
Evaluating Age and Growth Relationship to Ciguatoxicity in Five Coral Reef Fish Species from French Polynesia

 ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
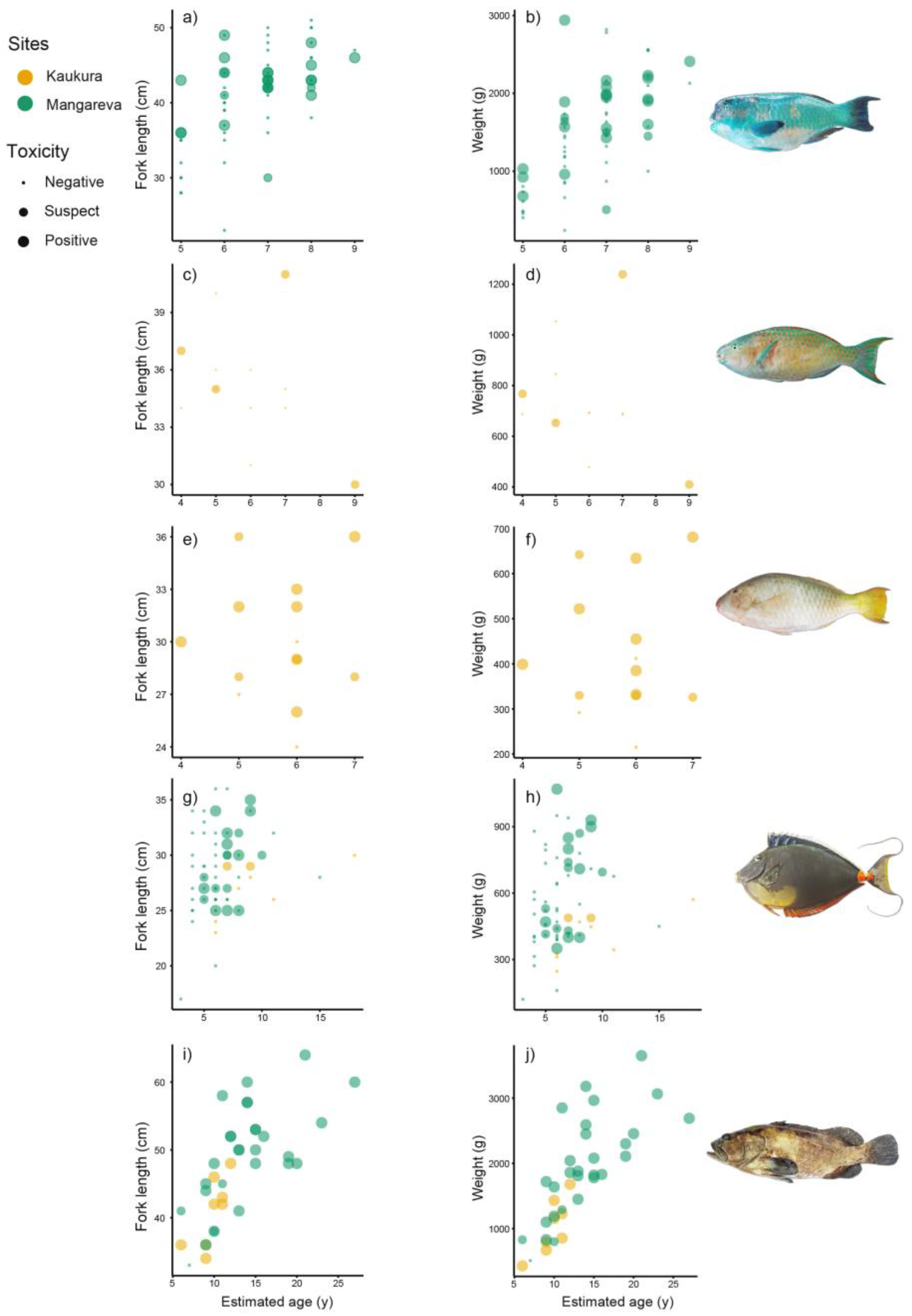
2.1. Fish Demographic Data
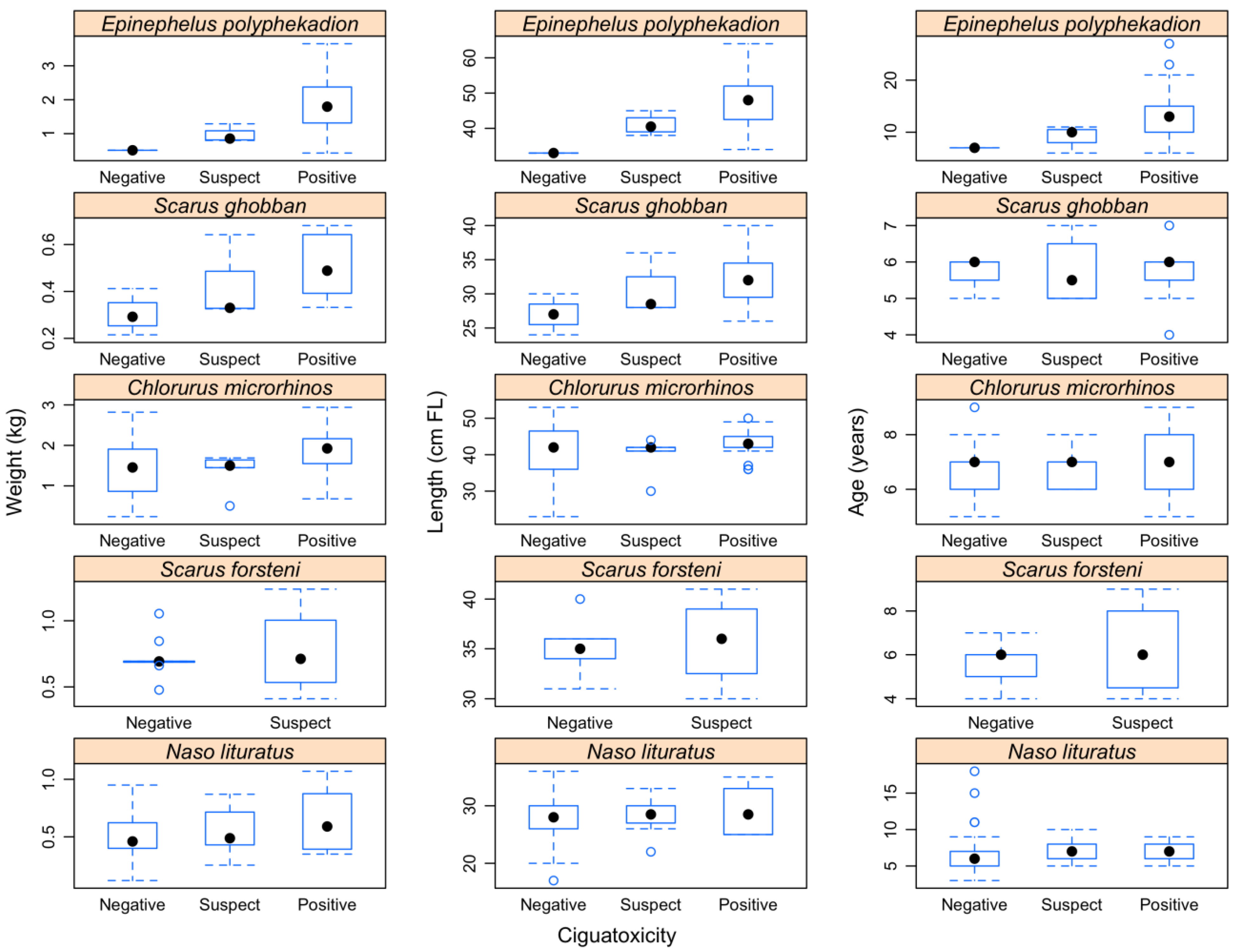
2.2. Relationship between Size, Weight, and Age and Fish Ciguatoxicity
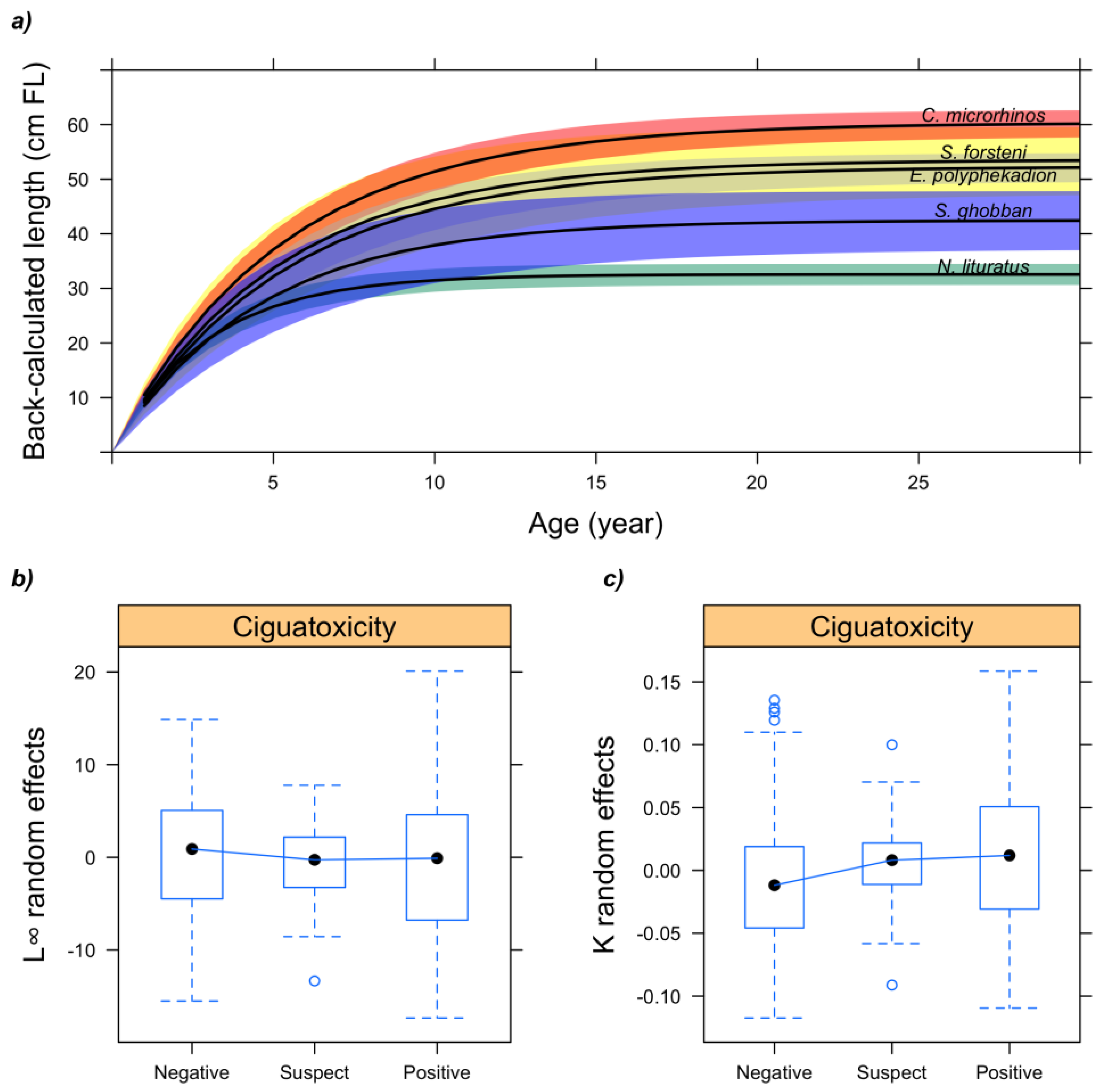
2.3. Relationship between Fish Ciguatoxicity and Growth Characteristics

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) | Model | Fixed Effect | Random Effect | df | AIC | BIC | logLik | Test | L.Ratio | p-Value |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Intercept | K, L∞ | 6 | 7440 | 7472 | −3714 | ||||
| 2 | Species K, L∞ | K, L∞ | 14 | 7247 | 7323 | −3609 | 1 vs. 2 | 208 | <0.0001 | |
| 3 | Species K, L∞ | L∞ | 12 | 7869 | 7934 | −3922 | 2 vs. 3 | 625 | <0.0001 | |
| 4 | Species K, L∞ | K | 12 | 7863 | 7928 | −3919 | 2 vs. 4 | 619 | <0.0001 | |
| (b) | Fixed Effects | numDF | denDF | F-Value | p-Value | |||||
| L∞.(Intercept) | 1 | 1531 | 21,467 | <0.0001 | ||||||
| L∞.Species | 4 | 1531 | 139 | <0.0001 | ||||||
| K.(Intercept) | 1 | 1531 | 1941 | <0.0001 | ||||||
| K.Species | 4 | 1531 | 37 | <0.0001 |
2.4. Influence of Other Factors on Fish Ciguatoxicity
2.4.1. Sampling Season
2.4.2. Environmental Features of Sampling Areas
2.4.3. Trophic Level
2.4.4. CTX Bioaccumulation and Depuration Kinetics
2.5. Fish Ciguatoxicity versus Epidemiological Data in the Two Study Sites
3. Materials and Methods
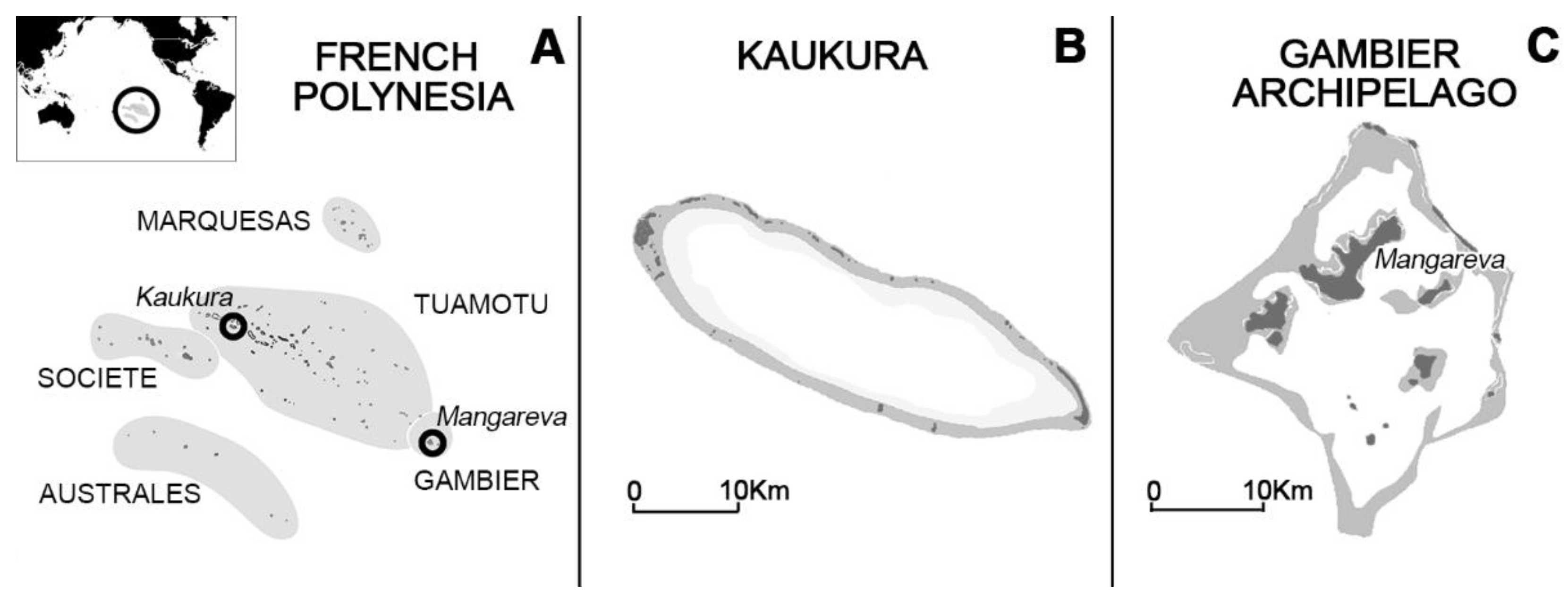
3.1. Study Area
3.2. Fish Sampling
3.3. CTXs Chemical Extraction
3.4. Neuroblastoma Cell-Based Assay (CBA-N2a)
3.5. Age Estimation
3.6. Modeling Fish Growth
3.7. Covariates of Ciguatoxicity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Friedman, M.A.; Fernandez, M.; Backer, L.C.; Dickey, R.W.; Bernstein, J.; Schrank, K.; Kibler, S.; Stephan, W.; Gribble, M.O.; Bienfang, P.; et al. An updated review of ciguatera fish poisoning: Clinical, epidemiological, environmental, and public health management. Mar. Drugs 2017, 15, 72. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Arellano, J.L.; Luzardo, O.P.; Perez Brito, A.; Hernandez Cabrera, M.; Zumbado, M.; Carranza, C.; Angel-Moreno, A.; Dickey, R.W.; Boada, L.D. Ciguatera fish poisoning, Canary Islands. Emerg. Infect. Dis. 2005, 11, 1981–1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boada, L.D.; Zumbado, M.; Luzardo, O.P.; Almeida-Gonzalez, M.; Plakas, S.M.; Granade, H.R.; Abraham, A.; Jester, E.L.E.; Dickey, R.W. Ciguatera fish poisoning on the West Africa coast: An emerging risk in the Canary Islands (Spain). Toxicon 2010, 56, 1516–1519. [Google Scholar] [CrossRef] [PubMed]
- Nunez, D.; Matute, P.; Garcia, A.; Garcia, P.; Abadía, N. Outbreak of ciguatera food poisoning by consumption of amberjack (Seriola spp.) in the Canary Islands, May 2012. Eurosurveillance 2012, 17, 20188. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=20188 (accessed on 27 March 2022). [PubMed]
- Bravo, J.; Suarez, F.C.; Ramirez, A.S.; Acosta, F. Ciguatera, an emerging human poisoning in Europe. J. Aquac. Mar. Biol. 2015, 3, 00053. [Google Scholar] [CrossRef]
- Rajeish, M.; Shekar, M.; Madhushree, H.N.; Venugopal, M.N. Presumptive case of ciguatera fish poisoning in Mangalore, India. Curr. Sci. 2016, 111, 1543–1547. [Google Scholar] [CrossRef]
- Rajisha, R.; Kishore, P.; Panda, S.K.; Ravishankar, C.N.; Kumar, K.A. Confirmation of ciguatoxin fish poisoning in red snapper, Lutjanus bohar (forsskål, 1775) by mouse bioassay. Fish. Technol. 2017, 54, 287–290. [Google Scholar]
- Skinner, M.P.; Brewer, T.D.; Johnstone, R.; Fleming, L.E.; Lewis, R.J. Ciguatera fish poisoning in the Pacific Islands (1998 to 2008). PLoS Negl. Trop. Dis. 2011, 5, e1416. [Google Scholar] [CrossRef]
- Chinain, M.; Gatti, C.M.i.; Darius, H.T.; Quod, J.P.; Tester, P.A. Ciguatera poisonings: A global review of occurrences and trends. Harmful Algae 2021, 102, 101873. [Google Scholar] [CrossRef]
- Annual Reports of Ciguatera Poisoning in French Polynesia. Available online: https://www.ciguatera.pf/index.php/fr/la-ciguatera/surveillance-et-statistiques (accessed on 9 December 2021).
- Gatti, C.M.; Lonati, D.; Darius, H.T.; Zancan, A.; Roué, M.; Schicchi, A.; Locatelli, C.A.; Chinain, M. Tectus niloticus (Tegulidae, Gastropod) as a novel vector of ciguatera poisoning: Clinical characterization and follow-up of a mass poisoning event in Nuku Hiva Island (French Polynesia). Toxins 2018, 10, 102. [Google Scholar] [CrossRef] [Green Version]
- Chinain, M.; Gatti, C.M.; Roué, M.; Darius, H.T. Ciguatera-causing dinoflagellates in the genera Gambierdiscus and Fukuyoa: Distribution, ecophysiology and toxicology. In Dinoflagellates: Morphology, Life History and Ecological Significance; Subba Rao, D.V., Ed.; The Nova Science Publishers, Inc.: New York, NY, USA, 2020; pp. 405–457. [Google Scholar]
- Lehane, L.; Lewis, R.J. Ciguatera: Recent advances but the risk remains. Int. J. Food Microbiol. 2000, 61, 91–125. [Google Scholar] [CrossRef]
- Ikehara, T.; Kuniyoshi, K.; Oshiro, N.; Yasumoto, T. Biooxidation of ciguatoxins leads to species-specific toxin profiles. Toxins 2017, 9, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soliño, L.; Costa, P.R. Differential toxin profiles of ciguatoxins in marine organisms: Chemistry, fate and global distribution. Toxicon 2018, 150, 124–143. [Google Scholar] [CrossRef] [PubMed]
- Pasinszki, T.; Lako, J.; Dennis, T.E. Advances in detecting ciguatoxins in fish. Toxins 2020, 12, 494. [Google Scholar] [CrossRef]
- Chinain, M.; Gatti, C.M.; Martin-Yken, H.; Roué, M.; Darius, H.T. Ciguatera poisoning: An increasing burden for Pacific Islands communities in light of climate change? In Climate Change and Marine and Freshwater Toxins, 2nd ed.; Botana, L.M., Louzao, M.C., Vilariño, N., Eds.; De Gruyter: Berlin, Germany; Munich, Germany; Boston, MA, USA, 2021; pp. 369–428. [Google Scholar]
- Mak, Y.L.; Wai, T.-C.; Murphy, M.B.; Chan, W.H.; Wu, J.J.; Lam, J.C.W.; Chan, L.L.; Lam, P.K.S. Pacific ciguatoxins in food web components of coral reef systems in the Republic of Kiribati. Environ. Sci. Technol. 2013, 47, 14070–14079. [Google Scholar] [CrossRef]
- Randall, J.E. A review of ciguatera, tropical fish poisoning, with a tentative explanation of its cause. Bull. Mar. Sci. 1958, 8, 236–267. [Google Scholar]
- Lewis, R.J.; Holmes, M.J. Origin and transfer of toxins involved in ciguatera. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 1993, 106, 615–628. [Google Scholar] [CrossRef]
- Food and Drug Administration (FDA). Appendix 5-FDA and EPA safety levels in regulations and guidance. In Fish and Fishery Products Hazards and Control Guidance, 4th ed.; University of Florida: Gainesville, FL, USA, 2020. Available online: https://www.fda.gov/media/80400/downloadpp.A5-1 (accessed on 11 February 2022).
- EFSA. Scientific opinion on marine biotoxins in shellfish–Emerging toxins: Ciguatoxin group. EFSA J. 2010, 8, 1627. [Google Scholar] [CrossRef]
- Lewis, N.D. Ciguatera-parameters of a tropical health problem. Hum. Ecol. 1984, 12, 253–273. [Google Scholar] [CrossRef]
- Darius, H.T.; Drescher, O.; Ponton, D.; Pawlowiez, R.; Laurent, D.; Dewailly, E.; Chinain, M. Use of folk tests to detect ciguateric fish: A scientific evaluation of their effectiveness in Raivavae Island (Australes, French Polynesia). Food Addit. Contam. Part A 2013, 30, 550–566. [Google Scholar] [CrossRef]
- Arrêté n°125 du 03-11-1939. Liste interdisant la vente de certaines espèces dangereuses de poissons. J. Off. Etabl. Français L’océanie 1939, 1939, 542–543. [Google Scholar]
- Lewis, N.D. Ciguatera-implications for nutrition and marine ressource development in the Pacific islands. Société Océanistes 1983, 39, 89–104. [Google Scholar] [CrossRef]
- Sumner, J.; Turnbull, A.; Dowsett, N. Hazards Affecting Australian Seafood; SARDI: Urrbrae, SA, Australia, 2014; pp. 15–20.
- Sydney Fish Market. Seafood Handling Guidelines, 5th ed.; Locked Bag 247; Sydney Fish Market Pty Ltd.: Pyrmont, NSW, Australia, 2015; pp. 35–36. [Google Scholar]
- Stewart, I.; Lewis, R.J.; Eaglesham, G.K.; Graham, G.C.; Poole, S.; Craig, S.B. Emerging tropical diseases in Australia. Part 2. Ciguatera fish poisoning. Ann. Trop. Med. Parasitol. 2010, 104, 557–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clua, E.; Brena, P.F.; Lecasble, C.; Ghnassia, R.; Chauvet, C. Prevalence and proposal for cost-effective management of the ciguatera risk in the Noumea fish market, New Caledonia (South Pacific). Toxicon 2011, 58, 591–601. [Google Scholar] [CrossRef]
- Chan, T.Y.K. Ciguatoxic potential of brown-marbled grouper in relation to fish size and geographical origin. Am. J. Trop. Med. Hyg. 2015, 93, 1117–1121. [Google Scholar] [CrossRef] [Green Version]
- Farrell, H.; Zammit, A.; Manning, J.; Shadbolt, C.; Szabo, L.; Harwood, T.; Mc Nabb, P.; Turahui, J.A.; Van der Berg, D.J. Clinical diagnosis and chemical confirmation of ciguatera fish poisoning in New South Wales, Australia. Commun. Dis. Intell. Q. Rep. 2016, 40, E1–E6. [Google Scholar]
- Kohli, G.S.; Haslauer, K.; Sarowar, C.; Kretzschmar, A.L.; Boulter, M.; Harwood, D.T.; Laczka, O.; Murray, S.A. Qualitative and quantitative assessment of the presence of ciguatoxin, P-CTX-1B, in Spanish mackerel (Scomberomorus commerson) from waters in New South Wales (Australia). Toxicol. Rep. 2017, 4, 328–334. [Google Scholar] [CrossRef]
- Edwards, A.; Zammit, A.; Farrell, H. Four recent ciguatera fish poisoning incidents in New South Wales, Australia linked to imported fish. Commun. Dis. Intell. 2019, 43, 1–9. [Google Scholar] [CrossRef]
- Caillaud, A.; Eixarch, H.; de la Iglesia, P.; Rodriguez, M.; Dominguez, L.; Andree, K.B.; Diogène, J. Towards the standardisation of the neuroblastoma (neuro-2a) cell-based assay for ciguatoxin-like toxicity detection in fish: Application to fish caught in the Canary Islands. Food Addit. Contam. Part A 2012, 29, 1000–1010. [Google Scholar] [CrossRef]
- Costa, P.R.; Estevez, P.; Castro, D.; Soliño, L.; Gouveia, N.; Santos, C.; Rodrigues, S.M.; Leao, J.M.; Gago-Martínez, A. New insights into the occurrence and toxin profile of ciguatoxins in Selvagens Islands (Madeira, Portugal). Toxins 2018, 10, 524. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Henao, J.A.; García-Álvarez, N.; Fernández, A.; Saavedra, P.; Silva Sergent, F.; Padilla, D.; Acosta-Hernández, B.; Martel Suárez, M.; Diogène, J.; Real, F. Predictive score and probability of CTX-like toxicity in fish samples from the official control of ciguatera in the Canary Islands. Sci. Total Environ. 2019, 673, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Viallon, J.; Chinain, M.; Darius, H.T. Revisiting the neuroblastoma cell-based assay (CBA-N2a) for the improved detection of marine toxins active on voltage gated sodium channels (VGSCs). Toxins 2020, 12, 281. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.M.; Choat, J.H. Comparative demography of commercially important parrotfish species from Micronesia. J. Fish Biol. 2014, 84, 383–402. [Google Scholar] [CrossRef] [PubMed]
- Ebisawa, A.; Kanashiro, K.; Ohta, I.; Uehara, M.; Nakamura, H. Changes of group construction accompanying with growth and maturity in blue-barred parrotfish (Scarus ghobban), and influences of the fishing targeting the immature group to the stock. Reg. Stud. Mar. Sci. 2016, 7, 32–42. [Google Scholar] [CrossRef]
- Ebisawa, A.; Ohta, I.; Uehara, M.; Nakamura, H.; Kanashiro, K.; Yasui, R. Life history variables, annual change in sex ratios with age, and total mortality observed on commercial catch on Pacific steephead parrotfish, Chlorurus microrhinos in waters off the Okinawa Island, southwestern Japan. Reg. Stud. Mar. Sci. 2016, 8, 65–76. [Google Scholar] [CrossRef]
- Taylor, B.M.; Rhodes, K.L.; Marshell, A.; McIlwain, J.L. Age-based demographic and reproductive assessment of orangespine Naso lituratus and bluespine Naso unicornis unicornfishes. J. Fish Biol. 2014, 85, 901–916. [Google Scholar] [CrossRef]
- Pardee, C.; Taylor, B.M.; Felise, S.; Ochavillo, D.; Cuetos-Bueno, J. Growth and maturation of three commercially important coral reef species from American Samoa. Fish. Sci. 2020, 86, 985–993. [Google Scholar] [CrossRef]
- Ohta, I.; Akita, Y.; Uehara, M.; Ebisawa, A. Age-based demography and reproductive biology of three Epinephelus groupers, E. polyphekadion, E. tauvina, and E. howlandi (Serranidae), inhabiting coral reefs in Okinawa. Environ. Biol. Fishes 2017, 100, 1451–1467. [Google Scholar] [CrossRef]
- Rhodes, K.L.; Baremore, I.E.; Taylor, B.M.; Cuetos-Bueno, J.; Hernandez, D. Aligning fisheries management with life history in two commercially important groupers in Chuuk, Federated States of Micronesia. Aquatic Conserv. Mar. Freshw. Ecosyst. 2021, 31, 605–619. [Google Scholar] [CrossRef]
- Taylor, B.M.; Trip, E.D.; Choat, J.H. Dynamic demography: Investigations of life-history variation in the parrotfishes. In Biology of Parrotfishes; Hoey, A.S., Bonaldo, R.M., Eds.; CRC Press; Taylor & Francis Group: Boca Raton, FL, USA; London, UK, 2018; pp. 69–98. [Google Scholar]
- Gaboriau, M.; Ponton, D.; Darius, H.T.; Chinain, M. Ciguatera fish toxicity in French Polynesia: Size does not always matter. Toxicon 2014, 84, 41–50. [Google Scholar] [CrossRef]
- Montojo, U.M.; Tanyag, B.E.; Perelonia, K.B.S.; Cambia, F.D.; Oshiro, N. Ciguatera in the Philippines: Examining reef fish vectors and its causative benthic dinoflagellates in Visayan and Sibuyan seas. Philipp. J. Fish. 2020, 27, 19–29. [Google Scholar] [CrossRef]
- Bienfang, P.; DeFelice, S.; Laws, E.; Wallsgrove, N.; Caldwell, P. Ciguatoxicity in the main Hawaiian Islands: Spatial and temporal variability in the introduced reef carnivore Cephalopholis argus. J. Res. Environ. Sci. Toxicol. 2012, 1, 47–57. [Google Scholar]
- Chinain, M.; Gatti, C.M.i.; Ung, A.; Cruchet, P.; Revel, T.; Viallon, J.; Sibat, M.; Varney, P.; Laurent, V.; Hess, P.; et al. Evidence for the range expansion of ciguatera in French Polynesia: A revisit of the 2009 mass-poisoning outbreak in Rapa Island (Australes Archipelago). Toxins 2020, 12, 759. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, C.R.; Abraham, A.; Stopa, J.E.; Flores Quintana, H.A.; Jester, E.L.E.; La Pinta, J.; Deeds, J.; Benner, R.A.; Adolf, J. Ciguatoxin in Hawai’i: Fisheries forecasting using geospatial and environmental analyses for the invasive Cephalopholis argus (Epinephelidae). Environ. Res. 2022, 207, 112164. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, C.R.; Robertson, A.; Flores Quintana, H.A.; Silander, M.C.; Smith, T.B.; Olsen, D. Ciguatoxin prevalence in 4 commercial fish species along an oceanic exposure gradient in the US Virgin Islands. Environ. Toxicol. Chem. 2018, 37, 1852–1863. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Henao, A.; García-Álvarez, N.; Silva Sergent, F.; Estévez, P.; Gago-Martínez, A.; Martín, F.; Ramos-Sosa, M.; Fernández, A.; Diogène, J.; Real, F. Presence of CTXs in moray eels and dusky groupers in the marine environment of the Canary Islands. Aquat. Toxicol. 2020, 221, 105427. [Google Scholar] [CrossRef]
- Vernoux, J.P.; Lejeune, J. Ciguatera in the French West Indies. Mem. Qld. Mus. Brisb. 1994, 34, 631–638. [Google Scholar]
- O’Toole, A.C.; Dechraoui Bottein, M.-Y.; Danylchuk, A.J.; Ramsdell, J.S.; Cooke, S.J. Linking ciguatera poisoning to spatial ecology of fish: A novel approach to examining the distribution of biotoxin levels in the great barracuda by combining non-lethal blood sampling and biotelemetry. Sci. Total Environ. 2012, 427–428, 98–105. [Google Scholar] [CrossRef] [Green Version]
- Pottier, I.; Vernoux, J.P.; Jones, A.; Lewis, R.J. Characterisation of multiple Caribbean ciguatoxins and congeners in individual specimens of horse-eye jack (Caranx latus) by high-performance liquid chromatography/mass spectrometry. Toxicon 2002, 40, 929–939. [Google Scholar] [CrossRef]
- Soliño, L.; Widgy, S.; Pautonnier, A.; Turquet, J.; Loeffler, C.R.; Flores Quintana, H.A.; Diogène, J. Prevalence of ciguatoxins in lionfish (Pterois spp.) from Guadeloupe, Saint Martin, and Saint Barthélmy Islands (Caribbean). Toxicon 2015, 102, 62–68. [Google Scholar] [CrossRef]
- Oshiro, N.; Nagasawa, H.; Watanabe, M.; Nishimura, M.; Kuniyoshi, K.; Kobayashi, N.; Sugita-Konishi, Y.; Asakura, H.; Tachihara, K.; Yasumoto, T. An extensive survey of ciguatoxins on grouper Variola louti from the Ryukyu Islands, Japan, using liquid chromatography-tandem mass spectrometry (LC-MS/MS). J. Mar. Sci. Eng. 2022, 10, 423. [Google Scholar] [CrossRef]
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-PLUS; Springer: New York, NY, USA, 2000; p. 528. [Google Scholar] [CrossRef]
- DeMartini, E.E.; Andrews, A.H.; Howard, K.G.; Taylor, B.M.; Lou, D.-C.; Donovan, M.K. Comparative growth, age at maturity and sex change, and longevity of Hawaiian parrotfishes, with bomb radiocarbon validation. Can. J. Fish. Aquat. Sci. 2018, 75, 580–589. [Google Scholar] [CrossRef] [Green Version]
- Parsons, M.L.; Aligizaki, K.; Bottein, M.-Y.D.; Fraga, S.; Morton, S.L.; Penna, A.; Rhodes, L. Gambierdiscus and Ostreopsis: Reassessment of the state of knowledge of their taxonomy, geography, ecophysiology, and toxicology. Harmful Algae 2012, 14, 107–129. [Google Scholar] [CrossRef]
- Darius, H.T.; Roué, M.; Ung, A.; Cruchet, P.; Revel, T.; Viallon, J.; Suhas, E.; Gatti, C.M.; Chinain, M. Ecotoxicological characterization of 3 different lagoons of French Polynesia: Tikehau, Kaukura and Mangareva. In Proceedings of the 16th International Conference on Harmful Algae, Wellington, New Zealand, 27–31 October 2014; p. 207. [Google Scholar]
- Chinain, M.; Darius, H.T.; Gatti, C.M.; Roué, M. Update on ciguatera research in French Polynesia. SPC Fish. 2016, 150, 42–51. [Google Scholar]
- Longo, S.; Sibat, M.; Viallon, J.; Darius, H.T.; Hess, P.; Chinain, M. Intraspecific variability in the toxin production and toxin profiles of in vitro cultures of Gambierdiscus polynesiensis (Dinophyceae) from French Polynesia. Toxins 2019, 11, 735. [Google Scholar] [CrossRef] [Green Version]
- Longo, S.; Sibat, M.; Darius, H.T.; Hess, P.; Chinain, M. Effects of pH and nutrients (nitrogen) on growth and toxin profile of the ciguatera-causing dinoflagellate Gambierdiscus polynesiensis (Dinophyceae). Toxins 2020, 12, 767. [Google Scholar] [CrossRef]
- Chinain, M.; Faust, M.; Pauillac, S. Morphology and molecular analyses of three toxic species of Gambierdiscus (Dinophyceae): G. pacificus, sp. nov., G. australes, sp. nov., and G. polynesiensis, sp. nov. J. Phycol. 1999, 35, 1282–1296. [Google Scholar] [CrossRef]
- Litaker, R.W.; Vandersea, M.W.; Faust, M.A.; Kibler, S.R.; Nau, A.W.; Holland, W.C.; Chinain, M.; Holmes, M.J.; Tester, P.A. Global distribution of ciguatera causing dinoflagellates in the genus Gambierdiscus. Toxicon 2010, 56, 711–730. [Google Scholar] [CrossRef]
- Lyu, Y.; Richlen, M.L.; Sehein, T.R.; Chinain, M.; Adachi, M.; Nishimura, T.; Xu, Y.; Parsons, M.L.; Smith, T.B.; Zheng, T.; et al. LSU rDNA based RFLP assays for the routine identification of Gambierdiscus species. Harmful Algae 2017, 66, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Yogi, K.; Sakugawa, S.; Oshiro, N.; Ikehara, T.; Sugiyama, K.; Yasumoto, T. Determination of toxins involved in ciguatera fish poisoning in the Pacific by LC/MS. J. AOAC Internat. 2014, 97, 398–402. [Google Scholar] [CrossRef]
- Bienfang, P.K.; Trapido-Rosenthal, H.; Laws, E.A. Bioaccumulation/biomagnifications in food chains. In Environmental Toxicology: Selected Entries from the Encyclopedia of Sustainability Science and Technology; Laws, E.A., Ed.; Springer Science: New York, NY, USA, 2013; pp. 35–69. [Google Scholar] [CrossRef]
- Chinain, M.; Darius, H.T.; Ung, A.; Fouc, M.T.; Revel, T.; Cruchet, P.; Pauillac, S.; Laurent, D. Ciguatera risk management in French Polynesia: The case study of Raivavae Island (Australes Archipelago). Toxicon 2010, 56, 674–690. [Google Scholar] [CrossRef] [PubMed]
- Rongo, T.; van Woesik, R. Ciguatera poisoning in Rarotonga, Southern Cook Islands. Harmful Algae 2011, 10, 345–355. [Google Scholar] [CrossRef]
- Rongo, T.; van Woesik, R. Socioeconomic consequences of ciguatera poisoning in Rarotonga, Southern Cook Islands. Harmful Algae 2012, 20, 92–100. [Google Scholar] [CrossRef]
- Darius, H.T.; Ponton, D.; Revel, T.; Cruchet, P.; Ung, A.; Tchou Fouc, M.; Chinain, M. Ciguatera risk assessment in two toxic sites of French Polynesia using the receptor-binding assay. Toxicon 2007, 50, 612–626. [Google Scholar] [CrossRef]
- Pawlowiez, R.; Darius, H.T.; Cruchet, P.; Rossi, F.; Caillaud, A.; Laurent, D.; Chinain, M. Evaluation of seafood toxicity in the Australes Archipelago (French Polynesia) using the neuroblastoma cell-based assay. Food Add. Contam. Part A 2013, 30, 567–586. [Google Scholar] [CrossRef]
- Banner, A.H.; Helfrich, P.; Piyakarnchana, T. Retention of ciguatera toxin by the red snapper, Lutjanus bohar. Copeia 1966, 2, 297–301. [Google Scholar] [CrossRef]
- Lewis, R.J.; Sellin, M.; Street, R.; Holmes, M.J.; Gillespie, N.C. Excretion of ciguatoxin from moray eels (Muraenidae) of the central Pacific. In Proceedings of Third International Conference on Ciguatera Fish Poisoning; Polysciences: Laval, QC, Canada, 1992; pp. 131–143. [Google Scholar]
- Ledreux, A.; Brand, H.; Chinain, M.; Dechraoui-Bottein, M.-Y.; Ramsdell, J.S. Dynamics of ciguatoxins from Gambierdiscus polynesiensis in the benthic herbivore Mugil cephalus: Trophic transfer implications. Harmful Algae 2014, 39, 165–172. [Google Scholar] [CrossRef]
- Clausing, R.J.; Losen, B.; Oberhaensli, F.R.; Darius, H.T.; Sibat, M.; Hess, P.; Swarzenski, P.W.; Chinain, M.; Dechraoui Bottein, M.-Y. Experimental evidence of dietary ciguatoxin accumulation in an herbivorous coral reef fish. Aquat. Toxicol. 2018, 200, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Mak, Y.L.; Chang, Y.-H.; Xiao, C.; Chen, Y.-M.; Shen, J.; Wang, Q.; Ruan, Y.; Lam, P.K.S. Uptake and depuration kinetics of Pacific ciguatoxins in orange-spotted grouper (Epinephelus coioides). Environ. Sci. Technol. 2020, 54, 4475–4483. [Google Scholar] [CrossRef]
- Bennett, C.T.; Robertson, A. Depuration kinetics and growth dilution of Caribbean ciguatoxin in the omnivore Lagodon rhomboides: Implications for trophic transfer and ciguatera risk. Toxins 2021, 13, 774. [Google Scholar] [CrossRef]
- Holmes, M.J.; Venables, B.; Lewis, R.J. Critical review and conceptual and quantitative models for the transfer and depuration of ciguatoxins in fishes. Toxins 2021, 13, 515. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Luo, Q.; Liang, Y.; Mazumder, A. Processes and pathways of ciguatoxin in aquatic foodwebs and fish poisoning of seafood consumers. Environ. Rev. 2016, 24, 144–150. [Google Scholar] [CrossRef] [Green Version]
- Nadon, M.O.; Ault, J.S.; Williams, I.D.; Smith, S.G.; DiNardo, G.T. Length-based assessment of coral reef fish populations in the main and northwestern Hawaiian islands. PLoS ONE 2015, 10, e0133960. [Google Scholar] [CrossRef] [PubMed]
- Vernoux, J.P.; Lahlou, N.; Abbad el Andaloussi, S.; Riyeche, N.; Magras, L.P. A study of the distribution of ciguatoxin in individual Caribbean fish. Acta Trop 1985, 42, 225–233. [Google Scholar]
- Oshiro, N.; Nagasawa, H.; Kuniyoshi, K.; Kobayashi, N.; Sugita-Konishi, Y.; Asakura, H.; Yasumoto, T. Characteristic distribution of ciguatoxins in the edible parts of a grouper, Variola louti. Toxins 2021, 13, 218. [Google Scholar] [CrossRef]
- Hardison, D.R.; Holland, W.C.; McCall, J.R.; Bourdelais, A.J.; Baden, D.G.; Darius, H.T.; Chinain, M.; Tester, P.A.; Shea, D.; Flores Quintana, H.A.; et al. Fluorescent receptor binding assay for detecting ciguatoxins in fish. PLoS ONE 2016, 11, e0153348. [Google Scholar] [CrossRef]
- Sibat, M.; Herrenknecht, C.; Darius, H.T.; Roué, M.; Chinain, M.; Hess, P. Detection of Pacific ciguatoxins using liquid chromatography coupled to either low or high resolution mass spectrometry (LC-MS/MS). J. Chromatogr. A 2018, 1571, 16–28. [Google Scholar] [CrossRef]
- Periou, C.; Natier, S.; Damon, N. L’archipel des Gambier; Etudes Thématiques; Institut d’Emision d’Outre-Mer: Papeete, Tahiti, Polynésie Française, 2020; No. 298; p. 4. [Google Scholar]
- Bagnis, R.; Bennet, J.; Barsinas, M.; Chebret, M.; Jacquet, G.; Lechat, I.; Mitermite, Y.; Perolat, P.H.; Rongeras, S. Epidemiology of ciguatera in French Polynesia from 1960 to 1984. In Proceedings of the Fifth International Coral Reef Congress, Tahiti, French Polynesia, 27 May–1 June 1985; pp. 475–482. [Google Scholar]
- Chateau-Degat, M.-L.; Chinain, M.; Darius, H.T.; Dewailly, E.; Mallet, H.-P. Epidemiological surveillance of ciguatera in French Polynesia. BEH Thématique 2009, 48, 522–525. [Google Scholar]
- Chinain, M.; Gatti, C.M.; Roué, M.; Darius, H.T. Ciguatera poisoning in French Polynesia: Insights into the novel trends of an ancient disease. New Microbes New Infect. 2019, 31, 100565. [Google Scholar] [CrossRef]
- Bacchet, P.; Zysman, T.; Lefèvre, Y. Guide des Poissons de Tahiti et Ses Îles; Au Vent des Iles: Pirae, Tahiti, Polynésie Française, 2006; p. 608. [Google Scholar]
- Green, A.L.; Maypa, A.P.; Almany, G.R.; Rhodes, K.L.; Weeks, R.; Abesamis, R.A.; Gleason, M.G.; Mumby, P.J.; White, A.T. Larval dispersal and movement patterns of coral reef fishes, and implications for marine reserve network design. Biol. Rev. 2015, 90, 1215–1247. [Google Scholar] [CrossRef]
- Darius, H.T.; Roué, M.; Sibat, M.; Viallon, J.; Gatti, C.M.; Vandersea, M.W.; Tester, P.A.; Litaker, R.W.; Amzil, Z.; Hess, P.; et al. Tectus niloticus (Tegulidae, Gastropod) as a novel vector of ciguatera poisoning: Detection of Pacific ciguatoxins in toxic samples from Nuku Hiva Island (French Polynesia). Toxins 2018, 10, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caillaud, A.; de la Iglesia, P.; Darius, H.T.; Pauillac, S.; Aligizaki, K.; Fraga, S.; Chinain, M.; Diogène, J. Update on methodologies available for ciguatoxin determination: Perspectives to confront the onset of ciguatera fish poisoning in europe. Mar. Drugs 2010, 8, 1838–1907. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, C.R.; Bodi, D.; Tartaglione, L.; Dell’Aversano, C.; Preiss-Weigert, A. Improving in vitro ciguatoxin and brevetoxin detection: Selecting neuroblastoma (Neuro-2a) cells with lower sensitivity to ouabain and veratridine (OV-LS). Harmful Algae 2021, 103, 101994. [Google Scholar] [CrossRef] [PubMed]
- Vigliola, L.; Meekan, M.G. Size at hatching and planktonic growth determine post-settlement survivorship of a coral reef fish. Oecologia 2002, 131, 89–93. [Google Scholar] [CrossRef]
- Panfili, J.; de Pontual, H.; Troadec, H.; Wright, P. Manual of Fish Sclerochronology; IFREMER-et IRD Coedition: Brest, France, 2002; p. 464. [Google Scholar]
- Lecomte-Finiger, R. Growth history and age at recruitment of European glass eels (Anguilla anguilla) as revealed by otolith microstructure. Mar. Biol. 1992, 114, 205–210. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Vigliola, L.; Meekan, M.G. The back-calculation of fish growth from otoliths. In Tropical Fish Otoliths: Information for Assessment, Management and Ecology. Reviews: Methods and Technologies in Fish Biology and Fisheries; Green, B.S., Mapstone, B.D., Carlos, G., Begg, G.A., Eds.; Springer: Dordrecht, The Netherland, 2009; Volume 11, pp. 174–211. [Google Scholar]
- Lea, E. On the methods used in the herring investigations. Publ. Circ. Cons. Perm. Int. Explor. Mer. 1910, 53, 1–175. [Google Scholar]
- Enberg, K.; Dunlop, E.S.; Jørgensen, C. Fish Growth. In Encyclopedia of Ecology; Jørgensen, S.E., Fath, B.D., Eds.; Academic Press: Oxford, UK, 2008; pp. 1564–1572. [Google Scholar] [CrossRef]
- Mercier, L.; Panfili, J.; Paillon, C.; N’Diaye, A.; Mouillot, D.; Darnaude, A.M. Otolith reading and multi-model inference for improved estimation of age and growth in the gilthead seabream Sparus aurata (L.). Estuar. Coast Shelf. Sci. 2011, 92, 534–545. [Google Scholar] [CrossRef]
- Pinheiro, J.C. Model Building Using Covariates in Nonlinear Mixed-Effects Models. J. Soc. Fr. Stat. 2002, 143, 79–101. Available online: http://www.numdam.org/item?id=JSFS_2002__143_1-2_79_0 (accessed on 27 March 2022).
- R Core Team. R: A Language and Environment for Statistical Computing, Version 3.5. 2.; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. NLME: Linear and Nonlinear Mixed Effects Models, R Package Version 3.1-108; 2013. Available online: https://CRAN.R-project.org/package=nlme (accessed on 11 February 2022).
- FAO; WHO. Report of the Expert Meeting on Ciguatera Poisoning: Rome, 19–23 November 2018; Food Safety and Quality No. 9; World Health Organization: Rome, Italy, 2020; pp. 1–156. [Google Scholar] [CrossRef]
- Oshiro, N.; Tomikawa, T.; Kuniyoshi, K.; Ishikawa, A.; Toyofuku, H.; Kojima, T.; Asakura, H. LC–MS/MS analysis of ciguatoxins revealing the regional and species distinction of fish in the tropical Western Pacific. J. Mar. Sci. Eng. 2021, 9, 299. [Google Scholar] [CrossRef]
- Hardison, D.R.; Holland, W.C.; Darius, H.T.; Chinain, M.; Tester, P.A.; Shea, D.; Bogdanoff, A.K.; Morris, J.A., Jr.; Flores Quintana, H.A.; Loeffler, C.R.; et al. Investigation of ciguatoxins in invasive lionfish from the greater Caribbean region: Implications for fishery development. PLoS ONE 2018, 13, e0198358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diogène, J.; Rambla, M.; Campàs, M.; Fernández, M.; Andree, K.; Tudó, A.; Rey, M.; Sagristà, N.; Aguayo, P.; Leonardo, S.; et al. Evaluation of ciguatoxins in seafood and the environment in Europe. EFSA Supporting Publ. 2021, 18, 6648E. [Google Scholar] [CrossRef]
- Litaker, R.W.; Holland, W.C.; Hardison, D.R.; Pisapia, F.; Hess, P.; Kibler, S.R.; Tester, P.A. Ciguatoxicity of Gambierdiscus and Fukuyoa species from the Caribbean and Gulf of Mexico. PLoS ONE 2017, 12, e0185776. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Legrand, A.M.; Ishibashi, Y.; Fukui, M.; Yasumoto, T. Structures and configurations of ciguatoxin from the moray eel Gymnothorax javanicus and its likely precursor from the dinoflagellate Gambierdiscus toxicus. J. Am. Chem. Soc. 1990, 112, 4380–4386. [Google Scholar] [CrossRef]
- Legrand, A.M.; Fukui, M.; Cruchet, P.; Ishibashi, Y.; Yasumoto, T. Characterization of ciguatoxins from different fish species and wild Gambierdiscus toxicus. In Proceedings of Third International Conference on Ciguatera Fish Poisoning; Polysciences: Laval, QC, Canada, 1992; pp. 25–32. [Google Scholar]
- Satake, M.; Murata, M.; Yasumoto, T. The structure of CTX3C, a ciguatoxin congener isolated from cultured Gambierdiscus toxicus. Tetrahedron Lett. 1993, 34, 1975–1978. [Google Scholar] [CrossRef]
- Satake, M.; Ishibashi, Y.; Legrand, A.-M.; Yasumoto, T. Isolation and structure of ciguatoxin-4A, a new ciguatoxin precursor, from cultures of dinoflagellate Gambierdiscus toxicus and parrotfish Scarus gibbus. Biosci. Biotech. Biochem. 1997, 60, 2103–2105. [Google Scholar] [CrossRef] [Green Version]
- Satake, M.; Morohashi, A.; Oguri, H.; Oishi, T.; Hirama, M.; Harada, N.; Yasumoto, T. The absolute configuration of ciguatoxin. J. Am. Chem. Soc. 1997, 119, 11325–11326. [Google Scholar] [CrossRef]
- Yasumoto, T.; Igarashi, T.; Legrand, A.-M.; Cruchet, P.; Chinain, M.; Fujita, T.; Naoki, H. Structural elucidation of ciguatoxin congeners by fast-atom bombardment tandem mass spectroscopy. J. Am. Chem. Soc. 2000, 122, 4988–4989. [Google Scholar] [CrossRef]
- Chinain, M.; Darius, H.T.; Ung, A.; Cruchet, P.; Wang, Z.; Ponton, D.; Laurent, D.; Pauillac, S. Growth and toxin production in the ciguatera-causing dinoflagellate Gambierdiscus polynesiensis (Dinophyceae) in culture. Toxicon 2010, 56, 739–750. [Google Scholar] [CrossRef]
- Yogi, K.; Oshiro, N.; Inafuku, Y.; Hirama, M.; Yasumoto, T. Detailed LC-MS/MS analysis of ciguatoxins revealing distinct regional and species characteristics in fish and causative alga from the Pacific. Anal. Chem. 2011, 83, 8886–8891. [Google Scholar] [CrossRef]
- Rhodes, L.; Harwood, T.; Smith, K.; Argyle, P.; Munday, R. Production of ciguatoxin and maitotoxin by strains of Gambierdiscus australes, G. pacificus and G. polynesiensis (Dinophyceae) isolated from Rarotonga, Cook Islands. Harmful Algae 2014, 39, 185–190. [Google Scholar] [CrossRef]
- Roué, M.; Darius, H.T.; Picot, S.; Ung, A.; Viallon, J.; Gaertner-Mazouni, N.; Sibat, M.; Amzil, Z.; Chinain, M. Evidence of the bioaccumulation of ciguatoxins in giant clams (Tridacna maxima) exposed to Gambierdiscus spp. cells. Harmful Algae 2016, 57, 78–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, K.F.; Rhodes, L.; Verma, A.; Curley, B.G.; Harwood, D.T.; Kohli, G.S.; Solomona, D.; Rongo, T.; Munday, R.; Murray, S.A. A new Gambierdiscus species (Dinophyceae) from Rarotonga, Cook Islands: Gambierdiscus cheloniae sp. nov. Harmful Algae 2016, 60, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Munday, R.; Murray, S.; Rhodes, L.; Larsson, M.; Harwood, D. Ciguatoxins and maitotoxins in extracts of sixteen Gambierdiscus isolates and one Fukuyoa isolate from the South Pacific and their toxicity to mice by intraperitoneal and oral administration. Mar. Drugs 2017, 15, 208. [Google Scholar] [CrossRef] [Green Version]
- Roué, M.; Darius, H.T.; Ung, A.; Viallon, J.; Sibat, M.; Hess, P.; Amzil, Z.; Chinain, M. Tissue distribution and elimination of ciguatoxins in Tridacna maxima (Tridacnidae, Bivalvia) fed Gambierdiscus polynesiensis. Toxins 2018, 10, 189. [Google Scholar] [CrossRef] [Green Version]
- Murray, J.S.; Boundy, M.J.; Selwood, A.I.; Harwood, D.T. Development of an LC–MS/MS method to simultaneously monitor maitotoxins and selected ciguatoxins in algal cultures and P-CTX-1B in fish. Harmful Algae 2018, 80, 80–87. [Google Scholar] [CrossRef]
- Legrand, A.M.; Litaudon, M.; Genthon, J.N.; Bagnis, R.; Yasumoto, T. Isolation and some properties of ciguatoxin. J. App. Phycol. 1989, 1, 183–188. [Google Scholar] [CrossRef]
- Murata, M.; Legrand, A.M.; Yasumoto, T. A probable partial structure of ciguatoxin isolated from the moray eel Gymnothorax javanicus. Tetrahedron Lett. 1989, 30, 3793–3796. [Google Scholar] [CrossRef]
- Lewis, R.J.; Sellin, M.; Poli, M.A.; Norton, R.S.; MacLeod, J.K.; Sheil, M.M. Purification and characterization of ciguatoxins from moray eel (Lycodontis javanicus, Muraenidae). Toxicon 1991, 29, 1115–1127. [Google Scholar] [CrossRef]
- Satake, M.; Fukui, M.; Legrand, A.-M.; Cruchet, P.; Yasumoto, T. Isolation and structures of new ciguatoxin analogs, 2,3-dihydroxyCTX3C and 51-hydroxyCTX3C, accumulated in tropical reef fish. Tetrahedron Lett. 1998, 39, 1197–1198. [Google Scholar] [CrossRef]
- Lewis, R.J.; Yang, A.; Jones, A. Rapid extraction combined with LC-tandem mass spectrometry (CREM-LC/MS/MS) for the determination of ciguatoxins in ciguateric fish flesh. Toxicon 2009, 54, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, B.; Whittle, N.; Shaw, G.; Eaglesham, G.; Moore, M.R.; Lewis, R.J. Human fatality associated with Pacific ciguatoxin contaminated fish. Toxicon 2010, 56, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Stewart, I.; Eaglesham, G.K.; Poole, S.; Graham, G.; Paulo, C.; Wickramasinghe, W.; Sadler, R.; Shaw, G.R. Establishing a public health analytical service based on chemical methods for detecting and quantifying Pacific ciguatoxin in fish samples. Toxicon 2010, 56, 804–812. [Google Scholar] [CrossRef]
- Otero, P.; Pérez, S.; Alfonso, A.; Vale, C.; Rodriguez, P.; Gouveia, N.N.; Gouveia, N.; Delgado, J.; Vale, P.; Hirama, M.; et al. First toxin profile of ciguateric fish in Madeira Arquipelago (Europe). Anal. Chem. 2010, 82, 6032–6039. [Google Scholar] [CrossRef] [PubMed]
- Oshiro, N.; Yogi, K.; Asato, S.; Sasaki, T.; Tamanaha, K.; Hirama, M.; Yasumoto, T.; Inafuku, Y. Ciguatera incidence and fish toxicity in Okinawa, Japan. Toxicon 2010, 56, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.J.; Mak, Y.L.; Murphy, M.B.; Lam, J.C.W.; Chan, W.C.; Wang, M.; Chan, L.L.; Lam, P.K.S. Validation of an accelerated solvent extraction liquid chromatography tandem mass spectrometry method for Pacific ciguatoxin-1 in fish flesh and comparison with the mouse neuroblastoma assay. Anal. Bioanal. Chem. 2011, 400, 3165–3175. [Google Scholar] [CrossRef] [PubMed]
- Yogi, K.; Oshiro, N.; Matsuda, S.; Sakugawa, S.; Matsuo, K.; Yasumoto, T. Toxin profiles in fish implicated in Ciguatera Fish Poisoning in Amami and Kakeroma Islands, Kagoshima Prefecture, Japan. Shokuhin Eiseigaku Zasshi. J. Food Hyg. Soc. Jpn. 2013, 54, 385–391. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.-K.; Hung, P.; Lo, J.Y.C. Ciguatera fish poisoning in Hong Kong -A 10-year perspective on the class of ciguatoxins. Toxicon 2014, 86, 96–106. [Google Scholar] [CrossRef]
- Farrell, H.; Murray, S.A.; Zammit, A.; Edwards, A.W. Management of ciguatoxin risk in Eastern Australia. Toxins 2017, 9, 367. [Google Scholar] [CrossRef] [Green Version]
- Ha, D.; Uesugi, A.; Uchida, H.; Ky, P.; Minh, D.; Watanabe, R.; Matsushima, R.; Oikawa, H.; Nagai, S.; Iwataki, M.; et al. Identification of causative ciguatoxins in red snappers Lutjanus bohar implicated in Ciguatera Fish Poisonings in Vietnam. Toxins 2018, 10, 420. [Google Scholar] [CrossRef] [Green Version]
- Leonardo, S.; Gaiani, G.; Tsumuraya, T.; Hirama, M.; Turquet, J.; Sagristà, N.; Rambla-Alegre, M.; Flores, C.; Caixach, J.; Diogène, J.; et al. Addressing the analytical challenges for the detection of ciguatoxins using an electrochemical biosensor. Anal. Chem. 2020, 92, 4858–4865. [Google Scholar] [CrossRef] [PubMed]
- Nagae, M.; Igarashi, T.; Mizukoshi, K.; Yasumoto, T.; Kuniyoshi, K.; Oshiro, N. Development and validation of an LC-MS/MS method for the ultra-trace analysis of Pacific ciguatoxins in fish. J. AOAC Internat. 2021, 104, 1272–1281. [Google Scholar] [CrossRef] [PubMed]
- Darius, H.T.; Revel, T.; Cruchet, P.; Viallon, J.; Gatti, C.M.i.; Sibat, M.; Hess, P.; Chinain, M. Deep-water fish are potential vectors of ciguatera poisoning in the Gambier Islands, French Polynesia. Marine Drugs 2021, 19, 644. [Google Scholar] [CrossRef] [PubMed]
- Spielmeyer, A.; Loeffler, C.R.; Bodi, D. Extraction and LC-MS/MS Analysis of Ciguatoxins: A Semi-Targeted Approach Designed for Fish of Unknown Origin. Toxins 2021, 13, 630. [Google Scholar] [CrossRef] [PubMed]
- Dao, H.V.; Uesugi, A.; Uchida, H.; Watanabe, R.; Matsushima, R.; Lim, Z.F.; Jipanin, S.J.; Pham, K.X.; Phan, M.-T.; Leaw, C.P.; et al. Identification of fish species and toxins implicated in a snapper food poisoning event in Sabah, Malaysia, 2017. Toxins 2021, 13, 657. [Google Scholar] [CrossRef] [PubMed]
- Vernoux, J.-P.; Lewis, R.J. Isolation and characterisation of Caribbean ciguatoxins from the horse-eye jack (Caranx latus). Toxicon 1997, 35, 889–900. [Google Scholar] [CrossRef]
- Lewis, R.J.; Vernoux, J.-P.; Brereton, I.M. Structure of Caribbean ciguatoxin Isolated from Caranx latus. J. Am. Chem. Soc. 1998, 120, 5914–5920. [Google Scholar] [CrossRef]
- Pottier, I.; Vernoux, J.P.; Jones, A.; Lewis, R.J. Analysis of toxin profiles in three different fish species causing ciguatera fish poisoning in Guadeloupe, French West Indies. Food Add. Contam. Part A 2002, 19, 1034–1042. [Google Scholar] [CrossRef]
- Pottier, I.; Hamilton, B.; Jones, A.; Lewis, R.J.; Vernoux, J.P. Identification of slow and fast-acting toxins in a highly ciguatoxic barracuda (Sphyraena barracuda) by HPLC/MS and radiolabelled ligand binding. Toxicon 2003, 42, 663–672. [Google Scholar] [CrossRef]
- Abraham, A.; Jester, E.L.E.; Granade, H.R.; Plakas, S.M.; Dickey, R.W. Caribbean ciguatoxin profile in raw and cooked fish implicated in ciguatera. Food Chem. 2012, 131, 192–198. [Google Scholar] [CrossRef]
- Hossen, V.; Soliño, L.; Leroy, P.; David, E.; Velge, P.; Dragacci, S.; Krys, S.; Flores Quintana, H.; Diogène, J. Contribution to the risk characterization of ciguatoxins: LOAEL estimated from eight ciguatera fish poisoning events in Guadeloupe (French West Indies). Environ. Res. 2015, 143, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Robertson, A.; Garcia, A.C.; Quintana, H.A.F.; Smith, T.B.; Castillo Ii, B.F.; Reale-Munroe, K.; Gulli, J.A.; Olsen, D.A.; Hooe-Rollman, J.I.; Jester, E.L.E.; et al. Invasive lionfish (Pterois volitans): A potential human health threat for ciguatera fish poisoning in tropical waters. Mar. Drugs 2014, 12, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estevez, P.; Castro, D.; Pequeño-Valtierra, A.; Leao, J.M.; Vilariño, O.; Diogène, J.; Gago-Martínez, A. An attempt to characterize the ciguatoxin profile in Seriola fasciata causing ciguatera fish poisoning in Macaronesia. Toxins 2019, 11, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estevez, P.; Castro, D.; Manuel Leao, J.; Yasumoto, T.; Dickey, R.; Gago-Martinez, A. Implementation of liquid chromatography tandem mass spectrometry for the analysis of ciguatera fish poisoning in contaminated fish samples from Atlantic coasts. Food Chem. 2019, 280, 8–14. [Google Scholar] [CrossRef]
- Loeffler, C.R.; Handy, S.M.; Flores Quintana, H.A.; Deeds, J.R. Fish hybridization leads to uncertainty regarding ciguatera fish poisoning risk; confirmation of hybridization and ciguatoxin accumulation with implications for stakeholders. J. Mar. Sci. Eng. 2019, 7, 105. [Google Scholar] [CrossRef] [Green Version]
- Kryuchkov, F.; Robertson, A.; Miles, C.O.; Mudge, E.M.; Uhlig, S. LC–HRMS and chemical derivatization strategies for the structure elucidation of caribbean ciguatoxins: Identification of C-CTX-3 and -4. Mar. Drugs 2020, 18, 182. [Google Scholar] [CrossRef] [Green Version]
- Canals, A.; Martínez, C.V.; Diogène, J.; Gago-Martínez, A.; Cebadera-Miranda, L.; de Vasconcelos, F.M.; Gómez, I.L.; Sánchez, E.V.M.; Alférez, R.C.; Núñez, D.; et al. Risk characterisation of ciguatera poisoning in Europe. EFSA Support. Publ. 2021, 18, 6647E. [Google Scholar] [CrossRef]
- Costa, P.R.; Estévez, P.; Soliño, L.; Castro, D.; Rodrigues, S.M.; Timoteo, V.; Leao-Martins, J.M.; Santos, C.; Gouveia, N.; Diogène, J.; et al. An update on ciguatoxins and CTX-like toxicity in fish from different trophic levels of the Selvagens Islands (NE Atlantic, Madeira, Portugal). Toxins 2021, 13, 580. [Google Scholar] [CrossRef]
- Hamilton, B.; Hurbungs, M.; Jones, A.; Lewis, R.J. Multiple ciguatoxins present in Indian Ocean reef fish. Toxicon 2002, 40, 1347–1353. [Google Scholar] [CrossRef]
- Hamilton, B.; Hurbungs, M.; Vernoux, J.P.; Jones, A.; Lewis, R.J. Isolation and characterisation of Indian Ocean ciguatoxin. Toxicon 2002, 40, 685–693. [Google Scholar] [CrossRef]
- Darius, H.T.; Roué, M.; Sibat, M.; Viallon, J.; Gatti, C.M.i.; Vandersea, M.W.; Tester, P.A.; Litaker, R.W.; Amzil, Z.; Hess, P.; et al. Toxicological investigations on the sea urchin Tripneustes gratilla (Toxopneustidae, Echinoid) from Anaho Bay (Nuku Hiva, French Polynesia): Evidence for the presence of Pacific ciguatoxins. Mar. Drugs 2018, 16, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diogène, J.; Reverté, L.; Rambla-Alegre, M.; del Rio, V.; de la Iglesia, P.; Campas, M.; Palacios, O.; Flores, C.; Caixach, J.; Ralijaona, C.; et al. Identification of ciguatoxins in a shark involved in a fatal food poisoning in the Indian Ocean. Sci. Rep. 2017, 7, 8240. [Google Scholar] [CrossRef] [PubMed]
- Dierking, J.; Campora, C.E. Ciguatera in the introduced fish Cephalopholis argus (Serranidae) in Hawai‘i and implications for fishery management. Pac. Sci. 2009, 63, 193–204. [Google Scholar] [CrossRef] [Green Version]




| Response | Factor | DF | Sum Sq | Mean Sq | Iter | p-Value |
|---|---|---|---|---|---|---|
| Size | Species | 4 | 3852.3 | 963.08 | 107 | <2 × 10−16 *** |
| Ciguatoxicity | 2 | 234.2 | 117.09 | 107 | 0.01639 * | |
| Species:Ciguatoxicity | 7 | 258.6 | 36.95 | 107 | 0.24569 | |
| Residuals | 206 | 5835.8 | 28.33 | |||
| Weight | Species | 4 | 23,076,135 | 5,769,034 | 107 | <2 × 10−16 *** |
| Ciguatoxicity | 2 | 2,143,875 | 1,071,937 | 107 | 0.01918 * | |
| Species:Ciguatoxicity | 7 | 2,642,863 | 377,552 | 107 | 0.19061 | |
| Residuals | 206 | 54,247,969 | 263,340 | |||
| Age | Species | 4 | 77.17 | 19.2921 | 107 | 0.0406 * |
| Ciguatoxicity | 2 | 28.73 | 14.3655 | 107 | 0.1126 | |
| Species:Ciguatoxicity | 7 | 77.95 | 11.1362 | 107 | 0.1247 | |
| Residuals | 2 | 28.73 | 14.3655 | 0.1126 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Darius, H.T.; Paillon, C.; Mou-Tham, G.; Ung, A.; Cruchet, P.; Revel, T.; Viallon, J.; Vigliola, L.; Ponton, D.; Chinain, M. Evaluating Age and Growth Relationship to Ciguatoxicity in Five Coral Reef Fish Species from French Polynesia. Mar. Drugs 2022, 20, 251. https://doi.org/10.3390/md20040251
Darius HT, Paillon C, Mou-Tham G, Ung A, Cruchet P, Revel T, Viallon J, Vigliola L, Ponton D, Chinain M. Evaluating Age and Growth Relationship to Ciguatoxicity in Five Coral Reef Fish Species from French Polynesia. Marine Drugs. 2022; 20(4):251. https://doi.org/10.3390/md20040251
Chicago/Turabian StyleDarius, Hélène Taiana, Christelle Paillon, Gérard Mou-Tham, André Ung, Philippe Cruchet, Taina Revel, Jérôme Viallon, Laurent Vigliola, Dominique Ponton, and Mireille Chinain. 2022. "Evaluating Age and Growth Relationship to Ciguatoxicity in Five Coral Reef Fish Species from French Polynesia" Marine Drugs 20, no. 4: 251. https://doi.org/10.3390/md20040251