
Unusual Secondary Metabolites from the Mangrove Ecosystems: Structures, Bioactivities, Chemical, and Bio-Syntheses


Abstract
:
1. Introduction
2. Unusual Natural Products from Mangrove Flora
2.1. Limonoids
2.2. Diterpenoids
3. Unusual Natural Products from Mangrove-Associated Microorganisms
3.1. Polyketides
3.1.1. Coumarins and Isocoumarins
3.1.2. Chromones
3.1.3. Azaphilones
3.1.4. Benzophenones Derivatives
3.1.5. Macrolides
3.1.6. Others
3.2. Terpenoids
3.2.1. Sesquiterpenoids
3.2.2. Sesterterpenoids
3.2.3. Meroterpenoids
3.3. Alkaloids and Other Nitrogen-Containing Metabolites
3.3.1. Diketopiperazines
3.3.2. Indole and Isoindole Alkaloids Derivatives
3.3.3. Pyridines
3.3.4. Others
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zhou, J.; Feng, Z.; Zhang, W.; Xu, J. Evaluation of the antimicrobial and cytotoxic potential of endophytic fungi extracts from mangrove plants Rhizophora stylosa and R. mucronata. Sci. Rep. 2022, 12, 2733–2744. [Google Scholar] [CrossRef] [PubMed]
- Ser, H.L.; Tan, L.T.; Law, J.W.; Chan, K.-G.; Duangjai, A.; Saokaew, S.; Pusparajah, P.; Mutalib, N.-S.; Khan, T.M.; Goh, B.-H.; et al. Focused Review: Cytotoxic and Antioxidant Potentials of Mangrove-Derived Streptomyces. Front. Microbiol. 2017, 8, 2065–2075. [Google Scholar] [CrossRef]
- Azman, A.-S.; Othman, I.; Velu, S.S.; Chan, K.-G.; Lee, L.-H. Mangrove rare actinobacteria: Taxonomy, natural compound, and discovery of bioactivity. Front. Microbiol. 2015, 6, 856–870. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti, R.; Salvatore, M.M.; Andolfi, A. Secondary Metabolites of Mangrove-Associated Strains of Talaromyces. Mar. Drugs 2018, 16, 12–26. [Google Scholar] [CrossRef] [Green Version]
- Xu, J. Bioactive natural products derived from mangrove-associated microbes. RSC Adv. 2015, 5, 841–892. [Google Scholar] [CrossRef]
- Chen, S.; Cai, R.; Liu, Z.; Cui, H.; She, Z. Secondary metabolites from mangrove-associated fungi: Source, chemistry and bioactivities. Nat. Prod. Rep. 2022, 39, 560–595. [Google Scholar] [CrossRef]
- Ancheeva, E.; Daletos, G.; Proksch, P. Lead Compounds from Mangrove-Associated Microorganisms. Mar. Drugs 2018, 16, 319–349. [Google Scholar] [CrossRef] [Green Version]
- Zeng, W.; Huang, G.; Wang, B.; Cai, J.; Zheng, C. Secondary Metabolites and Bioactivities of Penicillium sp. Sourced from Mangrove from 2007 to 2020. Chin. J. Org. Chem. 2021, 41, 4255–4278. [Google Scholar] [CrossRef]
- Xu, D.B.; Ye, W.-W.; Han, Y.; Deng, Z.-X.; Hong, K. Natural products from mangrove actinomycetes. Mar. Drugs 2014, 12, 2590–2613. [Google Scholar] [CrossRef] [Green Version]
- Deshmukh, S.K.; Gupta, M.K.; Prakash, V.; Reddy, M.S. Mangrove-Associated Fungi: A Novel Source of Potential Anticancer Compounds. J. Fungi 2018, 4, 101–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Mao, Z.-G.; Song, B.-B.; Chen, C.-H.; Xiao, W.-W.; Hu, B.; Wang, J.-W.; Jiang, X.-B.; Zhu, Y.-H.; Wang, H.-J. Advances in the study of the structures and bioactivities of metabolites isolated from mangrove-derived fungi in the South China Sea. Mar. Drugs 2013, 11, 3601–3616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manohar, S.M. A Review of the Botany, Phytochemistry and Pharmacology of Mangrove Lumnitzera racemosa Willd. Pharmacogn. Rev. 2021, 15, 107–116. [Google Scholar] [CrossRef]
- Wang, K.-W.; Wang, S.-W.; Wu, B.; Wei, J.-G. Bioactive Natural Compounds from the Mangrove Endophytic Fungi. Mini. Rev. Med. Chem. 2014, 14, 370–391. [Google Scholar] [CrossRef] [PubMed]
- Cadamuro, R.D.; Bastos, I.M.A.S.; Silva, I.T.; Cruz, A.C.C.; Robl, D.; Sandjo, L.P.; Alves, S.; Lorenzo, J.M.; Rodríguez-Lázaro, D.; Treichel, H.; et al. Bioactive Compounds from Mangrove Endophytic Fungus and Their Uses for Microorganism Control. J. Fungi 2021, 7, 455–473. [Google Scholar] [CrossRef]
- Li, K.; Chen, S.; Pang, X.; Cai, J.; Zhang, X.; Liu, Y.; Zhu, Y.; Zhou, X. Natural products from mangrove sediments-derived microbes: Structural diversity, bioactivities, biosynthesis, and total synthesis. Eur. J. Med. Chem. 2022, 230, 114117–114157. [Google Scholar] [CrossRef]
- Wu, J.; Xiao, Q.; Xu, J.; Li, M.-Y.; Pan, J.-Y.; Yang, M.-H. Natural products from true mangrove flora: Source, chemistry and bioactivities. Nat. Prod. Rep. 2008, 25, 955–981. [Google Scholar] [CrossRef]
- Li, M.Y.; Xiao, Q.; Pan, J.-Y.; Wu, J. Natural products from semi-mangrove flora: Source, chemistry and bioactivities. Nat. Prod. Rep. 2009, 26, 281–298. [Google Scholar] [CrossRef]
- Fang, X.; Di, Y.T.; Hao, X.J. The Advances in the Limonoid Chemistry of the Meliaceae Family. Curr. Org. Chem. 2011, 15, 1363–1391. [Google Scholar] [CrossRef]
- Luo, J.; Sun, Y.; Kong, L. Research progress of meliaceous limonoids from 2011 to 2021. Nat. Prod. Rep. 2022, 39, 1325–1365. [Google Scholar] [CrossRef]
- Tan, Q.-G.; Luo, X.-D. Meliaceous limonoids: Chemistry and biological activities. Chem. Rev. 2011, 111, 7437–7522. [Google Scholar] [CrossRef]
- Li, J.; Li, M.-Y.; Bruhn, T.; Katele, F.X.; Xiao, Q.; Pedpradab, P.; Wu, J.; Bringmann, G. Thaixylomolins A-C: Limonoids Featuring Two New Motifs from the Thai Xylocarpus Moluccensis. Org. Lett. 2013, 15, 3682–3685. [Google Scholar] [CrossRef]
- Dai, Y.-G.; Li, W.-S.; Pedpradab, P.; Liu, J.-J.; Wu, J.; Shen, L. Thaixylomolins O–R: Four new limonoids from the Trang mangrove, Xylocarpus moluccensis. RSC Adv. 2016, 6, 85978–85984. [Google Scholar] [CrossRef]
- Wu, Y.-B.; Qing, X.; Huo, C.-H.; Yan, H.-M.; Shi, Q.-W.; Sauriol, F.; Gu, Y.-C.; Kiyota, H. Xylomexicanins E-H, new limonoids from Xylocarpus granatum. Tetrahedron 2014, 70, 4557–4562. [Google Scholar] [CrossRef]
- Pan, J.-Y.; Chen, S.-L.; Li, M.-Y.; Li, J.; Yang, M.-H.; Wu, J. Limonoids from the Seeds of a Hainan Mangrove, Xylocarpus Granatum. J. Nat. Prod. 2010, 73, 1672–1679. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, S.; Bruhn, T.; Xiao, Q.; Ding, H.; Bringmann, G. Xylogranatins F-R: Antifeedants from the Chinese mangrove, Xylocarpus granatum, a new biogenetic pathway to tetranortriterpenoids. Chem. Eur. J. 2008, 14, 1129–1144. [Google Scholar] [CrossRef]
- Sheng, Y.; Yin, S.; Wang, X.-N.; Lin, L.-P.; Ding, J.; Yue, J.-M. Xylogranatins A−D: Novel Tetranortriterpenoids with an Unusual 9,10-seco Scaffold from Marine Mangrove Xylocarpus granatum. Org. Lett. 2006, 8, 4935–4938. [Google Scholar] [CrossRef]
- Li, W.-S.; Shen, L.; Bruhn, T.; Pedpradab, P.; Wu, J.; Bringmann, G. Trangmolins A-F with an Unprecedented Structural Plasticity of the Rings A and B: New Insight into Limonoid Biosynthesis. Chem. Eur. J. 2016, 22, 11719–11727. [Google Scholar] [CrossRef]
- He, C.L.; Li, W.-S.; Wu, J.; Shen, L. Krishnolides E-K: New limonoids from the Krishna mangrove Xylocarpus moluccensis. Fitoterapia 2021, 150, 104835–104843. [Google Scholar] [CrossRef]
- Li, J.; Li, M.-Y.; Bruhn, T.; Gotz, D.C.G.; Xiao, Q.; Satyanandamurty, T.; Wu, J.; Bringmann, G. Andhraxylocarpins A-E: Structurally intriguing limonoids from the true mangroves Xylocarpus granatum and Xylocarpus moluccensis. Chem. Eur. J. 2012, 18, 14342–14351. [Google Scholar] [CrossRef]
- Li, W.-S.; Wu, J.; Li, J.; Satyanandamurty, T.; Shen, L.; Bringmann, G. Krishnadimer A, an Axially Chiral Non-biaryl Natural Product: Discovery and Biomimetic Synthesis. Org. Lett. 2017, 19, 182–185. [Google Scholar] [CrossRef]
- Li, W.-S.; Yang, Y.; Liu, J.-J.; Shen, L.; Shi, Z.; Wu, J. Scaffold diversity-oriented synthesis of limonoid dimers: Discovery of an axially chiral agent within vivoanti-breast cancer activity. Org. Chem. Front. 2018, 5, 1079–1091. [Google Scholar] [CrossRef]
- Zhou, Z.-F.; Liu, H.-L.; Zhang, W.; Kurtán, T.; Mándi, A.; Bényei, A.; Li, J.; Taglialatela-Scafati, O.; Guo, Y.-W. Bioactive rearranged limonoids from the Chinese mangrove Xylocarpus granatum Koenig. Tetrahedron 2014, 70, 6444–6449. [Google Scholar] [CrossRef]
- Wu, Y.-B.; Wang, Y.-Z.; Ni, Z.-Y.; Qing, X.; Shi, Q.-W.; Sauriol, F.; Vavricka, C.J.; Gu, Y.-C.; Kiyota, H. Xylomexicanins I and J: Limonoids with Unusual B/C Rings from Xylocarpus granatum. J. Nat. Prod. 2017, 80, 2547–2550. [Google Scholar] [CrossRef]
- Li, W.-S.; Mandi, A.; Liu, J.-J.; Shen, L.; Kurtan, T.; Wu, J. Xylomolones A-D from the Thai Mangrove Xylocarpus moluccensis: Assignment of Absolute Stereostructures and Unveiling a Convergent Strategy for Limonoid Biosynthesis. J. Org. Chem. 2019, 84, 2596–2606. [Google Scholar] [CrossRef] [PubMed]
- Ponnapalli, M.G.; Ankireddy, M.; Annam, S.C.V.A.R.; Ravirala, S.; Sukki, S.; Tuniki, V.R. Unusual ent-isopimarane-type diterpenoids from the wood of Excoecaria agallocha. Tetrahedron Lett. 2013, 54, 2942–2945. [Google Scholar] [CrossRef]
- Wang, H.; Li, M.-Y.; Katele, F.Z.; Satyanandamurty, T.; Wu, J.; Bringmann, G. Decandrinin, an unprecedented C9-spiro-fused 7,8-seco-ent-abietane from the Godavari mangrove Ceriops decandra. Beilstein J. Org. Chem. 2014, 10, 276–281. [Google Scholar] [CrossRef] [Green Version]
- Chooi, Y.-H.; Tang, Y. Navigating the fungal polyketide chemical space: From genes to molecules. J. Org. Chem. 2012, 77, 9933–9953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vederas, J.C. Explorations of fungal biosynthesis of reduced polyketides—A personal viewpoint. Nat. Prod. Rep. 2014, 31, 1253–1259. [Google Scholar] [CrossRef]
- Hang, L.; Liu, N.; Tang, Y. Coordinated and Iterative Enzyme Catalysis in Fungal Polyketide Biosynthesis. ACS Catal. 2016, 6, 5935–5945. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Guo, J.; Cheng, F.; Li, S. Cytochrome P450 enzymes in fungal natural product biosynthesis. Nat. Prod. Rep. 2021, 38, 1072–1099. [Google Scholar] [CrossRef]
- Little, R.F.; Hertweck, C. Chain release mechanisms in polyketide and non-ribosomal peptide biosynthesis. Nat. Prod. Rep. 2022, 39, 163–205. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, G.A.; Spillere, A.R.; Neves, G.M.; Kagami, L.P.; Poser, G.L.; Canto, R.F.S.; Eifler-Lima, V.L. Natural and synthetic coumarins as antileishmanial agents: A review. Eur. J. Med. Chem. 2020, 203, 112514–112533. [Google Scholar] [CrossRef] [PubMed]
- Shabir, G.; Saeed, A.; El-Seedi, H.R. Natural isocoumarins: Structural styles and biological activities, the revelations carry on. Phytochemistry 2021, 181, 112568–112590. [Google Scholar] [CrossRef]
- Cai, R.; Wu, Y.; Chen, S.; Cui, H.; Liu, Z.; Li, C.; She, Z. Peniisocoumarins A-J: Isocoumarins from Penicillium commune QQF-3, an Endophytic Fungus of the Mangrove Plant Kandelia candel. J. Nat. Prod. 2018, 81, 1376–1383. [Google Scholar] [CrossRef]
- Darsih, C.; Prachyawarakorn, V.; Wiyakrutta, S.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. Cytotoxic metabolites from the endophytic fungus Penicillium chermesinum: Discovery of a cysteine-targeted Michael acceptor as a pharmacophore for fragment-based drug discovery, bioconjugation and click reactions. RSC Adv. 2015, 5, 70595–70603. [Google Scholar] [CrossRef]
- Huang, J.; She, J.; Yang, X.; Liu, J.; Zhou, X.; Yang, B. A New Macrodiolide and Two New Polycyclic Chromones from the Fungus Penicillium sp. SCSIO041218. Molecules 2019, 24, 1686–1692. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.-M.; Yang, S.-X.; Qin, J.-C. Azaphilones: Chemistry and biology. Chem. Rev. 2013, 113, 4755–4811. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Li, D.; Peng, J.; Zhu, T.; Gu, Q.; Li, D. Penicitols A-C and penixanacid A from the mangrove-derived Penicillium chrysogenum HDN11-24. J. Nat. Prod. 2015, 78, 306–310. [Google Scholar] [CrossRef]
- Blanc, P.J.; Laussac, J.P.; Bars, J.L.; Bars, P.L.; Loret, M.O.; Pareilleux, A.; Prome, D.; Prome, J.C.; Santerre, A.L.; Goma, G. Characterization of monascidin A from Monascus as citrinin. Int. J. Food Microbiol. 1995, 27, 201–213. [Google Scholar] [CrossRef]
- Ohashi, T.; Hosokawa, S. Total Syntheses of Stoloniferol B and Penicitol A, and Structural Revision of Fusaraisochromanone. Org. Lett. 2018, 20, 3021–3024. [Google Scholar] [CrossRef]
- Wu, S.-B.; Long, C.; Kennelly, E.J. Structural diversity and bioactivities of natural benzophenones. Nat. Prod. Rep. 2014, 31, 1158–1174. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Liu, Y.; Li, J.; Huang, X.; Yan, T.; Cao, W.; Liu, H.; Long, Y.; She, Z. Diaporindenes A-D: Four Unusual 2,3-Dihydro-1 H-indene Analogues with Anti-inflammatory Activities from the Mangrove Endophytic Fungus Diaporthe sp. SYSU-HQ3. J. Org. Chem. 2018, 83, 11804–11813. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Ondeyka, J.G.; Herath, K.B.; Guan, Z.; Collado, J.; Platas, G.; Pelaez, F.; Leavitt, P.S.; Gurnett, A.; Nare, B.; et al. Tenellones A and B from a Diaporthe sp.: Two Highly Substituted Benzophenone Inhibitors of Parasite cGMP-Dependent Protein Kinase Activity. J. Nat. Prod. 2005, 68, 611–613. [Google Scholar] [CrossRef] [PubMed]
- Dinos, G.P. The macrolide antibiotic renaissance. Br. J. Pharmacol. 2017, 174, 2967–2983. [Google Scholar] [CrossRef]
- Meng, L.-H.; Li, X.-M.; Lv, C.-T.; Li, C.-S.; Xu, G.-M.; Huang, C.-G.; Wang, B.-G. Sulfur-containing cytotoxic curvularin macrolides from Penicillium sumatrense MA-92, a fungus obtained from the rhizosphere of the mangrove Lumnitzera racemosa. J. Nat. Prod. 2013, 76, 2145–2149. [Google Scholar] [CrossRef]
- Ding, L.; Maier, A.; Fiebig, H.-H.; Gorls, H.; Lin, W.-H.; Peschel, G.; Hertweck, G. Divergolides A-D from a mangrove endophyte reveal an unparalleled plasticity in ansa-macrolide biosynthesis. Angew. Chem. Int. Ed. 2011, 50, 1630–1634. [Google Scholar] [CrossRef]
- Wiley, P.F.; Kelly, R.B.; Caron, E.L.; Wiley, V.H.; Johnson, J.H.; MacKellar, F.A.; Mizsak, S.A. Structure of Nogalamycin. J. Am. Chem. Soc. 1977, 99, 542–549. [Google Scholar] [CrossRef]
- Pontius, A.; Krick, A.; Kehraus, S.; Brun, R.; König, G.M. Antiprotozoal Activities of Heterocyclic-Substituted Xanthones from the Marine-Derived Fungus Chaetomium sp. J. Nat. Prod. 2008, 71, 1579–1584. [Google Scholar] [CrossRef]
- Xu, Z.; Baunach, M.; Ding, L.; Peng, H.; Franke, J.; Hertweck, C. Biosynthetic code for divergolide assembly in a bacterial mangrove endophyte. ChemBioChem 2014, 15, 1274–1279. [Google Scholar] [CrossRef]
- Li, S.-R.; Zhao, G.-S.; Sun, M.-W.; He, H.-G.; Wang, H.-X.; Li, Y.-Y.; Lu, C.-H.; Shen, Y.-M. Identification and characterization of the biosynthetic gene cluster of divergolides from Streptomyces sp. W112. Gene 2014, 544, 93–99. [Google Scholar] [CrossRef]
- Zhao, G.; Li, S.; Guo, Z.; Sun, M.; Lu, C. Overexpression of div8 increases the production and diversity of divergolides in Streptomyces sp. W112. RSC Adv. 2015, 5, 98209–98214. [Google Scholar] [CrossRef]
- Zhao, G.; Wu, J.; Dai, W.-M. Toward a Total Synthesis of Divergolide, A.; Synthesis of the Amido Hydro-quinone Core and the C10–C15 Fragment. Synlett 2012, 23, 2845–2849. [Google Scholar] [CrossRef]
- Rasapalli, S.; Jarugumilli, G.; Yarrapothu, G.R.; Golen, J.A.; Rheingold, A.L. Studies toward Total Synthesis of Divergolides C and D. Org. Lett. 2013, 15, 1736–1739. [Google Scholar] [CrossRef] [PubMed]
- Nawrat, C.C.; Kitson, R.R.; Moody, C.J. Toward the total synthesis of hygrocin B and divergolide C: Construction of the naphthoquinone-azepinone core. Org. Lett. 2014, 16, 1896–1899. [Google Scholar] [CrossRef]
- Trauner, D.; Hager, A.; Kuttruff, C.; Hager, D.; Terwilliger, D. Toward the Total Synthesis of Divergolides C and D. Synlett 2013, 24, 1915–1920. [Google Scholar] [CrossRef]
- Rasapalli, S.; Jarugumilli, G.; Yarrapothu, G.R.; Golen, J.A.; Rheingold, A.L. Synthesis of the naphthoquinone core of divergolides (C–D) and model studies for elaboration of the ansabridge. Tetrahedron Lett. 2013, 54, 2615–2618. [Google Scholar] [CrossRef]
- Tian, E.-L.; Gu, B.-B.; Han, Y.; Qu, X.-D.; Lin, H.-W.; Deng, Z.-X.; Hong, K. Hainanmycin A, a cyclo-heptadeca macrolide bearing a cyclopentenone moiety from the mangrove-derived Streptomyces sp. 219807. Tetrahedron Lett. 2017, 58, 4348–4351. [Google Scholar] [CrossRef]
- Zhou, T.; Komaki, H.; Ichikawa, N.; Hosoyama, A.; Sato, S.; Igarashi, Y. Biosynthesis of akaeolide and lorneic acids and annotation of type I polyketide synthase gene clusters in the genome of Streptomyces sp. NPS554. Mar. Drugs 2015, 13, 581–596. [Google Scholar] [CrossRef] [Green Version]
- Choi, B.-K.; Park, S.-Y.; Choi, D.-K.; Shin, B.; Shin, Y.-H.; Oh, D.-C.; Lee, H.-S.; Lee, H.-S.; Lee, Y.-J.; Lee, J.S.; et al. Streptoglycerides A-D with a Rare 6/5/5 Tricyclic Ring Skeleton from a Marine Actinomycete Streptomyces species. Org. Lett. 2018, 20, 6037–6040. [Google Scholar] [CrossRef]
- Shin, H.J.; Heo, C.-S.; Anh, C.V.; Yoon, Y.D.; Kang, J.S. Streptoglycerides E-H, Unsaturated Polyketides from the Marine-Derived Bacterium Streptomyces specialis and Their Anti-Inflammatory Activity. Mar. Drugs 2022, 20, 44–55. [Google Scholar] [CrossRef]
- Choi, B.-K.; Cho, D.-Y.; Choi, D.-K.; Shin, H.J. Miharadienes A–D with unique cyclic skeletons from a marine-derived Streptomyces miharaensis. Org. Chem. Front. 2021, 8, 4845–4852. [Google Scholar] [CrossRef]
- Zhao, W.-Y.; Yi, J.; Chang, Y.-B.; Sun, C.-P.; Ma, X.-C. Recent studies on terpenoids in Aspergillus fungi: Chemical diversity, biosynthesis, and bioactivity. Phytochemistry 2022, 193, 113011–113035. [Google Scholar] [CrossRef]
- Jiang, M.; Wu, Z.; Guo, H.; Liu, L.; Chen, S. A Review of Terpenes from Marine-Derived Fungi: 2015–2019. Mar. Drugs 2020, 18, 321–368. [Google Scholar] [CrossRef] [PubMed]
- Wibowo, M.; Prachyawarakorn, V.; Aree, T.; Wiyakrutta, S.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. Tricyclic and Spirobicyclic Norsesquiterpenes from the Endophytic Fungus Pseudolagarobasidium acaciicola. Eur. J. Org. Chem. 2014, 19, 3976–3980. [Google Scholar] [CrossRef]
- Wibowo, M.; Prachyawarakorn, V.; Aree, T.; Mahidol, C.; Ruchirawat, S.; Kittakoopet, P. Cytotoxic sesquiterpenes from the endophytic fungus Pseudolagarobasidium acaciicola. Phytochemistry 2016, 122, 126–138. [Google Scholar] [CrossRef]
- Liu, D.-Z.; Dong, Z.-J.; Wang, F.; Liu, J.-K. Two novel norsesquiterpene peroxides from basidiomycete Steccherinum ochraceum. Tetrahedron Lett. 2010, 51, 3152–3153. [Google Scholar] [CrossRef]
- Meng, L.-H.; Li, X.-M.; Liu, Y.; Wang, B.-G. Penicibilaenes A and B, sesquiterpenes with a tricyclo[6.3.1.01,5]dodecane skeleton from the marine isolate of Penicillium bilaiae MA-267. Org. Lett. 2014, 16, 6052–6255. [Google Scholar] [CrossRef]
- Xue, Y.; Dong, G. Total Synthesis of Penicibilaenes via C-C Activation-Enabled Skeleton Deconstruction and Desaturation Relay-Mediated C-H Functionalization. J. Am. Chem. Soc. 2021, 143, 8272–8277. [Google Scholar] [CrossRef]
- Matsuo, R.; Watanabe, A.; Kamo, S.; Matsuzawa, A.; Sugita, K. Total syntheses of (±)-penicibilaenes A and B via intramolecular aldol condensation. Org. Chem. Front. 2021, 8, 6063–6066. [Google Scholar] [CrossRef]
- Huang, X.; Huang, H.; Li, H.; Sun, X.; Huang, H.; Lu, Y.; Lin, Y.; Long, Y.; She, Z. Asperterpenoid A, a New Sesterterpenoid as an Inhibitor of Mycobacterium tuberculosis Protein Tyrosine Phosphatase B from the Culture of Aspergillus sp. 16-5c. Org. Lett. 2013, 15, 721–723. [Google Scholar] [CrossRef]
- Xiao, Z.; Huang, H.; Shao, C.; Xia, X.; Ma, L.; Huang, X.; Lu, Y.; Lin, Y.; Long, Y.; She, Z. Asperterpenols A and B, New Sesterterpenoids Isolated from a Mangrove Endophytic Fungus Aspergillus sp. 085242. Org. Lett. 2013, 15, 2522–2525. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chen, Y.; Chen, S.; Liu, Y.; Lu, Y.; Chen, D.; Lin, Y.; Huang, X.; She, Z. Aspterpenacids A and B, Two Sesterterpenoids from a Mangrove Endophytic Fungus Aspergillus terreus H010. Org. Lett. 2016, 18, 1406–1409. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Wu, Z.; Liu, L.; Chen, S. The chemistry and biology of fungal meroterpenoids (2009–2019). Org. Biomol. Chem. 2021, 19, 1644–1704. [Google Scholar] [CrossRef]
- Meng, L.-H.; Li, X.-M.; Li, H.-L.; Wang, B.-G. Chermebilaenes A and B, New Bioactive Meroterpenoids from Co-Cultures of Marine-Derived Isolates of Penicillium bilaiae MA-267 and Penicillium chermesinum EN-480. Mar. Drugs 2020, 18, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-L.; Xu, R.; Li, X.-M.; Yang, S.-Q.; Meng, L.-H.; Wang, B.-G. Simpterpenoid A, a Meroterpenoid with a Highly Functionalized Cyclohexadiene Moiety Featuring gem-Propane-1,2-dione and Methylformate Groups, from the Mangrove-Derived Penicillium simplicissimum MA-332. Org. Lett. 2018, 20, 1465–1468. [Google Scholar] [CrossRef]
- Bai, M.; Zheng, C.-J.; Huang, G.-L.; Mei, R.-Q.; Wang, B.; Luo, Y.-P.; Zheng, C.; Niu, Z.-G.; Chen, G.-Y. Bioactive Meroterpenoids and Isocoumarins from the Mangrove-Derived Fungus Penicillium sp. TGM112. J. Nat. Prod. 2019, 82, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Horikoshi, A.; Tsuchida, M.; Tsujiuchi, T.; Oyama, K.; Mitomi, M. Substance, Its Production Method, Production Strain, and Agricultural and Horticultural Insecticide Containing the Same as an Active Ingredient. JP Patent JP-2010018586-A, 10 July 2008. [Google Scholar]
- Hemberger, Y.; Xu, J.; Wray, V.; Proksch, P.; Wu, J.; Bringmann, G. Pestalotiopens A and B: Stereochemically Challenging Flexible Sesquiterpene-Cyclopaldic Acid Hybrids from Pestalotiopsis sp. Chem. Eur. J. 2013, 19, 15556–15564. [Google Scholar] [CrossRef] [PubMed]
- Che, Q.; Zhu, T.; Qi, X.; Mándi, A.; Kurtán, T.; Mo, X.; Li, J.; Gu, Q.; Li, D. Hybrid Isoprenoids from a Reeds Rhizosphere Soil Derived Actinomycete Streptomyces sp. CHQ-64. Org. Lett. 2012, 14, 3438–3441. [Google Scholar] [CrossRef]
- Che, Q.; Zhu, T.; Keyzers, R.A.; Liu, X.; Li, J.; Gu, Q.; Li, D. Polycyclic hybrid isoprenoids from a reed rhizosphere soil derived Streptomyces sp. CHQ-64. J. Nat. Prod. 2013, 76, 759–763. [Google Scholar] [CrossRef]
- Che, Q.; Li, J.; Li, D.; Gu, Q.; Zhu, T. Structure and absolute configuration of drimentine I, an alkaloid from Streptomyces sp. CHQ-64. J. Antibiot. 2016, 69, 467–469. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Wang, Y.; Fu, P.; Chairoungdua, A.; Piyachaturawat, P.; Zhu, W. Secopaxilline A, an indole-diterpenoid derivative from an aciduric Penicillium fungus, its identification and semisynthesis. Org. Chem. Front. 2018, 5, 2835–2839. [Google Scholar] [CrossRef]
- Byrne, K.M.; Smith, S.K.; Ondeyka, J.G. Biosynthesis of Nodulisporic Acid A: Precursor Studies. J. Am. Chem. Soc. 2002, 124, 7055–7060. [Google Scholar] [CrossRef]
- Gao, S.-S.; Li, X.-M.; Williams, K.; Proksch, P.; Ji, N.-Y.; Wang, B.-G. Rhizovarins A-F, Indole-Diterpenes from the Mangrove-Derived Endophytic Fungus Mucor irregularis QEN-189. J. Nat. Prod. 2016, 79, 2066–2074. [Google Scholar] [CrossRef] [PubMed]
- Jesus, A.D.; Gorst-Allman, C.P.; Steyn, P.S.; Heerden, F.V.; Vleggaar, R.; Wessels, P.L.; Hull, W.E. Tremorgenic Mycotoxins from Penicillium crustosum. Biosynthesis of Penitrem A. J. Chem. Soc. 1983, 1863. [Google Scholar] [CrossRef]
- Hu, Y.; MacMillan, J.B. Erythrazoles A-B, Cytotoxic Benzothiazoles from a Marine-Derived Erythrobacter sp. Org. Lett. 2011, 13, 6580–6583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, R.M.; Yi, X.-X.; Zhou, Y.; Su, X.; Peng, Y.; Gao, C.-H. An update on 2,5-diketopiperazines from marine organisms. Mar. Drugs 2014, 12, 6213–6235. [Google Scholar] [CrossRef]
- Gao, H.; Liu, W.; Zhu, T.; Mo, X.; Mándi, A.; Kurtán, T.; Li, J.; Ai, J.; Gu, Q.; Li, D. Diketopiperazine alkaloids from a mangrove rhizosphere soil derived fungus Aspergillus effuses H1-1. Org. Biomol. Chem. 2012, 10, 9501–9506. [Google Scholar] [CrossRef] [Green Version]
- Meng, L.-H.; Wang, C.Y.; Mándi, A.; Li, X.-M.; Hu, X.-Y.; Kassack, M.U.; Kurtán, T.; Wang, B.-G. Three Diketopiperazine Alkaloids with Spirocyclic Skeletons and One Bisthiodiketopiperazine Derivative from the Mangrove-Derived Endophytic Fungus Penicillium brocae MA-231. Org. Lett. 2016, 18, 5304–5307. [Google Scholar] [CrossRef]
- Niu, S.; Liu, D.; Shao, Z.; Proksch, P.; Lin, W. Eutypellazines N−S, new thiodiketopiperazines from a deep sea sediment derived fungus Eutypella sp. with anti-VRE activities. Tetrahedron Lett. 2017, 58, 3695–3699. [Google Scholar] [CrossRef]
- Zhu, M.; Yang, Z.; Wang, H.; Gan, Q.; Zhang, G.; Che, Q.; Zhu, T.; Gu, Q.; Han, B.; Li, D. Penispirozines A-H, Three Classes of Dioxopiperazine Alkaloids with Spirocyclic Skeletons Isolated from the Mangrove-Derived Penicillium janthinellum. J. Nat. Prod. 2020, 83, 2647–2654. [Google Scholar] [CrossRef] [PubMed]
- Cai, R.; Jiang, H.; Xiao, Z.; Cao, W.; Yan, T.; Liu, Z.; Lin, S.; Long, Y.; She, Z. (−)- and (+)-Asperginulin A, a Pair of Indole Diketopiperazine Alkaloid Dimers with a 6/5/4/5/6 Pentacyclic Skeleton from the Mangrove Endophytic Fungus Aspergillus sp. SK-28. Org. Lett. 2019, 21, 9633–9636. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Sun, C.; Che, Q.; Zhu, T.; Gu, Q.; Guan, H.; Zhang, G.; Li, D. Pyrazinopyrimidine alkaloids from a mangrove-derived fungus Aspergillus versicolor HDN11-84. Phytochemistry 2021, 188, 112817–112823. [Google Scholar] [CrossRef]
- Duan, Y.; Liu, Y.; Huang, T.; Zou, Y.; Huang, T.; Hu, K.; Deng, Z.; Lin, S. Divergent biosynthesis of indole alkaloids FR900452 and spiro-maremycins. Org. Biomol. Chem. 2018, 16, 5446–5451. [Google Scholar] [CrossRef]
- Luo, X.; Chen, C.; Tao, H.; Lin, X.; Yang, B.; Zhou, X.; Liu, Y. Structurally diverse diketopiperazine alkaloids from the marine-derived fungus Aspergillus versicolor SCSIO 41016. Org. Chem. Front. 2019, 6, 736–740. [Google Scholar] [CrossRef]
- Huang, S.; Chen, H.; Li, W.; Zhu, X.; Ding, W.; Li, C. Bioactive Chaetoglobosins from the Mangrove Endophytic Fungus Penicillium chrysogenum. Mar. Drugs 2016, 14, 172–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Zhou, D.; Liang, F.; Wu, Z.; She, Z.; Li, C. Penochalasin K, a new unusual chaetoglobosin from the mangrove endophytic fungus Penicillium chrysogenum V11 and its effective semi-synthesis. Fitoterapia 2017, 123, 23–28. [Google Scholar] [CrossRef]
- Yang, B.; Dong, J.; Lin, X.; Zhou, X.; Zhang, Y.; Liu, Y. New prenylated indole alkaloids from fungus Penicillium sp. derived of mangrove soil sample. Tetrahedron 2014, 70, 3859–3863. [Google Scholar] [CrossRef]
- Cui, H.; Lin, Y.; Luo, M.; Lu, Y.; Huang, X.; She, Z. Diaporisoindoles A-C: Three Isoprenylisoindole Alkaloid Derivatives from the Mangrove Endophytic Fungus Diaporthe sp. SYSU-HQ3. Org. Lett. 2017, 19, 5621–5624. [Google Scholar] [CrossRef]
- Yu, G.; Zhou, G.; Zhu, M.; Wang, W.; Zhu, T.; Gu, Q.; Li, D. Neosartoryadins A and B, Fumiquinazoline Alkaloids from a Mangrove-Derived Fungus Neosartorya udagawae HDN13-313. Org. Lett. 2016, 18, 244–247. [Google Scholar] [CrossRef]
- Fu, P.; Yang, C.; Wang, Y.; Liu, P.; Ma, Y.; Xu, L.; Su, M.; Hong, K.; Zhu, W. Streptocarbazoles A and B, Two Novel Indolocarbazoles from the Marine-Derived Actinomycete Strain Streptomyces sp. FMA. Org. Lett. 2012, 14, 2422–2425. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Du, Y.; Cui, Q.; Zhang, J.; Zhu, W.; Hong, K.; Li, W. Cloning, characterization and heterologous expression of the indolocarbazole biosynthetic gene cluster from marine-derived Streptomyces sanyensis FMA. Mar. Drugs 2013, 11, 466–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Liang, Z.; Chen, W.; Luo, X.; Fang, W.; Liao, S.; Lin, X.; Yang, B.; Wang, J.; Tang, L.; et al. Iakyricidins A-D, antiproliferative piericidin analogs bearing carbonyl group or cyclic skeleton from Streptomyces iakyrus SCSIO NS104. J. Org. Chem. 2019, 84, 12626–12631. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Zhang, X.; Feng, H.; Che, Q.; Zhu, T.; Gu, Q.; Li, D. Campyridones A–D, pyridone alkaloids from a mangrove endophytic fungus Campylocarpon sp. HDN13-307. Tetrahedron 2016, 72, 5679–5683. [Google Scholar] [CrossRef]
- Zhou, Z.-F.; Kurtán, T.; Yang, X.-H.; Mándi, A.; Geng, M.-Y.; Ye, B.-P.; Taglialatela-Scafati, O.; Guo, Y.-W. Penibruguieramine A, a novel pyrrolizidine alkaloid from the endophytic fungus Penicillium sp. GD6 associated with Chinese mangrove Bruguiera gymnorrhiza. Org. Lett. 2014, 16, 1390–1393. [Google Scholar] [CrossRef]
- Chen, S.; He, L.; Chen, D.; Cai, R.; Long, Y.; Lu, Y.; She, Z. Talaramide A, an Unusual Alkaloid from the Mangrove Endophytic Fungus Talaromyces sp. (HZ-YX1) as Inhibitor of Mycobacterial PknG. New J. Chem. 2017, 4273–4277. [Google Scholar] [CrossRef]
- Gressler, M.; Zaehle, C.; Scherlach, K.; Hertweck, C.; Brock, M. Multifactorial induction of an orphan PKS-NRPS gene cluster in Aspergillus terreus. Chem. Biol. 2011, 18, 198–209. [Google Scholar] [CrossRef] [Green Version]


































































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Secondary Metabolites | Reported Activities | Ref |
|---|---|---|---|
| X. moluccensis | Thaixylomolin A–C (1, 19 and 20) | Anti-inflammatory | [21] |
| X. moluccensis | Thaixylomolin R (2) | NR a | [22] |
| X. granatum | Xylomexicanins E and F (23 and 3) | Cytotoxicity | [23] |
| X. granatum | Hainangrantum I and J (4 and 5) | NR | [24] |
| X. moluccensis | Trangmolins A–F (6–10, 17) | NR | [27] |
| X. moluccensis | Krishnolide J (11) | NR | [28] |
| X. moluccensis | Andhraxylocarpins A and C (12, 14) | NR | [29] |
| X. granatum | Andhraxylocarpins A–B and D–E (12–13 and 15–16) | NR | [29] |
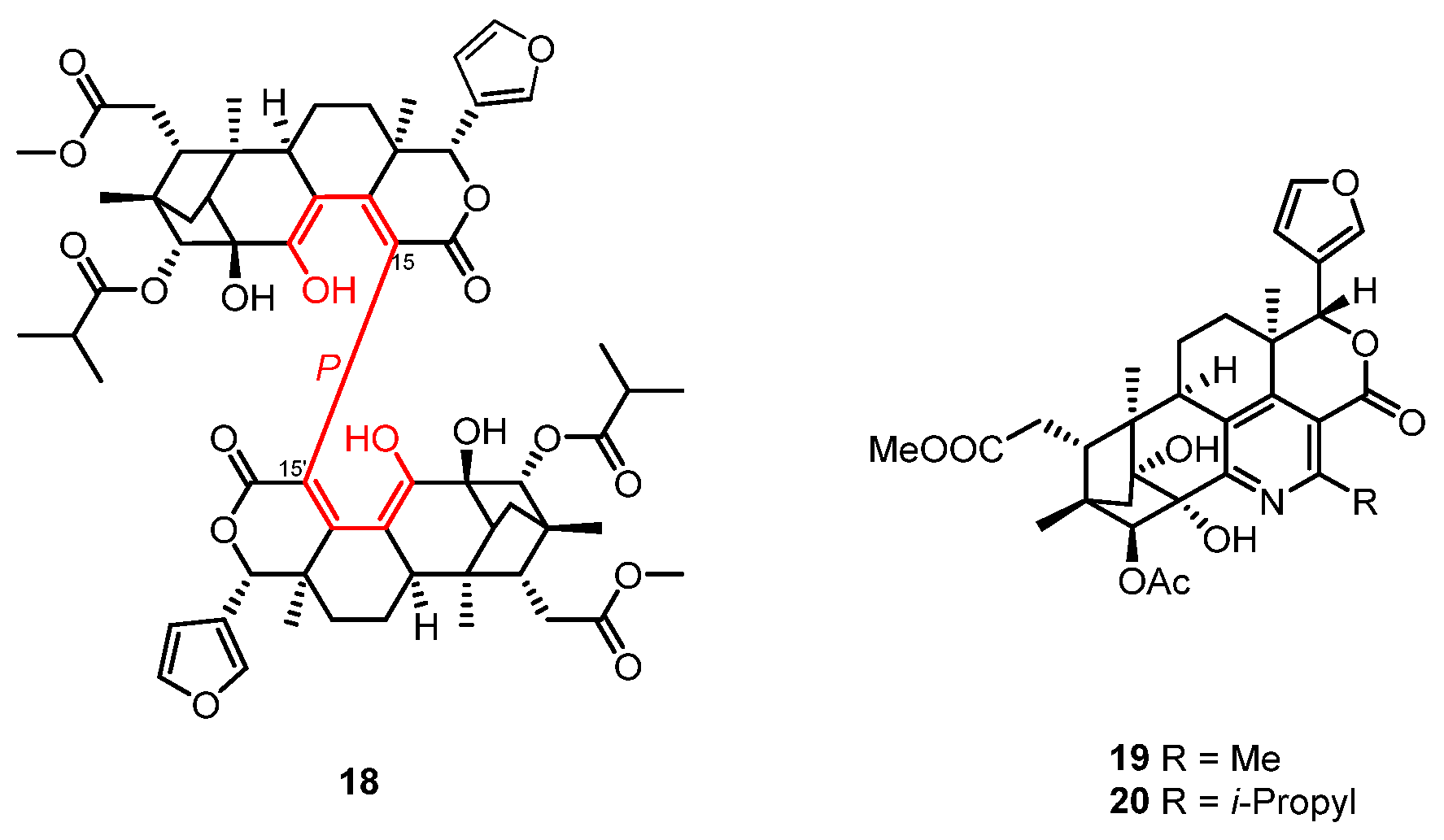
| X. moluccensis | Krishnadimer A (18) | NR | [30] |
| X. granatum | Xylogranatopyridines A and B (21 and 22) | PTP1B inhibitory | [32] |
| X. granatum | Xylomexicanins I and J (24 and 25) | NR | [33] |
| X. moluccensis | Xylomolones A–C (26–28) | NR | [34] |
| Excoecaria agallocha | Agallochaexceorins D–E (29–30) | NR | [35] |
| Ceriops decandra | Decandrinin (31) | NR | [36] |
| Microorganisms | Hosts | Secondary Metabolites | Reported Activities | Ref |
|---|---|---|---|---|
| Penicillium commune | Kandelia candel | Peniisocoumarins A and B (32 and 33) | NR a | [44] |
| Penicillium chermesinum | Mangrove forest | Penicilliumolide A (34) | NR | [45] |
| Penicillium sp. | Mangrove sediment | Penixanthones C and D (35 and 36) | NR | [46] |
| Penicillium chrysogenum | Acanthus ilicifolius | Penicitols A and B (37-38) | Cytotoxicity | [48] |
| Diaporthe sp. | Excoecaria agallocha | Diaporindenes A–D (39-42) | Anti-inflammatory | [52] |
| Diaporthe sp. | Excoecaria agallocha | Isoprenylisobenzofuran A (43) | Anti-inflammatory | [52] |
| Penicillium sumatrense | Lumnitzera racemose | Sumalarins A−C (44–46) | Cytotoxicity | [55] |
| Streptomyces sp. | Bruguiera gymnorrhiza | Divergolide A-D (47–50) | Antimicrobial, cytotoxicity | [56] |
| Streptomyces sp. | Mangrove soil | Hainanmycin A (51) | NR | [67] |
| Streptomyces sp. | Mangrove sample | Streptoglycerides A–H (52–59) | Anti-inflammatory | [69,70] |
| Streptomyces sp. | Mangrove sample | Miharadiened A–D (60–63) | NR | [71] |
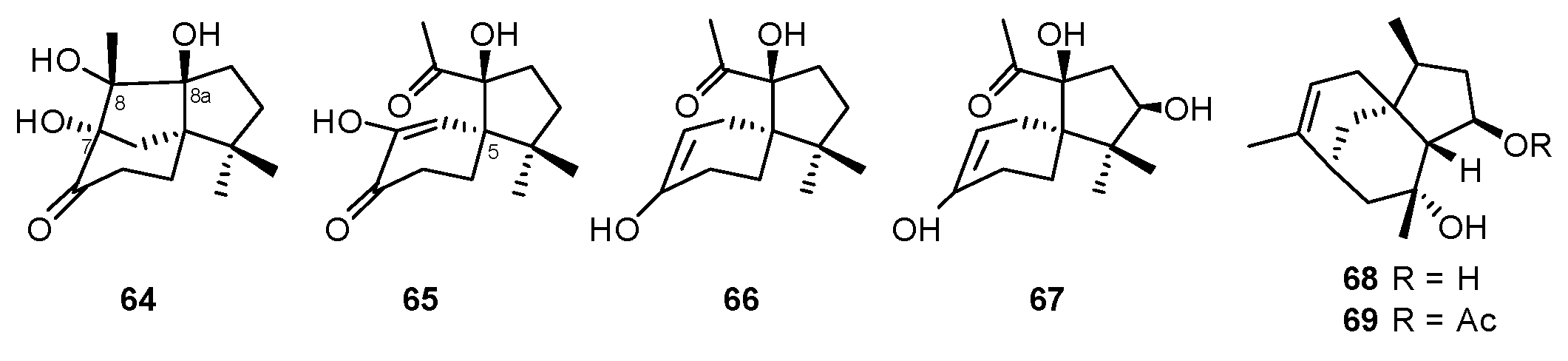
| Pseudolagarobasidium acaciicola | Bruguiera gymnorrhiza | Acaciicolin A (64), Spiroacaciicolides A–C (65–67) | NR | [74,75] |
| Penicillium bilaiae | Lumnitzera racemosa | Penicibilaenes A–B (68–69) | Anti-fungal | [77] |
| Aspergillus sp. | Mangrove endophytic | Asperterpenoid A (70) | Antituberculosis | [80] |
| Aspergillus sp. | Mangrove endophytic | Asperterpenols A–B (71–72) | Acetylcholinesterase inhibition | [81] |
| Aspergillus terreus | Kandelia obovate | Aspterpenacids A–B (73–74) | NR | [82] |
| Penicillium bilaiae | Lumnitzera racemosa | Chermebilaenes A–B (75–76) | Antibiotic | [84] |
| Penicillium simplicissimum | Bruguiera sexangula | Simpterpenoid A (77) | Antiviral | [85] |
| Penicillium sp. | Bruguiera sexangula | Penicianstinoids A–B (78–79) | Insecticidal activity | [86] |
| Pestalotiopsis sp. | Rhizophora mucronata | Pestalotiopens A–B (80–81) | NR | [88] |
| Streptomyces sp. | Mangrove rhizosphere soil | Indotertine A (82), indotertine B (83a/83b) | Antitumor | [89,90] |
| Streptomyces sp. | Mangrove rhizosphere soil | Drimentine I (84) | Antitumor | [91] |
| Penicillium camemberti | Rhizosphere soil of Rhizophora apiculata | Secopaxilline A (85) | NR | [92] |
| Mucor irregularis | Rhizophora stylosa | Rhizovarin A–D (86–89) | Antitumor | [94] |
| Erythrobacter sp. | Mangrove sediment | Erythrazoles A–B (90–91) | NR | [96] |
| Aspergillus effuses | Mangrove rhizosphere soil | Effusin A (92), dihydrocryptoechinulin D (93) | Cytotoxicity | [98] |
| Penicillium brocae | Avicennia marina | Spirobrocazines A–B (94–95) | NR | [99] |
| Eutypella sp. | Deep sea sediment | Eutypellazines N–P (96–98) | NR | [100] |
| Penicillium janthinellum | Mangrove | Penispirozines A–H (99–106) | Antioxidant | [101] |
| Aspergillus sp. | Mangrove endophytic | (±)-Asperginulin A (107a/b) | Antifouling | [102] |
| Aspergillus versicolor | Rhizosphere soil of Thespesia populnea | Pyrasplorines A–C (108–110) | NR | [103] |
| Penicillium chrysogenum | Mangrove endophytic | Penochalasin I and K (111–112) | Antibiotic, cytotoxicity | [106,107] |
| penicillium sp. | Mangrove sediment | Mangrovamides A–C (113–115) | Acetylcholinesterase inhibition | [108] |
| Diaporthe sp. | Excoecaria agallocha | Diaporisoindoles A–B (116–117), diaporisoindoles D–E (118–119), diaporisoindole C (120) | Antituberculosis, anti-inflammatory | [52,109] |
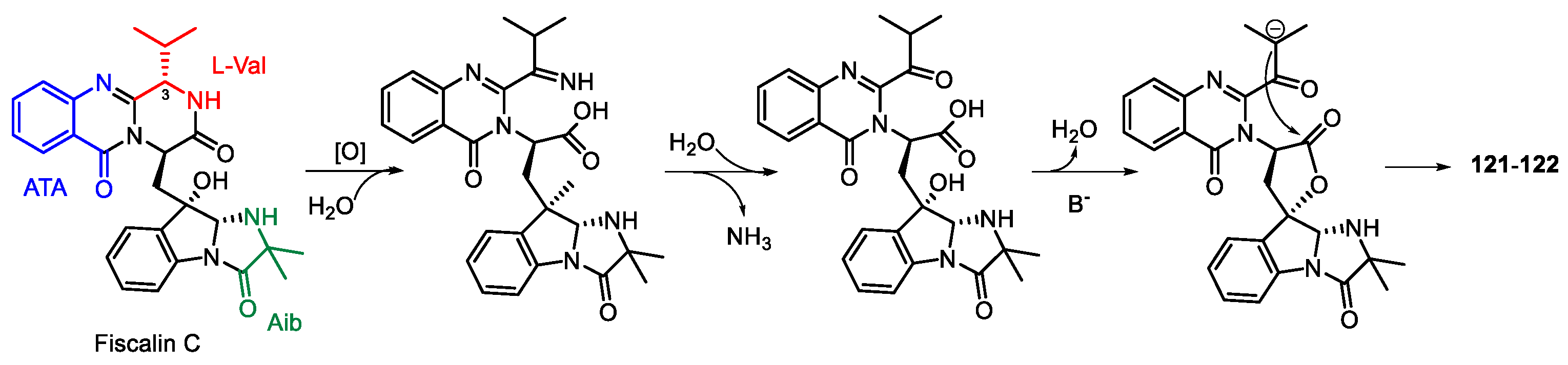
| Neosartorya udagawae | Mangrove endophytic | Neosartoryadins A–B (121–122) | NR | [110] |
| Streptomyces sp. | Mangrove soil | Streptocarbazoles A–B (123–124) | Antitumor | [111] |
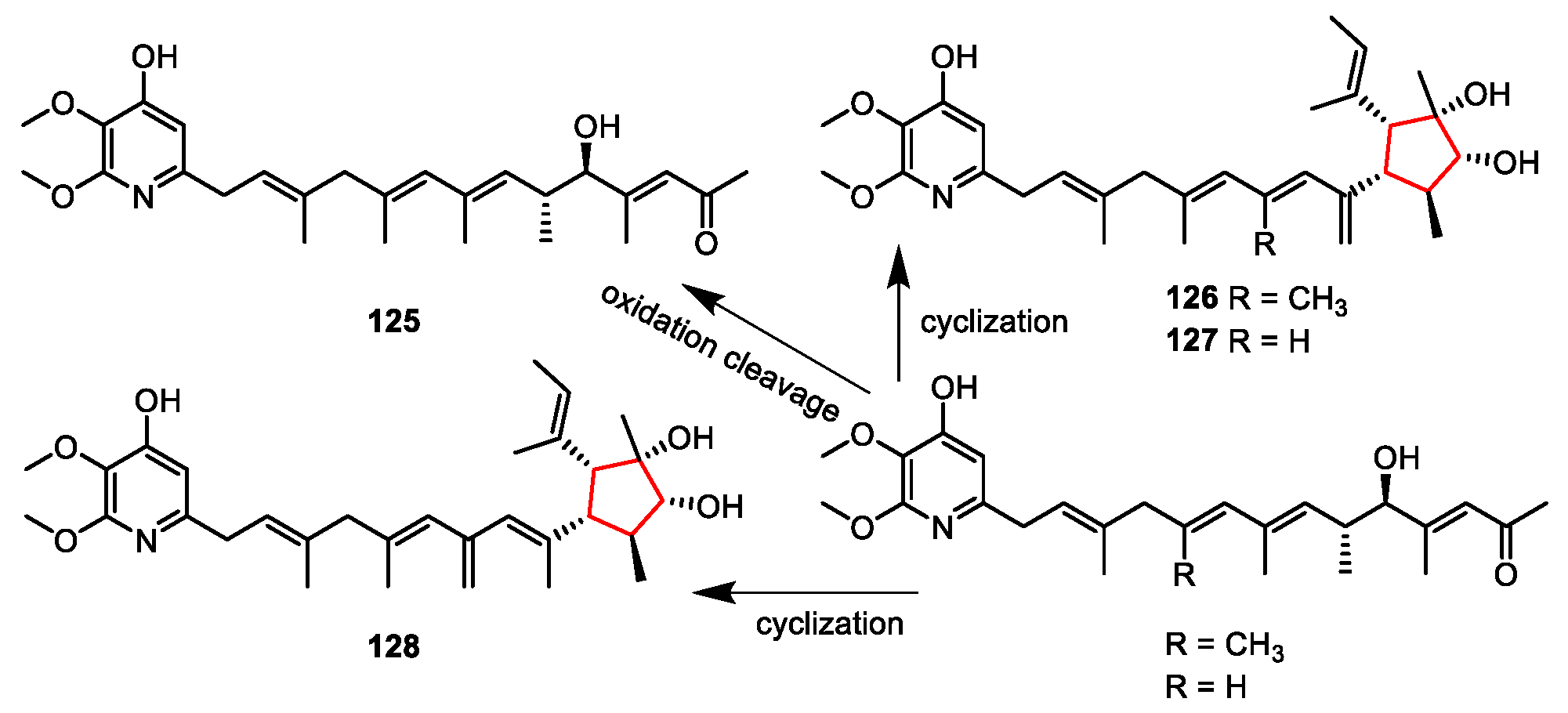
| Streptomyces iakyrus | Mangrove sediment | Iakyricidins A–D (125–128) | NR | [113] |
| Campylocarpon sp. | Sonneratia caseolaris | Campyridones A–D (129–132) | Antitumor | [114] |
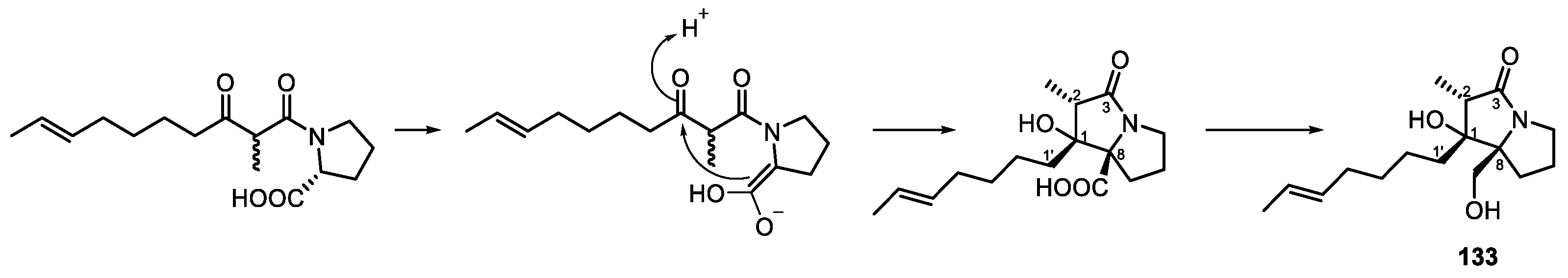
| Penicillium sp. | Bruguiera gymnorrhiza | Penibruguieramine A (133) | NR | [115] |
| Talaromyces sp. | Mangrove endophytic | Talaramide A (134) | NR | [116] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, M.-J.; Xu, B.; Guo, Y.-W. Unusual Secondary Metabolites from the Mangrove Ecosystems: Structures, Bioactivities, Chemical, and Bio-Syntheses. Mar. Drugs 2022, 20, 535. https://doi.org/10.3390/md20080535
Wu M-J, Xu B, Guo Y-W. Unusual Secondary Metabolites from the Mangrove Ecosystems: Structures, Bioactivities, Chemical, and Bio-Syntheses. Marine Drugs. 2022; 20(8):535. https://doi.org/10.3390/md20080535
Chicago/Turabian StyleWu, Meng-Jun, Baofu Xu, and Yue-Wei Guo. 2022. "Unusual Secondary Metabolites from the Mangrove Ecosystems: Structures, Bioactivities, Chemical, and Bio-Syntheses" Marine Drugs 20, no. 8: 535. https://doi.org/10.3390/md20080535