
Epoxinnamide: An Epoxy Cinnamoyl-Containing Nonribosomal Peptide from an Intertidal Mudflat-Derived Streptomyces sp.

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
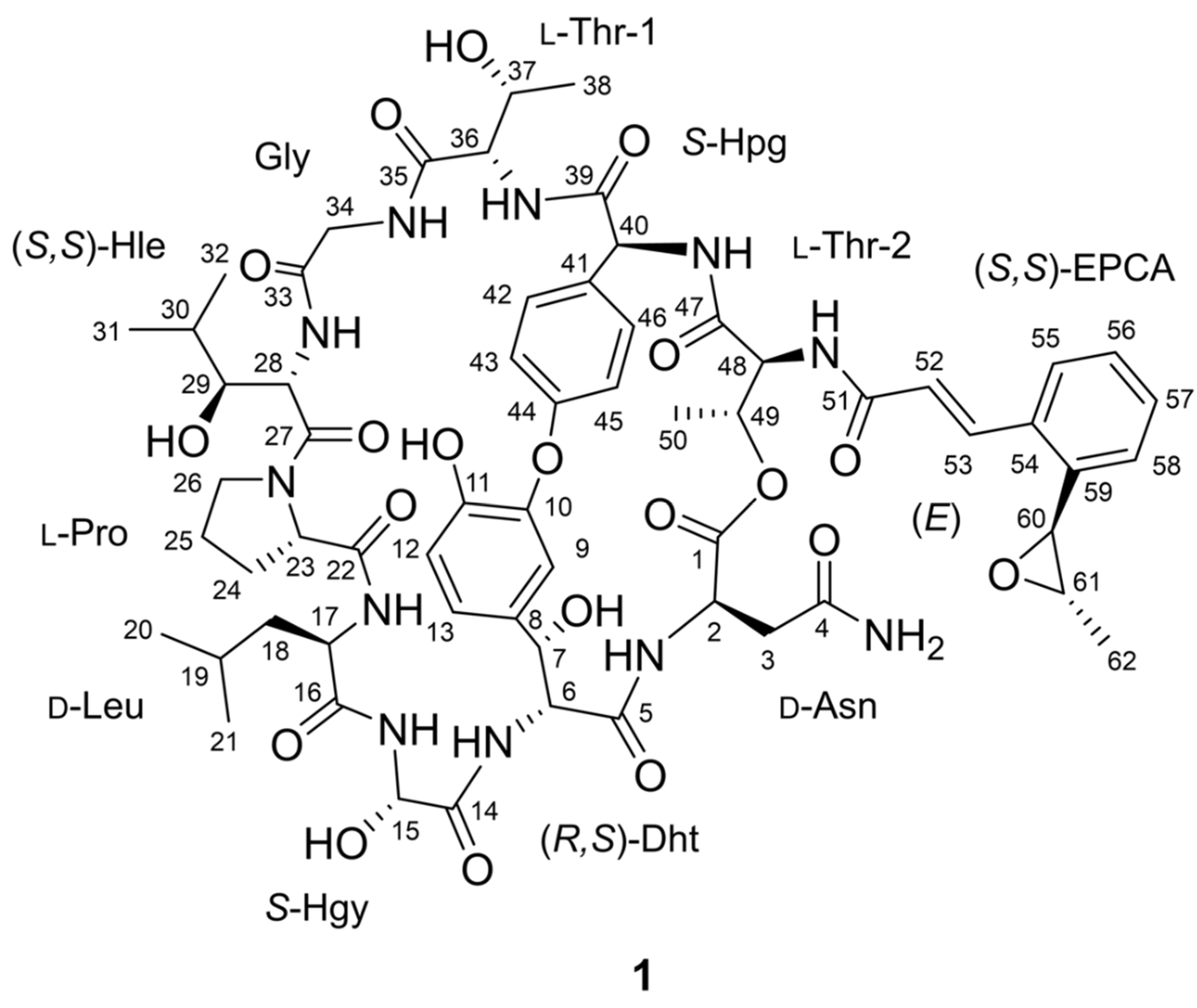
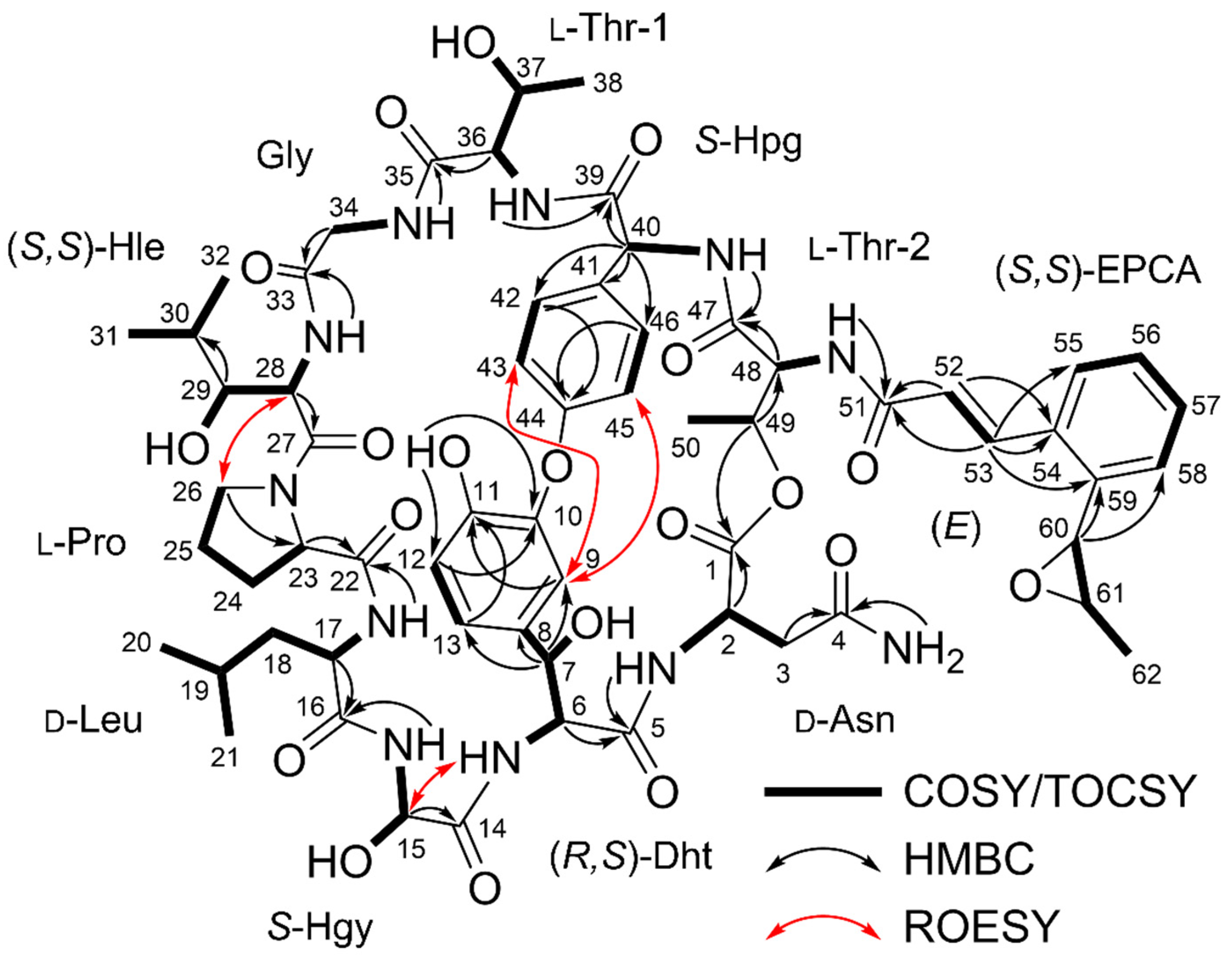
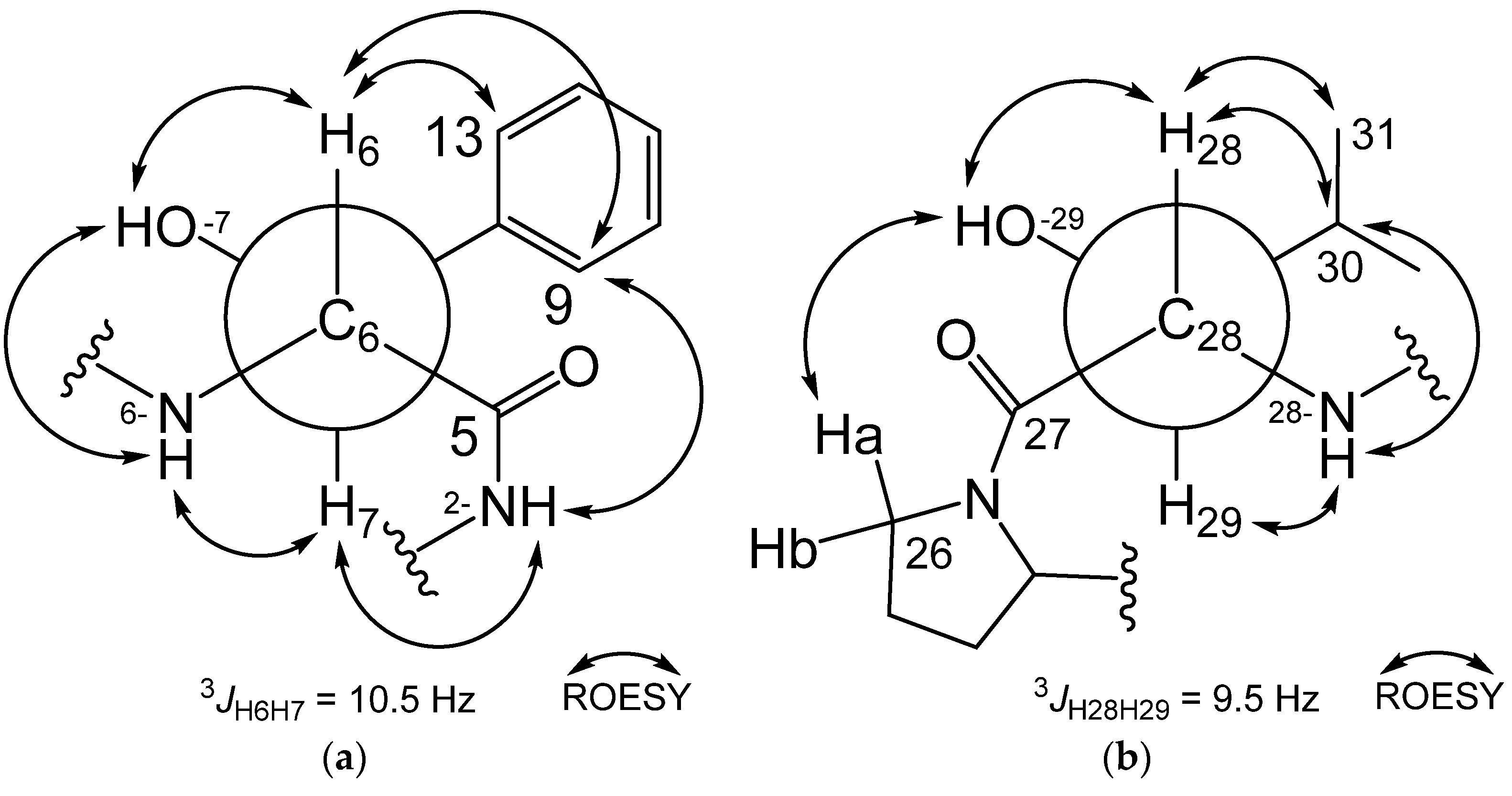
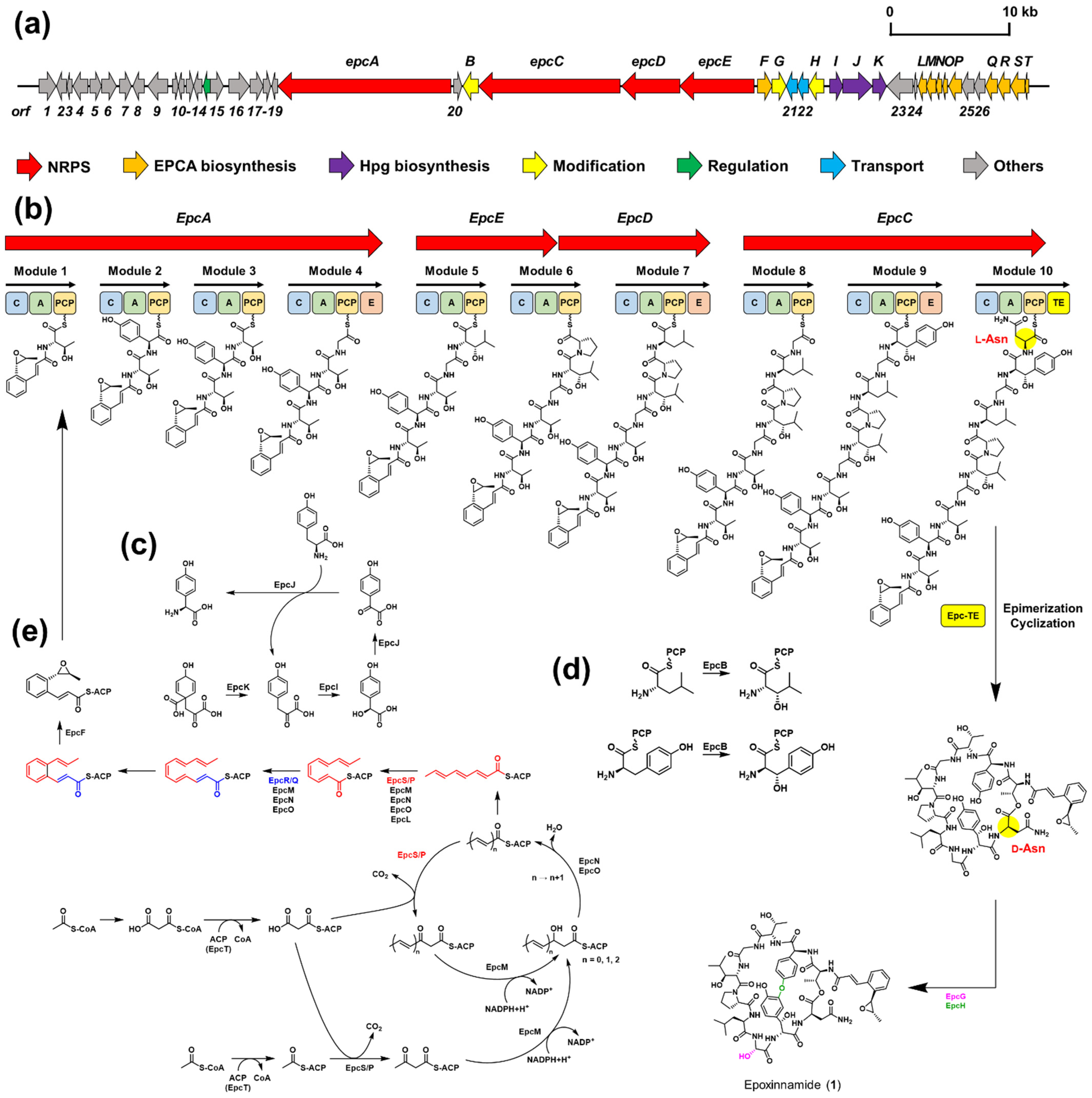
2.1. Structure Elucidation and Putative Biosynthetic Pathway of Epoxinnamide (1)
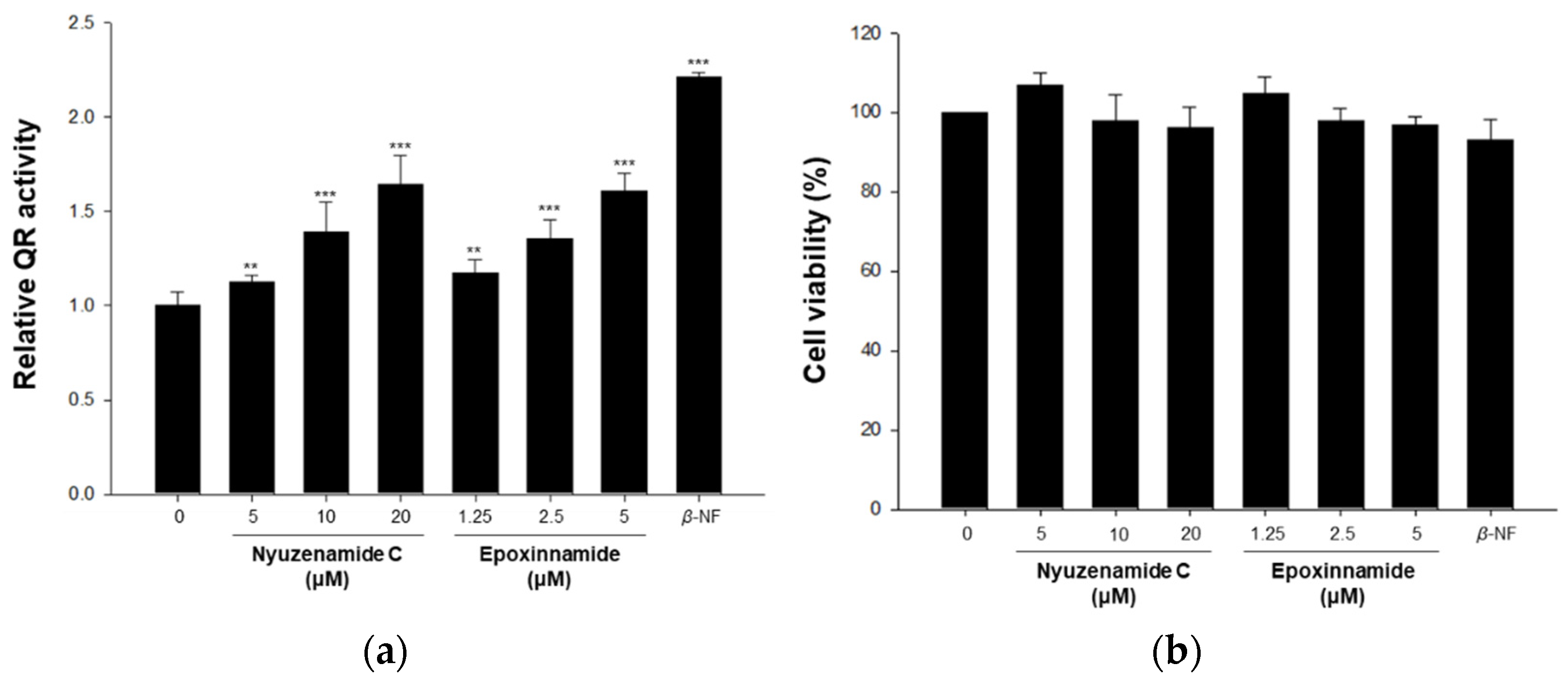
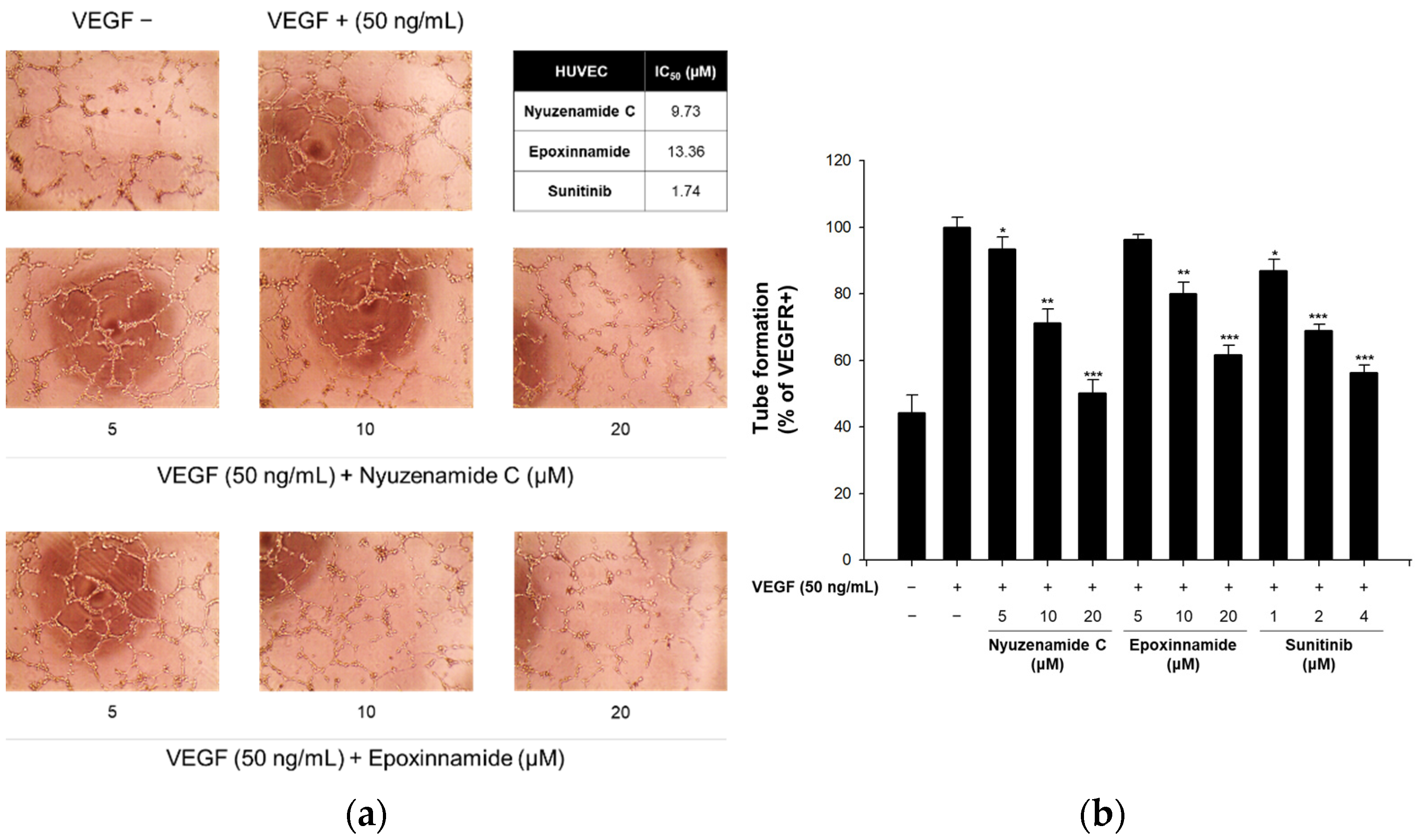
2.2. Bioactivity of Epoxinnamide (1)
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Bacterial Isolation
3.3. Phylogenetic Analysis of Streptomyces sp. OID44
3.4. Large-Scale Culture and Extraction
3.5. Purification of Epoxinnamide (1)
3.6. Determination of the Absolute Configuration at the α- and β-Carbons of the Amino Acid Units
3.7. Conformation Search and DP4 Analysis
3.8. Genome Analysis of Streptomyces sp. OID44
3.9. Quinone Reductase Assay
3.10. In Vitro Capillary Tube Formation Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fischbach, M.A.; Walsh, C.T. Assembly-Line Enzymology for Polyketide and Nonribosomal Peptide Antibiotics: Logic, Machinery, and Mechanisms. Chem. Rev. 2006, 106, 3468–3496. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Liu, C.L.; Zhang, B.; Guo, W.J.; Zhu, J.; Chang, C.-Y.; Zhao, E.J.; Jiao, R.H.; Tan, R.X.; Ge, H.M. Genome Mining and Biosynthesis of Kitacinnamycins as a STING Activator. Chem. Sci. 2019, 10, 4839–4846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, K.; Hashimoto, M.; Shigematsu, N.; Nishikawa, M.; Ezaki, M.; Yamashita, M.; Kiyoto, S.; Okuhara, M.; Kohsaka, M.; Imanaka, H. WS9326A, a Novel Tachykinin Antagonist Isolated from Streptomyces violaceusniger No. 9326. I. Taxonomy, Fermentation, Isolation, Physico-chemical Properties and Biological Activities. J. Antibiot. 1992, 45, 1055–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Zhu, J.; Zechel, D.L.; Jessen-Trefzer, C.; Eastman, R.T.; Paululat, T.; Bechthold, A. New WS9326A Derivatives and One New Annimycin Derivative with Antimalarial Activity are Produced by Streptomyces asterosporus DSM 41452 and Its Mutant. Chembiochem 2018, 19, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Vodanovic-Jankovic, S.; Kron, M.; Shen, B. New WS9326A Congeners from Streptomyces sp. 9078 Inhibiting Brugia malayi Asparaginyl-tRNA Synthetase. Org. Lett. 2012, 14, 4946–4949. [Google Scholar] [CrossRef] [Green Version]
- Shiomi, K.; Yang, H.; Inokoshi, J.; Van der Pyl, D.; Nakagawa, A.; Takeshima, H.; Omura, S. Pepticinnamins, New Farnesyl-protein Transferase Inhibitors Produced by an Actinomycete. II. Structural Elucidation of Pepticinnamin E. J. Antibiot. 1993, 46, 229–234. [Google Scholar] [CrossRef] [Green Version]
- Toki, S.; Agatsuma, T.; Ochiai, K.; Saitoh, Y.; Ando, K.; Nakanishi, S.; Lokker, N.A.; Giese, N.A.; Matsuda, Y. RP-1776, a Novel Cyclic Peptide Produced by Streptomyces sp., inhibits the Binding of PDGF to the Extracellular Domain of Its Receptor. J. Antibiot. 2001, 54, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Pohle, S.; Appelt, C.; Roux, M.; Fiedler, H.-P.; Süssmuth, R.D. Biosynthetic Gene Cluster of the Non-Ribosomally Synthesized Cyclodepsipeptide Skyllamycin: Deciphering Unprecedented Ways of Unusual Hydroxylation Reactions. J. Am. Chem. Soc. 2011, 133, 6194–6205. [Google Scholar] [CrossRef]
- Navarro, G.; Cheng, A.T.; Peach, K.C.; Bray, W.M.; Bernan, V.S.; Yildiz, F.H.; Linington, R.G. Image-Based 384-Well High-Throughput Screening Method for the Discovery of Skyllamycins A to C as Biofilm Inhibitors and Inducers of Biofilm Detachment in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2014, 58, 1092–1099. [Google Scholar] [CrossRef] [Green Version]
- Bracegirdle, J.; Hou, P.; Nowak, V.V.; Ackerley, D.F.; Keyzers, R.A.; Owen, J.G. Skyllamycins D and E, Non-Ribosomal Cyclic Depsipeptides from Lichen-Sourced Streptomyces anulatus. J. Nat. Prod. 2021, 84, 2536–2543. [Google Scholar] [CrossRef]
- Zhang, F.; Adnani, N.; Vazquez-Rivera, E.; Braun, D.R.; Tonelli, M.; Andes, D.R.; Bugni, T.S. Application of 3D NMR for Structure Determination of Peptide Natural Products. J. Org. Chem. 2015, 80, 8713–8719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Liu, N.; Shang, F.; Huang, Y. Activation and Identification of NC-1: A Cryptic Cyclodepsipeptide from Red Soil-Derived Streptomyces sp. FXJ1.172. Eur. J. Org. Chem. 2016, 2016, 3943–3948. [Google Scholar] [CrossRef]
- Sun, C.; Yang, Z.; Zhang, C.; Liu, Z.; He, J.; Liu, Q.; Zhang, T.; Ju, J.; Ma, J. Genome Mining of Streptomyces atratus SCSIO ZH16: Discovery of Atratumycin and Identification of Its Biosynthetic Gene Cluster. Org. Lett. 2019, 21, 1453–1457. [Google Scholar] [CrossRef]
- Liu, Q.; Liu, Z.; Sun, C.; Shao, M.; Ma, J.; Wei, X.; Zhang, T.; Li, W.; Ju, J. Discovery and Biosynthesis of Atrovimycin, an Antitubercular and Antifungal Cyclodepsipeptide Featuring Vicinal-Dihydroxylated Cinnamic Acyl Chain. Org. Lett. 2019, 21, 2634–2638. [Google Scholar] [CrossRef]
- Zhang, C.; Seyedsayamdost, M.R. Discovery of a Cryptic Depsipeptide from Streptomyces ghanaensis via MALDI-MS-Guided High-Throughput Elicitor Screening. Angew. Chem. Int. Ed. 2020, 59, 23005–23009. [Google Scholar] [CrossRef]
- Karim, R.U.; In, Y.; Zhou, T.; Harunari, E.; Oku, N.; Igarashi, Y. Nyuzenamides A and B: Bicyclic Peptides with Antifungal and Cytotoxic Activity from a Marine-Derived Streptomyces sp. Org. Lett. 2021, 23, 2109–2113. [Google Scholar] [CrossRef] [PubMed]
- Bae, M.; Kim, H.; Moon, K.; Nam, S.-J.; Shin, J.; Oh, K.-B.; Oh, D.-C. Mohangamides A and B, New Dilactone-Tethered Pseudo-Dimeric Peptides Inhibiting Candida Albicans Isocitrate Lyase. Org. Lett. 2015, 17, 712–715. [Google Scholar] [CrossRef] [PubMed]
- Bae, M.; Oh, J.; Bae, E.S.; Oh, J.; Hur, J.; Suh, Y.-G.; Lee, S.K.; Shin, J.; Oh, D.-C. WS9326H, an Antiangiogenic Pyrazolone-Bearing Peptide from an Intertidal Mudflat Actinomycete. Org. Lett. 2018, 20, 1999–2002. [Google Scholar] [CrossRef]
- Um, S.; Park, S.H.; Kim, J.; Park, H.J.; Ko, K.; Bang, H.-S.; Lee, S.K.; Shin, J.; Oh, D.-C. Coprisamides A and B, New Branched Cyclic Peptides from a Gut Bacterium of the Dung Beetle Copris tripartitus. Org. Lett. 2015, 17, 1272–1275. [Google Scholar] [CrossRef]
- Shin, Y.-H.; Ban, Y.H.; Kim, T.H.; Bae, E.S.; Shin, J.; Lee, S.K.; Jang, J.; Yoon, Y.J.; Oh, D.-C. Structures and Biosynthetic Pathway of Coprisamides C and D, 2-Alkenylcinnamic Acid-Containing Peptides from the Gut Bacterium of the Carrion Beetle Silpha perforata. J. Nat. Prod. 2021, 84, 239–246. [Google Scholar] [CrossRef]
- An, J.S.; Kim, M.-S.; Han, J.; Jang, S.C.; Im, J.H.; Cui, J.; Lee, Y.; Nam, S.-J.; Shin, J.; Lee, S.K.; et al. Nyuzenamide C, an Antiangiogenic Epoxy Cinnamic Acid-Containing Bicyclic Peptide from a Riverine Streptomyces sp. J. Nat. Prod. 2022, 85, 804–814. [Google Scholar] [CrossRef] [PubMed]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine Natural Products. Nat. Prod. Rep. 2022, 39, 1122–1171. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.; Fujii, K.; Mayumi, T.; Hibino, Y.; Suzuki, M.; Ikai, Y.; Oka, H. A Method Using LC/MS for Determination of Absolute Configuration of Constituent Amino Acids in Peptide—Advanced Marfey’s Method. Tetrahedron Lett. 1995, 36, 1515–1518. [Google Scholar] [CrossRef]
- MacMillan, J.B.; Ernst-Russell, M.A.; de Ropp, J.S.; Molinski, T.F. Lobocyclamides A–C, Lipopeptides from a Cryptic Cyanobacterial Mat Containing Lyngbya confervoides. J. Org. Chem. 2002, 67, 8210–8215. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.G.; Urry, D.W. Proton Magnetic Resonance Assignments of the Polypeptide Antibiotic Telomycin. Biochemistry 1973, 12, 3811–3817. [Google Scholar] [CrossRef]
- Wyche, T.P.; Ruzzini, A.C.; Beemelmanns, C.; Kim, K.H.; Klassen, J.L.; Cao, S.; Poulsen, M.; Bugni, T.S.; Currie, C.R.; Clardy, J. Linear Peptides Are the Major Products of a Biosynthetic Pathway That Encodes for Cyclic Depsipeptides. Org. Lett. 2017, 19, 1772–1775. [Google Scholar] [CrossRef]
- Raju, R.; Khalil, Z.G.; Piggott, A.M.; Blumenthal, A.; Gardiner, D.L.; Skinner-Adams, T.S.; Capon, R.J. Mollemycin A: An Antimalarial and Antibacterial Glyco-Hexadepsipeptide-Polyketide from an Australian Marine-Derived Streptomyces sp. (CMB-M0244). Org. Lett. 2014, 16, 1716–1719. [Google Scholar] [CrossRef]
- McDonald, L.A.; Barbieri, L.R.; Carter, G.T.; Lenoy, E.; Lotvin, J.; Petersen, P.J.; Siegel, M.M.; Singh, G.; Williamson, R.T. Structures of the Muraymycins, Novel Peptidoglycan Biosynthesis Inhibitors. J. Am. Chem. Soc. 2002, 124, 10260–10261. [Google Scholar] [CrossRef]
- Shimokawa, K.; Mashima, I.; Asai, A.; Ohno, T.; Yamada, K.; Kita, M.; Uemura, D. Biological Activity, Structural Features, and Synthetic Studies of (−)-Ternatin, a Potent Fat-Accumulation Inhibitor of 3T3-L1 Adipocytes. Chem.—Asian J. 2008, 3, 438–446. [Google Scholar] [CrossRef]
- Nam, S.-J.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Isolation and Characterization of Actinoramides A–C, Highly Modified Peptides from a Marine Streptomyces sp. Tetrahedron 2011, 67, 6707–6712. [Google Scholar] [CrossRef] [Green Version]
- Machida, K.; Arai, D.; Katsumata, R.; Otsuka, S.; Yamashita, J.K.; Ye, T.; Tang, S.; Fusetani, N.; Nakao, Y. Sameuramide A, a New Cyclic Depsipeptide Isolated from an Ascidian of the Family Didemnidae. Bioorganic Med. Chem. 2018, 26, 3852–3857. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, M.; Suzumura, K.-I.; Nagai, K.; Kawasaki, T.; Takasaki, J.; Sekiguchi, M.; Moritani, Y.; Saito, T.; Hayashi, K.; Fujita, S.; et al. YM-254890 Analogues, Novel Cyclic Depsipeptides with Gαq/11 Inhibitory Activity from Chromobacterium sp. QS3666. Bioorganic Med. Chem. 2004, 12, 3125–3133. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Kemmoku, K.; Satoh, Y.; Ogasawara, Y.; Shin-ya, K.; Dairi, T. N-Phenylacetylation and Nonribosomal Peptide Synthetases with Substrate Promiscuity for Biosynthesis of Heptapeptide Variants, JBIR-78 and JBIR-95. ACS Chem. Biol. 2017, 12, 1813–1819. [Google Scholar] [CrossRef] [PubMed]
- Giltrap, A.M.; Haeckl, F.P.J.; Kurita, K.L.; Linington, R.G.; Payne, R.J. Synthetic Studies Toward the Skyllamycins: Total Synthesis and Generation of Simplified Analogues. J. Org. Chem. 2018, 83, 7250–7270. [Google Scholar] [CrossRef] [PubMed]
- Boyaud, F.; Mahiout, Z.; Lenoir, C.; Tang, S.; Wdzieczak-Bakala, J.; Witczak, A.; Bonnard, I.; Banaigs, B.; Ye, T.; Inguimbert, N. First Total Synthesis and Stereochemical Revision of Laxaphycin B and Its Extension to Lyngbyacyclamide A. Org. Lett. 2013, 15, 3898–3901. [Google Scholar] [CrossRef] [PubMed]
- Bae, M.; Mevers, E.; Pishchany, G.; Whaley, S.G.; Rock, C.O.; Andes, D.R.; Currie, C.R.; Pupo, M.T.; Clardy, J. Chemical Exchanges between Multilateral Symbionts. Org. Lett. 2021, 23, 1648–1652. [Google Scholar] [CrossRef]
- Treadwell, E.M.; Yan, Z.; Xiao, X. Epoxidation with Possibilities: Discovering Stereochemistry in Organic Chemistry via Coupling Constants. J. Chem. Educ. 2017, 94, 640–643. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. AntiSMASH 6.0: Improving Cluster Detection and Comparison Capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 Web Portal for Protein Modeling, Prediction and Analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Song, J.; Chi, C.; Liu, T.; Geng, T.; Cai, Z.; Dong, W.; Shi, C.; Ma, X.; Zhang, Z.; et al. Functional Characterization and Crystal Structure of the Bifunctional Thioesterase Catalyzing Epimerization and Cyclization in Skyllamycin Biosynthesis. ACS Catal. 2021, 11, 11733–11741. [Google Scholar] [CrossRef]
- Al Toma, R.S.; Brieke, C.; Cryle, M.J.; Süssmuth, R.D. Structural Aspects of Phenylglycines, Their Biosynthesis and Occurrence in Peptide Natural Products. Nat. Prod. Rep. 2015, 32, 1207–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubbard, B.K.; Thomas, M.G.; Walsh, C.T. Biosynthesis of L-p-Hydroxyphenylglycine, a Non-Proteinogenic Amino Acid Constituent of Peptide Antibiotics. Chem. Biol. 2000, 7, 931–942. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Zabriskie, T.M. The Enduracidin Biosynthetic Gene Cluster from Streptomyces fungicidicus. Microbiology 2006, 152, 2969–2983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hojati, Z.; Milne, C.; Harvey, B.; Gordon, L.; Borg, M.; Flett, F.; Wilkinson, B.; Sidebottom, P.J.; Rudd, B.A.; A Hayes, M.; et al. Structure, Biosynthetic Origin, and Engineered Biosynthesis of Calcium-Dependent Antibiotics from Streptomyces coelicolor. Chem. Biol. 2002, 9, 1175–1187. [Google Scholar] [CrossRef]
- Uhlmann, S.; Süssmuth, R.D.; Cryle, M.J. Cytochrome P450sky Interacts Directly with the Nonribosomal Peptide Synthetase to Generate Three Amino Acid Precursors in Skyllamycin Biosynthesis. ACS Chem. Biol. 2013, 8, 2586–2596. [Google Scholar] [CrossRef]
- Fu, C.; Keller, L.; Bauer, A.; Brönstrup, M.; Froidbise, A.; Hammann, P.; Herrmann, J.; Mondesert, G.; Kurz, M.; Schiell, M.; et al. Biosynthetic Studies of Telomycin Reveal New Lipopeptides with Enhanced Activity. J. Am. Chem. Soc. 2015, 137, 7692–7705. [Google Scholar] [CrossRef]
- Deng, Z.; Liu, J.; Li, T.; Li, H.; Liu, Z.; Dong, Y.; Li, W. An Unusual Type II Polyketide Synthase System Involved in Cinnamoyl Lipid Biosynthesis. Angew. Chem. Int. Ed. 2021, 60, 153–158. [Google Scholar] [CrossRef]
- Shi, J.; Shi, Y.; Li, J.C.; Wei, W.; Chen, Y.; Cheng, P.; Liu, C.L.; Zhang, H.; Wu, R.; Zhang, B.; et al. In Vitro Reconstitution of Cinnamoyl Moiety Reveals Two Distinct Cyclases for Benzene Ring Formation. J. Am. Chem. Soc. 2022, 144, 7939–7948. [Google Scholar] [CrossRef]
- Hertweck, C. The Biosynthetic Logic of Polyketide Diversity. Angew. Chem. Int. Ed. 2009, 48, 4688–4716. [Google Scholar] [CrossRef]
- Arthur, C.J.; Szafranska, A.; Evans, S.E.; Findlow, S.C.; Burston, S.G.; Owen, P.; Clark-Lewis, I.; Simpson, T.J.; Crosby, J.; Crump, M.P. Self-Malonylation Is an Intrinsic Property of a Chemically Synthesized Type II Polyketide Synthase Acyl Carrier Protein. Biochemistry 2005, 44, 15414–15421. [Google Scholar] [CrossRef]
- Gómez, C.; Olano, C.; Palomino-Schätzlein, M.; Pineda-Lucena, A.; Carbajo, R.J.; Braña, A.F.; Méndez, C.; Salas, J.A. Novel compounds produced by Streptomyces lydicus NRRL 2433 engineered mutants altered in the biosynthesis of streptolydigin. J. Antibiot. 2012, 65, 341–348. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Gu, K.; Xia, S.-S.; Zhang, D.-J.; Li, Y.-G. Dolyemycins A and B, two novel cyclopeptides isolated from Streptomyces griseus subsp. griseus HYS31. J. Antibiot. 2018, 71, 838–845. [Google Scholar] [CrossRef] [PubMed]
- Peschke, M.; Brieke, C.; Cryle, M.J. F-O-G Ring Formation in Glycopeptide Antibiotic Biosynthesis is Catalysed by OxyE. Sci. Rep. 2016, 6, 35584. [Google Scholar] [CrossRef] [PubMed]
- Haslinger, K.; Maximowitsch, E.; Brieke, C.; Koch, A.; Cryle, M.J. Cytochrome P450 OxyBtei Catalyzes the First Phenolic Coupling Step in Teicoplanin Biosynthesis. ChemBioChem 2014, 15, 2719–2728. [Google Scholar] [CrossRef] [PubMed]
- Carlson, J.C.; Fortman, J.L.; Anzai, Y.; Li, S.; Burr, D.A.; Sherman, D.H. Identification of the Tirandamycin Biosynthetic Gene Cluster from Streptomyces sp. 307-9. ChemBioChem 2010, 11, 564–572. [Google Scholar] [CrossRef] [Green Version]
- Cuendet, M.; Oteham, C.P.; Moon, R.C.; Pezzuto, J.M. Quinone Reductase Induction as a Biomarker for Cancer Chemoprevention. J. Nat. Prod. 2006, 69, 460–463. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Choi, S.W.; Cho, J.; Been, J.-H.; Choi, K.; Jiang, W.; Han, J.; Oh, J.; Park, C.; Choi, S.; et al. Discovery of Novel Small-Molecule Antiangiogenesis Agents to Treat Diabetic Retinopathy. J. Med. Chem. 2021, 64, 5535–5550. [Google Scholar] [CrossRef]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A Taxonomically United Database of 16S rRNA Gene Sequences and Whole-Genome Assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Pruesse, E.; Peplies, J.; Glöckner, F.O. SINA: Accurate High-Throughput Multiple Sequence Alignment of Ribosomal RNA Genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Halgren, T.A. Merck Molecular Force Field. I. Basis, Form, Scope, Parameterization, and Performance of MMFF94. J. Comput. Chem. 1996, 17, 490–519. [Google Scholar] [CrossRef]
- Smith, S.G.; Goodman, J.M. Assigning Stereochemistry to Single Diastereoisomers by GIAO NMR Calculation: The DP4 Probability. J. Am. Chem. Soc. 2010, 132, 12946–12959. [Google Scholar] [CrossRef]
- Steffen, C.; Thomas, K.; Huniar, U.; Hellweg, A.; Rubner, O.; Schroer, A. TmoleX-A Graphical User Interface for TURBOMOLE. J. Comput. Chem. 2010, 31, 2967–2970. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, A.; Au, K.F. PacBio Sequencing and Its Applications. Genom. Proteom. Bioinform. 2015, 13, 278–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, C.-S.; Alexander, D.H.; Marks, P.; Klammer, A.A.; Drake, J.; Heiner, C.; Clum, A.; Copeland, A.; Huddleston, J.; Eichler, E.E.; et al. Nonhybrid, Finished Microbial Genome Assemblies from Long-Read SMRT Sequencing Data. Nat. Methods. 2013, 10, 563–569. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Meng, E.C.; Couch, G.S.; Croll, T.I.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Structure Visualization for Researchers, Educators, and Developers. Protein Sci. 2021, 30, 70–82. [Google Scholar] [CrossRef] [PubMed]
- An, J.S.; Lee, J.Y.; Kim, E.; Ahn, H.; Jang, Y.-J.; Shin, B.; Hwang, S.; Shin, J.; Yoon, Y.J.; Lee, S.K.; et al. Formicolides A and B, Antioxidative and Antiangiogenic 20-Membered Macrolides from a Wood Ant Gut Bacterium. J. Nat. Prod. 2020, 83, 2776–2784. [Google Scholar] [CrossRef]
- Wang, C.Y.; Kim, D.; Zhu, Y.K.; Oh, D.-C.; Huang, R.Z.; Wang, H.-S.; Liang, D.; Lee, S.K. Glechomanamides A–C, Germacrane Sesquiterpenoids with an Unusual Δ8-7,12-Lactam Moiety from Salvia scapiformis and Their Antiangiogenic Activity. J. Nat. Prod. 2019, 82, 3056–3064. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δC, Type | δH (J in Hz) | Position | δC, Type | δH (J in Hz) |
|---|---|---|---|---|---|
| d-Asn | 28-NH | 7.69 a, d (9.5) | |||
| 1 | 169.6, C | 29 | 74.4, CH | 3.22, ddd (9.5, 2.0, 2.0) | |
| 2 | 49.0, CH | 3.99, ddd (9.5, 7.0, 3.5) | 29-OH | 5.09, br d (2.0) | |
| 2-NH | 8.23, d (7.0) | 30 | 27.9, CH | 1.74, m | |
| 3a | 35.1, CH2 | 2.65, dd (15.5, 3.5) | 31 | 14.4, CH3 | 0.72, d (7.0) |
| 3b | 2.57, dd (15.5, 9.5) | 32 | 20.2, CH3 | 0.74, d (7.0) | |
| 4 | 170.64, C | Gly | |||
| 4-NH2a | 7.43 a, br s | 33 | 168.4, C | ||
| 4-NH2b | 7.03, br s | 34a | 42.60, CH2 | 4.18, dd (17.5, 8.5) | |
| (R,S)-Dht | 34b | 3.39, dd (17.5, 5.0) | |||
| 5 | 170.63, C | 34-NH | 7.68 a, dd (8.5, 5.0) | ||
| 6 | 63.8, CH | 3.49, dd (10.5, 3.5) | l-Thr-1 | ||
| 6-NH | 8.14 a, br s | 35 | 170.0, C | ||
| 7 | 69.5, CH | 4.69, dd (10.5, 6.5) | 36 | 57.7, CH | 4.44, dd (10.0, 3.5) |
| 7-OH | 6.15, d (6.5) | 36-NH | 8.16, d (10.0) | ||
| 8 | 132.3, C | 37 | 65.8, CH | 4.40, m | |
| 9 | 120.1, CH | 6.27, d (2.0) | 37-OH | 4.52, d (7.0) | |
| 10 | 147.3, C | 38 | 20.6, CH3 | 1.08, d (6.5) | |
| 11 | 147.4, C | S-Hpg | |||
| 11-OH | 9.40, s | 39 | 169.1, C | ||
| 12 | 116.1, CH | 6.87, d (8.5) | 40 | 60.1, CH | 4.83, d (3.0) |
| 13 | 120.2, CH | 7.08 a, dd (8.5, 2.0) | 40-NH | 7.55, d (3.0) | |
| S-Hgy | 41 | 131.6, C | |||
| 14 | 170.9, C | 42 | 128.5, CH | 7.07 a, dd (8.5, 2.0) | |
| 15 | 71.3, CH | 5.79, dd (9.5, 5.5) | 43 | 123.6, CH | 6.42, dd (8.5, 2.5) |
| 15-OH | 7.17 a, br s | 44 | 159.6, C | ||
| 15-NH | 9.07, d (9.5) | 45 | 122.3, CH | 7.18, dd (8.0, 2.5) | |
| d-Leu | 46 | 131.3, CH | 7.37, dd (8.0, 2.0) | ||
| 16 | 171.5, C | l-Thr-2 | |||
| 17 | 50.3, CH | 4.64, td (9.5, 5.0) | 47 | 173.3, C | |
| 17-NH | 8.44, d (9.5) | 48 | 61.0, CH | 5.03, d (4.5) | |
| 18a | 42.58, CH2 | 1.45 a, m | 48-NH | 8.96, d (4.5) | |
| 18b | 1.31, m | 49 | 68.7, CH | 5.19, q (7.0) | |
| 19 | 24.1, CH | 1.46 a, m | 50 | 17.3, CH3 | 1.23, d (7.0) |
| 20 | 23.2, CH3 | 0.86, d (6.5) | (S,S)-EPCA | ||
| 21 | 21.8, CH3 | 0.85, d (6.5) | 51 | 169.2, C | |
| l-Pro | 52 | 124.6, CH | 6.94, d (15.5) | ||
| 22 | 171.8, C | 53 | 136.69, CH | 8.11, d (15.5) | |
| 23 | 60.2, CH | 4.37, dd (7.5, 7.0) | 54 | 134.1, C | |
| 24a | 30.0, CH2 | 2.21, m | 55 | 126.9, CH | 7.42 a, d (7.5) |
| 24b | 1.56, m | 56 | 127.8, CH | 7.00, ddd (7.5, 7.5, 1.0) | |
| 25 | 24.7, CH2 | 1.79, m | 57 | 129.6, CH | 7.33, ddd (7.5, 7.5, 1.0) |
| 26a | 47.3, CH2 | 3.72, m | 58 | 124.4, CH | 7.09 a, dd (7.5, 1.0) |
| 26b | 3.52, m | 59 | 136.74, C | ||
| (S,S)-Hle | 60 | 54.8, CH | 4.07, d (2.0) | ||
| 27 | 170.1, C | 61 | 58.9, CH | 2.97, qd (5.0, 2.0) | |
| 28 | 52.5, CH | 4.59, dd (9.5, 9.5) | 62 | 17.6, CH3 | 1.37, d (5.0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, S.; Han, J.; Jang, S.C.; An, J.S.; Kang, I.; Kwon, Y.; Nam, S.-J.; Shim, S.H.; Cho, J.-C.; Lee, S.K.; et al. Epoxinnamide: An Epoxy Cinnamoyl-Containing Nonribosomal Peptide from an Intertidal Mudflat-Derived Streptomyces sp. Mar. Drugs 2022, 20, 455. https://doi.org/10.3390/md20070455
Kang S, Han J, Jang SC, An JS, Kang I, Kwon Y, Nam S-J, Shim SH, Cho J-C, Lee SK, et al. Epoxinnamide: An Epoxy Cinnamoyl-Containing Nonribosomal Peptide from an Intertidal Mudflat-Derived Streptomyces sp. Marine Drugs. 2022; 20(7):455. https://doi.org/10.3390/md20070455
Chicago/Turabian StyleKang, Sangwook, Jaeho Han, Sung Chul Jang, Joon Soo An, Ilnam Kang, Yun Kwon, Sang-Jip Nam, Sang Hee Shim, Jang-Cheon Cho, Sang Kook Lee, and et al. 2022. "Epoxinnamide: An Epoxy Cinnamoyl-Containing Nonribosomal Peptide from an Intertidal Mudflat-Derived Streptomyces sp." Marine Drugs 20, no. 7: 455. https://doi.org/10.3390/md20070455