
Alzheimer’s Disease and Toxins Produced by Marine Dinoflagellates: An Issue to Explore



Abstract
:1. Introduction
2. Alzheimer’s Disease Worldwide and a Discussion of Major Influencing Factors
Possible Factors Influencing the Geographic Variability of Alzheimer’s Disease
3. Applications of Marine Natural Products in Alzheimer’s Disease
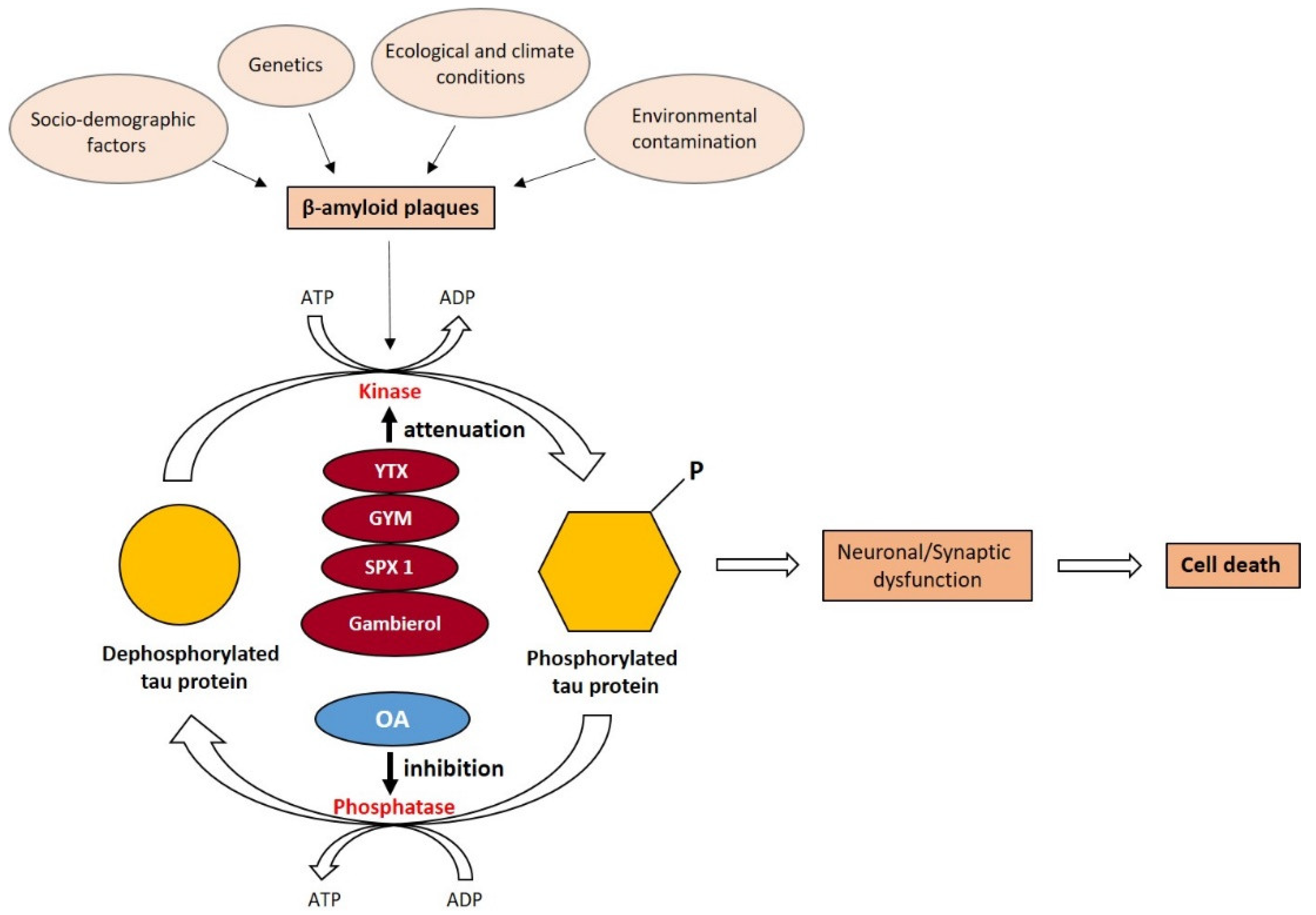
4. Potentialities of Toxins Produced by Marine Dinoflagellates
4.1. Yessotoxins
4.2. Gymnodimine
4.3. Spirolides
4.4. Gambierol
4.5. Okadaic Acid and Its Derivatives
5. Challenges to Alzheimer Disease Therapies from Toxins Produced by Marine Dinoflagellates
5.1. Chemical Synthesis of Marine Toxins
5.2. Large-Scale Cultivation of Producer Organisms
6. Summary and Future Works
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alzheimer’s Association. Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2021, 17, 327. [Google Scholar]
- Bekris, L.M.; Yu, C.E.; Bird, T.D.; Tsuang, D.W. Genetics of Alzheimer disease. J. Geriatr. Psychiatry Neurol. 2010, 23, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isik, A.T. Late onset Alzheimer’s disease in older people. Clin. Interv. Aging 2010, 5, 307–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabir, M.T.; Uddin, M.S.; Jeandet, P.; Emran, T.B.; Mitra, S.; Albadrani, G.M.; Sayed, A.A.; Abdel-Daim, M.M.; Simal-Gandara, J. Anti-Alzheimer’s molecules derived from marine life: Understanding molecular mechanisms and therapeutic potential. Mar. Drugs 2021, 19, 251. [Google Scholar] [CrossRef] [PubMed]
- European Federation of Pharmaceutical Industries and Associations. Alzheimer’s Disease Health System Readiness—The Time to Act Is Now. EFPIA. 2020, pp. 1–6. Available online: www.efpia.eu (accessed on 15 October 2021).
- Xia, X.; Wang, Y.; Zheng, J. COVID-19 and Alzheimer’s disease: How one crisis worsens the other. Transl. Neurodegener. 2021, 10, 15. [Google Scholar] [CrossRef]
- Xiong, N.; Schiller, M.R.; Li, J.; Chen, X.; Lin, Z. Severe COVID-19 in Alzheimer’s disease: APOE4’s fault again? Alzheimer’s Res. Ther. 2021, 13, 111. [Google Scholar] [CrossRef]
- Adefegha, S.A.; Oboh, G.; Olasehinde, T.A. Alkaloid extracts from shea butter and breadfruit as potential inhibitors of monoamine oxidase, cholinesterases, and lipid peroxidation in rats’ brain homogenates: A comparative study. Comp. Clin. Pathol. 2016, 25, 1213–1219. [Google Scholar]
- Olasehinde, T.A.; Olaniran, A.O.; Okoh, A.I. Therapeutic potentials of microalgae in the treatment of Alzheimer’s disease. Molecules 2017, 22, 480. [Google Scholar] [CrossRef] [Green Version]
- Botana, L.M.; Alonso, E.; Vale., C. Use of YTX and Analogues and Derivatives for Treating and/or Preventing Neurodegenerative diseases Linked to Tau and β-Amyloid. European Patent Application EP 2 535 335 A1, 29 August 2012. [Google Scholar]
- Huang, H.C.; Jiang, Z.F. Accumulated amyloid-beta peptide and hyperphosphorylated tau protein: Relationship and links in Alzheimer’s disease. J. Alzheimer’s Dis. 2009, 16, 15–27. [Google Scholar] [CrossRef]
- Castaneda, A.; Ferraz, R.; Vieira, M.; Cardoso, I.; Vasconcelos, V.; Martins, R. Bridging Cyanobacteria to neurodogenerative diseases: A new potential source of bioactive compounds against Alzheimer’s disease. Mar. Drugs 2021, 19, 343. [Google Scholar] [CrossRef]
- Lanctôt, K.L.; Herrmann, N.; Yau, K.K.; Khan, L.R.; Liu, B.A.; Lou, M.M.; Einarson, T.R. Efficacy and safety of cholinesterase inhibitors in Alzheimer’s disease: A meta-analysis. CMAJ 2003, 169, 557–564. [Google Scholar] [PubMed]
- Casey, D.A.; Antimisiaris, D.; O’Brien, J. Drugs for Alzheimer’s Disease: Are they effective? Pharm. Ther. 2010, 35, 208–211. [Google Scholar]
- Verma, A.; Waiker, D.K.; Bhardwaj, B.; Saraf, P.; Shrivastava, S.K. The molecular mechanism, targets, and novel molecules in the treatment of Alzheimer’s disease. Bioinorg. Chem. 2022, 119, 105562. [Google Scholar] [CrossRef] [PubMed]
- Tatulian, A.S. Challenges and hopes for Alzheimer’s disease. Drug Discov. Today 2022, 27, 1027–1043. [Google Scholar] [CrossRef] [PubMed]
- Cotrina, E.Y.; Santos, L.M.; Rivas, J.; Blasi, D.; Leite, J.P.; Liz, M.A.; Busquets, M.A.; Planas, A.; Prohens, R.; Gimeno, A.; et al. Targeting transthyretin in Alzheimer’s disease: Drug discovery of small-molecule chaperones as disease-modifying drug candidates for Alzheimer’s disease. Eur. J. Med. Chem. 2021, 226, 113847. [Google Scholar] [CrossRef]
- Liu, Y.; Han, C.; Dai, R.; Li, B. Rational design, synthesis and activities of hydroxylated chalcones as highly potent dual functional agents against Alzheimer’s disease. Bioorganic Chem. 2022, 122, 105662. [Google Scholar] [CrossRef]
- Cummings, J.; Lee, G.; Zhong, K.; Fonseca, J.; Taghva, K. Alzheimer’s disease drug development pipeline: 2021. Alzheimer’s Dement. Transl. Res. Clin. Interv. 2021, 7, e12179. [Google Scholar] [CrossRef]
- Habtemariam, S. Natural products in Alzheimer’s disease therapy: Would old therapeutic approaches fix the broken promise of modern medicines? Molecules 2019, 24, 1519. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Drew, J.; Berney, W.; Lei, W. Neuroprotective natural products for Alzheimer’s disease. Cells 2021, 10, 1309. [Google Scholar] [CrossRef]
- Noori, T.; Dehpour, A.R.; Sureda, A.; Sobarzo-Sanchez, E.; Shirooie, S. Role of natural products for the treatment of Alzheimer’s disease. Eur. J. Pharmacol. 2021, 898, 173974. [Google Scholar] [CrossRef]
- Leggieri, M.; Thaut, M.H.; Fornazzari, L.; Schweizer, T.A.; Barfett, J.; Munoz, D.G.; Fischer, C.E. Music intervention approaches for Alzheimer’s disease: A review of the literature. Front. Neurosci. 2019, 13, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Haj, M.; Antoine, P.; Nandrino, J.L.; Gély-Nargeot, M.C.; Raffard, S. Self-defining memories during exposure to music in Alzheimer’s disease. Int. Psychogeriatr. 2015, 27, 1719–1730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casaletto, K.; Ramos-Miguel, A.; Vandebunte, A.; Memel, M.; Buchman, A.; Bennett, D.; Honer, W. Late-life physical activity relates to brain tissue synaptic integrity markers in older adults. Alzheimer’s Dement. 2021, 1–13. [Google Scholar] [CrossRef]
- Russo, P.; Kisialiou, A.; Lamonaca, P.; Moroni, R.; Prinzi, G.; Fini, M. New drugs from marine organisms in Alzheimer’s disease. Mar. Drugs 2016, 14, 5. [Google Scholar] [CrossRef] [Green Version]
- Lakshmi, S.; Prakash, P.; Essa, M.M.; Qoronfleh, W.M.; Akbar, M.; Song, B.J.; Kumar, S.; Elimali, P. Marine derived bioactive compounds for treatment of Alzheimer’s disease. Front. Biosci. 2018, 10, 537–548. [Google Scholar]
- Ghoran, S.H.; Kijjoa, A. Marine-derived compounds with anti-Alzheimer’s disease activities. Mar. Drugs 2021, 19, 410. [Google Scholar] [CrossRef]
- Rahman, M.A.; Dash, R.; Sohag, A.A.M.S.; Alam, M.; Rhim, H.; Ha, H.; Moon, I.S.; Uddin, M.J.; Hannan, M.A. Prospects of marine sterols against pathobiology of Alzheimer’s disease: Pharmacological insights and technological advances. Mar. Drugs 2021, 19, 167. [Google Scholar] [CrossRef]
- Lima, E.; Medeiros, J. Marine Organisms as Alkaloid Biosynthesizers of Potential Anti-Alzheimer Agents. Mar. Drugs 2022, 20, 75. [Google Scholar] [CrossRef]
- Barbosa, M.; Valentão, P.; Andrade, P.B. Bioactive compounds from macroalgae in the New Millenium: Implications for neurodegenerative diseases. Mar. Drugs 2014, 12, 4934. [Google Scholar]
- Hielscher-Michael, S.; Griehl, C.; Buchholz, M.; Demuth, H.U.; Arnold, N.; Wessjohann, L.A. Natural products from microalgae with potential against Alzheimer’s disease: Sulfolipids are potent glutaminyl cyclase inhibitors. Mar. Drugs 2016, 14, 203. [Google Scholar] [CrossRef]
- Olasehinde, T.A.; Olaniran, A.O.; Okoh, A.I. Macroalgae as a valuable source of naturally occurring bioactive compounds for the treatment of Alzheimer’s disease. Mar. Drugs 2019, 17, 609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, S.; Jin, W.; Zhang, F.; Linhardt, R.J. The application of seaweed polysaccharides and their derived products with potential for the treatment of Alzheimer’s disease. Mar. Drugs 2021, 19, 89. [Google Scholar] [CrossRef] [PubMed]
- Carreto, J.I.; Carignan, M.O.; Montoya, N.G. Comparative studies on mycosporine-like amino acids, paralytic shellfish toxins and pigment profiles of the toxic dinoflagellates Alexandrium tamarense, A. catenella and A. minutum. Mar. Ecol. Prog. Ser. 2001, 223, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Gallacher, S.; Flynn, K.J.; Franco, J.M.; Brueggemann, E.E.; Hines, H.B. Evidence for production of paralytic shellfish toxins by bacteria associated with Alexandrium spp. (Dinophyta) in culture. Appl. Environ. Microbiol. 1997, 63, 239–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kodama, M.; Ogata, T.; Sakamoto, S. Possible association of marine bacteria with paralytic shellfish toxicity of bivalves. Mar. Ecol. Prog. Ser. 1990, 61, 203–206. [Google Scholar] [CrossRef]
- Gerssen, A.; Pol-Hofstad, I.E.; Poelman, M.; Mulder, P.P.J.; Van den Top, H.J.; De Boer, J. Marine toxins: Chemistry, toxicity, occurrence and detection, with special reference to the Dutch situation. Toxins 2010, 2, 878–904. [Google Scholar] [CrossRef] [Green Version]
- Bricelj, V.M.; Shumway, S.E. Paralytic shellfish toxins in bivalve molluscs: Occurrence, transfer kinetics, and biotransformation. Rev. Fish. Sci. 1998, 6, 315–383. [Google Scholar] [CrossRef]
- Zhang, Z.; Simpkins, J.W. An okadaic acid-induced model of tauopathy and cognitive deficiency. Brain Res. 2010, 1359, 233–246. [Google Scholar] [CrossRef]
- Zhang, Z.; Simpkins, J.W. Okadaic acid induces tau phosphorylation in SH-SY5Y cells in an estrogen-preventable manner. Brain Res. 2010, 1345, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Alonso, E.; Vale, C.; Vieytes, M.R.; Laferla, F.M.; Giménez-Llort, L.; Botana, L.M. 13-Desmethyl spirolide-C is neuroprotective and reduces intracellular Aβ and hyperphosphorylated tau in vitro. Neurochem. Int. 2011, 59, 1056–1065. [Google Scholar] [CrossRef]
- Alonso, E.; Vale, C.; Vieytes, M.R.; Laferla, F.M.; Giménez-Llort, L.; Botana, L.M. The Cholinergic Antagonist Gymnodimine Improves Aβ and Tau Neuropathology in an in Vitro Model of Alzheimer’s Disease. Cell Physiol. Biochem. 2011, 27, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Botana, L.M.; Alonso, E.; Vale, C. Use of Gymnodimine, Analogues, and Derivatives for the Treatment and/or Prevention of Neurodegenerative Diseases Associated to Tau and β-Amyloid. U.S. Patent 2012/0245223 A1, 27 September 2012. [Google Scholar]
- Alonso, E.; Vale, C.; Vieytes, M.R.; Botana, L.M. Translocation of PKC by yessotoxin in an in vitro model of Alzheimer’s disease with Improvement of tau and β-amyloid pathology. ACS Chem. Neurosci. 2013, 4, 1062–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamat, P.K.; Rai, S.; Nath, C. Okadaic acid induced neurotoxicity: An emerging tool to study Alzheimer’s disease pathology. Neurotoxicology 2013, 37, 163–172. [Google Scholar] [CrossRef]
- Alonso, E.; Vieira, A.C.; Rodriguez, I.; Alvariño, R.; Gegunde, S.; Fuwa, H.; Suga, Y.; Sasaki, M.; Alfonso, A.; Cifuentes, J.M.; et al. Tetracyclic truncated analogue of the marine toxin gambierol modifies NMDA, tau, and amyloid β expression in mice brains: Implications in AD pathology. ACS Chem. Neurosci. 2017, 8, 1358–1367. [Google Scholar] [CrossRef] [PubMed]
- World Life Expectancy. Available online: https://www.worldlifeexpectancy.com/cause-of-death/alzheimers-dementia/by-country/ (accessed on 17 March 2022).
- Population Reference Bureau. The Top 50 Countries with the Largest Percentage of Older Adults. 2021. Available online: https://www.prb.org/resources/countries-with-the-oldest-populations-in-the-world/ (accessed on 15 October 2021).
- Eiser, A.R. Why does Finland have the highest dementia mortality rate? Environmental factors may be generalizable. Brain Res. 2017, 1671, 14–17. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Kiselova, N.; Rosen, J.; Ilag, L.L. Quantification of neurotoxin BMAA (b-N-methylamino-L-alanine) in seafood from Swedish markets. Sci. Rep. 2014, 4, 6931. [Google Scholar] [CrossRef] [Green Version]
- Piemontese, L. Plant food supplements with antioxidant properties for the treatment of chronic and neurodegenerative diseases: Benefits or risks? J. Diet. Suppl. 2017, 14, 478–484. [Google Scholar] [CrossRef]
- Crapper, D.R.; Krishnan, A.J.; Dalton, A.J. Brain aluminium distribution in Alzheimer’s disease and experimental neurofibrillary degeneration. Science 1973, 180, 511–513. [Google Scholar] [CrossRef]
- Pick, F.R. Blooming algae: A Canadian perspective on the rise of toxic cyanobacteria. Can. J. Fish. Aquat. Sci. 2016, 73, 1149. [Google Scholar] [CrossRef] [Green Version]
- Perl, D.P.; Brody, A.R. Alzheimer’s disease—X-ray spectrometric evidence of aluminium accumulation in neurofibrillary tangle-bearing neurons. Science 1980, 208, 297–299. [Google Scholar] [CrossRef]
- Exley, C. What is the risk of aluminium as a neurotoxin? Expert Rev. Neurother. 2014, 14, 589–591. [Google Scholar] [CrossRef] [PubMed]
- Van Dyke, N.; Yenugadhati, N.; Birkett, N.J.; Lindsay, J.; Turner, M.C.; Willhite, C.C.; Krewski, D. Association between aluminum in drinking water and incident Alzheimer’s disease in the Canadian study of health and aging cohort. Neurotoxicology 2021, 83, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Guo, J.-L.; Wang, L.; Zhong, X.; Yao, W.-F.; Gao, H.; Liu, M.-Y. Natural products as pharmacological modulators of mitochondrial dysfunctions for the treatments of Alzheimer’s disease: A comprehensive review. Eur. J. Med. Chem. 2021, 218, 113401. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Seijas, P.; Otero, P. Exploitation of marine molecules to manage Alzheimer’s disease. Mar. Drugs 2021, 19, 373. [Google Scholar] [CrossRef]
- Bui, T.T.; Nguyen, T.H. Natural product for the treatment of Alzheimer’s disease. J. Basic Clin. Physiol. Pharmacol. 2017, 28, 413–423. [Google Scholar] [CrossRef]
- Leiteritz, A.; Dilberger, B.; Wenzel, U.; Fitzenberger, E. Betaine reduces β-amyloid-induced paralysis through activation of cystathionine-β-synthase in an Alzheimer model of Caenorhabditis elegans. Genes Nutr. 2018, 13, 21. [Google Scholar] [CrossRef] [Green Version]
- Schrott, L.M.; Jackson, K.; Yi, P.; Dietz, F.; Johnson, G.S.; Basting, T.F.; Purdum, G.; Tyler, T.; Rios, J.D.; Castor, T.P.; et al. Acute oral bryostatin-1 administration improves learning deficits in the APP/PS1 transgenic mouse model of Alzheimer’s disease. Curr. Alzheimer Res. 2015, 12, 22–31. [Google Scholar] [CrossRef]
- Bazan, N.G. Cellular and molecular events mediated by docosahexaenoic acid-derived neuroprotectin D1 signaling in photoreceptor cell survival and brain protection. Prostaglandins Leukot. Essent. Fat. Acids 2009, 81, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Castro-Silva, E.S.; Bello, M.; Rosales-Hernández, M.C.; Correa-Basurto, J.; Hernández-Rodríguez, M.; Villalobos-Acosta, D.; Méndez-Méndez, J.V.; Estrada-Pérez, A.; Murillo-Álvarez, J.; Muñoz-Ochoa, M. Fucosterol from Sargassum horridum as an amyloid-beta (Aβ1-42) aggregation inhibitor: In vitro and in silico studies. J. Biomol. Struct. Dyn. 2021, 39, 1271–1283. [Google Scholar] [CrossRef]
- Hannan, M.A.; Dash, R.; Haque, M.N.; Mohibbullah, M.; Sohag, A.A.M.; Rahman, M.A.; Uddin, M.J.; Alam, M.; Moon, I.S. Neuroprotective Potentials of Marine Algae and Their Bioactive Metabolites: Pharmacological Insights and Therapeutic Advances. Mar. Drugs 2020, 18, 347. [Google Scholar] [CrossRef]
- Cotas, J.; Leandro, A.; Pacheco, D.; Gonçalves, A.M.M.; Pereira, L. A comprehensive review of the nutraceutical and therapeutic applications of red seaweeds (Rhodophyta). Life 2020, 10, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, D.; Yao, L.; Lin, Z.; Chi, L.; Li, H.; Xu, H.; Du, X.; Liu, Q.; Hu, Z.; Lu, J.; et al. Unsaturated mannuronate oligosaccharide ameliorates β-amyloid pathology through autophagy in Alzheimer’s disease cell models. Carbohydr. Polym. 2021, 251, 117124. [Google Scholar] [CrossRef] [PubMed]
- Kellmann, R.; Stüken, A.; Orr, R.J.S.; Svendsen, H.M.; Jakobsen, K.S. Biosynthesis and molecular genetics of polyketides in marine dinoflagellates. Mar. Drugs 2010, 8, 1011–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cembella, A.D. Chemical ecology of eukaryotic microalgae in marine ecosystems. Phycologia 2003, 42, 420–447. [Google Scholar] [CrossRef]
- Camacho-Muñoz, D.; Praptiwi, R.A.; Lawton, L.A.; Edwards, C. High value phycotoxins from the dinoflagellate Prorocentrum. Front. Mar. Sci. 2021, 8, 638739. [Google Scholar] [CrossRef]
- Satake, M.; MacKenzie, L.; Yasumoto, T. Identification of Protoceratium reticulatum as the biogenetic origin of yessotoxin. Nat. Toxins 1998, 5, 164–167. [Google Scholar] [CrossRef]
- Paz, B.; Riobó, P.; Fernández, M.L.; Fraga, S.; Franco, J.M. Production and release of yessotoxins by the dinoflagellates Protoceratium reticulatum and Lingulodinium polyedrum in culture. Toxicon 2004, 44, 251–258. [Google Scholar] [CrossRef]
- Armstrong, M.; Kudela, R. Evaluation of California isolates of Lingulodinium polyedrum for the production of yessotoxin. Afr. J. Mar. Sci. 2006, 28, 399–401. [Google Scholar] [CrossRef]
- Rhodes, L.; McNabb, P.; de Salas, M.; Briggs, L.; Beuzenberg, V.; Gladstone, M. Yessotoxin production by Gonyaulax spinifera. Harmful Algae 2006, 5, 148–155. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Marine biotoxins in shellfish—Yessotoxin group. EFSA J. 2008, 907, 1–62. [Google Scholar]
- Tubaro, A.; Giangaspero, A.; Ardizzone, M.; Soranzo, M.R.; Vita, F.; Yasumoto, T.; Maucher, J.M.; Ramsdelle, J.S.; Sosa, S. Ultrastructural damage to heart tissue from repeated oral exposure to yessotoxin resolves in 3 months. Toxicon 2008, 51, 1225–1235. [Google Scholar] [CrossRef] [PubMed]
- Ferreiro, S.F.; Carrera, C.; Vilariño, N.; Louza, M.C.; Santamarina, G.; Cantalapiedra, A.G.; Botana, L.M. Acute cardiotoxicity evaluation of the marine biotoxins OA, DTX-1 and YTX. Toxins 2015, 7, 1030–1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfonso, A.; Vieytes, M.R.; Botana, L.M. Yessotoxin, a Promising Therapeutic Tool. Mar. Drugs 2016, 14, 30. [Google Scholar] [CrossRef] [Green Version]
- Martín-López, A.; Rodríguez, J.G.; Mirón, A.S.; García-Camacho, F.; Grima, E.M. Immunoregulatory potential of marine algal toxins yessotoxin and okadaic acid in mouse T lymphocyte cell line EL-4. Toxicol. Lett. 2011, 207, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Cembella, A.; Krock, B. Cyclic imine toxins: Chemistry, biogeography, biosynthesis and pharmacology. In Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection, 2nd ed.; Botana, L.M., Ed.; CRC Press (Taylor and Francis Group): Boca Raton, FL, USA, 2008; pp. 561–580. [Google Scholar]
- Selwood, A.I.; Miles, C.O.; Wilkins, A.L.; van Ginkel, R.; Munday, R.; Rise, F.; McNabb, P. Isolation, structural determination and acute toxicity of pinnatoxins E, F and G. J. Agric. Food Chem. 2010, 58, 6532–6542. [Google Scholar] [CrossRef]
- Stivala, C.E.; Benoit, E.; Aráoz, R.; Servent, D.; Novikov, A.; Molgó, J.; Zakarian, A. Synthesis and biology of cyclic imine toxins, an emerging class of potent, globally distributed marine toxins. Nat. Prod. Rep. 2015, 32, 411–435. [Google Scholar] [CrossRef]
- Seki, T.; Satake, M.; Mackenzie, L.; Kaspar, H.F.; Yasumoto, T. Gymnodimine, a new marine toxin of unprecedented structure isolated from New Zealand oysters and the dinoflagellate, Gymnodinium sp. Tetrahedron Lett. 1995, 36, 7093–7096. [Google Scholar] [CrossRef]
- Van Wagoner, R.M.; Misner, I.; Tomas, C.R.; Wright, J.L.C. Occurrence of 12-methylgymnodimine in a spirolide-producing dinoflagellate Alexandrium peruvianum and the biogenetic implications. Tetrahedron Lett. 2011, 52, 4243–4246. [Google Scholar] [CrossRef]
- Harju, K.; Koskela, H.; Kremp, A.; Sanna Suikkanen, S.; de la Iglesia, P.; Miles, C.O.; Krock, B.; Vanninen, P. Identification of gymnodimine D and presence of gymnodimine variants in the dinoflagellate Alexandrium ostenfeldii from the Baltic Sea. Toxicon 2016, 112, 68–76. [Google Scholar] [CrossRef] [Green Version]
- Bourne, Y.; Radi, Z.; Aráoz, R.; Talleyb, T.T.; Benoit, E.; Servent, D.; Taylor, P.; Jordi Molgó, J.; Marchot, P. Structural determinants in phycotoxins and AChBP conferring high affinity binding and nicotinic AChR antagonism. Proc. Natl. Acad. Sci. USA 2010, 107, 6076–6081. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Marine biotoxins in shellfish—Cyclic imines (spirolides, gymnodimines, pinnatoxins and pteriatoxins). EFSA J. 2010, 8, 1628. [Google Scholar]
- Sosa, S.; Pelin, M.; Cavion, F.; Hervé, F.; Hess, P.; Tubaro, A. Acute Oral Toxicity of Pinnatoxin G in Mice. Toxins 2020, 12, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botana, L.M.; Alonso, E.; Vale, C. Use of Gambierol for Treating and/or Preventing Neurodegenerative Diseases Related to Tau and Beta-Amyloid. U.S. Patent 2012/0283321 A1, 8 November 2012. [Google Scholar]
- Touzet, N.; Franco, J.M.; Raine, R. Morphogenetic diversity and biotoxin composition of Alexandrium (Dinophyceae) in Irish coastal waters. Harmful Algae 2008, 7, 782–797. [Google Scholar] [CrossRef]
- Hu, T.; Burton, I.W.; Cembella, A.D.; Curtis, J.M.; Quilliam, M.A.; John, A.; Walter, J.A.; Wright, J.L.C. Characterization of Spirolides A, C, and 13-Desmethyl C, New Marine Toxins Isolated from Toxic Plankton and Contaminated Shellfish. J. Nat. Prod. 2001, 64, 308–312. [Google Scholar] [CrossRef] [PubMed]
- Otero, P.; Alfonso, A.; Rodríguez, P.; Rubiolo, J.A.; Cifuentes, J.M.; Roberto Bermúdez, R.; Vieytes, M.R.; Luis, M.; Botana, L.M. Pharmacokinetic and toxicological data of spirolides after oral and intraperitoneal administration. Food Chem. Toxicol. 2012, 50, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, M.; Hellström-Lindahl, E. Nicotinic receptor agonists and antagonists increase sAPPa secretion and decrease Ab levels in vitro. Neurochem. Int. 2009, 54, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Satake, M.; Michio Murata, M.; Takeshi Yasumoto, T. Gambierol: A new toxic polyether compound isolated from the marine dinoflagellate Gambierdiscus toxicus. J. Am. Chem. Soc. 1993, 115, 361–362. [Google Scholar] [CrossRef]
- Morohashi, A.; Satake, M.; Nagai, H.; Oshima, Y. The Absolute configuration of gambieric acids A–D, potent antifungal polyethers, isolated from the marine dinoflagellate Gambierdiscus toxicus. Tetrahedron 2000, 56, 8995–9001. [Google Scholar] [CrossRef]
- Chinain, M.; Gatti, C.M.I.; Darius, H.T.; Quod, J.P.; Tester, P.A. Ciguatera poisonings: A global review of occurrences and trends. Harmful Algae 2021, 102, 101873. [Google Scholar] [CrossRef]
- Yasumoto, T. The chemistry and biological function of natural marine toxins. Chem. Rec. 2001, 1, 228–242. [Google Scholar] [CrossRef]
- Ito, A.; Sazuki, F.; Toshimori, K.; Fuwa, H.; Tachibana, K.; Satake, M.; Sazaki, M. Pathological effects on mice by gambierol, possibly one of the ciguatera toxins. Toxicon 2003, 42, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Cuypers, E.; Abdel-Mottaleb, Y.; Kopljar, I.; Rainier, J.D.; Raes, A.L.; Snyders, D.J.; Tytgat, J. Gambierol, a toxin produced by the dinoflagellate Gambierdiscus toxicus, is a potent blocker of voltage-gated potassium channels. Toxicon 2008, 51, 974–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghiaroni, V.; Sasaki, M.; Fuwa, H.; Rossini, G.P.; Scalera, G.; Takeshi Yasumoto, T.; Pietra, P.; Bigiani, A. Inhibition of voltage-gated potassium currents by gambierol in mouse taste cells. Toxicol. Sci. 2005, 85, 657–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assunção, J.; Guedes, A.C.; Malcata, F.X. Biotechnological and pharmacological applications of biotoxins and other bioactive molecules from dinoflagellates. Mar. Drugs 2017, 15, 393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morton, S.L.; Vershinin, A.; Smith, L.L.; Leighfield, T.A.; Pankov, S.; Quilliam, M.A. Seasonality of Dinophysis spp. and Prorocentrum lima in Black Sea phytoplankton and associated shellfish toxicity. Harmful Algae 2009, 8, 629–636. [Google Scholar] [CrossRef]
- Reguera, B.; Velo-Suárez, L.; Raine, R.; Park, M.G. Harmful Dinophysis species: A review. Harmful Algae 2012, 14, 87–106. [Google Scholar] [CrossRef]
- Yasumoto, T.; Oshima, Y.; Sugawara, W.; Fukuyo, Y.; Oguri, H.; Igarashi, T.; Fujita, N. Identification of Dinophysis fortii as the causative organism of diarrhetic shellfish poisoning. Nippon. Suisan Gakkaishi 1980, 46, 1405–1411. [Google Scholar] [CrossRef] [Green Version]
- Garcia, C.; Truan, D.; Lagos, M.; Santelices, J.P.; Diaz, J.C.; Lagos, N. Metabolic transformation of dinophysistoxin-3 into dinophysistoxin-1 causes human intoxication by consumption of O-acyl-derivatives dinophysistoxins contaminated shellfish. J. Toxicol. Sci. 2005, 30, 287–296. [Google Scholar] [CrossRef] [Green Version]
- Bialojan, C.; Takai, A. Inhibitory effect of a marine-sponge toxin, okadaic acid, on protein phosphatases. Specificity and kinetics. Biochem. J. 1988, 256, 283–290. [Google Scholar] [CrossRef]
- Wera, S.; Hemmings, B.A. Serine/threonine protein phosphatases. Biochem. J. 1995, 311, 17–29. [Google Scholar] [CrossRef]
- Arias, C.; Becerra-García, F.; Arrieta, I.; Tapia, R. The protein phosphatase inhibitor okadaic acid induces heat shock protein expression and neurodegeneration in rat hippocampus in vivo. Exp. Neurol. 1998, 153, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.X.G.; Singh, T.J.; Grundke-Iqbal, I.; Iqbal, K. Phosphoprotein phosphatase activities in Alzheimer disease brain. J. Neurochem. 1993, 61, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Torrent, L.; Ferrer, I. PP2A and Alzheimer disease. Curr. Alzheimer Res. 2012, 9, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, E.S.; Shin, R.-W.; Billingsley, M.L.; Van de Voorde, A.; O’Connor, M.; Trojanowski, J.Q.; Lee, V.M.Y. Biopsy-derived adult human brain tau is phosphorylated at many of the same sites as Alzheimer’s disease paired helical filament tau. Neuron 1994, 13, 989–1002. [Google Scholar] [CrossRef]
- Costa, A.P.; Tramontina, A.C.; Biasibetti, R.; Batassini, C.; Lopes, M.W.; Wartchow, K.M.; Bernardi, C.; Tortorelli, L.S.; Leal, R.B.; Gonçalves, C.A. Neuroglial alterations in rats submitted to the okadaic acid-induced model of dementia. Behav. Brain Res. 2012, 226, 420–427. [Google Scholar] [CrossRef]
- Koehler, D.; Shah, Z.A.; Hensley, K.; Williams, F.E. Lanthionine ketimine-5-ethyl ester provides neuroprotection in a zebrafish model of okadaic acid-induced Alzheimer’s disease. Neurochem. Int. 2018, 115, 61–68. [Google Scholar] [CrossRef]
- Koehler, D.; Shah, Z.A.; Williams, F.E. The GSK3β inhibitor, TDZD-8, rescues cognition in a zebrafish model of okadaic acid-induced Alzheimer’s disease. Neurochem. Int. 2019, 122, 31–37. [Google Scholar] [CrossRef]
- Smayda, T.J.; Trainer, V.L. Dinoflagellate blooms in upwelling systems: Seeding, variability, and contrasts with diatom bloom behaviour. Prog. Oceanogr. 2010, 85, 92–107. [Google Scholar] [CrossRef]
- Díaz, P.A.; Reguera, B.; Ruiz-Villarreal, M.; Pazos, Y.; Velo-Suárez, L.; Berger, H.; Sourisseau, M. Climate variability and oceanographic settings associated with interannual variability in the initiation of Dinophysis acuminata blooms. Mar. Drugs 2013, 11, 2964–2981. [Google Scholar] [CrossRef] [Green Version]
- Gallardo-Rodríguez, J.J.; Sánchez-Mirón, A.; García-Camacho, F.; López-rosales, L.; Chisti, Y.; Molina-Grima, E. Bioactives from microalgal dinoflagellates. Biotechnol. Adv. 2012, 30, 1673–1684. [Google Scholar] [CrossRef]
- Isobe, M.; Ichikawa, Y.; Goto, T. Synthetic studies toward marine toxic polyethers: The total synthesis of okadaic acid. Tetrahedron Lett. 1986, 27, 963–966. [Google Scholar] [CrossRef]
- Nicolaou, K.C.; Rutjes, F.P.J.T.; Theodorakis, E.A.; Tiebes, J.; Sato, M.; Untersteller, E. Total synthesis of brevetoxin B. 3. Final strategy and completion. J. Am. Chem. Soc. 1995, 117, 10252–10263. [Google Scholar] [CrossRef] [Green Version]
- Nicolaou, K.C.; Gunzner, J.L.; Shi, G.; Agrios, K.A.; Gärtner, P.; Yang, Z. Total synthesis of brevetoxin A: Part 4: Final stages and completion. Chem. Eur. J. 1999, 5, 646–658. [Google Scholar] [CrossRef]
- Forsyth, C.J.; Sabes, S.F.; Urbanek, R.A. An efficient total synthesis of okadaic acid. J. Am. Chem. Soc. 1997, 119, 8381–8382. [Google Scholar] [CrossRef]
- Pang, Y.; Fang, C.; Twiner, M.J.; Miles, C.O.; Forsyth, C.J. Total synthesis of dinophysistoxin-2 and 2-epi-dinophysistoxin-2 and their PPase inhibition. Angew. Chem. Int. 2011, 50, 7631–7635. [Google Scholar] [CrossRef]
- Fuwa, H.; Ishigai, K.; Hashizume, K.; Sasaki, M. Total synthesis and complete stereostructure of gambieric acid A. J. Am. Chem. Soc. 2012, 134, 11984–11987. [Google Scholar] [CrossRef]
- Ley, S.V.; Humphries, A.C.; Eick, H.; Downham, R.; Ross, A.R.; Boyce, R.; Pavey, J.B.J.; Pietrusza, J. Total synthesis of the protein phosphatase inhibitor okadaic acid. J. Chem. Soc. Perkin Trans. 1998, 1, 3907–3912. [Google Scholar] [CrossRef]
- Nicholas, G.M.; Phillips, A.J. Marine natural products: Synthetic aspects. Nat. Prod. Rep. 2006, 23, 79–99. [Google Scholar] [CrossRef]
- Armstrong, R.W.; Beau, J.M.; Cheon, S.H.; Christ, W.J.; Fujioka, H.; Ham, W.H.; Hawkins, L.D.; Jin, H.; Kang, S.H. Total synthesis of palytoxin carboxylic acid and palytoxin amide. J. Am. Chem. Soc. 1989, 111, 7530–7533. [Google Scholar] [CrossRef]
- Doblin, M.A.; Thompson, P.A.; Revilla, A.T.; Butler, E.C.V.; Blackburn, S.I.; Hallegraeff, G.M. Vertical migration of the toxic dinoflagellate Gymnodinium catenatum under different concentrations of nutrients and humic substances in culture. Harmful Algae 2006, 5, 665–677. [Google Scholar] [CrossRef]
- Pan, Y.; Cembella, A.D.; Quilliam, M.A. Cell cycle and toxin production in the benthic dinoflagellate Prorocentrum lima. Mar. Biol. 1999, 134, 541–549. [Google Scholar] [CrossRef]
- Pistocchi, R.; Guerrini, F.; Pezzolesi, L.; Riccardi, M.; Vanucci, S.; Ciminiello, P.; Dell’Aversano, C.; Forino, M.; Fattorusso, E.; Tartaglione, L.; et al. Toxin levels and profiles in microalgae from the north-western adriatic sea—15 years of studies on cultured species. Mar. Drugs 2012, 10, 140–162. [Google Scholar] [CrossRef] [Green Version]
- Gallardo-Rodríguez, J.J.; Sánchez-Mirón, A.; Cerón-Garcia, M.C.; Belarbia, E.; García-Camacho, F.; Chisti, Y.; Molina-Grima, E. Macronutrients requirements of the dinoflagellate Protoceratium reticulatum. Harmful Algae 2009, 8, 239–246. [Google Scholar] [CrossRef]
- Gallardo-Rodríguez, J.J.; Sánchez-Mirón, A.; García-Camacho, F.; Cerón-García, M.; Belarbi, E.H.; Molina-Grima, E. Culture of dinoflagellates in a fed-batch and continuous stirred-tank photobioreactors: Growth, oxidative stress and toxin production. Process Biochem. 2010, 45, 660–666. [Google Scholar] [CrossRef]
- Wang, S.; Chen, J.; Li, Z.; Wang, Y.; Fu, B.; Han, X.; Zheng, L. Cultivation of the benthic microalga Prorocentrum lima for the production of diarrhetic shellfish poisoning toxins in a vertical flat photobioreactor. Bioresour. Technol. 2015, 179, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Loader, J.I.; Hawkes, A.D.; Beuzenberg, V.; Jensen, D.J.; Cooney, J.M.; Wilkins, A.L.; Fitzgerald, J.M.; Briggs, L.R.; Miles, C.O. Convenient large-scale purification of yessotoxin from Protoceratium reticulatum culture and isolation of a novel furano yessotoxin. J. Agric. Food Chem. 2007, 55, 11093–11100. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-J. Genomic understanding of dinoflagellates. Res. Microbiol. 2011, 162, 551–569. [Google Scholar] [CrossRef]
- Verma, A.; Barua, A.; Ruvindy, R.; Savela, H.; Ajani, P.A.; Murray, S.A. The genetic basis of toxin biosynthesis in dinoflagellates. Microorganisms 2019, 7, 222. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Classification | Country | Rate |
|---|---|---|
| Very high | Turkey | 58 |
| Lebanon | 56 | |
| Libya | 53 | |
| Finland, Equatorial Guinea, Tunisia, Yemen, Jordan, Saudi Arabia, Morocco, Nigeria, Qatar, Iran | 51–42 | |
| Indonesia, Syria, Cambodia, Kiribati, Thailand, Laos, Timor-Leste, Mali, Myanmar, United Kingdom, Algeria | 41–38 | |
| Mauritania, Gabon, Malaysia, Gambia, Egypt, Arab Emirates, Maldives, Sri Lanka, China, Burkina Faso, Sierra Leone | 37–35 | |
| Afghanistan, Namibia, Sudan, Comoros, Togo, Angola, Netherlands, Bahrain | 34–33 | |
| High | United States, Micronesia, Djibouti, DR Congo, Brunei, Oman, Senegal, Congo, Seychelles, Guinea, Cote d’Ivoire, Paraguay, Iraq | 32–31 |
| South Africa, Ghana, Niger, Malawi, El Salvador, Iceland, Nicaragua, Zimbabwe, Belize, Rwanda, Tonga | 30–29 | |
| Botswana, Samoa, Sweden, Cameroon, Liberia, Mozambique, Vietnam, Chad, Benin, Cape Verde, Central Africa, Ireland, Uganda, Tanzania | 28–27 | |
| Georgia, Solomon Islands, North Korea, Peru, Vanuatu | 26 | |
| Medium | Nepal, Eritrea, Switzerland, Suriname, Denmark, Ethiopia, Albania, Guinea-Bissau, Norway, Swaziland, Canada, Bosnia and Herzegovina, Montenegro | 25–24 |
| Burundi, Australia, Zambia, South Sudan, Honduras, Kazakhstan, Slovakia, Somalia, Belgium, Tajikistan, Lesotho, Spain | 23–22 | |
| Armenia, Turkmenistan, Pakistan, Sao Tome, New Zealand, Cuba, Kenya, Haiti, New Guinea, Mongolia, Bolivia, Azerbaijan, France, Dominican Republic, Belarus, Madagascar, Ukraine, Jamaica | 20–18 | |
| Low | Barbados, Bhutan, Russia, India, Luxembourg, Bahamas, Uruguay, Portugal, Germany, Cyprus, Hungary, Israel, South Korea, Italy, Malta, Bangladesh | 17–14 |
| Chile, Czech Republic, Brazil, Serbia, Costa Rica, Austria, Croatia, Trinidad/Tobago | 12–8 | |
| Panama, Greece, Japan, Latvia, Antigua and Barbuda, Argentina, Grenada, Lithuania, Estonia, Romania, Ecuador, Saint Vincent, Guatemala, Guyana, Colombia, Moldova | 6–4 | |
| Mexico, Poland, Slovenia, Venezuela, Mauritius, Saint Lucia, Uzbekistan, Philippines, North Macedonia, Kyrgyzstan, Bulgaria | 3–1 | |
| Kuwait, Fiji, Singapore | <1 |
| Source | Compound | Chemical Structure | Anti-Alzheimer Activity Mechanism | Reference |
|---|---|---|---|---|
| Amphiporus spp. | 3-(2,4-Dimethoxybenzylidene)-anabaseine |  | Stimulation of nicotinic acetylcholine receptors (nAChR) | [4] |
| Seafood Spinach Seaweeds | Betaine |  | Trimethyl derivative of glycine, labile donor of methyl groups. Improve sulphur aminoacids metabolism protecting against oxidative stress | [61] |
| Bugula neritina | Bryostatin-1 |  | Reduces amyloid plaque. Activator of PKC revealed to prevent synaptic loss with increasing synaptic maturation | [62] |
| Seafood | Docosahexaenoic acid (DHA) |  | Protective against the dendritic pathology associated with expression of mutated amyloid precursor protein (APP). | [22] |
| Sargassum horridum | Fucosterol |  | Neuroprotective agent with anti-amyloid properties | [64] |
| Caulerpa racemosa | Caulerpenyne |  | Anti-inflammatory activity | [65] |
| Dictyopteris undulate | Zonarol |  | Neuroprotective activity | [65] |
| Brown seaweeds | Mannuronate oligosaccharide (MOS) |  | Inhibits the tau protein aggregation. Attenuates the phosphorylation of tau protein | [66] |
| Source | Compound | Chemical Structure | Relation with Anti-AD Activity | Reference |
|---|---|---|---|---|
| Protoceratium reticulatum; Lingulodinium polyedrum; Gonyaulax spinifera | Yessotoxin |  | Reduces amyloid plaque and the tau hyperphosphorylation | [45] |
| Karenia selliformis | Gymnodimine |  | Reduces amyloid plaque and the tau hyperphosphorylation | [43] |
| Alexandrium ostenfelii; A. peruvianum | 13-desmethyl spirolide C |  | Reduces the tau hyperphosphorylation; Neuroprotective activity | [42] |
| Gambierdiscus toxicus | Gambierol |  | Inhibits the voltage-dependent Na and K channels; Reduces the tau hyperphosphorylation and the overexpression of B-amyloid | [47] |
| Dinophysis acuta; D. acuminata; D. caudata;
D. fortii; D. infundibulum; D. miles; D. norvegica; D. ovum; D. sacculus; Prorocentrum arenarium P. belizeanium; P. concavum P. lima | Okadaic acid |  | Inhibits the serine/threonine protein phosphatases 1 and 2A | [40,41] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Botelho, M.J.; Milinovic, J.; Bandarra, N.M.; Vale, C. Alzheimer’s Disease and Toxins Produced by Marine Dinoflagellates: An Issue to Explore. Mar. Drugs 2022, 20, 253. https://doi.org/10.3390/md20040253
Botelho MJ, Milinovic J, Bandarra NM, Vale C. Alzheimer’s Disease and Toxins Produced by Marine Dinoflagellates: An Issue to Explore. Marine Drugs. 2022; 20(4):253. https://doi.org/10.3390/md20040253
Chicago/Turabian StyleBotelho, Maria João, Jelena Milinovic, Narcisa M. Bandarra, and Carlos Vale. 2022. "Alzheimer’s Disease and Toxins Produced by Marine Dinoflagellates: An Issue to Explore" Marine Drugs 20, no. 4: 253. https://doi.org/10.3390/md20040253