ruiz, D.L.; Garcia-Arraras, J.E. Extracellular matrix remodeling and metalloproteinase involvement during intestine regeneration in the sea cucumber Holothuria glaberrima. Dev. Biol. 2002, 250, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Boyko, A.V.; Girich, A.S.; Tkacheva, E.S.; Dolmatov, I.Y. The Eupentacta fraudatrix transcriptome provides insights into regulation of cell transdifferentiation. Sci. Rep. 2020, 10, 1522. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Hamel, J.-F.; Mercier, A. The Sea Cucumber Apostichopus Japonicus: History, Biology and Aquaculture; Academic Press: Cambridge, MA, USA, 2015; p. 454. [Google Scholar]
- Ortiz-Pineda, P.A.; Ramírez-Gómez, F.; Pérez-Ortiz, J.; González-Díaz, S.; Santiago-De Jesús, F.; Hernández-Pasos, J.; Del Valle-Avila, C.; Rojas-Cartagena, C.; Suárez-Castillo, E.C.; Tossas, K.; et al. Gene expression profiling of intestinal regeneration in the sea cucumber. BMC Genom. 2009, 10, 262. [Google Scholar] [CrossRef]
- Zheng, F.-X.; Sun, X.-Q.; Fang, B.-H.; Hong, X.-G.; Zhang, J.-X. Comparative analysis of genes expressed in regenerating intestine and non-eviscerated intestine of Apostichopus japonicus Selenka (Aspidochirotida: Stichopodidae) and cloning of ependymin gene. Hydrobiologia 2006, 571, 109–122. [Google Scholar] [CrossRef]
- Rojas-Cartagena, C.; Ortiz-Pineda, P.; Ramirez-Gomez, F.; Suarez-Castillo, E.C.; Matos-Cruz, V.; Rodriguez, C.; Ortiz-Zuazaga, H.; Garcia-Arraras, J.E. Distinct profiles of expressed sequence tags during intestinal regeneration in the sea cucumber Holothuria glaberrima. Physiol. Genom. 2007, 31, 203–215. [Google Scholar] [CrossRef]
- Sun, L.; Chen, M.; Yang, H.; Wang, T.; Liu, B.; Shu, C.; Gardiner, D.M. Large scale gene expression profiling during intestine and body wall regeneration in the sea cucumber Apostichopus japonicus. Comp. Biochem. Physiol. Part D Genom. Proteom. 2011, 6, 195–205. [Google Scholar] [CrossRef]
- Quispe-Parra, D.J.; Medina-Feliciano, J.G.; Cruz-Gonzalez, S.; Ortiz-Zuazaga, H.; Garcia-Arraras, J.E. Transcriptomic analysis of early stages of intestinal regeneration in Holothuria glaberrima. Sci. Rep. 2021, 11, 346. [Google Scholar] [CrossRef]
- Sun, L.; Yang, H.; Chen, M.; Ma, D.; Lin, C. RNA-seq reveals dynamic changes of gene expression in key stages of intestine regeneration in the sea cucumber Apostichopus japonicus. PLoS ONE 2013, 8, e69441. [Google Scholar]
- Zhang, X.; Sun, L.; Yuan, J.; Sun, Y.; Gao, Y.; Zhang, L.; Li, S.; Dai, H.; Hamel, J.-F.; Liu, C.; et al. The sea cucumber genome provides insights into morphological evolution and visceral regeneration. PLoS Biol. 2017, 15, e2003790. [Google Scholar] [CrossRef]
- Sun, L.; Sun, J.; Li, X.; Zhang, L.; Yang, H.; Wang, Q. Understanding regulation of microRNAs on intestine regeneration in the sea cucumber Apostichopus japonicus using high-throughput sequencing. Comp. Biochem. Physiol. Part D Genom. Proteom. 2017, 22, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bello, S.A.; Torres-Gutierrez, V.; Rodriguez-Flores, E.J.; Toledo-Roman, E.J.; Rodriguez, N.; Diaz-Diaz, L.M.; Vazquez-Figueroa, L.D.; Cuesta, J.M.; Grillo-Alvarado, V.; Amador, A.; et al. Insights into intestinal regeneration signaling mechanisms. Dev. Biol. 2019, 458, 12–31. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Gao, Y.; Sun, L.; Jin, S.; Zhang, X.; Liu, C.; Li, F.; Xiang, J. Wnt signaling pathway linked to intestinal regeneration via evolutionary patterns and gene expression in the sea cucumber Apostichopus japonicus. Front. Genet. 2019, 10, 112. [Google Scholar] [CrossRef]
- Dolmatov, I.Y.; Kalacheva, N.V.; Tkacheva, E.S.; Shulga, A.P.; Zavalnaya, E.G.; Shamshurina, E.V.; Girich, A.S.; Boyko, A.V.; Eliseikina, M.G. Expression of Piwi, MMP, TIMP, and Sox during gut regeneration in holothurian Eupentacta fraudatrix (Holothuroidea, Dendrochirotida). Genes 2021, 12, 1292. [Google Scholar] [CrossRef] [PubMed]
- Girich, A.S.; Issaeva, M.P.; Dolmatov, I.Y. Wnt and frizzled expression during regeneration of internal organs in the holothurian Eupentacta fraudatrix. Wound Repair Regen. 2017, 25, 828–835. [Google Scholar] [CrossRef]
- Mashanov, V.S.; Zueva, O.R.; Garcia-Arraras, J.E. Expression of Wnt9, TCTP, and Bmp1/Tll in sea cucumber visceral regeneration. Gene Expr. Patterns 2012, 12, 24–35. [Google Scholar] [CrossRef]
- Mashanov, V.S.; Zueva, O.R.; Rojas-Catagena, C.; García-Arrarás, J.E. Visceral regeneration in a sea cucumber involves extensive expression of survivin and mortalin homologs in the mesothelium. BMC Dev. Biol. 2010, 10, 117–141. [Google Scholar] [CrossRef]
- Santiago-Cardona, P.G.; Berrios, C.A.; Ramirez, F.; Garcia-Arraras, J.E. Lipopolysaccharides induce intestinal serum amyloid A expression in the sea cucumber Holothuria glaberrima. Dev. Comp. Immunol. 2003, 27, 105–110. [Google Scholar] [CrossRef]
- Soto-Acabá, A.; Ortíz-Pineda, P.A.; García-Arrarás, J.E. Characterization of two novel EF-hand proteins identifies a clade of putative Ca2+-binding protein specific to the Ambulacraria. J. Bioinform. Syst. Biol. 2022, 5, 1–25. [Google Scholar] [CrossRef]
- Kretsinger, R.H.; Nockolds, C.E. Carp muscle calcium-binding protein: II. structure determination and general description. J. Biol. Chem. 1973, 248, 3313–3326. [Google Scholar] [CrossRef]
- McPhalen, C.A.; Strynadka, N.C.J.; James, M.N.G. Calcium-binding sites in proteins—A structural perspective. Adv. Protein Chem. 1991, 42, 77–144. [Google Scholar] [PubMed]
- Nelson, M.R.; Chazin, W.J. Structures of EF-hand Ca2+-binding proteins: Diversity in the organization, packing and response to Ca2+ binding. Biometals 1998, 11, 297–318. [Google Scholar] [CrossRef] [PubMed]
- Persechini, A.; Moncrief, N.D.; Kretsinger, R.H. The Ef-hand family of calcium-modulated proteins. Trends Neurosci. 1989, 12, 462–467. [Google Scholar] [CrossRef]
- Yu, T.; Zhao, Y.; Yu, S. Progress on the EF-hand proteins. Biophysics 2013, 1, 1–10. [Google Scholar]
- Kretsinger, R.H.; Tolbert, D.; Nakayama, S.; Pearson, W. The EF-Hand, Homologs and Analogs. In Novel Calcium-Binding Proteins: Fundamentals and Clinical Implications; Heizmann, C.W., Ed.; Springer: Berlin/Heidelberg, Germany, 1991; pp. 17–37. [Google Scholar]
- Phan, E.; Ahluwalla, A.; Tarnawski, A.S. Role of SPARC-matricellular protein in pathophysiology and tissue injury healing. Implications for gastritis and gastric ulcers. Med. Sci. Monit. 2007, 13, RA25–RA30. [Google Scholar]
- Brekken, R.A.; Sage, E.H. SPARC, a matricellular protein: At the crossroads of cell-matrix communication. Matrix Biol. J. Int. Soc. Matrix Biol. 2001, 19, 816–827. [Google Scholar] [CrossRef]
- Latvala, T.; Puolakkainen, P.; Vesaluoma, M.; Tervo, T. Distribution of SPARC protein (osteonectin) in normal and wounded feline cornea. Exp. Eye Res. 1996, 63, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Reed, M.J.; Puolakkainen, P.; Lane, T.F.; Dickerson, D.; Bornstein, P.; Sage, E.H. Differential expression of sparc and thrombospondin 1 in wound repair: Immunolocalization and in-situ hybridization. J. Histochem. Cytochem. 1993, 41, 1467–1477. [Google Scholar] [CrossRef] [PubMed]
- Sage, H.; Vernon, R.B.; Decker, J.; Funk, S.; Iruelaarispe, M.L. Distribution of the calcium-binding protein SPARC in tissues of embryonic and adult mice. J. Histochem. Cytochem. 1989, 37, 819–829. [Google Scholar] [CrossRef]
- Tarnawski, A.S. Cellular and molecular mechanisms of gastrointestinal ulcer healing. Dig. Dis. Sci. 2005, 50, S24–S33. [Google Scholar] [CrossRef]
- Moe, A.M.; Golding, A.E.; Bement, W.M. Cell Healing: Calcium, Repair and Regeneration. In Seminars in Cell & Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2015; Volume 45, pp. 18–23. [Google Scholar]
- Huo, D.; Sun, L.; Zhang, L.; Ru, X.; Liu, S.; Yang, X.; Yang, H. Global-warming-caused changes of temperature and oxygen alter the proteomic profile of sea cucumber Apostichopus japonicus. J. Proteom. 2019, 193, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Sun, L.; Yang, H.; Zhang, L.; Miao, T.; Xing, L.; Huo, D. Identification and expression characterization of WntA during intestinal regeneration in the sea cucumber Apostichopus japonicus. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2017, 210, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Mashanov, V.S.; Dolmatov, I.Y. Regeneration of digestive tract in the pentactulae of the far-eastern holothurian Eupentacta fraudatrix (Holothuroidea, Dendrochirota). Invertebr. Reprod. Dev. 2001, 39, 143–151. [Google Scholar] [CrossRef]
- Odintsova, N.A.; Dolmatov, I.Y.; Mashanov, V.S. Regenerating holothurian tissues as a source of cells for long-term cell cultures. Mar. Biol. 2005, 146, 915–921. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11 molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Whelan, S.; Goldman, N. A general empirical model of protein evolution derived from multiple protein families using a maximum-likelihood approach. Mol. Biol. Evol. 2001, 18, 691–699. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence |
|---|---|
| GspAj-Orpin F1 | 5′- GACCCACCCCAACCCGATTC -3′ |
| GspAj-Orpin R1 | 5′- GCTTTGCCGCCCTCTTTCGT -3′ |
| GspAj-Orpin F2 | 5′- GGTGGATGGTTGAAGAGAAGTTGGAAG -3′ |
| GspAj-Orpin R2 | 5′- CCCCACTTCGTCACGTTGATATTTACC -3′ |
| UPM | Long5′-CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT-3′ Short5′-CTAATACGACTCACTATAGGGC-3′ |
| RT-Aj-Orpin F | 5′-AGTGGTGCTGTTGGTGTTAAGGTC-3′ |
| RT-Aj-Orpin R | 5′-CCGCCCTCTTTCGTCCATTCAC-3′ |
| NADHF | 5′-GTCCTACGACCCAATCTGGA-3′ |
| NADHR | 5′-ATGAGCCTTGGTTACGTTGG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, F.; Sun, L.; Li, X.; Cui, W.; Yang, H. Characterization and Expression Analysis of Regeneration-Associated Protein (Aj-Orpin) during Intestinal Regeneration in the Sea Cucumber Apostichopus japonicus. Mar. Drugs 2022, 20, 568. https://doi.org/10.3390/md20090568
Su F, Sun L, Li X, Cui W, Yang H. Characterization and Expression Analysis of Regeneration-Associated Protein (Aj-Orpin) during Intestinal Regeneration in the Sea Cucumber Apostichopus japonicus. Marine Drugs. 2022; 20(9):568. https://doi.org/10.3390/md20090568
Chicago/Turabian StyleSu, Fang, Lina Sun, Xiaoni Li, Wei Cui, and Hongsheng Yang. 2022. "Characterization and Expression Analysis of Regeneration-Associated Protein (Aj-Orpin) during Intestinal Regeneration in the Sea Cucumber Apostichopus japonicus" Marine Drugs 20, no. 9: 568. https://doi.org/10.3390/md20090568