
Active Targeting of P-Selectin by Fucoidan Modulates the Molecular Profiling of Metastasis in Docetaxel-Resistant Prostate Cancer

 , and
, and 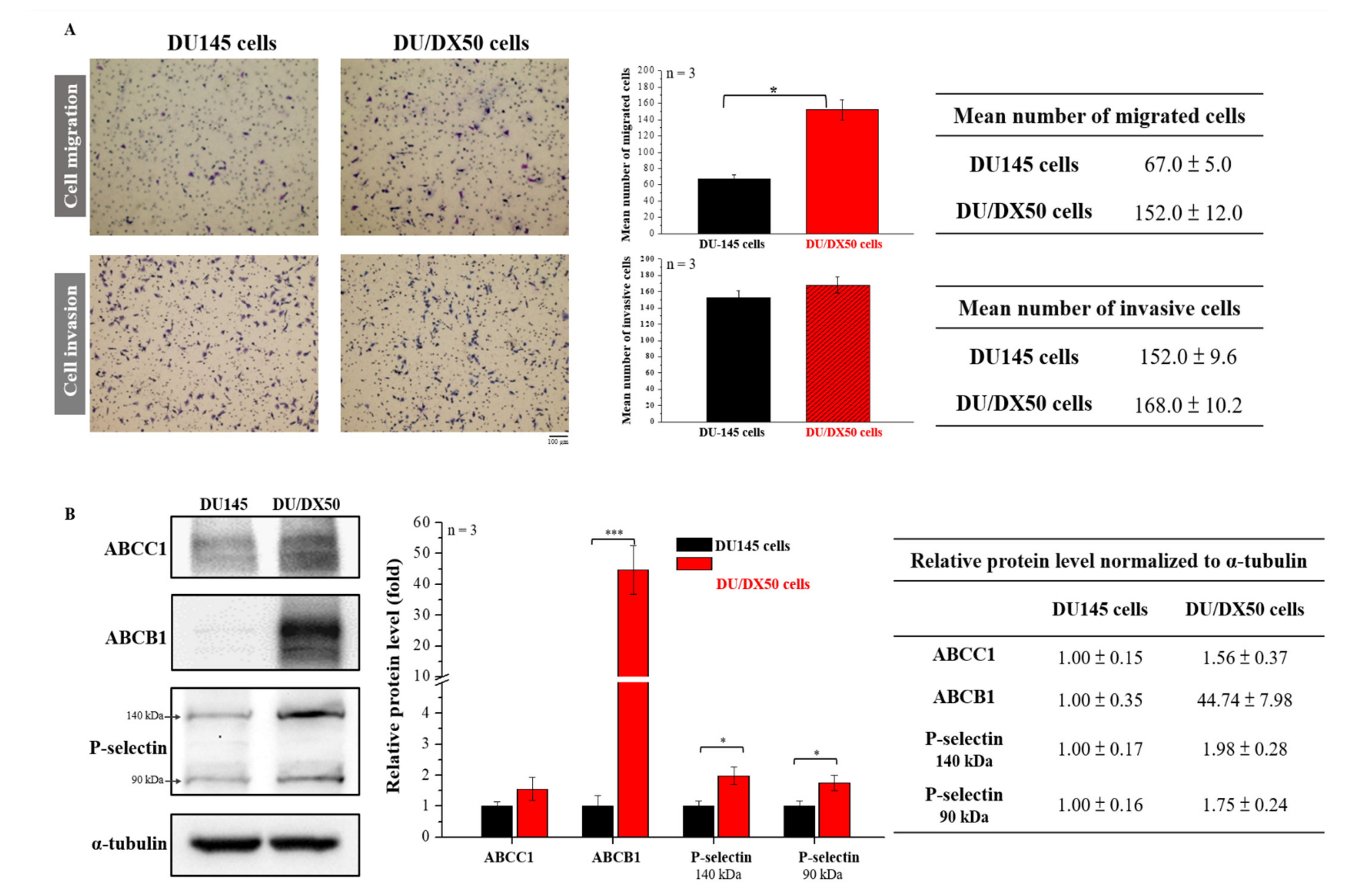{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Docetaxel-Resistant PCa Cells Exhibit High Motility and Malignancy-Associated Protein Expression
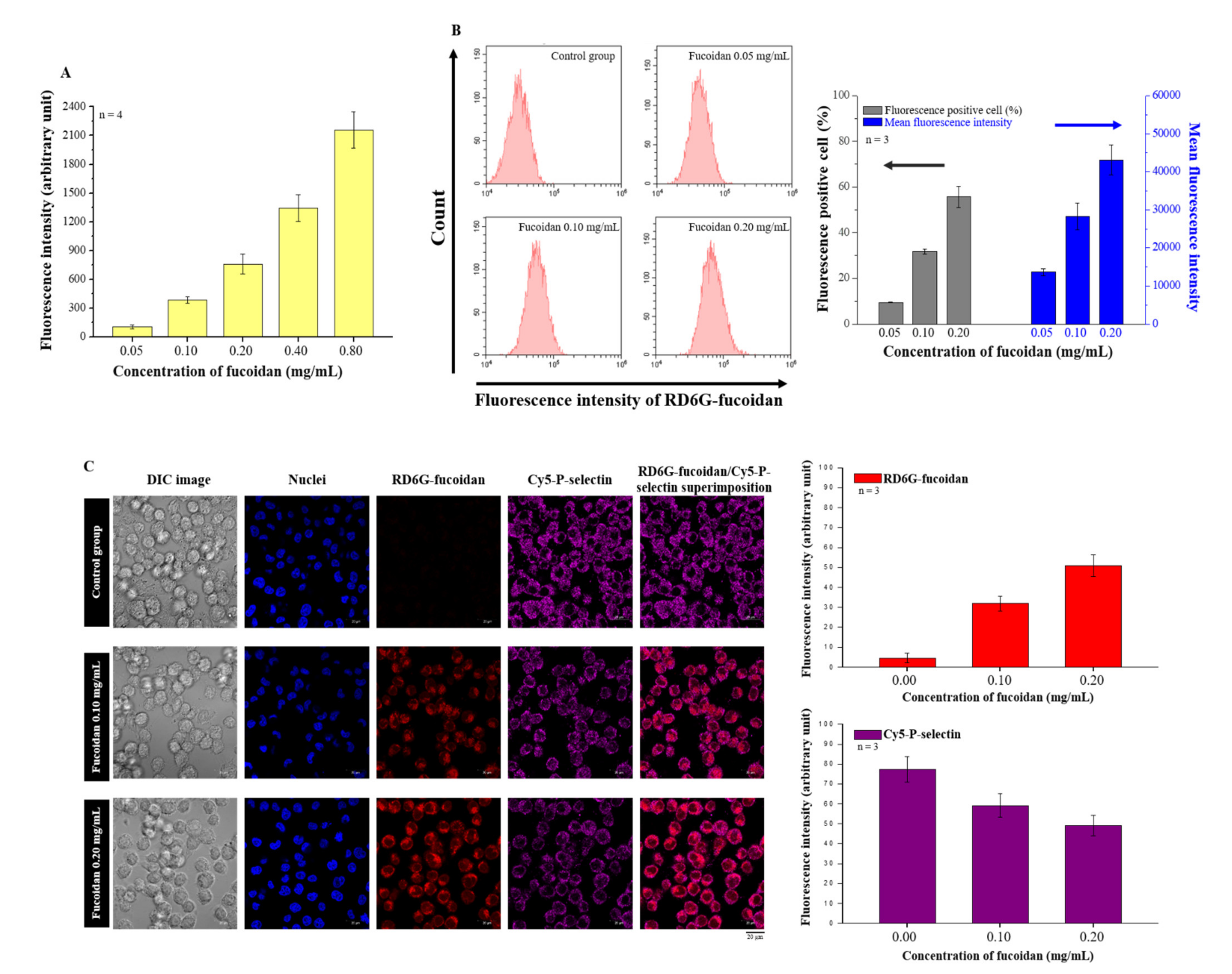
2.2. Effects of Fucoidan on P-Selectin Binding and Cellular Uptake
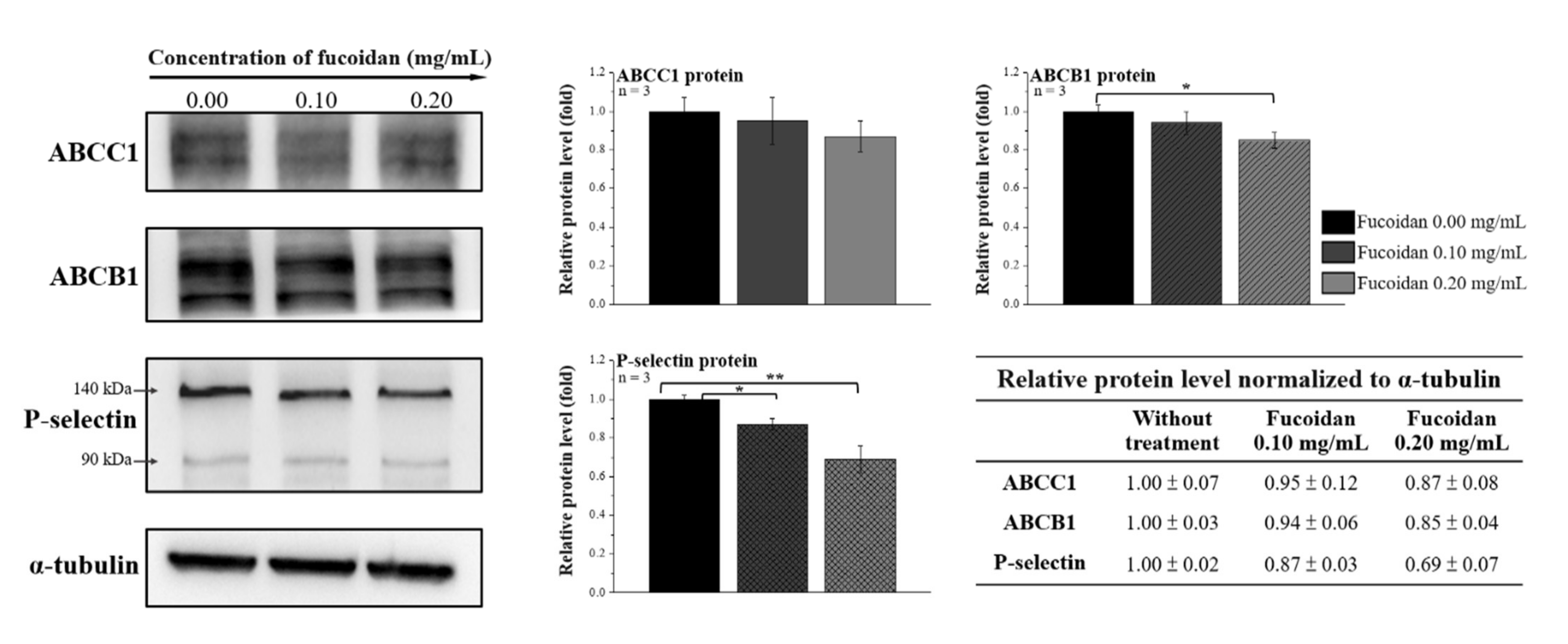
2.3. Fucoidan Reduces the Expression of the ABC Transporter Family and P-Selectin in Docetaxel-Resistant DU/DX50 Cells
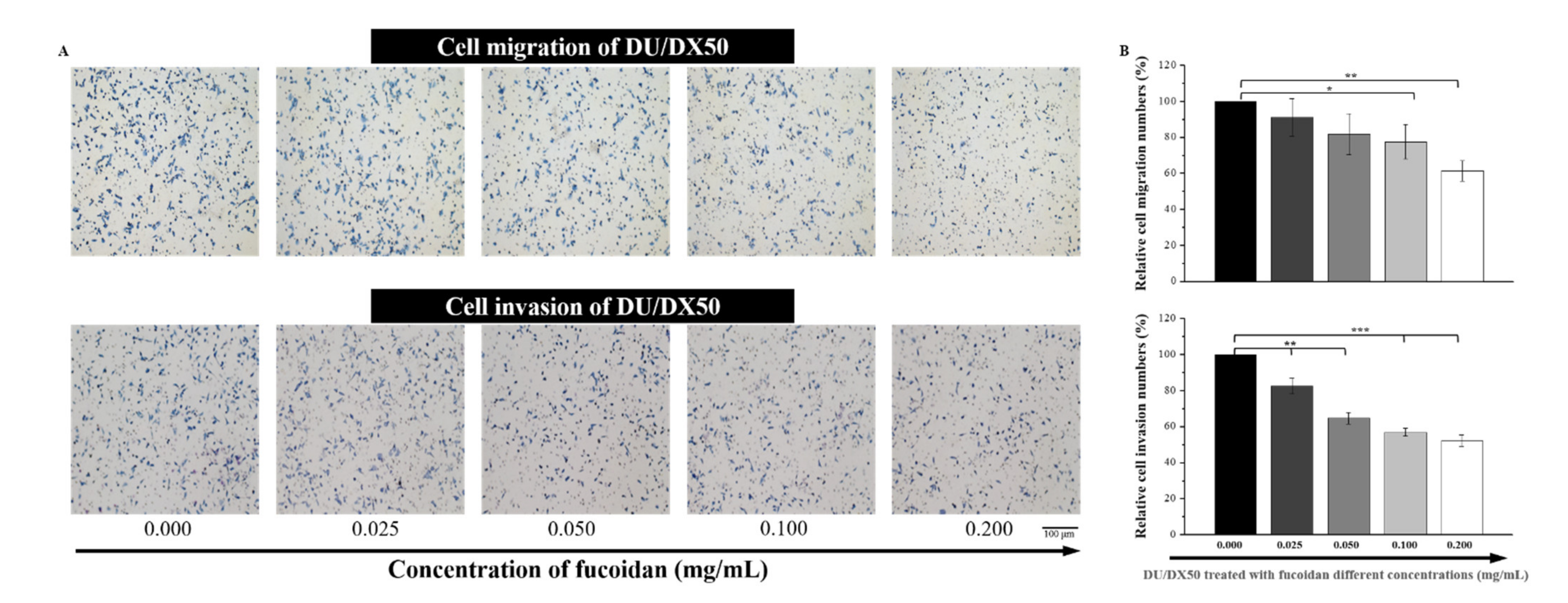
2.4. Fucoidan Attenuates the Abilities of Migration and Invasion in Docetaxel-Resistant DU/DX50 Cells
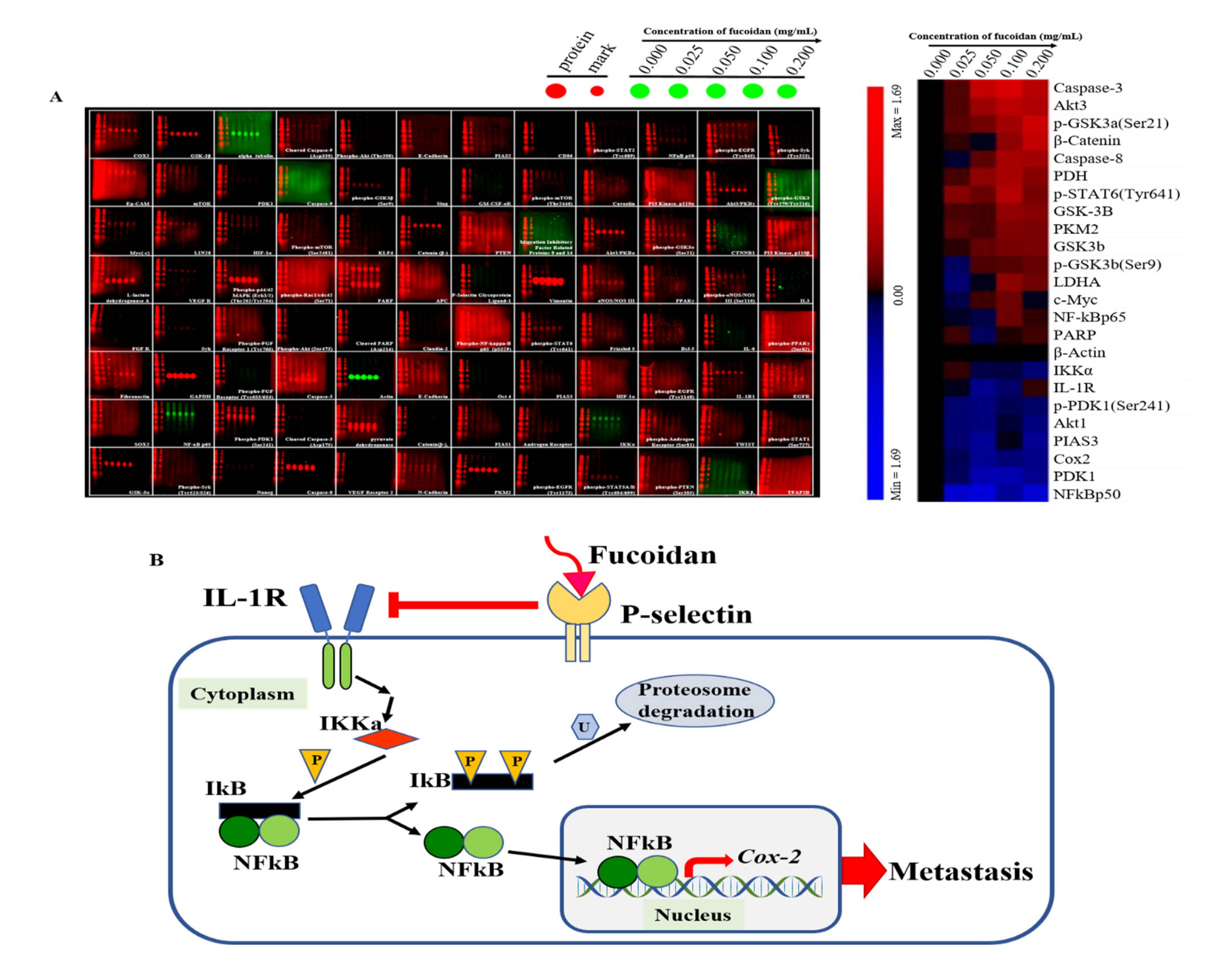
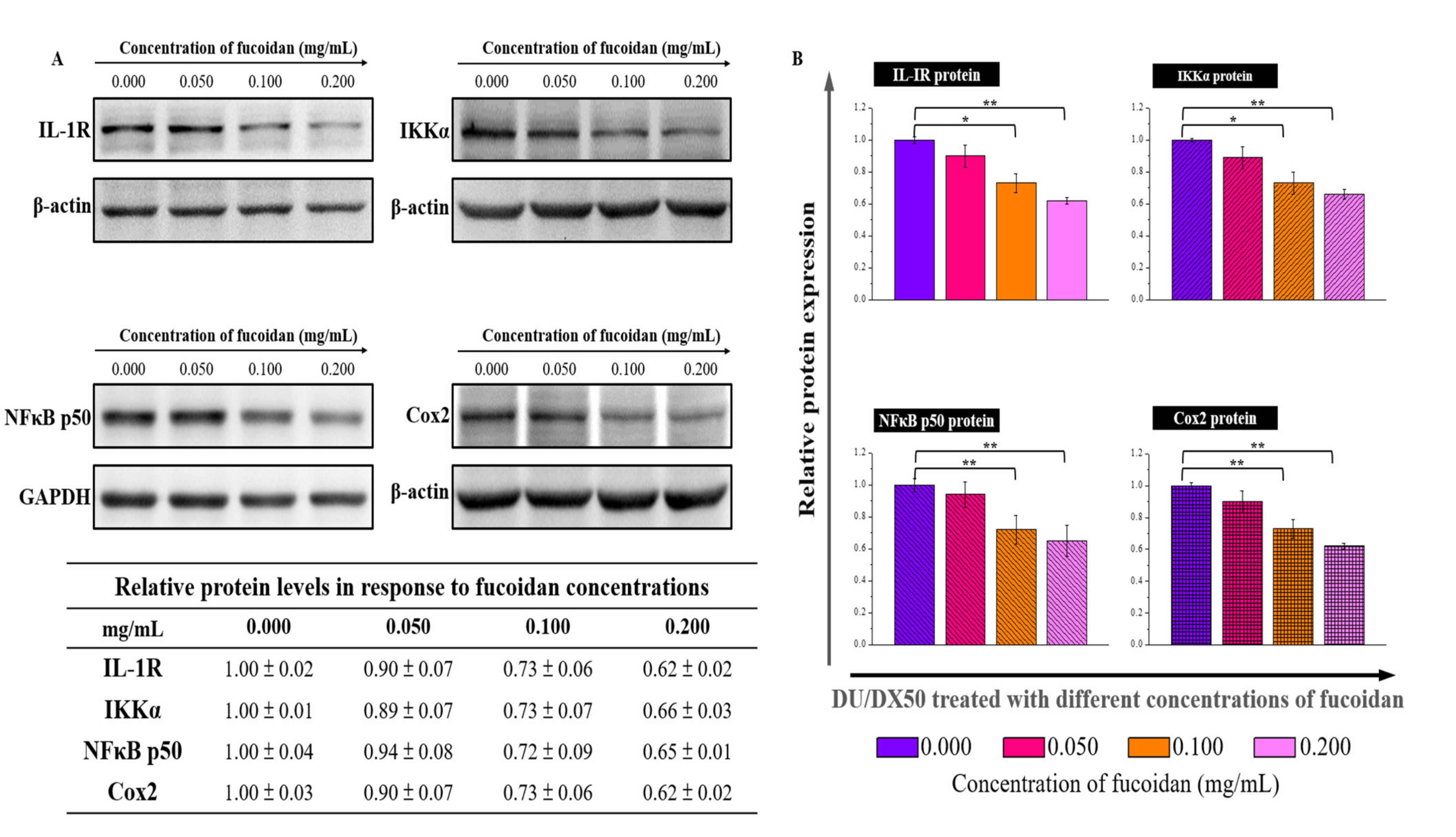
2.5. Expression of Hypothesized Signal Transduction Pathways and Metastasis-Related Protein in Fucoidan Treated Docetaxel-Resistant DU/DX50 Cells
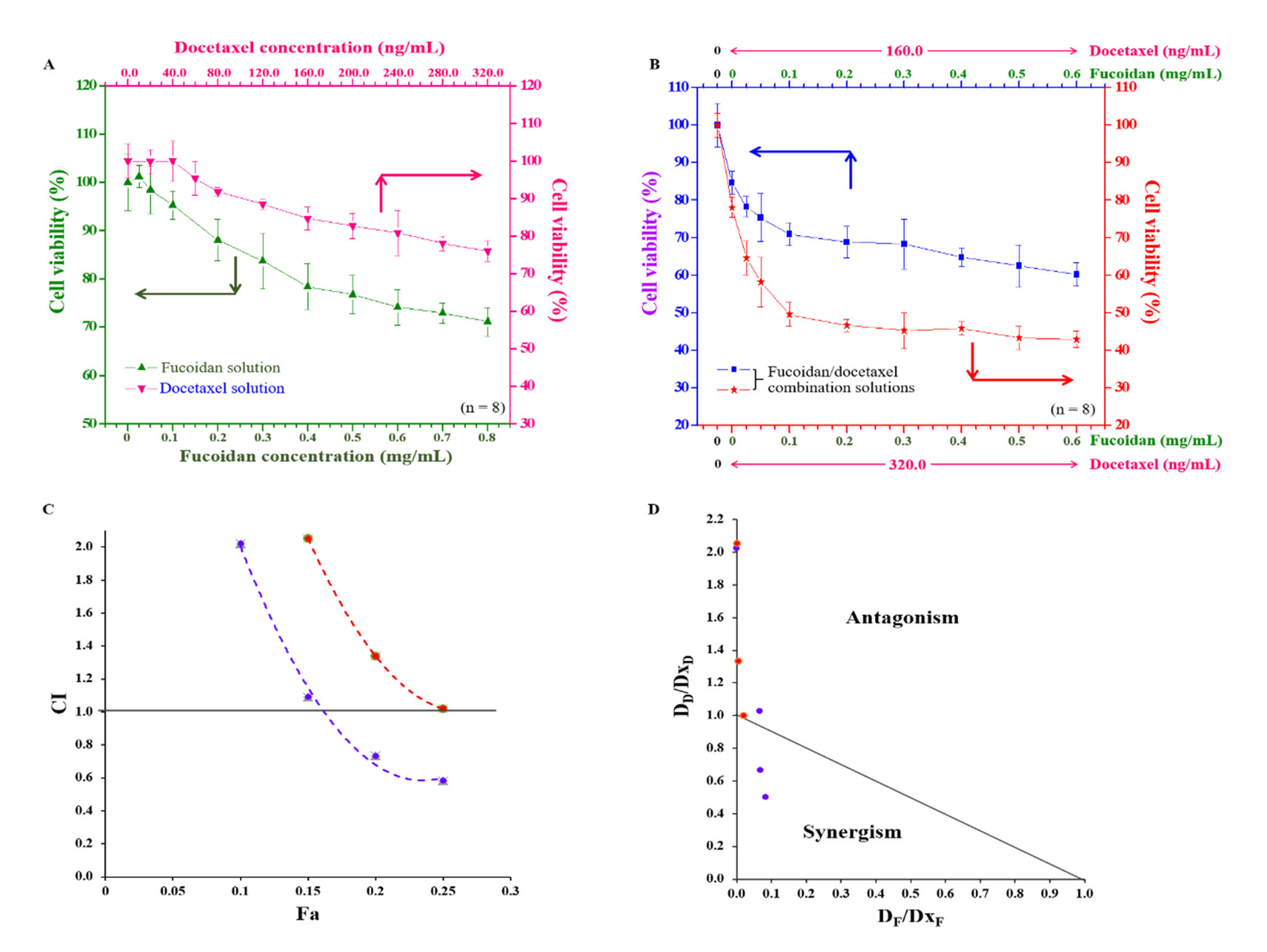
2.6. Anti-Cancer Effects of Fucoidan and Docetaxel and the Synergistic Effect of the Combination of Fucoidan/Docetaxel on Docetaxel-Resistant DU/DX50 Cells
3. Discussion
4. Materials and Methods
4.1. Transwell Migration, Invasion, and Western Blotting Arrays on Docetaxel-Resistant PCa Cells
4.2. Assay of Fucoidan Binding to the P-Selectin Protein and Cellular Uptake of Fucoidan
4.3. Determining the Binding Ability to P-Selectin and the Effect on the Expression of Related Proteins in Fucoidan-Treated PCa Cells
4.4. Determination of the Effect of Fucoidan on the Migration and Invasion of Docetaxel-Resistant PCa Cells
4.5. Analysis of the Effect of Fucoidan on the Expression of Metastasis-Related Proteins
4.6. Evaluation of the Anticancer Effect of Docetaxel-Resistant PCa Cells and the Determination of Combination Index
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Galsky, M.D.; Vogelzang, N.J. Docetaxel-based combination therapy for castration-resistant prostate cancer. Ann. Oncol. 2010, 21, 2135–2144. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Mo, M.; Wei, Y.; Wu, J.; Pan, J.; Freedland, S.J.; Zheng, Y.; Ye, D. Epidemiology and genomics of prostate cancer in Asian men. Nat. Rev. Urol. 2021, 18, 282–301. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Center, M.M.; DeSantis, C.; Ward, E.M. Global patterns of cancer incidence and mortality rates and trends. Cancer Epidemiol. Biomark. Prev. 2010, 19, 1893–1907. [Google Scholar] [CrossRef] [PubMed]
- Taitt, H.E. Global trends and prostate cancer: A review of incidence, detection, and mortality as influenced by race, ethnicity, and geographic location. Am. J. Mens Health 2018, 12, 1807–1823. [Google Scholar] [CrossRef]
- Shubina, L.K.; Makarieva, T.N.; Denisenko, V.A.; Popov, R.S.; Dyshlovoy, S.A.; Grebnev, B.B.; Dmitrenok, P.S.; von Amsberg, G.; Stonik, V.A. Gracilosulfates A–G, monosulfated polyoxygenated steroids from the marine sponge Haliclona gracilis. Mar. Drugs 2020, 18, 454. [Google Scholar] [CrossRef]
- Kim, T.J.; Lee, Y.H.; Koo, K.C. Current status and future perspectives of androgen receptor inhibition therapy for prostate cancer: A comprehensive review. Biomolecules 2021, 11, 492. [Google Scholar] [CrossRef]
- Murakami, M.; Ishikawa, H.; Shimizu, S.; Iwata, H.; Okimoto, T.; Takagi, M.; Murayama, S.; Akimoto, T.; Wada, H.; Arimura, T.; et al. Optimal androgen deprivation therapy combined with proton beam therapy for prostate cancer: Results from a multi-institutional study of the Japanese radiation oncology study group. Cancers 2020, 12, 1690. [Google Scholar] [CrossRef]
- Labriola, M.K.; Atiq, S.; Hirshman, N.; Bitting, R.L. Management of men with metastatic castration-resistant prostate cancer following potent androgen receptor inhibition: A review of novel investigational therapies. Prostate Cancer Prostatic Dis. 2021, 24, 301–309. [Google Scholar] [CrossRef]
- Seruga, B.; Tannock, I.F. Chemotherapy-based treatment for castration-resistant prostate cancer. J. Clin. Oncol. 2011, 29, 3686–3694. [Google Scholar] [CrossRef]
- Tannock, I.F.; de Wit, R.; Berry, W.R.; Horti, J.; Pluzanska, A.; Chi, K.N.; Oudard, S.; Théodore, C.; James, N.D.; Turesson, I.; et al. Docetaxel plus prednisone or mitoxantrone plus prednisone for advanced prostate cancer. N. Engl. J. Med. 2004, 351, 1502–1512. [Google Scholar] [CrossRef] [PubMed]
- Petrylak, D.P.; Tangen, C.M.; Hussain, M.H.; Lara, P.N., Jr.; Jones, J.A.; Taplin, M.E.; Burch, P.A.; Berry, D.; Moinpour, C.; Kohli, M.; et al. Docetaxel and estramustine compared with mitoxantrone and prednisone for advanced refractory prostate cancer. N. Engl. J. Med. 2004, 351, 1513–1520. [Google Scholar] [CrossRef] [PubMed]
- Dagher, R.; Li, N.; Abraham, S.; Rahman, A.; Sridhara, R.; Pazdur, R. Approval summary: Docetaxel in combination with prednisone for the treatment of androgen-independent hormone-refractory prostate cancer. Clin. Cancer Res. 2004, 10, 8147–8151. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.J.; Lin, C.Y.; Liao, W.Y.; Hour, T.C.; Wang, H.D.; Chuu, C.P. CD44 promotes migration and invasion of docetaxel-resistant prostate cancer cells likely via induction of Hippo-Yap signaling. Cells 2019, 8, 295. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.K.; Lee, S.J.; Kwon, W.A.; Jeong, K.C. Docetaxel-resistant prostate cancer cells become sensitive to gemcitabine due to the upregulation of ABCB1. Prostate 2020, 80, 453–462. [Google Scholar] [CrossRef]
- Choi, C.H. ABC transporters as multidrug resistance mechanisms and the development of chemosensitizers for their reversal. Cancer Cell Int. 2005, 5, 30. [Google Scholar] [CrossRef]
- Leslie, E.M.; Deeley, R.G.; Cole, S.P.C. Multidrug resistance proteins: Role of P-glycoprotein, MRP1, MRP2, and BCRP (ABCG2) in tissue defense. Toxicol. Appl. Pharmacol. 2005, 204, 216–237. [Google Scholar] [CrossRef]
- Haimeur, A.; Conseil, G.; Deeley, R.G.; Cole, S.P.C. The MRP-related and BCRP/ABCG2 multidrug resistance proteins: Biology, substrate specificity and regulation. Curr. Drug. Metab. 2004, 5, 21–53. [Google Scholar] [CrossRef]
- Zhou, H.H.; Chen, X.; Cai, L.Y.; Nan, X.W.; Chen, J.H.; Chen, X.X.; Yang, Y.; Xing, Z.H.; Wei, M.N.; Li, Y.; et al. Erastin reverses ABCB1-mediated docetaxel resistance in ovarian cancer. Front. Oncol. 2019, 9, 1398. [Google Scholar] [CrossRef]
- Fenner, A. Prostate cancer: Antiandrogens reverse docetaxel resistance via ABCB1 inhibition. Nat. Rev. Urol. 2015, 12, 361. [Google Scholar] [CrossRef]
- Mansoori, B.; Mohammadi, A.; Davudian, S.; Shirjang, S.; Baradaran, B. The different mechanisms of cancer drug resistance: A brief review. Adv. Pharm. Bull. 2017, 7, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hsia, S.; Wu, T.H.; Wu, C.J. Fish Oil, Se Yeast, and micronutrient-enriched nutrition as adjuvant treatment during target therapy in a murine model of lung cancer. Mar. Drugs 2021, 19, 262. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Madigan, M.C.; Khatri, A.; Power, C.A.; Hung, T.T.; Beretov, J.; Chang, L.; Xiao, W.; Cozzi, P.J.; Graham, P.H.; et al. In vitro and in vivo prostate cancer metastasis and chemoresistance can be modulated by expression of either CD44 or CD147. PLoS ONE 2012, 7, e40716. [Google Scholar]
- Zayed, A.; Avila-Peltroche, J.; El-Aasr, M.; Ulber, R. Sulfated galactofucans: An outstanding class of fucoidans with promising bioactivities. Mar. Drugs 2022, 20, 412. [Google Scholar] [CrossRef]
- Nagahawatta, D.P.; Liyanage, N.M.; Jayawardhana, H.H.A.C.K.; Lee, H.G.; Jayawardena, T.U.; Jeon, Y.J. Anti-Fine dust effect of fucoidan extracted from Ecklonia maxima laves in macrophages via inhibiting inflammatory signaling pathways. Mar. Drugs 2022, 20, 413. [Google Scholar] [CrossRef]
- Yao, Y.; Yim, E.K.F. Fucoidan for cardiovascular application and the factors mediating its activities. Carbohydr. Polym. 2021, 270, 118347. [Google Scholar] [CrossRef]
- Mabate, B.; Daub, C.D.; Malgas, S.; Edkins, A.L.; Pletschke, B.I. Fucoidan structure and its impact on glucose metabolism: Implications for diabetes and cancer therapy. Mar. Drugs 2021, 19, 30. [Google Scholar] [CrossRef]
- Atashrazm, F.; Lowenthal, R.M.; Woods, G.M.; Holloway, A.F.; Dickinson, J.L. Fucoidan and cancer: A multifunctional molecule with anti-tumor potential. Mar. Drugs 2015, 13, 2327–2346. [Google Scholar] [CrossRef]
- Novoyatleva, T.; Kojonazarov, B.; Owczarek, A.; Veeroju, S.; Rai, N.; Henneke, I.; Böhm, M.; Grimminger, F.; Ghofrani, H.A.; Seeger, W.; et al. Evidence for the fucoidan/p-selectin axis as a therapeutic target in hypoxia-induced pulmonary hypertension. Am. J. Respir. Crit. Care Med. 2019, 199, 1407–1420. [Google Scholar] [CrossRef]
- Läubli, H.; Borsig, L. Selectins promote tumor metastasis. Semin. Cancer Biol. 2010, 20, 169–177. [Google Scholar] [CrossRef]
- Cariello, M.; Piccinin, E.; Zerlotin, R.; Piglionica, M.; Peres, C.; Divella, C.; Signorile, A.; Villani, G.; Ingravallo, G.; Sabbà, C.; et al. Adhesion of platelets to colon cancer cells is necessary to promote tumor development in xenograft, genetic and inflammation models. Cancers 2021, 13, 4243. [Google Scholar] [CrossRef] [PubMed]
- Preobrazhenskaya, M.E.; Berman, A.E.; Mikhailov, V.I.; Ushakova, N.A.; Mazurov, A.V.; Semenov, A.V.; Usov, A.I.; Nifant'ev, N.E.; Bovin, N.V. Fucoidan inhibits leukocyte recruitment in a model peritoneal inflammation in rat and blocks interaction of p-selectin with Its carbohydrate ligand. Biochem. Mol. Biol. Int. 1997, 43, 443–451. [Google Scholar] [PubMed]
- Carvalho, A.C.; Sousa, R.B.; Franco, Á.X.; Costa, J.V.G.; Neves, L.M.; Ribeiro, R.A.; Sutton, R.; Criddle, D.N.; Soares, P.M.; de Souza, M.H. Protective effects of fucoidan, a p-and l-selectin inhibitor, in murine acute pancreatitis. Pancreas 2014, 43, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Shamay, Y.; Elkabets, M.; Li, H.; Shah, J.; Brook, S.; Wang, F.; Adler, K.; Baut, E.; Scaltriti, M.; Jena, P.V.; et al. P-selectin is a nanotherapeutic delivery target in the tumor microenvironment. Sci. Transl. Med. 2016, 8, 345ra87. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Kim, J.S.; Kim, E. Fucoidan from seaweed fucus vesiculosus inhibits migration and invasion of human lung cancer cell via PI3K-Akt-mTOR pathways. PLoS ONE 2012, 7, e50624. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki-Miyamoto, Y.; Yamasaki, M.; Tachibana, H.; Yamada, K. Fucoidan induces apoptosis through activation of caspase-8 on human breast cancer MCF-7 cells. J. Agric. Food Chem. 2009, 57, 8677–8682. [Google Scholar] [CrossRef]
- Hsu, H.Y.; Lin, T.Y.; Hu, C.H.; Shu, D.T.F.; Lu, M.K. Fucoidan upregulates TLR4/CHOP-mediated caspase-3 and PARP activation to enhance cisplatin-induced cytotoxicity in human lung cancer cells. Cancer Lett. 2018, 432, 112–120. [Google Scholar] [CrossRef]
- Gelfo, V.; Romaniello, D.; Mazzeschi, M.; Sgarzi, M.; Grilli, G.; Morselli, A.; Manzan, B.; Rihawi, K.; Lauriola, M. Roles of IL-1 in cancer: From tumor progression to resistance to targeted therapies. Int. J. Mol. Sci. 2020, 21, 6009. [Google Scholar] [CrossRef]
- Park, M.H.; Hong, J.T. Roles of NF-κB in cancer and inflammatory diseases and their therapeutic approaches. Cells 2016, 5, 15. [Google Scholar] [CrossRef]
- Pang, L.Y.; Hurst, E.A.; Argyle, D.J. Cyclooxygenase-2: A role in cancer stem cell survival and repopulation of cancer cells during therapy. Stem Cells Int. 2016, 2016, 2048731. [Google Scholar] [CrossRef]
- Fu, Y.K.; Wang, B.J.; Tseng, J.C.; Huang, S.H.; Lin, C.Y.; Kuo, Y.Y.; Hour, T.C.; Chuu, C.P. Combination treatment of docetaxel with caffeic acid phenethyl ester suppresses the survival and the proliferation of docetaxel-resistant prostate cancer cells via induction of apoptosis and metabolism interference. J. Biomed. Sci. 2022, 29, 16.3. [Google Scholar] [CrossRef] [PubMed]
- Hall, E.H.; Gurel, V.; Dahlberg, A.E.; McMichael, J.; Brautigan, D.L. Inhibition of human breast cancer Matrigel invasion by Streptolysin O activation of the EGF receptor ErbB1. Cell. Signal. 2011, 23, 1972–1977. [Google Scholar] [CrossRef] [PubMed]
- Rima, M.; Trognon, J.; Latapie, L.; Chbani, A.; Roques, C.; El Garah, F. Seaweed Extracts: A Promising Source of Antibiofilm Agents with Distinct Mechanisms of Action against Pseudomonas aeruginosa. Mar. Drugs 2022, 20, 92. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Yuan, X.; Yu, T.; Huang, H.; Yang, C.; Zhang, L.; Yang, S.; Luo, X.; Luo, J. Lycorine inhibits cell proliferation, migration and invasion, and primarily exerts in vitro cytostatic effects in human colorectal cancer via activating the ROS/p38 and AKT signaling pathways. Oncol. Rep. 2021, 45, 19. [Google Scholar] [CrossRef] [PubMed]
- Bachelet, L.; Bertholon, I.; Lavigne, D.; Vassy, R.; Jandrot-Perrus, M.; Chaubet, F.; Letourneur, D. Affinity of low molecular weight fucoidan for P-selectin triggers its binding to activated human platelets. Biochim. Biophys. Acta 2009, 1790, 141–146. [Google Scholar] [CrossRef]
- Frigault, M.M.; Lacoste, J.; Swift, J.L.; Brown, C.M. Live-cell microscopy—tips and tools. J. Cell Sci. 2009, 122, 753–767. [Google Scholar] [CrossRef]
- Ciaccio, M.F.; Wagner, J.P.; Chuu, C.P.; Lauffenburger, D.A.; Jones, R.B. Systems analysis of EGF receptor signaling dynamics with microwestern arrays. Nat. Methods 2010, 7, 148–155. [Google Scholar] [CrossRef]
- Zhou, D.H.; Wang, X.; Yang, M.; Shi, X.; Huang, W.; Feng, Q. Combination of low concentration of (−)-epigallocatechin gallate (EGCG) and curcumin strongly suppresses the growth of non-small cell lung cancer in vitro and in vivo through causing cell cycle arrest. Int. J. Mol. Sci. 2013, 14, 12023–12036. [Google Scholar] [CrossRef]
- Huang, W.Y.; Lai, C.H.; Peng, S.L.; Hsu, C.Y.; Hsu, P.H.; Chu, P.Y.; Feng, C.L.; Lin, Y.H. Targeting tumor cells with nanoparticles for enhanced co-drug delivery in cancer treatment. Pharmaceutics 2021, 13, 1327. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ho, C.-H.; Chen, M.-L.; Huang, H.-L.; Lai, C.-J.; Liu, C.-H.; Chuu, C.-P.; Lin, Y.-H. Active Targeting of P-Selectin by Fucoidan Modulates the Molecular Profiling of Metastasis in Docetaxel-Resistant Prostate Cancer. Mar. Drugs 2022, 20, 542. https://doi.org/10.3390/md20090542
Ho C-H, Chen M-L, Huang H-L, Lai C-J, Liu C-H, Chuu C-P, Lin Y-H. Active Targeting of P-Selectin by Fucoidan Modulates the Molecular Profiling of Metastasis in Docetaxel-Resistant Prostate Cancer. Marine Drugs. 2022; 20(9):542. https://doi.org/10.3390/md20090542
Chicago/Turabian StyleHo, Chang-Hsun, Mei-Lin Chen, Hau-Lun Huang, Chih-Jen Lai, Chih-Hsin Liu, Chih-Pin Chuu, and Yu-Hsin Lin. 2022. "Active Targeting of P-Selectin by Fucoidan Modulates the Molecular Profiling of Metastasis in Docetaxel-Resistant Prostate Cancer" Marine Drugs 20, no. 9: 542. https://doi.org/10.3390/md20090542