
The Discovery, Enzymatic Characterization and Functional Analysis of a Newly Isolated Chitinase from Marine-Derived Fungus Aspergillus fumigatus df347

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Identification of ITS rDNA Sequence of the Strain
2.2. Sequence Analysis of AfChi28
2.3. Gene Coning of AfChi28
2.4. Expression and Purification of AfChi28
2.5. Enzyme Activity Assay of AfChi28
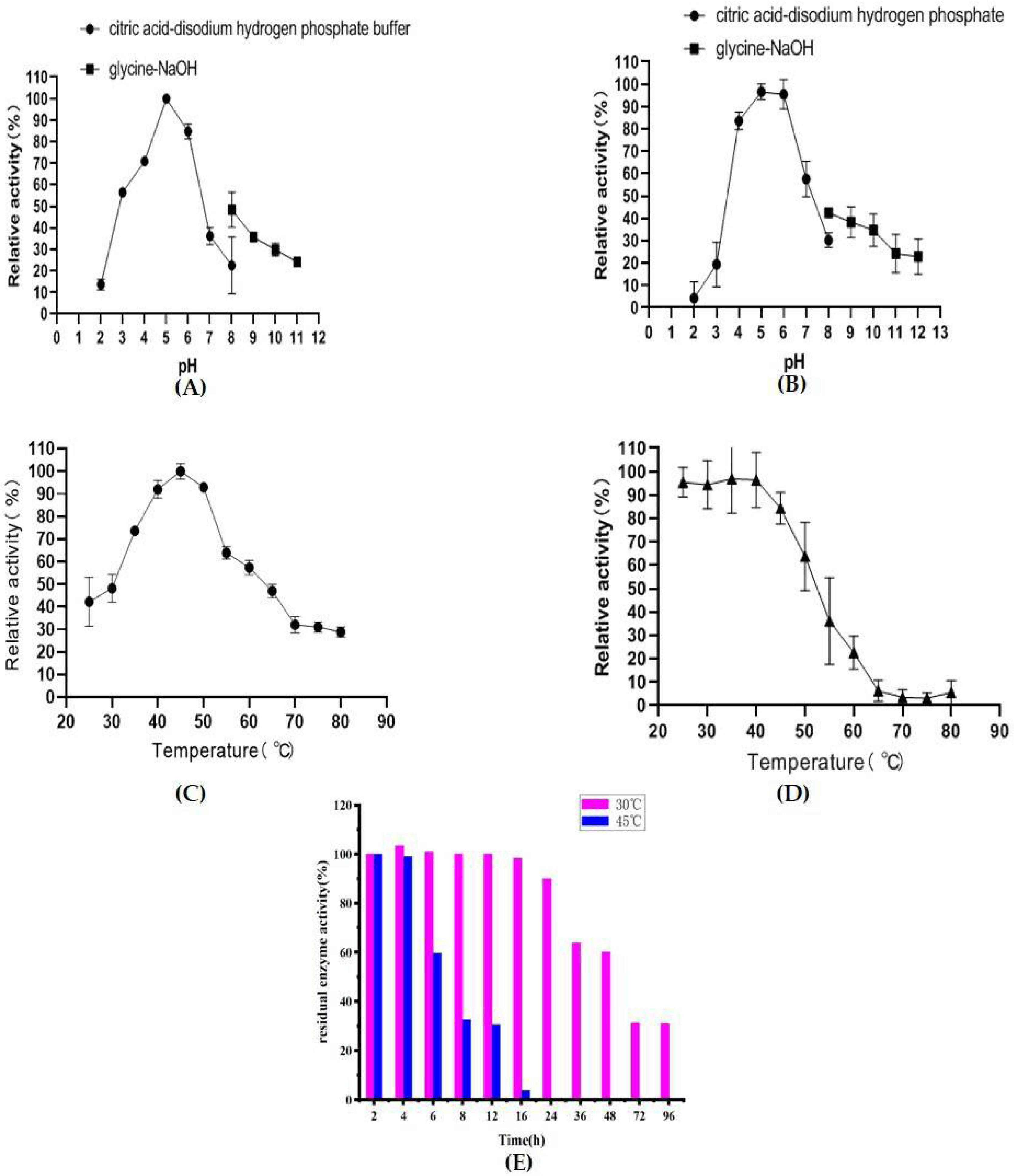
2.6. Biochemical Characterization of AfChi28
2.7. Effect of Metal Ions and Chemical Reagents on Enzyme Activity
2.8. The Substrate Specificity of AfChi28
2.9. Kinetic Parameters
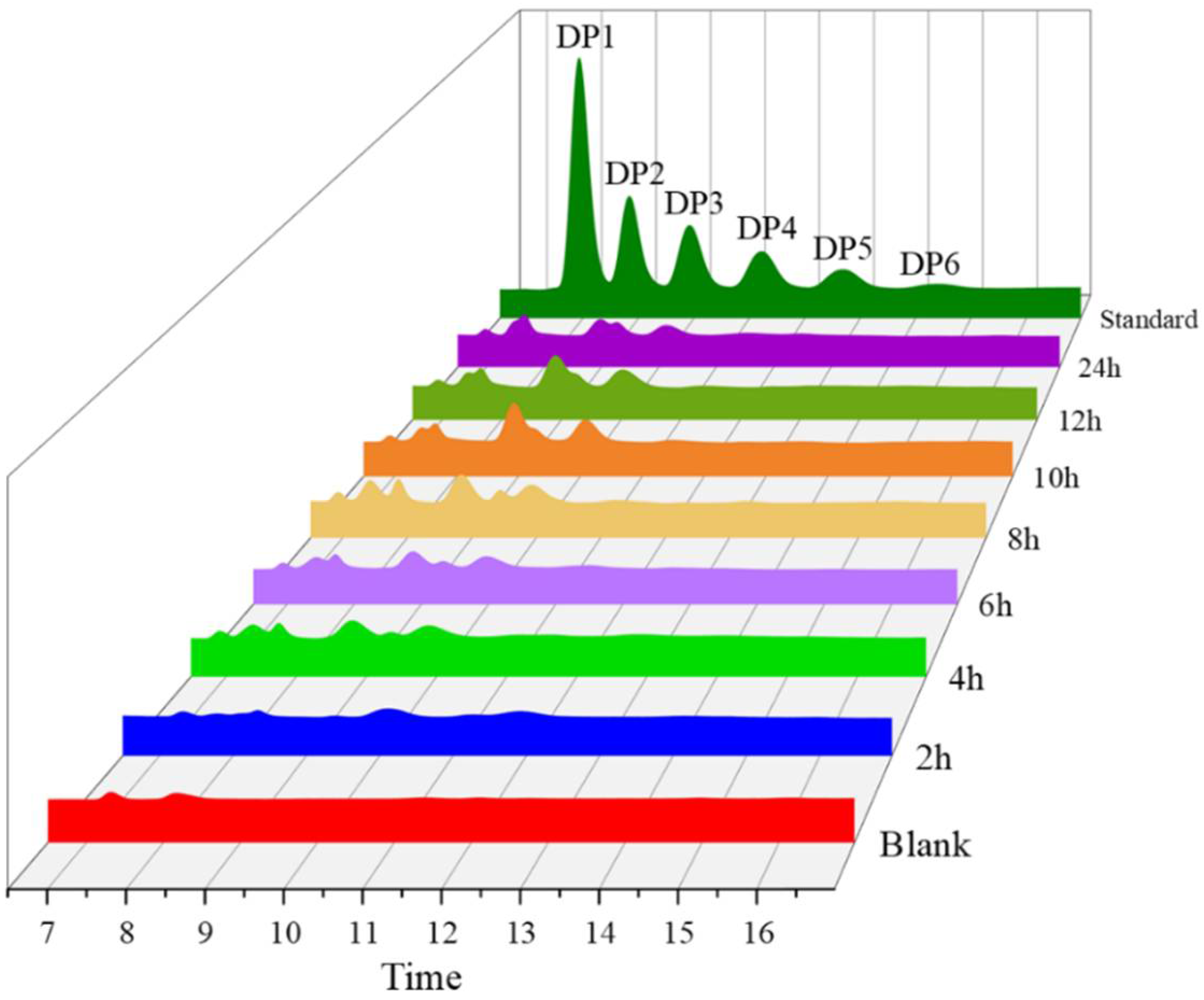
2.10. Hydrolysis Patterns of AfChi28
2.11. HPLC and Q Exactive LC-MS Analysis of Hydrolysates Produced by AfChi28
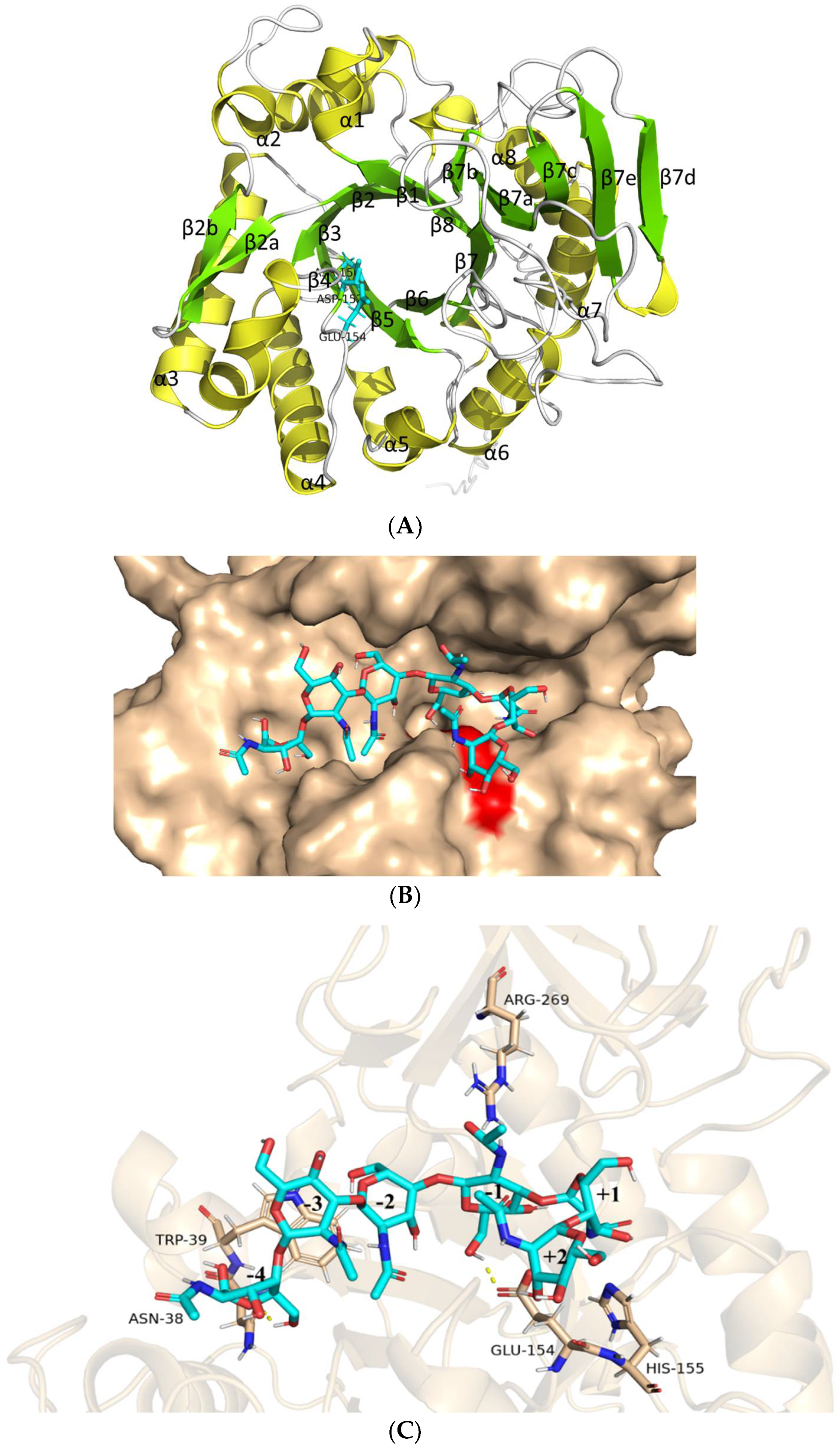
2.12. Structural Analysis of AfChi28 and Molecular Docking Simulation
3. Materials and Methods
3.1. Strains and Materials
3.2. Sequence Analysis
3.3. Isolation of Strains with Chitinolytic Activity
3.4. Strain Identification
3.5. Preparation of cDNA
3.6. Gene Cloning of AfChi28
3.7. Construction of Expression Vector in E. coli BL21 Codon plus (DE3) RIL
3.8. Purification of Recombinant AfChi28
3.9. Enzyme Activity Assay of AfChi28
3.10. Biochemical Characteristics of AfChi28
3.11. Effect of Metal Ions and Various Chemicals on Enzyme Activity
3.12. Substrate Specificity and Kinetic Parameters
3.13. TLC Analysis of Colloidal Chitin Hydrolysates Produced by AfChi28
3.14. HPLC and Q Exactive LC-MS Analysis of Colloidal Chitin Hydrolysates Produced by AfChi28
3.15. Structure Prediction and Molecular Docking
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Wan, A.C.A.; Tai, B.C.U. CHITIN—A Promising Biomaterial for Tissue Engineering and Stem Cell Technologies. Biotechnol. Adv. 2013, 31, 1776–1785. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, H.; Velayutham, K.; Ravichandran, R. Chitin and Chitosan Preparation from Shrimp Shells Penaeus Monodon and Its Human Ovarian Cancer Cell Line, PA-1. Int. J. Biol. Macromol. 2018, 107, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Song, Y.; Han, Y.S.; Jo, Y.H.; Choi, M.H.; Park, Y.; Kang, S.H.; Kim, S.; Choi, C.; Jung, W. Production of Chitin and Chitosan from the Exoskeleton of Adult Two-Spotted Field Crickets (Gryllus Bimaculatus). Entomol. Res. 2017, 47, 279–285. [Google Scholar] [CrossRef]
- Crini, G.; Hatchett, C. Historical Review on Chitin and Chitosan Biopolymers. Environ. Chem. Lett. 2019, 17, 1623–1643. [Google Scholar] [CrossRef]
- Rahman, M.A.; Halfar, J. First Evidence of Chitin in Calcified Coralline Algae: New Insights into the Calcification Process of Clathromorphum Compactum. Sci. Rep. 2014, 4, 6162. [Google Scholar] [CrossRef]
- Yadav, M.; Goswami, P.; Paritosh, K.; Kumar, M.; Pareek, N.; Vivekanand, V. Seafood Waste: A Source for Preparation of Commercially Employable Chitin/Chitosan Materials. Bioresour. Bioprocess. 2019, 6, 8. [Google Scholar] [CrossRef]
- Souza, C.P.; Almeida, B.C.; Colwell, R.R.; Rivera, I.N.G. The Importance of Chitin in the Marine Environment. Mar. Biotechnol. 2011, 13, 823–830. [Google Scholar] [CrossRef]
- Dhillon, G.S.; Kaur, S.; Brar, S.K.; Verma, M. Reen Synthesis Approach: Extraction of Chitosan from Fungus Mycelia. Crit. Rev. Biotechnol. 2012, 33, 379–403. [Google Scholar] [CrossRef]
- Aranaz, I.; Acosta, N.; Civera, C.; Elorza, B.; Mingo, J.; Castro, C.; Gandía, M.D.l.L.; Caballero, A.H. Cosmetics and Cosmeceutical Applications of Chitin, Chitosan and Their Derivatives. Polymers 2018, 10, 213. [Google Scholar] [CrossRef]
- Parhi, R. Drug Delivery Applications of Chitin and Chitosan: A Review. Environ. Chem. Lett. 2020, 18, 577–594. [Google Scholar] [CrossRef]
- Ueda, M.; Shioyama, T.; Nakadoi, K.; Nakazawa, M.; Sakamoto, T.; Iwamoto, T.; Sakaguchi, M. Cloning and Expression of a Chitinase Gene from Eisenia Fetida. Int. J. Biol. Macromol. 2017, 105, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Xia, W.; Yu, X. Purification and Characterization of Two Types of Chitosanase from Aspergillus Sp. CJ22-326. Food Res. Int. 2005, 38, 315–322. [Google Scholar] [CrossRef]
- Cohen-Kupiec, R.; Chet, I. The Molecular Biology of Chitin Digestion. Curr. Opin. Biotechnol. 1998, 9, 270–277. [Google Scholar] [CrossRef]
- Ding, B.; Huang, S.; Shen, K.; Hou, J.; Gao, H.; Duan, Y. Natural Rubber Bio-Nanocomposites Reinforced with Self-Assembled Chitin Nano Fi Bers from Aqueous KOH/Urea Solution. Carbohydr. Polym. 2019, 225, 115230. [Google Scholar] [CrossRef]
- Fang, Y.; Zhang, R.; Duan, B.; Liu, M.; Lu, A.; Zhang, L. Recyclable Universal Solvents for Chitin to Chitosan with Various Degrees of Acetylation and Construction of Robust Hydrogels. ACS Sustain. Chem. Eng. 2017, 5, 2725–2733. [Google Scholar] [CrossRef]
- Lin, C.W.; Lin, J.C. Characterization and Blood Coagulation Evaluation of the Water-Soluble Chitooligosaccharides Prepared by a Facile Fractionation Method. Biomacromolecules 2003, 4, 1691–1697. [Google Scholar] [CrossRef]
- Chang, K.L.B.; Tai, M.C.; Cheng, F.H. Kinetics and Products of the Degradation of Chitosan by Hydrogen Peroxide. J. Agric. Food Chem. 2001, 49, 4845–4851. [Google Scholar] [CrossRef]
- Adams, C.A.; Andrews, J.E.; Jickells, T. Nitrous Oxide and Methane Fluxes vs. Carbon, Nitrogen and Phosphorous Burial in New Intertidal and Saltmarsh Sediments. Sci. Total Environ. 2012, 434, 240–251. [Google Scholar] [CrossRef]
- Xing, R.; Liu, S.; Yu, H.; Guo, Z.; Wang, P.; Li, C.; Li, Z.; Li, P. Salt-Assisted Acid Hydrolysis of Chitosan to Oligomers under Microwave Irradiation. Carbohydr. Res. 2005, 340, 2150–2153. [Google Scholar] [CrossRef]
- Qin, Z.; Luo, S.; Li, Y.; Chen, Q.; Qiu, Y.; Zhao, L.; Jiang, L.; Zhou, J. Biochemical Properties of a Novel Chitosanase from Bacillus Amyloliquefaciens and Its Use in Membrane Reactor. Lwt 2018, 97, 9–16. [Google Scholar] [CrossRef]
- Zhou, J.; Liu, X.; Yuan, F.; Deng, B.; Yu, X. Biocatalysis of Heterogenously-Expressed Chitosanase for the Preparation of Desirable Chitosan Oligosaccharides Applied against Phytopathogenic Fungi. ACS Sustain. Chem. Eng. 2020, 8, 4781–4791. [Google Scholar] [CrossRef]
- Yang, S.; Fu, X.; Yan, Q.; Jiang, Z.; Wang, J. Biochemical Characterization of a Novel Acidic Exochitinase from Rhizomucor Miehei with Antifungal Activity. J. Agric. Food Chem. 2016, 64, 461–469. [Google Scholar] [CrossRef]
- Dai, Y.; Yang, F.; Liu, X.; Wang, H. The Discovery and Characterization of a Novel Chitinase with Dual Catalytic Domains from a Qinghai-Tibetan Plateau Wetland Soil Metagenome. Int. J. Biol. Macromol. 2021, 188, 482–490. [Google Scholar] [CrossRef]
- Hartl, L.; Zach, S.; Seidl-seiboth, V. Fungal Chitinases: Diversity, Mechanistic Properties and Biotechnological Potential. Appl. Microbiol. Biotechnol. 2012, 533–543. [Google Scholar] [CrossRef]
- Deng, J.J.; Shi, D.; Mao, H.H.; Li, Z.W.; Liang, S.; Ke, Y.; Luo, X.C. Heterologous Expression and Characterization of an Antifungal Chitinase (Chit46) from Trichoderma Harzianum GIM 3.442 and Its Application in Colloidal Chitin Conversion. Int. J. Biol. Macromol. 2019, 134, 113–121. [Google Scholar] [CrossRef]
- Ohnuma, T.; Numata, T.; Osawa, T.; Inanaga, H.; Okazaki, Y.; Shinya, S.; Kondo, K.; Fukuda, T.; Fukamizo, T. Crystal Structure and Chitin Oligosaccharide-Binding Mode of a ′Loopful’ Family GH19 Chitinase from Rye, Secale Cereale, Seeds. FEBS J. 2012, 279, 3639–3651. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, Y.; Ma, J.; Yan, Q.; Jiang, Z. Biochemical Characterization of a Bifunctional Chitinase/Lysozyme from Streptomyces Sampsonii Suitable for N-Acetyl Chitobiose Production. Biotechnol. Lett. 2020, 42, 1489–1499. [Google Scholar] [CrossRef]
- Beygmoradi, A.; Homaei, A.; Hemmati, R.; Santos-Moriano, P.; Hormigo, D.; Fernández-Lucas, J. Marine Chitinolytic Enzymes, a Biotechnological Treasure Hidden in the Ocean? Appl. Microbiol. Biotechnol. 2018, 102, 9937–9948. [Google Scholar] [CrossRef]
- Schmitz, C.; Auza, L.G.; Koberidze, D.; Rasche, S.; Fischer, R.; Bortesi, L. Conversion of Chitin to Defined Chitosan Oligomers: Current Status and Future Prospects. Mar. Drugs 2019, 17, 452. [Google Scholar] [CrossRef]
- Xie, X.; Fu, X.; Yan, X.; Peng, W.; Kang, L. A Broad-Specificity Chitinase from Penicillium Oxalicum K10 Exhibits Antifungal Activity and Biodegradation Properties of Chitin. Mar. Drugs 2021, 19, 356. [Google Scholar] [CrossRef]
- Tanaka, H.; Akutsu, H.; Yabuta, I.; Hara, M.; Sugimoto, H.; Ikegami, T.; Watanabe, T.; Fujiwara, T. A Novel Chitin-Binding Mode of the Chitin-Binding Domain of Chitinase A1 from Bacillus Circulans WL-12 Revealed by Solid-State NMR. FEBS Lett. 2018, 592, 3173–3182. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, J.; Liu, X.; Pan, X.; Hou, J.; Ran, C.; Zhou, Z. Improving Extracellular Production of Serratia Marcescens Lytic Polysaccharide Monooxygenase CBP21 and Aeromonas Veronii B565 Chitinase Chi92 in Escherichia Coli and Their Synergism. AMB Express 2017, 7, 170. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Fong, S.S. Bacterial Chitinase: Nature and Perspectives for Sustainable Bioproduction. Bioresour. Bioprocess. 2015, 2, 31. [Google Scholar] [CrossRef]
- Makhdoumi-Kakhki, A.; Amoozegar, M.A.; Ventosa, A. Salinibacter Iranicus Sp. Nov. and Salinibacter Luteus Sp. Nov., Isolated from a Salt Lake, and Emended Descriptions of the Genus Salinibacter and of Salinibacter Ruber. Int. J. Syst. Evol. Microbiol. 2012, 62, 1521–1527. [Google Scholar] [CrossRef] [PubMed]
- Qu, T.; Zhang, C.; Qin, Z.; Fan, L.; Jiang, L.; Zhao, L. A Novel GH Family 20 β-N-Acetylhexosaminidase With Both Chitosanase and Chitinase Activity From Aspergillus Oryzae. Front. Mol. Biosci. 2021, 8, 432. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Lan, X.; Li, X.; Huang, L.; Zhang, Y.; Wang, Z. Protein Expression and Puri Fi Cation High-Level Expression and Characterization of a Stereoselective Lipase from Aspergillus Oryzae in Pichia Pastoris. Protein Expr. Purif. 2019, 155, 1–7. [Google Scholar] [CrossRef]
- Xia, J.L.; Xiong, J.; Xu, T.; Zhang, C.G.; Zhang, R.Y.; Zhang, Q.; Wu, S.; Qiu, G.Z. Purification and Characterization of Extracellular Chitinase from a Novel Strain Aspergillus Fumigatus CS-01. J. Cent. South Univ. Technol. 2009, 16, 552–557. [Google Scholar] [CrossRef]
- Feller, I.C.; Lovelock, C.E.; Berger, U.; McKee, K.L.; Joye, S.B.; Ball, M.C. Biocomplexity in Mangrove Ecosystems. Annu. Rev. Mar. Sci. 2010, 2, 395–417. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Wang, M.; Fu, H.; Tang, C.; Wang, W. Predicting Changes in Molluscan Spatial Distributions in Mangrove Forests in Response to Sea-Level Rise. Ecol. Evolut. 2022, 12, e9033. [Google Scholar] [CrossRef]
- Kathiresan, K.; Bingham, B.L. Biology of mangroves and mangrove Ecosystems. In Advances in Marine Biology; Academic Press: London, UK, 2001; pp. 81–251. ISBN 0120261405. [Google Scholar]
- Ihrmark, K.; Asmail, N.; Ubhayasekera, W.; Melin, P.; Stenlid, J.; Karlsson, M. Comparative Molecular Evolution of Trichoderma Chitinases in Response to Mycoparasitic Interactions. Evol. Bioinform. 2010, 6, EBO-S4198. [Google Scholar] [CrossRef] [PubMed]
- Alcazar-Fuoli, L.; Clavaud, C.; Lamarre, C.; Aimanianda, V.; Seidl-Seiboth, V.; Mellado, E.; Latgé, J.P. Functional Analysis of the Fungal/Plant Class Chitinase Family in Aspergillus Fumigatus. Fungal Genet. Biol. 2011, 48, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI Search and Sequence Analysis Tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef]
- Muth, H. Fritz Schales 1916–1976. Radiat. Environ. Biophys. 1976, 13, 271–272. [Google Scholar] [CrossRef]
- Bouacem, K.; Laribi-Habchi, H.; Mechri, S.; Hacene, H.; Jaouadi, B.; Bouanane-Darenfed, A. Biochemical Characterization of a Novel Thermostable Chitinase from Hydrogenophilus Hirschii Strain KB-DZ44. Int. J. Biol. Macromol. 2018, 106, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Li, A.; Han, H.; Liu, T.; Yang, Q. A Potent Chitinase from Bacillus Subtilis for the Ef Fi Cient Bioconversion of Chitin-Containing Wastes. Int. J. Biol. Macromol. 2018, 116, 863–868. [Google Scholar] [CrossRef] [PubMed]
- Xia, G.; Jin, C.; Zhou, J.; Yang, S.; Zhang, S.; Jin, C. A Novel Chitinase Having a Unique Mode of Action from Aspergillus Fumigatus YJ-407. Eur. J. Biochem. 2001, 268, 4079–4085. [Google Scholar] [CrossRef]
- Wang, Y. Simulations of a Novel Recombinant Chitinase from Chitinibacter Tainanensis CT01 Appropriate for Chitin Enzymatic Hydrolysis. Polymers 2020, 12, 1648. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Yu, M.; Wu, Y.; Ran, L.; Liu, W.; Zhang, X.H. Two Highly Similar Chitinases from Marine Vibrio Species Have Different Enzymatic Properties. Mar. Drugs 2020, 18, 139. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Chen, W.T.; Lin, Z.H.; Kuo, Y.H.; Nguyen, A.D.; Pan, P.S.; Wang, S.L. An Amphiprotic Novel Chitosanase from Bacillus Mycoides and Its Application in the Production of Chitooligomers with Their Antioxidant and Anti-Inflammatory Evaluation. Int. J. Mol. Sci. 2016, 17, 1302. [Google Scholar] [CrossRef]
- Liu, C.; Shen, N.; Wu, J.; Jiang, M.; Shi, S. Cloning, Expression and Characterization of a Chitinase from Paenibacillus Chitinolyticus Strain UMBR 0002. PeerJ 2020, 8, e8964. [Google Scholar] [CrossRef] [PubMed]
- Kidibule, P.E.; Moriano, P.S.; Ortega, E.J.; Escudero, M.R.; Limón, M.C.; Remacha, M.; Plou, F.J.; Aparicio, J.S. Use of Chitin and Chitosan to Produce New Chitooligosaccharides by Chitinase Chit42: Enzymatic Activity and Structural Basis of Protein Specificity. Microb. Cell Factor. 2018, 17, 47. [Google Scholar] [CrossRef] [PubMed]
- Cramer, P. AlphaFold2 and the Future of Structural Biology. Nat. Struct. Mol. Biol. 2021, 28, 704–705. [Google Scholar] [CrossRef]
- Suzuki, K.; Taiyoji, M.; Sugawara, N.; Nikaidou, N.; Henrissat, B.; Watanabe, T. Relationship of Its Product To Other Bacterial Chitinases. Society 1999, 596, 587–596. [Google Scholar]
- Liu, T.; Han, H.; Wang, D.; Guo, X.; Zhou, Y.; Fukamizo, T.; Yang, Q. Potent Fungal Chitinase for the Bioconversion of Mycelial Waste. J. Agric. Food Chem. 2020, 68, 5384–5390. [Google Scholar] [CrossRef]
- Yang, J.; Gan, Z.; Lou, Z.; Tao, N.; Mi, Q.; Liang, L.; Sun, Y.; Guo, Y.; Huang, X.; Zou, C.; et al. Crystal Structure and Mutagenesis Analysis of Chitinase CrChi1 from the Nematophagous Fungus Clonostachys Rosea in Complex with the Inhibitor Caffeine. Microbiology (N. Y.) 2010, 156, 3566–3574. [Google Scholar] [CrossRef]
- Rao, F.V.; Andersen, O.A.; Vora, K.A.; DeMartino, J.A.; van Aalten, D.M.F. Methylxanthine Drugs Are Chitinase Inhibitors: Investigation of Inhibition and Binding Modes. Chem. Biol. 2005, 12, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; de Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB Bioinformatics Resource Portal. Nucleic Acids Res. 2012, 40, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Engeset, R.V.; Udnæs, H.C.; Guneriussen, T.; Koren, H.; Malnes, E.; Solberg, R.; Alfnes, E. Improving Runoff Simulations Using Satellite-Observed Time-Series of Snow Covered Area. Nord. Hydrol. 2003, 34, 281–294. [Google Scholar] [CrossRef]
- Bo, Y.Y.; Liang, L.D.; Hua, Y.J.; Zhao, Z.; Yao, M.S.; Shan, L.B.; Liang, C.Z. Benchmark High-Purity DNA Extraction from Animal Tissue Using Picking in the TRIzol-Based Method. Biotechniques 2020, 70, 187–191. [Google Scholar]
- Yuli, P.E.; Suhartono, M.T.; Rukayadi, Y.; Hwang, J.K.; Pyun, Y.R. Characteristics of Thermostable Chitinase Enzymes from the Indonesian Bacillus Sp.13.26. Enzym. Microb. Technol. 2004, 35, 147–153. [Google Scholar] [CrossRef]
- Kumar, M.; Madhuprakash, J.; Balan, V.; Kumar, A.; Vivekanand, V.; Pareek, N. Bioresource Technology Chemoenzymatic Production of Chitooligosaccharides Employing Ionic Liquids and Thermomyces Lanuginosus Chitinase. Bioresour. Technol. 2021, 337, 125399. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Step | Total Activity (mU) | Total Protein (mg) | Specific Activity (mU/mg) | Purification Factor (-Fold) | Recovery Yield (%) |
|---|---|---|---|---|---|
| Crude extract | 577.248 | 783.24 | 0.737 | 1.0 | 100 |
| purified protein eluate | 561.878 | 10.72 | 52.414 | 71.12 | 97.34 |
| Gel filtration chromatography | 156.837 | 0.242 | 648.088 | 879.36 | 27.17 |
| Additives | Relative Activity (%) | |
|---|---|---|
| 0.01 M | 0.05 M | |
| Control | 100.00 | 100.00 |
| Cobaltous (Co2+) | 75.40 | 131.35 |
| Sodium (Na+) | 181.79 | 173.34 |
| Magnesium (Mg2+) | 53.50 | 145.73 |
| Copper (Cu2+) | 24.53 | 62.19 |
| Iron (Fe3+) | 15.30 | 62.61 |
| Ammonium(NH4+) | 83.51 | 122.90 |
| Calcium (Ca2+) | 92.29 | 160.70 |
| Zinc (Zn2+) | 34.44 | 134.76 |
| Manganese(Mn2+) | 47.80 | 94.28 |
| Potassium (K+ ) | 111.03 | 147.61 |
| Barium(Ba2+) | 40.18 | 73.21 |
| Tris | 105.75 | 155.56 |
| SDS | 0 | 0 |
| EDTA | 52.56 | 92.34 |
| Carbamide | 106.49 | 120.77 |
| Substrate | Specific Activity (mU/mg) a |
|---|---|
| Colloidal chitin | 0.7134 |
| Chitin powder | 0.0137 |
| CMC | None |
| Chitosan powder | None |
| Ball milled crab shell powder | 0.0237 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.-L.; Wang, S.; Yang, D.-F.; Yang, L.-Y.; Wang, Q.-Y.; Yu, J.; Li, N.; Pan, L.-X. The Discovery, Enzymatic Characterization and Functional Analysis of a Newly Isolated Chitinase from Marine-Derived Fungus Aspergillus fumigatus df347. Mar. Drugs 2022, 20, 520. https://doi.org/10.3390/md20080520
Wu Y-L, Wang S, Yang D-F, Yang L-Y, Wang Q-Y, Yu J, Li N, Pan L-X. The Discovery, Enzymatic Characterization and Functional Analysis of a Newly Isolated Chitinase from Marine-Derived Fungus Aspergillus fumigatus df347. Marine Drugs. 2022; 20(8):520. https://doi.org/10.3390/md20080520
Chicago/Turabian StyleWu, Ya-Li, Sheng Wang, Deng-Feng Yang, Li-Yan Yang, Qing-Yan Wang, Jun Yu, Nan Li, and Li-Xia Pan. 2022. "The Discovery, Enzymatic Characterization and Functional Analysis of a Newly Isolated Chitinase from Marine-Derived Fungus Aspergillus fumigatus df347" Marine Drugs 20, no. 8: 520. https://doi.org/10.3390/md20080520