
Current Trends and New Challenges in Marine Phycotoxins

 , , , , ,
, , , , , 
Abstract
:1. Introduction
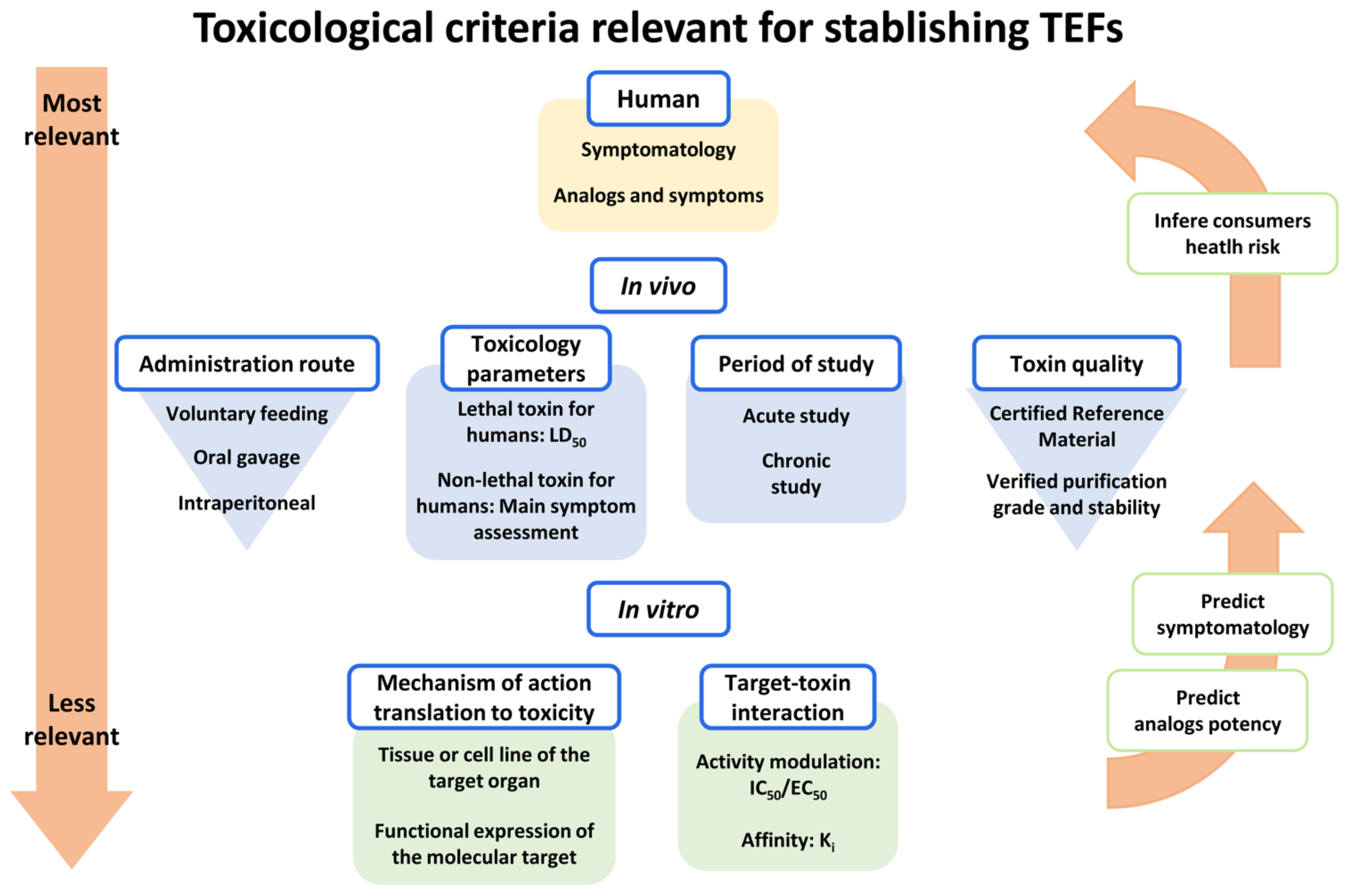
2. Lack of Traceability of Toxicity Values
- Establishment of TEFs based on acute oral toxicity data including analogues with toxic relevance at the levels in which they are present in mollusks;
- Information is needed on genotoxicity, oral toxicity, and toxicity mechanisms for some groups of toxins;
- Information is needed on the combined toxicity of different groups of toxins that are usually present in mollusks.
- Most of the current TEFs are based on the acute effects of toxin intraperitoneal (i.p.) injection to mice, but these values do not reflect the oral absorption, which is the relevant route for the effects of marine phycotoxins on human health;
- The majority of TEFs used today have been estimated using toxins of unknown origin and purity, and therefore generate discrepancies, recognized by EFSA [64]. In fact, the need for use of certified reference materials (CRM) for the different toxin analogues in TEFs determination is highlighted by several extensive reviews on marine toxins TEFs [48,70]. This is still a difficult problem to solve due to the absence of toxin CRMs for many analogues in the previously described toxin groups. In recent years, the commercialization of ISO 17034 certified reference materials for some marine toxins has been guaranteed in Europe through commercial channels (www.cifga.com; accessed on 11 January 2022);
- Current TEFs for marine neurotoxin proposed by EFSA have been obtained using differently purified toxins and quantifying the amount of toxin according to a different criterion in each laboratory, which increases the diversity and disparity of the data collected by EFSA. This fact should be amended at present. In the case of working with purified toxins from mollusk samples, the Standard Operating Procedures (SOPs) of the European marine biotoxin reference laboratory harmonize the extraction of toxins from mollusk samples and the realization of the corresponding analytical or biological determinations.
- First, the absence of correlation between the toxicity obtained by MBA and the acute oral toxicity is highlighted [71]. In general, compounds administered i.p. are absorbed quickly and completely from the peritoneal cavity, while the oral administration can decrease the absorption of many substances, and therefore the i.p. route would provide a much higher toxicity than the real one. The opposite occurs when the toxin is metabolized to a more toxic oral analogue: the MBA would give a lower toxicity, for example, in the case of neosaxitoxin that is more toxic than saxitoxin [72] and other toxins from the group of paralytic toxins [73]. In fact, these two studies with paralytic toxins and other studies with diarrheic toxins [39,40] emphasize the need to review actual TEFs using toxin CRMs and the oral route for toxin administration in order to determine reliable TEFs useful to use analytical methods for neurotoxin monitoring [48].
- In certain cases, TEFs have been established after measuring in vitro toxicity or cellular effects of the toxin [74,75,76]. Although these studies do not take into account neither the absorption nor the metabolism or elimination of the toxins in vivo, these data have also been taken into consideration by EFSA to establish the current TEFs [64] even when there is no approved in vitro model to evaluate the toxicity of paralytic toxins. In fact, the PSP TEFs reported by FAO/WHO are a combination of in vitro effect on human sodium channels with oral toxicity in mice [70].
- Although oral toxicity is the relevant parameter to establish TEFs for marine toxins, special caution is necessary, since, although the Organization for Economic Cooperation and Development (OECD) guidelines for determining acute toxicity OECD 420-Acute Oral Toxicity-Fixed Dose Procedure and OECD 425- Acute Oral Toxicity-Up-and-Down-Procedure establishes the administration of the chemical compound by gastric tube, the semisolid content of the stomach of the rodents can facilitate that the toxin is absorbed quickly in the duodenum instead of mixing with the stomach food. In the case of marine toxins, it seems more convenient to administer the toxins in the food to facilitate toxin ingestion with the food in a short time. In fact, TEFs obtained by forced feeding (gavage–gastric tube) and voluntary consumption of food [73] may show differences.
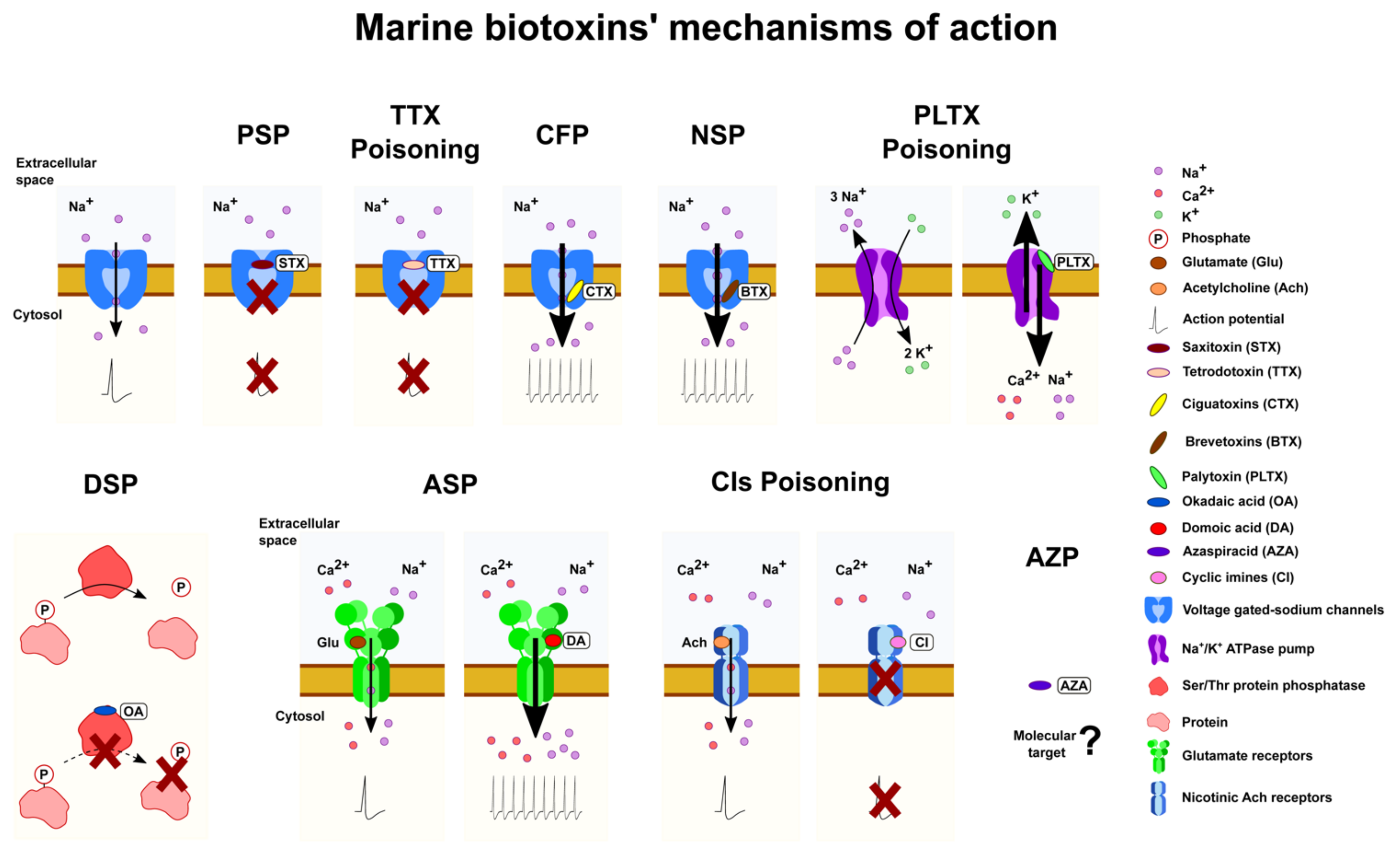
3. Mechanism of Action and Toxicity: The Need for Predefined Toxicological Criteria
3.1. Marine Phycotoxins Acting on Voltage-Gated Sodium Channel

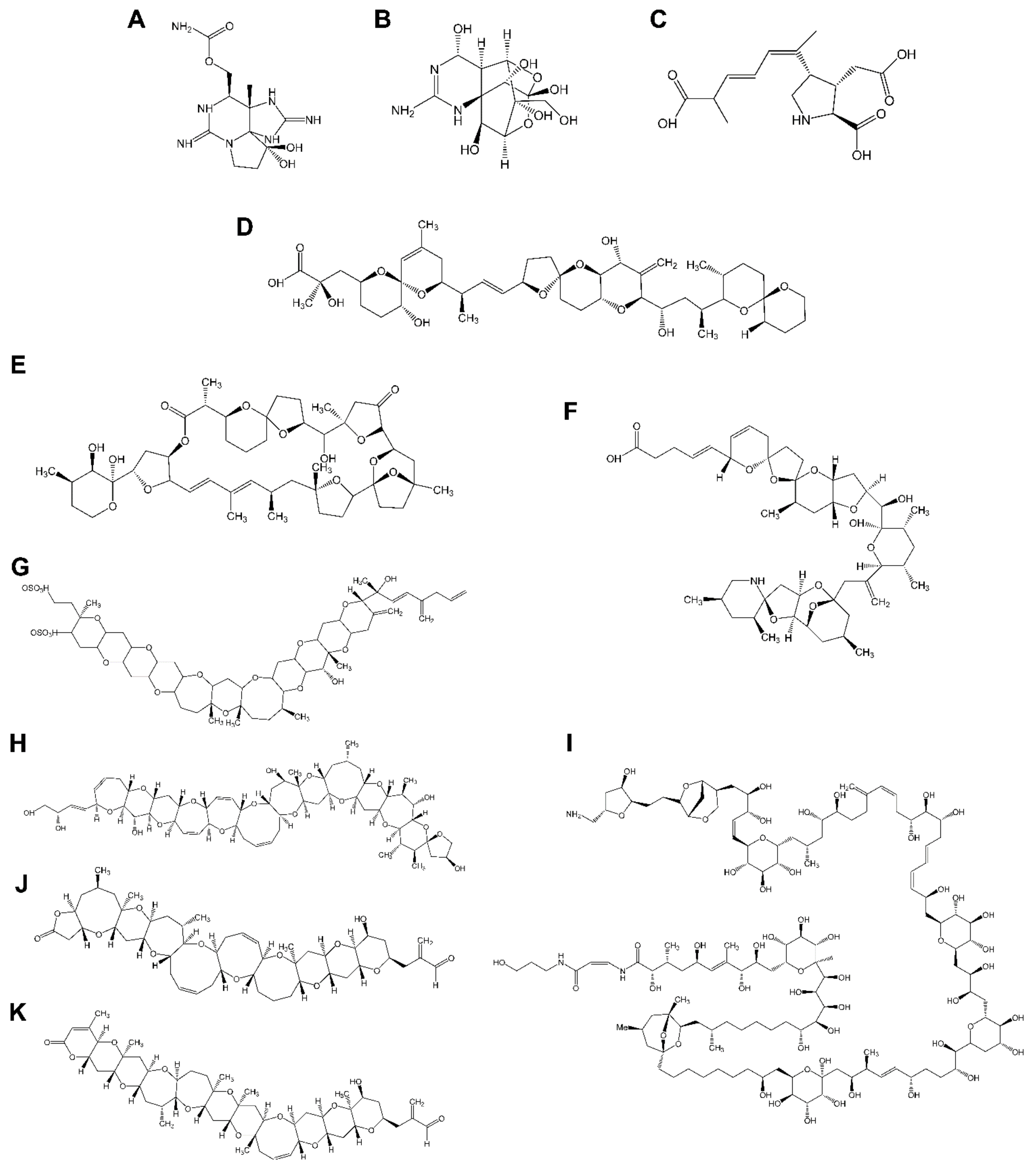{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
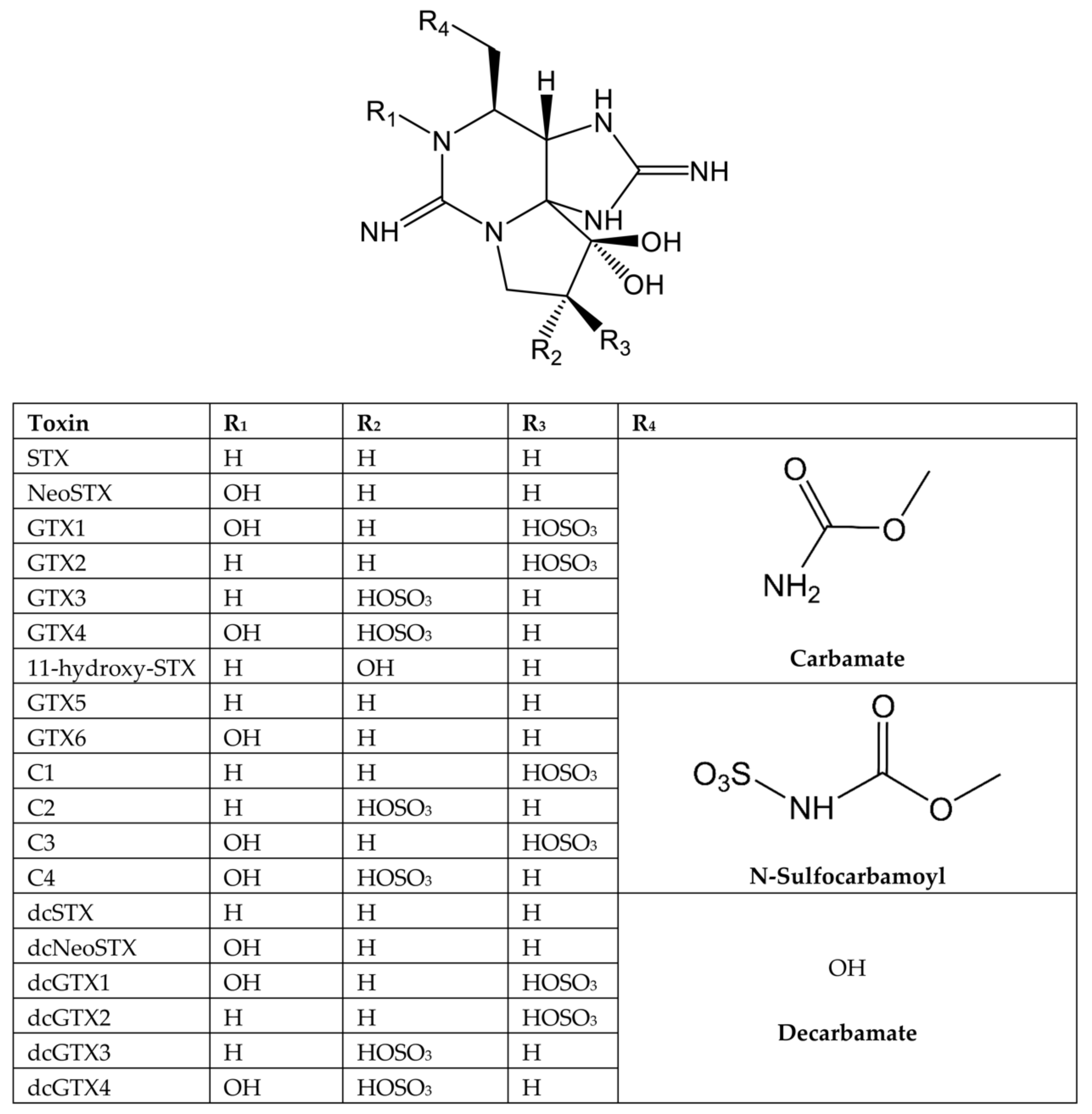
| Toxin | TEF EFSA [5] | TEF FAO [70] | TEFs Based on Oral Gavage | TEFs Based on Voluntary Feeding | TEFs Based on In Vitro IC50 for Each Nav Subunit [74] | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Nav1.1 | Nav1.2 | Nav1.3 | Nav1.4 | Nav1.5 | Nav1.6 | Nav1.7 | |||||
| STX | 1 | 1 | 1 a | 1 a | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| NeoSTX | 1 | 2 | 1.7 a | 2.54 a | 6.4 | 2.63 | 2.85 | 2.6 | 38.6 | 1.2 | 5 |
| GTX1 | 1 | 1 | |||||||||
| GTX2 | 0.4 | 0.4 | |||||||||
| GTX3 | 0.6 | 0.6 | |||||||||
| GTX4 | 0.7 | 0.7 | |||||||||
| GTX5 | 0.1 | 0.1 | 0.063 b | 0.064 b | 0.015 | 0.014 | 0.08 | 0.18 | 10.7 | 0.11 | 4.08 |
| GTX6 | 0.1 | 0.05 | 0.038 b | <0.017 b | |||||||
| GTX1&4 | 0.74 a | 0.93 a | 0.96 | 0.54 | 1.6 | 0.57 | 14.4 | 1.4 | 9.29 | ||
| GTX2&3 | 0.53 a | 0.57 a | 0.2 | 0.39 | 0.64 | 0.32 | 3.87 | 0.15 | 0.27 | ||
| dcSTX | 1 | 0.5 | 0.46 a | 0.37 a | 0.07 | 0.25 | 0.08 | 0.16 | 0.96 | 0.96 | 4.6 |
| dcNeoSTX | 0.4 | 0.2 | 0.22 b | 0.22 b | 0.001 | 0.1 | 0.024 | 0.001 | 0.73 | 0.25 | 0.33 |
| dcGTX2 | 0.2 | 0.2 | |||||||||
| dcGTX3 | 0.4 | 0.4 | |||||||||
| dcGTX1&4 | 0.1 c | ||||||||||
| dcGTX2&3 | 0.17 b | 0.11 b | 0.04 | 0.05 | 0.22 | 0.01 | 3.3. | 0.02 | 3.1 | ||
| C1 | 0.01 | ||||||||||
| C2 | 0.1 | 0.1 | |||||||||
| C3 | 0.01 | ||||||||||
| C4 | 0.1 | 0.1 | |||||||||
| C1&2 | 0.034 b | 0.043 b | 0.008 | 0.013 | 0.25 | ND | 2.6 | 0.09 | 0.1 | ||
| C3&4 | 0.028 b | ND b | |||||||||
| 11-hydroxy-STX | 0.3 | ||||||||||
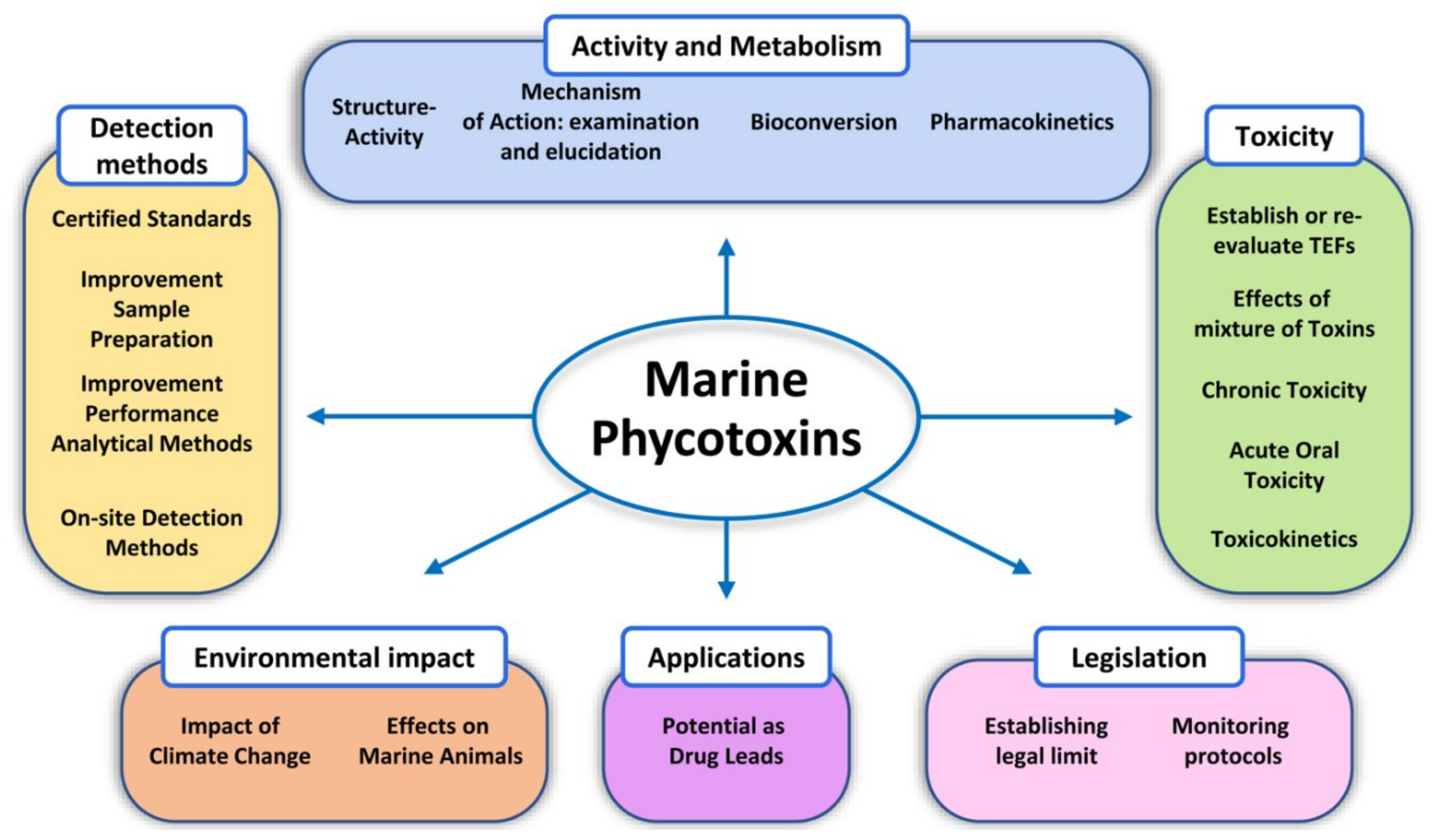
- Epidemiology studies;
- Studies of structure–activity relationship of toxins;
- Common criteria in the naming of toxins;
- Reevaluation of preestablished toxicity concepts based on false premises;
- Review mechanism of action responsible for the toxicity of compounds including mechanisms involved in the disturbances that can persist or reoccur many months or even years afterwards;
- Harmonization of criteria to set toxicity parameters to establish accurate TEF values especially of those toxin analogues commonly found in seafood or at relatively high levels;
- Research to better understand the toxins produced by bioconversion in the organisms and their toxicity;
- Information about pharmacokinetics of toxins;
- Toxicity studies with special focus on oral toxicity and on toxin mixtures;
- Studies related to chronic exposure of toxins;
- Information on the occurrence and factors conducive to the accumulation of toxins in marine organisms;
- Common legislative criteria: toxin regulation, implementation of effective toxin monitoring, and management programs for toxins;
- Climate change and its consequences;
- Evaluation of the therapeutic potential of these toxins based on the reversible interaction with the sodium channels.
3.2. Marine Phycotoxins Acting on Glutamate Receptors: Domoic Acid and Analogues
- Studies on the oral toxicity of DA isomers present in seafood.
- The health impacts associated with chronic, low-dose exposure to this prevalent neurotoxin. Results from these studies will also help reveal the human subpopulations with pre-existing conditions who may be more vulnerable to the toxic effects of this compound.
- Studies to further elucidate the toxicokinetic of DA and the role of drug transporters
- Research into DA effects other than neurotoxic (cardiac, renal, and gastrointestinal) especially considering chronic exposure.
- Research in humans and animal models should include studies during pregnancy and in exposed offspring to characterize the relationship between the increasing body burden of DA and related neurodevelopmental effects.
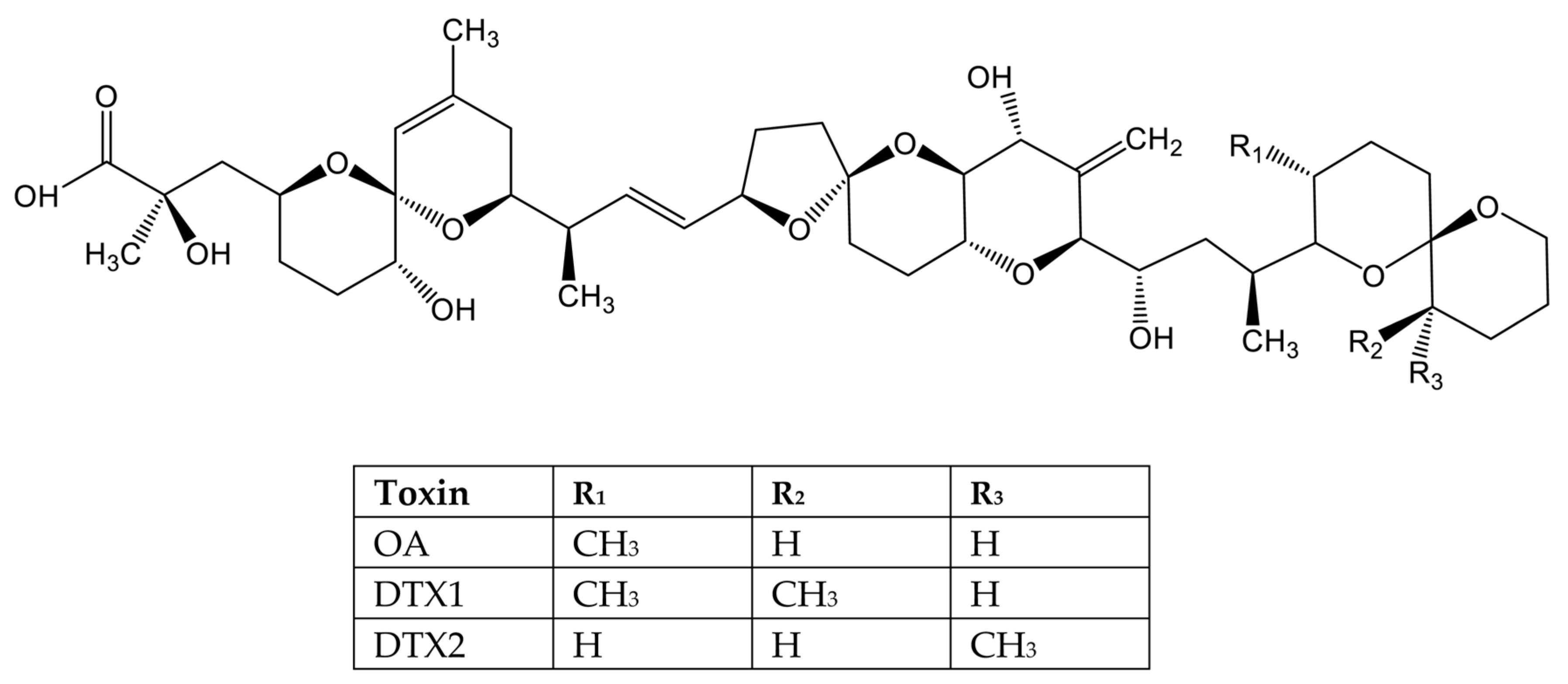
3.3. Lipophilic Marine Phycotoxins
- Well-characterized epidemiology studies;
- Further understanding of the molecular target of phycotoxins;
- Review the mechanisms of phycotoxins toxicity;
- Establishment of objective toxicity parameters to determine accurate TEF values;
- Advances on the knowledge of oral toxicity;
- Health effects associated to the chronic exposure of phycotoxins and studies of repeated exposure to toxins at levels below the current regulatory limit;
- Elucidation of the mechanisms of phycotoxins biotransformation in seafood and in the human body;
- Improvements in the information regarding toxicokinetics (i.e., absorption, distribution, metabolism, and elimination);
- Increase of data concerning the bioactivity of different toxins relevant to the assessment of toxicity;
- Further research on the effects of toxins mixtures;
- Identification of the impacts of phycotoxins on marine animals;
- Common legislative criteria: toxin regulation, implementation of effective toxin monitoring, and management programs for toxins.

3.4. Toxins Acting on Ion Pumps: Palytoxins, Ostreocins, and Ovatoxins
- Studies to assess the real hazard they present to humans;
- Detailed epidemiological studies are needed to better evaluate safety levels and to promote regulations that will protect human health and reduce economic losses;
- Pharmacological and toxicological effects of each PLTX analogue to carry out reliable structure–activity relationship;
- Evaluation of PLTX and analogues oral toxicity;
- Studies exploring treatments for PLTX including search for effective antidotes;
- Common legislative criteria: toxin regulation, implementation of effective toxin monitoring, and management programs for toxins.
4. Marine Toxins as a Source of Drugs
5. Detection Methods
5.1. Analytical Detection Methods
5.2. Molecular Interaction/Function-Based Assays
5.3. Cell-Based Assays
| Marine Toxin | Exposure Time | Cell Line | Detection Method | LOD (IC50) | Reference |
|---|---|---|---|---|---|
| OA DTX-1, DTX-2 | 24 h | V79 cells | MTT assay | (27 nM) | [414] |
| 48 h | HepG2 cells | MTT assay | (30.2 nM) | [415] | |
| 24 h | Neuro2a cells | MTT assay | (11.20 nM) | [203] | |
| AZAs | 48 h | HepG2 cells | MTT assay | (4.3 nM) | [415] |
| PSTs | 24 h | Neuro2a cells | MTT assay | 0.91 nM (8.6 nM) | [403] |
| 24 h | NG108-15 cells | MTT assay | 4.2 nM (8.2 nM) | [403] | |
| 24 h | Neuro2a cells | Electrical impedance | 0.1 nM | [409] | |
| 24 h | Neuro2a cells | qRT-PCR | 17 nM | [412] | |
| 15 min | BE(2)-M17 cells | Fluorimetry | 4.36 nM | [410] | |
| Immediately | Rat cortical neurones | Spectrofluorimeter | 1 nM | [416] | |
| CTXs | 20 h | Neuro2a cells | MTS assay | (0.02 nM C-CTX-1) | [417] |
| PLTXs | 24 h | Erythrocytes | Spectrometry | 0.37 pM | [405] |
| 24 h | BE(2)-M17 cells | Spectrofluorimetry | 0.07 nM | [401] |
5.4. Animal Bioassays
- On-site, easy-to-use, efficient methods for detection of multiple toxin groups are not yet available;
- Certified analytical standards of some toxin classes are urgently needed;
- Improvement of sample preparation procedures for further testing or extended automation of routine monitoring;
- Reliable TEF estimation for many analogues of these toxin groups is still missing;
- Improvement of performance of analytical methods, especially for ciguatoxins and palytoxins.
6. Climate Change Uncertainties
7. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Botana, L.M.; Louzao, M.C.; Vilariño, N. Climate Change and Marine and Freshwater Toxins, 2nd ed.; De Gruyter: Berlin, Germany, 2021; p. 661. [Google Scholar]
- Farabegoli, F.; Blanco, L.; Rodriguez, L.P.; Vieites, J.M.; Cabado, A.G. Phycotoxins in Marine Shellfish: Origin, Occurrence and Effects on Humans. Mar. Drugs 2018, 16, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deeds, J.R.; Landsberg, J.H.; Etheridge, S.M.; Pitcher, G.C.; Longan, S.W. Non-traditional vectors for paralytic shellfish poisoning. Mar. Drugs 2008, 6, 308–348. [Google Scholar] [CrossRef] [PubMed]
- Leal, J.F.; Cristiano, M.L.S. Marine paralytic shellfish toxins: Chemical properties, mode of action, newer analogues, and structure-toxicity relationship. Nat. Prod. Rep. 2022, 39, 33–57. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific Opinion of the Panel on Contaminants in the Food Chain on a request from the European Commission on Marine Biotoxins in Shellfish—Saxitoxin Group. EFSA J. 2009, 1019, 1–76. [Google Scholar]
- WHO. Cyanobacterial Toxins: Saxitoxins. In Background Document for Development of WHO Guidelines for Drinking-Water Quality and Guidelines for Safe Recreational Water Environments; WHO: Genoa, Italy, 2020. [Google Scholar]
- Catterall, W.A. Forty Years of Sodium Channels: Structure, Function, Pharmacology, and Epilepsy. Neurochem. Res. 2017, 42, 2495–2504. [Google Scholar] [CrossRef]
- Katikou, P.; Gokbulut, C.; Kosker, A.R.; Campas, M.; Ozogi, R. An updated review of Tetrodotoxin and its peculiarities. Mar. Drugs 2022, 20, 47. [Google Scholar] [CrossRef]
- Bates, S.S.; Hubbard, K.A.; Lundholm, N.; Montresor, M.; Leaw, C.P. Pseudo-nitzschia, Nitzschia, and domoic acid: New research since 2011. Harmful Algae 2018, 79, 3–43. [Google Scholar] [CrossRef]
- Lefebvre, K.A.; Bargu, S.; Kieckhefer, T.; Silver, M.W. From sanddabs to blue whales: The pervasiveness of domoic acid. Toxicon 2002, 40, 971–977. [Google Scholar] [CrossRef]
- Visciano, P.; Schirone, M.; Berti, M.; Milandri, A.; Tofalo, R.; Suzzi, G. Marine Biotoxins: Occurrence, Toxicity, Regulatory Limits and Reference Methods. Front. Microbiol. 2016, 7, 1051. [Google Scholar] [CrossRef] [Green Version]
- Bresnan, E.; Arevalo, F.; Belin, C.; Branco, M.A.C.; Cembella, A.D.; Clarke, D.; Correa, J.; Davidson, K.; Dhanji-Rapkova, M.; Lozano, R.F.; et al. Diversity and regional distribution of harmful algal events along the Atlantic margin of Europe. Harmful Algae 2021, 102, 101976. [Google Scholar] [CrossRef]
- Reguera, B.; Riobo, P.; Rodriguez, F.; Diaz, P.A.; Pizarro, G.; Paz, B.; Franco, J.M.; Blanco, J. Dinophysis toxins: Causative organisms, distribution and fate in shellfish. Mar. Drugs 2014, 12, 394–461. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.L.; Zhao, X.Y.; Ji, L.D.; Xu, J. Okadaic acid (OA): Toxicity, detection and detoxification. Toxicon 2019, 160, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Molgo, J.; Marchot, P.; Araoz, R.; Benoit, E.; Iorga, B.I.; Zakarian, A.; Taylor, P.; Bourne, Y.; Servent, D. Cyclic imine toxins from dinoflagellates: A growing family of potent antagonists of the nicotinic acetylcholine receptors. J. Neurochem. 2017, 142 (Suppl. 2), 41–51. [Google Scholar] [CrossRef] [PubMed]
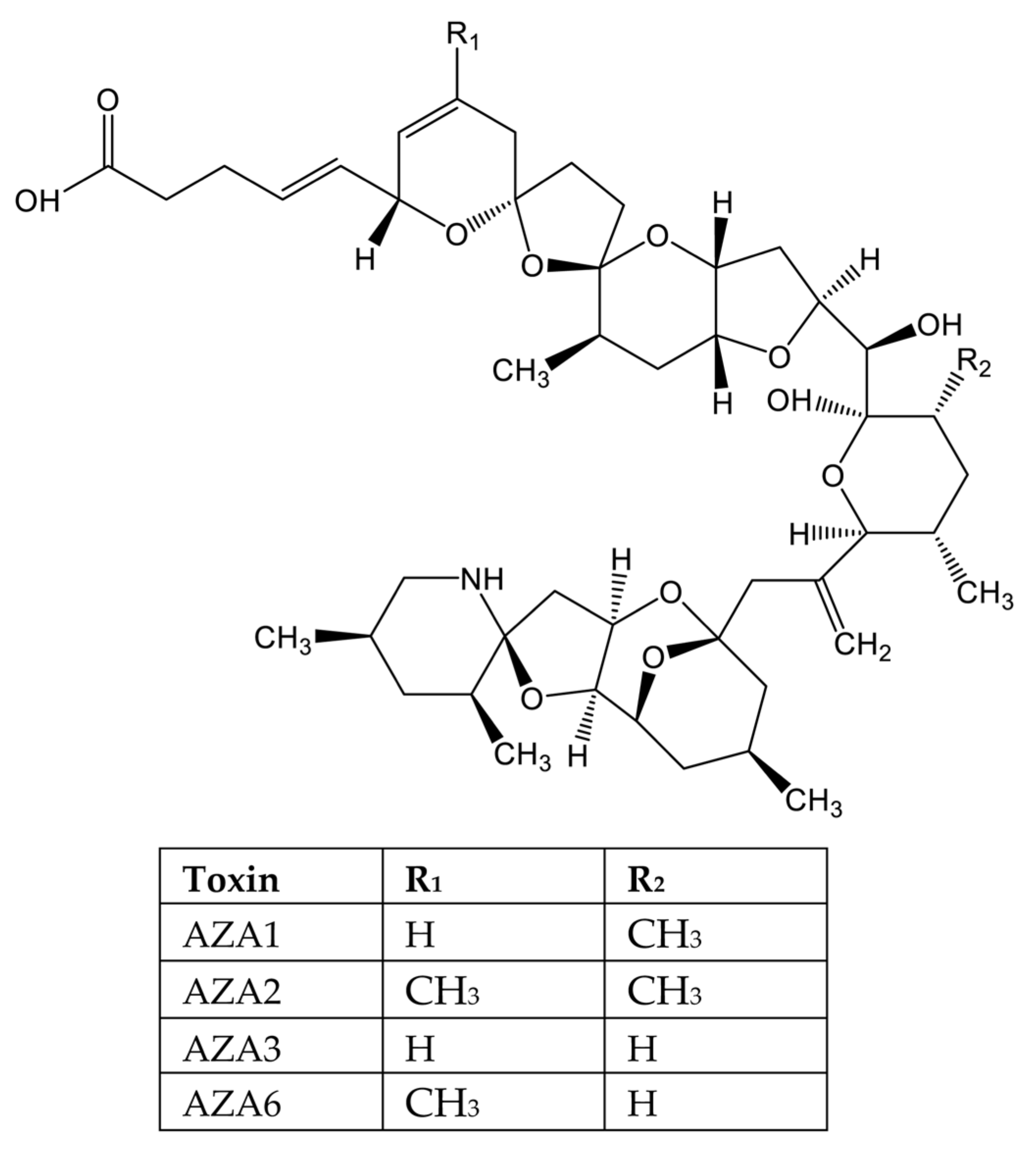
- Abal, P.; Louzao, M.C.; Fraga, M.; Vilarino, N.; Ferreiro, S.; Vieytes, M.R.; Botana, L.M. Absorption and Effect of Azaspiracid-1 Over the Human Intestinal Barrier. Cell. Physiol. Biochem. 2017, 43, 136–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furey, A.; O’Doherty, S.; O’Callaghan, K.; Lehane, M.; James, K.J. Azaspiracid poisoning (AZP) toxins in shellfish: Toxicological and health considerations. Toxicon 2010, 56, 173–190. [Google Scholar] [CrossRef]
- FAO/WHO. Report of the Joint FAO/IOC/WHO As Hoc Expert Consultation on Biotoxins in Bivalve Molluscs; Food and Agriculture Organization of the United Nations: Rome, Italy, 2004. [Google Scholar]
- EFSA. Opinion of the Scientific Panel on Contaminants in the Food chain on a request from the European Commission on marine biotoxins in shellfish—Azaspiracids. EFSA J. 2008, 723, 1–52. [Google Scholar]
- Kim, J.H.; Tillmann, U.; Adams, N.G.; Krock, B.; Stutts, W.L.; Deeds, J.R.; Han, M.S.; Trainer, V.L. Identification of Azadinium species and a new azaspiracid from Azadinium poporum in Puget Sound, Washington State, USA. Harmful Algae 2017, 68, 152–167. [Google Scholar] [CrossRef] [Green Version]
- Krock, B.; Tillmann, U.; Tebben, J.; Trefault, N.; Gu, H. Two novel azaspiracids from Azadinium poporum, and a comprehensive compilation of azaspiracids produced by Amphidomataceae, (Dinophyceae). Harmful Algae 2019, 82, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Vale, P.; Bire, R.; Hess, P. Confirmation by LC-MS/MS of azaspiracids in shellfish from the Portuguese north-western coast. Toxicon 2008, 51, 1449–1456. [Google Scholar] [CrossRef]
- Blanco, J.; Arevalo, F.; Morono, A.; Correa, J.; Muniz, S.; Marino, C.; Martin, H. Presence of azaspiracids in bivalve molluscs from Northern Spain. Toxicon 2017, 137, 135–143. [Google Scholar] [CrossRef]
- Bacchiocchi, S.; Siracusa, M.; Ruzzi, A.; Gorbi, S.; Ercolessi, M.; Cosentino, M.A.; Ammazzalorso, P.; Orletti, R. Two-year study of lipophilic marine toxin profile in mussels of the North-central Adriatic Sea: First report of azaspiracids in Mediterranean seafood. Toxicon 2015, 108, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Goya, A.B. Occurrence and profiles of lipophilic toxins in shellfish harvested from Argentina. Toxicon 2015, 102, 32–42. [Google Scholar] [CrossRef] [PubMed]
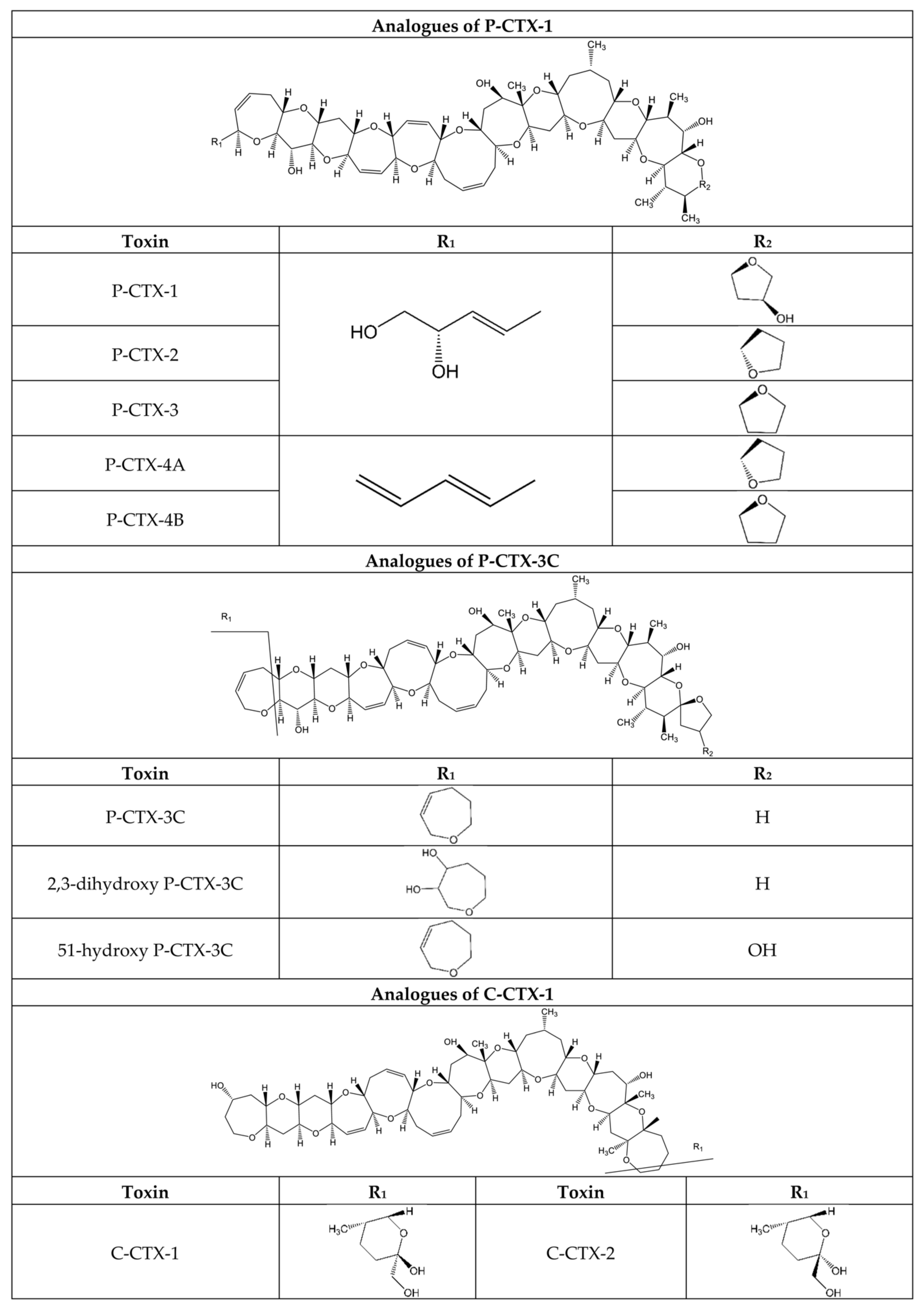
- Friedman, M.A.; Fernandez, M.; Backer, L.C.; Dickey, R.W.; Bernstein, J.; Schrank, K.; Kibler, S.; Stephan, W.; Gribble, M.O.; Bienfang, P.; et al. An Updated Review of Ciguatera Fish Poisoning: Clinical, Epidemiological, Environmental, and Public Health Management. Mar. Drugs 2017, 15, 72. [Google Scholar] [CrossRef] [PubMed]
- Katikou, P. Digital Technologies and Open Data Sources in Marine Biotoxins’ Risk Analysis: The Case of Ciguatera Fish Poisoning. Toxins 2021, 13, 692. [Google Scholar] [CrossRef] [PubMed]
- Chinain, M.; Gatti, C.M.I.; Darius, H.T.; Quod, J.P.; Tester, P.A. Ciguatera poisonings: A global review of occurrences and trends. Harmful Algae 2021, 102, 101873. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, I.; Genta-Jouve, G.; Alfonso, C.; Calabro, K.; Alonso, E.; Sanchez, J.A.; Alfonso, A.; Thomas, O.P.; Botana, L.M. Gambierone, a Ladder-Shaped Polyether from the Dinoflagellate Gambierdiscus belizeanus. Org. Lett. 2015, 17, 2392–2395. [Google Scholar] [CrossRef] [PubMed]
- FAO/WHO. Report of the Expert Meeting on Ciguatera Poisoning: Rome, 19–23 November 2018; Food Safety and Quality Series 9; FAO: Rome, Italy, 2020. [Google Scholar]
- Konoki, K.; Baden, D.G.; Scheuer, T.; Catterall, W.A. Molecular Determinants of Brevetoxin Binding to Voltage-Gated Sodium Channels. Toxins 2019, 11, 513. [Google Scholar] [CrossRef] [Green Version]
- Mackieh, R.; Abou-Nader, R.; Wehbe, R.; Mattei, C.; Legros, C.; Fajloun, Z.; Sabatier, J.M. Voltage-Gated Sodium Channels: A Prominent Target of Marine Toxins. Mar. Drugs 2021, 19, 562. [Google Scholar] [CrossRef]
- Turner, A.D.; Higgins, C.; Davidson, K.; Veszelovszki, A.; Payne, D.; Hungerford, J.; Higman, W. Potential threats posed by new or emerging marine biotoxins in UK waters and examination of detection methodology used in their control: Brevetoxins. Mar. Drugs 2015, 13, 1224–1254. [Google Scholar] [CrossRef] [Green Version]
- Arnich, N.; Abadie, E.; Amzil, Z.; Bottein, M.Y.D.; Comte, K.; Chaix, E.; Delcourt, N.; Hort, V.; Mattei, C.; Molgo, J.; et al. Guidance Level for Brevetoxins in French Shellfish. Mar. Drugs 2021, 19, 520. [Google Scholar] [CrossRef]
- Hort, V.; Abadie, E.; Arnich, N.; Dechraoui Bottein, M.Y.; Amzil, Z. Chemodiversity of Brevetoxins and Other Potentially Toxic Metabolites Produced by Karenia spp. and Their Metabolic Products in Marine Organisms. Mar. Drugs 2021, 19, 656. [Google Scholar] [CrossRef] [PubMed]
- Accoroni, S.; Tartaglione, L.; Dello Iacovo, E.; Pichierri, S.; Marini, M.; Campanelli, A.; Dell’Aversano, C.; Totti, C. Influence of environmental factors on the toxin production of Ostreopsis cf. ovata during bloom events. Mar. Pollut. Bull. 2017, 123, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Lounas, R.; Kasmi, H.; Chernai, S.; Amarni, N.; Hamdi, B. Dynamics of the genus Ostreopsis (Gonyaulacales, Dinophyceae) in a Mediterranean fish farm. Environ. Monit. Assess. 2021, 193, 333. [Google Scholar] [CrossRef] [PubMed]
- Botana, L.M. Toxicological Perspective on Climate Change: Aquatic Toxins. Chem. Res. Toxicol. 2016, 29, 619–625. [Google Scholar] [CrossRef] [Green Version]
- Abal, P.; Louzao, M.C.; Antelo, A.; Alvarez, M.; Cagide, E.; Vilarino, N.; Vieytes, M.R.; Botana, L.M. Acute Oral Toxicity of Tetrodotoxin in Mice: Determination of Lethal Dose 50 (LD50) and No Observed Adverse Effect Level (NOAEL). Toxins 2017, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abal, P.; Louzao, M.C.; Suzuki, T.; Watanabe, R.; Vilarino, N.; Carrera, C.; Botana, A.M.; Vieytes, M.R.; Botana, L.M. Toxic Action Reevaluation of Okadaic Acid, Dinophysistoxin-1 and Dinophysistoxin-2: Toxicity Equivalency Factors Based on the Oral Toxicity Study. Cell. Physiol. Biochem. 2018, 49, 743–757. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, K.A.; Robertson, A. Domoic acid and human exposure risks: A review. Toxicon 2010, 56, 218–230. [Google Scholar] [CrossRef]
- Pearn, J. Neurology of ciguatera. J. Neurol. Neurosurg. Psychiatry 2001, 70, 4–8. [Google Scholar] [CrossRef] [Green Version]
- Boente-Juncal, A.; Otero, P.; Rodriguez, I.; Camina, M.; Rodriguez-Vieytes, M.; Vale, C.; Botana, L.M. Oral Chronic Toxicity of the Safe Tetrodotoxin Dose Proposed by the European Food Safety Authority and Its Additive Effect with Saxitoxin. Toxins 2020, 12, 312. [Google Scholar] [CrossRef]
- Boente-Juncal, A.; Raposo-Garcia, S.; Vale, C.; Louzao, M.C.; Otero, P.; Botana, L.M. In Vivo Evaluation of the Chronic Oral Toxicity of the Marine Toxin Palytoxin. Toxins 2020, 12, 489. [Google Scholar] [CrossRef]
- Boente-Juncal, A.; Vale, C.; Cifuentes, M.; Otero, P.; Camina, M.; Rodriguez-Vieytes, M.; Botana, L.M. Chronic In Vivo Effects of Repeated Exposure to Low Oral Doses of Tetrodotoxin: Preliminary Evidence of Nephrotoxicity and Cardiotoxicity. Toxins 2019, 11, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vetter, I.; Mozar, C.A.; Durek, T.; Wingerd, J.S.; Alewood, P.F.; Christie, M.J.; Lewis, R.J. Characterisation of Na(v) types endogenously expressed in human SH-SY5Y neuroblastoma cells. Biochem. Pharmacol. 2012, 83, 1562–1571. [Google Scholar] [CrossRef] [PubMed]
- Viallon, J.; Chinain, M.; Darius, H.T. Revisiting the Neuroblastoma Cell-Based Assay (CBA-N2a) for the Improved Detection of Marine Toxins Active on Voltage Gated Sodium Channels (VGSCs). Toxins 2020, 12, 281. [Google Scholar] [CrossRef]
- Botana, L.; Hess, P.; Munday, R.; Nathalie, A.; DeGrasse, S.; Feeley, M.; Suzuki, T.; van den Berg, M.; Fattori, V.; Gamarro, E.; et al. Derivation of toxicity equivalency factors for marine biotoxins associated with Bivalve Molluscs. Trends Food Sci. Technol. 2017, 59, 15–24. [Google Scholar] [CrossRef] [Green Version]
- FAO/WHO. Codex Stan 292-2008: Standard for live and raw bivalve molluscs. Codex Alimentarius. FAO/WHO: Rome, Italy, 2015; pp. 1–9. [Google Scholar]
- AOAC. AOAC Official method 959.08. Paralytic shellfish poison. Biological method. In Official Methods of Analysis of AOAC International, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Rodriguez, I.; Vieytes, M.R.; Alfonso, A. Analytical challenges for regulated marine toxins. Detection methods. Curr. Opin. Food Sci. 2017, 19, 29–36. [Google Scholar] [CrossRef]
- Hwang, D.F.; Noguchi, T. Tetrodotoxin poisoning. Adv. Food Nutr. Res. 2007, 52, 141–236. [Google Scholar]
- Noguchi, T.; Onuki, K.; Arakawa, O. Tetrodotoxin poisoning due to pufferfish and gastropods, and their intoxication mechanism. ISRN Toxicol. 2011, 2011, 276939. [Google Scholar] [CrossRef] [Green Version]
- Vlamis, A.; Katikou, P.; Rodriguez, I.; Rey, V.; Alfonso, A.; Papazachariou, A.; Zacharaki, T.; Botana, A.M.; Botana, L.M. First Detection of Tetrodotoxin in Greek Shellfish by UPLC-MS/MS Potentially Linked to the Presence of the Dinoflagellate Prorocentrum minimum. Toxins 2015, 7, 1779–1807. [Google Scholar] [CrossRef] [Green Version]
- EFSA. Scientific opinion on the risk for public health related to the presence of tetrodotoxin (TTX) and TTX analogues in marine bivalves and gastropods. EFSA J. 2017, 15, 65. [Google Scholar]
- Abal, P.; Louzao, M.C.; Vilarino, N.; Vieytes, M.R.; Botana, L.M. Acute Toxicity Assessment: Macroscopic and Ultrastructural Effects in Mice Treated with Oral Tetrodotoxin. Toxins 2019, 11, 305. [Google Scholar] [CrossRef] [Green Version]
- Costa, P.R.; Estevez, P.; Solino, L.; Castro, D.; Rodrigues, S.M.; Timoteo, V.; Leao-Martins, J.M.; Santos, C.; Gouveia, N.; Diogene, J.; et al. An Update on Ciguatoxins and CTX-like Toxicity in Fish from Different Trophic Levels of the Selvagens Islands (NE Atlantic, Madeira, Portugal). Toxins 2021, 13, 580. [Google Scholar] [CrossRef] [PubMed]
- Inserra, M.C.; Israel, M.R.; Caldwell, A.; Castro, J.; Deuis, J.R.; Harrington, A.M.; Keramidas, A.; Garcia-Caraballo, S.; Maddern, J.; Erickson, A.; et al. Multiple sodium channel isoforms mediate the pathological effects of Pacific ciguatoxin-1. Sci. Rep. 2017, 7, 42810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raposo-Garcia, S.; Louzao, M.C.; Fuwa, H.; Sasaki, M.; Vale, C.; Botana, L.M. Determination of the toxicity equivalency factors for ciguatoxins using human sodium channels. Food Chem. Toxicol. 2022, 160, 112812. [Google Scholar] [CrossRef] [PubMed]
- Accoroni, S.; Romagnoli, T.; Penna, A.; Capellacci, S.; Ciminiello, P.; Dell’Aversano, C.; Tartaglione, L.; Abboud-Abi Saab, M.; Giussani, V.; Asnaghi, V.; et al. Ostreopsis fattorussoi sp. nov. (Dinophyceae), a new benthic toxic Ostreopsis species from the eastern Mediterranean Sea. J. Phycol. 2016, 52, 1064–1084. [Google Scholar] [CrossRef]
- Carnicer, O.; Guallar, C.; Andree, K.B.; Diogene, J.; Fernandez-Tejedor, M. Ostreopsis cf. ovata dynamics in the NW Mediterranean Sea in relation to biotic and abiotic factors. Environ. Res. 2015, 143 Pt B, 89–99. [Google Scholar] [CrossRef]
- EU. Commission Regulation (EU) No 15/2011 of 10 January 2011 amending Regulation (EC) No 2074/2005 as regards recognised testing methods for detecting marine biotoxins in live bivalve molluscs (Text with EEA relevance). Off. J. Eur. Union 2011, L6, 3–6. [Google Scholar]
- Otero, P.; Alfonso, A.; Alfonso, C.; Rodriguez, P.; Vieytes, M.R.; Botana, L.M. Effect of uncontrolled factors in a validated liquid chromatography-tandem mass spectrometry method question its use as a reference method for marine toxins: Major causes for concern. Anal. Chem. 2011, 83, 5903–5911. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion of the Panel on Contaminants in the Food Chain on a request from the European Commission on Marine Biotoxins in Shellfish—Summary on regulated marine biotoxins. EFSA J. 2009, 1306, 1–23. [Google Scholar]
- EU. Commission Delegated Regulation (EU) 2021/1374 of 12 April 2021 Amending Annex III to Regulation (EC) No 853/2004 of the European Parliament and of the Council on Specific Hygiene Requirements for Food of Animal Origin (Text with EEA Rele-Vance). Off. J. Eur. Union 2021, L297, 1–15. [Google Scholar]
- EFSA. Scientific Opinion. Marine biotoxins in shellfish—Pectenotoxin group. Scientific Opinion of the Panel on Contaminants in the Food chain. EFSA J. 2009, 1109, 1–47. [Google Scholar]
- Botana, L.M.; Alfonso, A.; Botana, A.M.; Vieytes, M.R.; Vale, C.; Vilariño, N.; Louzao, M.C. Functional assays for marine toxins as an alternative, high-throughput-screening solution to animal tests. Trends Anal. Chem. 2009, 28, 603–611. [Google Scholar] [CrossRef]
- Botana, L.M.; Vilariño, N.; Alfonso, A.; Vale, C.; Louzao, M.C.; Elliott, C.T.; Campbell, K.; Botana, A.M. The problem of toxicity equivalent factors in developing alternative methods to animal bioassays for marine-toxin detection. Trends Anal. Chem. 2010, 29, 1316–1325. [Google Scholar] [CrossRef]
- Campbell, K.; Vilariño, N.; Botana, L.M.; Elliott, C.T. A European perspective on progress in moving away from the mouse bioassay for marine-toxin analysis. Trends Anal. Chem. 2011, 30, 239–253. [Google Scholar] [CrossRef]
- FAO/WHO. Technical Paper on Toxicity Equivalency Factors for Marine Biotoxins Associated with Bivalve Molluscs; FAO: Rome, Italy, 2016. [Google Scholar]
- Botana, L.M. A perspective on the toxicology of marine toxins. Chem. Res. Toxicol. 2012, 25, 1800–1804. [Google Scholar] [CrossRef]
- Munday, R.; Thomas, K.; Gibbs, R.; Murphy, C.; Quilliam, M.A. Acute toxicities of saxitoxin, neosaxitoxin, decarbamoyl saxitoxin and gonyautoxins 1&4 and 2&3 to mice by various routes of administration. Toxicon 2013, 76, 77–83. [Google Scholar]
- Selwood, A.I.; Waugh, C.; Harwood, D.T.; Rhodes, L.L.; Reeve, J.; Sim, J.; Munday, R. Acute Toxicities of the Saxitoxin Congeners Gonyautoxin 5, Gonyautoxin 6, Decarbamoyl Gonyautoxin 2&3, Decarbamoyl Neosaxitoxin, C-1&2 and C-3&4 to Mice by Various Routes of Administration. Toxins 2017, 9, 73. [Google Scholar]
- Alonso, E.; Alfonso, A.; Vieytes, M.R.; Botana, L.M. Evaluation of toxicity equivalent factors of paralytic shellfish poisoning toxins in seven human sodium channels types by an automated high throughput electrophysiology system. Arch. Toxicol. 2016, 90, 479–488. [Google Scholar] [CrossRef]
- Perez, S.; Vale, C.; Botana, A.M.; Alonso, E.; Vieytes, M.R.; Botana, L.M. Determination of toxicity equivalent factors for paralytic shellfish toxins by electrophysiological measurements in cultured neurons. Chem. Res. Toxicol. 2011, 24, 1153–1157. [Google Scholar] [CrossRef]
- Vale, C.; Wandscheer, C.; Nicolaou, K.C.; Frederick, M.O.; Alfonso, C.; Vieytes, M.R.; Botana, L.M. Cytotoxic effect of azaspiracid-2 and azaspiracid-2-methyl ester in cultured neurons: Involvement of the c-Jun N-terminal kinase. J. Neurosci. Res. 2008, 86, 2952–2962. [Google Scholar] [CrossRef]
- Xu, L.; Ding, X.; Wang, T.; Mou, S.; Sun, H.; Hou, T. Voltage-gated sodium channels: Structures, functions, and molecular modeling. Drug Discov. Today 2019, 24, 1389–1397. [Google Scholar] [CrossRef]
- Duran-Riveroll, L.M.; Cembella, A.D. Guanidinium Toxins and Their Interactions with Voltage-Gated Sodium Ion Channels. Mar. Drugs 2017, 15, 303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartshorne, R.P.; Catterall, W.A. Purification of the saxitoxin receptor of the sodium channel from rat brain. Proc. Natl. Acad. Sci. USA 1981, 78, 4620–4624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, J.R.; Novick, P.A.; Parsons, W.H.; McGregor, M.; Zablocki, J.; Pande, V.S.; Du Bois, J. Marked difference in saxitoxin and tetrodotoxin affinity for the human nociceptive voltage-gated sodium channel (Nav1.7). Proc. Natl. Acad. Sci. USA 2012, 109, 18102–18107. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Liu, D.; Wu, K.; Lei, J.; Yan, N. Structures of human Nav1.7 channel in complex with auxiliary subunits and animal toxins. Science 2019, 363, 1303–1308. [Google Scholar]
- Tikhonov, D.B.; Zhorov, B.S. Predicting Structural Details of the Sodium Channel Pore Basing on Animal Toxin Studies. Front. Pharmacol. 2018, 9, 880. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Li, Z.; Zhou, Q.; Shen, H.; Wu, K.; Huang, X.; Chen, J.; Zhang, J.; Zhu, X.; Lei, J.; et al. Structure of the human voltage-gated sodium channel Nav1.4 in complex with beta1. Science 2018, 362, eaau2486. [Google Scholar] [CrossRef]
- Lee, C.H.; Ruben, P.C. Interaction between voltage-gated sodium channels and the neurotoxin, tetrodotoxin. Channels 2008, 2, 407–412. [Google Scholar] [CrossRef]
- Numano, S.; Kudo, Y.; Cho, Y.; Konoki, K.; Kaga, Y.; Nagasawa, K.; Yotsu-Yamashita, M. Two new skeletal analogues of saxitoxin found in the scallop, Patinopecten yessoensis, as possible metabolites of paralytic shellfish toxins. Chemosphere 2021, 278, 130224. [Google Scholar] [CrossRef]
- Vilarino, N.; Louzao, M.C.; Abal, P.; Cagide, E.; Carrera, C.; Vieytes, M.R.; Botana, L.M. Human Poisoning from Marine Toxins: Unknowns for Optimal Consumer Protection. Toxins 2018, 10, 324. [Google Scholar] [CrossRef] [Green Version]
- Alarcan, J.; Bire, R.; Le Hegarat, L.; Fessard, V. Mixtures of Lipophilic Phycotoxins: Exposure Data and Toxicological Assessment. Mar. Drugs 2018, 16, 46. [Google Scholar] [CrossRef] [Green Version]
- Prakash, A.; Medcof, J.C.; Tennant, A.D. Paralytic Shellfish Poisoning in eastern Canada. Fish. Res. Board Can. 1971, 177, 1–87. [Google Scholar]
- Wang, J.; Salata, J.J.; Bennett, P.B. Saxitoxin is a gating modifier of HERG K+ channels. J. Gen. Physiol. 2003, 121, 583–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Z.; Sheets, M.; Ishida, H.; Li, F.; Barry, W.H. Saxitoxin blocks L-type ICa. J. Pharmacol. Exp. Ther. 2004, 308, 324–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, A.D.; Dhanji-Rapkova, M.; Dean, K.; Milligan, S.; Hamilton, M.; Thomas, J.; Poole, C.; Haycock, J.; Spelman-Marriott, J.; Watson, A.; et al. Fatal Canine Intoxications Linked to the Presence of Saxitoxins in Stranded Marine Organisms Following Winter Storm Activity. Toxins 2018, 10, 94. [Google Scholar] [CrossRef] [Green Version]
- Ben-Gigirey, B.; Rossignoli, A.E.; Riobo, P.; Rodriguez, F. First Report of Paralytic Shellfish Toxins in Marine Invertebrates and Fish in Spain. Toxins 2020, 12, 723. [Google Scholar] [CrossRef]
- Rodriguez-Cabo, T.; Morono, A.; Arevalo, F.; Correa, J.; Lamas, J.P.; Rossignoli, A.E.; Blanco, J. Paralytic Shellfish Poisoning (PSP) in Mussels from the Eastern Cantabrian Sea: Toxicity, Toxin Profile, and Co-Occurrence with Cyclic Imines. Toxins 2021, 13, 761. [Google Scholar] [CrossRef]
- Garcia, C.; del Carmen Bravo, M.; Lagos, M.; Lagos, N. Paralytic shellfish poisoning: Post-mortem analysis of tissue and body fluid samples from human victims in the Patagonia fjords. Toxicon 2004, 43, 149–158. [Google Scholar] [CrossRef]
- Watanabe, R.; Kanamori, M.; Yoshida, H.; Okumura, Y.; Uchida, H.; Matsushima, R.; Oikawa, H.; Suzuki, T. Development of Ultra-Performance Liquid Chromatography with Post-Column Fluorescent Derivatization for the Rapid Detection of Saxitoxin Analogues and Analysis of Bivalve Monitoring Samples. Toxins 2019, 11, 573. [Google Scholar] [CrossRef] [Green Version]
- Turnbull, A.R.; Harwood, D.T.; Boundy, M.J.; Holland, P.T.; Hallegraeff, G.; Malhi, N.; Quilliam, M.A. Paralytic shellfish toxins—Call for uniform reporting units. Toxicon 2020, 178, 59–60. [Google Scholar] [CrossRef]
- Qiu, J.; Meng, F.; Ding, L.; Che, Y.; McCarron, P.; Beach, D.G.; Li, A. Dynamics of paralytic shellfish toxins and their metabolites during timecourse exposure of scallops Chlamys farreri and mussels Mytilus galloprovincialis to Alexandrium pacificum. Aquat. Toxicol. 2018, 200, 233–240. [Google Scholar] [CrossRef]
- Boundy, M.J.; Harwood, D.T.; Tommasi, E.; Burger, E.; van Ginkel, R.; Waugh, C.; Selwood, A.I.; Finch, S. Acute toxicity of decarbamoyl gonyautoxin 1&4 to mice by various routes of administration. Toxicon 2021, 204, 56–63. [Google Scholar] [PubMed]
- Munday, R. Is protein phosphatase inhibition responsible for the toxic effects of okadaic Acid in animals? Toxins 2013, 5, 267–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shearn-Bochsler, V.; Lance, E.W.; Corcoran, R.; Piatt, J.; Bodenstein, B.; Frame, E.; Lawonn, J. Fatal paralytic shellfish poisoning in Kittlitz’s Murrelet (Brachyramphus brevirostris) nestlings, Alaska, USA. J. Wildl. Dis. 2014, 50, 933–937. [Google Scholar] [CrossRef] [PubMed]
- Ben-Gigirey, B.; Solino, L.; Bravo, I.; Rodriguez, F.; Casero, M.V.M. Paralytic and Amnesic Shellfish Toxins Impacts on Seabirds, Analyses and Management. Toxins 2021, 13, 454. [Google Scholar] [CrossRef]
- Dusek, R.J.; Smith, M.M.; Van Hemert, C.; Shearn-Bochsler, V.I.; Hall, S.; Ridge, C.D.; Hardison, D.R.; Kaler, R.S.A.; Bodenstein, B.L.; Hofmeister, E.K.; et al. Acute oral toxicity and tissue residues of saxitoxin in the mallard (Anas platyrhynchos). Harmful Algae 2021, 109, 102109. [Google Scholar] [CrossRef]
- EU. Regulation (EC) No 853/2004 of the European Parliament and of the Council of 29 April 2004 laying down specific hygiene rules for food of animal origin. Off. J. Eur. Union 2004, L139, 55–205. [Google Scholar]
- Finch, S.C.; Webb, N.G.; Boundy, M.J.; Harwood, D.T.; Munday, J.S.; Sprosen, J.M.; Cave, V.M.; Broadhurst, R.B.; Nicolas, J. Sub-Acute Feeding Study of Saxitoxin to Mice Confirms the Effectiveness of Current Regulatory Limits for Paralytic Shellfish Toxins. Toxins 2021, 13, 627. [Google Scholar] [CrossRef]
- Lima-Filho, C.M.; Nogaroli, L.; Hedin-Pereira, C.; Azevedo, S.; Soares, R.M. Effects of saxitoxins exposure on oligodendrocyte development in mouse neonates. Toxicon 2020, 188, 89–94. [Google Scholar] [CrossRef]
- Sun, Q.; Chen, X.; Liu, W.; Li, S.; Zhou, Y.; Yang, X.; Liu, J. Effects of long-term low dose saxitoxin exposure on nerve damage in mice. Aging 2021, 13, 17211–17226. [Google Scholar] [CrossRef]
- Turner, A.D.; Powell, A.; Schofield, A.; Lees, D.N.; Baker-Austin, C. Detection of the pufferfish toxin tetrodotoxin in European bivalves, England, 2013 to 2014. Euro Surveill. 2015, 20, 21009. [Google Scholar] [CrossRef]
- Rodriguez, P.; Alfonso, A.; Vale, C.; Alfonso, C.; Vale, P.; Tellez, A.; Botana, L.M. First toxicity report of tetrodotoxin and 5,6,11-trideoxyTTX in the trumpet shell Charonia lampas lampas in Europe. Anal. Chem. 2008, 80, 5622–5629. [Google Scholar] [CrossRef] [PubMed]
- Bane, V.; Lehane, M.; Dikshit, M.; O’Riordan, A.; Furey, A. Tetrodotoxin: Chemistry, toxicity, source, distribution and detection. Toxins 2014, 6, 693–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botana, L.M.; Alfonso, A.; Rodriguez, I.; Botana, A.M.; Louzao Mdel, C.; Vieytes, M.R. How Safe Is Safe for Marine Toxins Monitoring? Toxins 2016, 8, 208. [Google Scholar] [CrossRef] [Green Version]
- Katikou, P. Human impact in Mediterraneal coastal ecosystems and climate change: Emerging toxins. In Climate Change and Marine and Freshwater Toxins, 2nd ed.; Botana, L.M., Louzao, M.C., Vilariño, N., Eds.; De Gruyter: Berlin, Germany, 2021; pp. 223–301. [Google Scholar]
- Van Theemsche, K.M.; Van de Sande, D.V.; Snyders, D.J.; Labro, A.J. Hydrophobic Drug/Toxin Binding Sites in Voltage-Dependent K(+) and Na(+) Channels. Front. Pharmacol. 2020, 11, 735. [Google Scholar] [CrossRef]
- Hidalgo, J.; Liberona, J.L.; Molgo, J.; Jaimovich, E. Pacific ciguatoxin-1b effect over Na+ and K+ currents, inositol 1,4,5-triphosphate content and intracellular Ca2+ signals in cultured rat myotubes. Br. J. Pharmacol. 2002, 137, 1055–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- L’Herondelle, K.; Talagas, M.; Mignen, O.; Misery, L.; Le Garrec, R. Neurological Disturbances of Ciguatera Poisoning: Clinical Features and Pathophysiological Basis. Cells 2020, 9, 2291. [Google Scholar] [CrossRef] [PubMed]
- Leung, P.T.Y.; Yan, M.; Lam, V.T.T.; Yiu, S.K.F.; Chen, C.Y.; Murray, J.S.; Harwood, D.T.; Rhodes, L.L.; Lam, P.K.S.; Wai, T.C. Phylogeny, morphology and toxicity of benthic dinoflagellates of the genus Fukuyoa (Goniodomataceae, Dinophyceae) from a subtropical reef ecosystem in the South China Sea. Harmful Algae 2018, 74, 78–97. [Google Scholar] [CrossRef]
- Diogene, J.; Reverte, L.; Rambla-Alegre, M.; Del Rio, V.; de la Iglesia, P.; Campas, M.; Palacios, O.; Flores, C.; Caixach, J.; Ralijaona, C.; et al. Identification of ciguatoxins in a shark involved in a fatal food poisoning in the Indian Ocean. Sci. Rep. 2017, 7, 8240. [Google Scholar] [CrossRef] [PubMed]
- Ikehara, T.; Kuniyoshi, K.; Oshiro, N.; Yasumoto, T. Biooxidation of Ciguatoxins Leads to Species-Specific Toxin Profiles. Toxins 2017, 9, 205. [Google Scholar] [CrossRef] [Green Version]
- Lewis, R.J. The changing face of ciguatera. Toxicon 2001, 39, 97–106. [Google Scholar] [CrossRef]
- Rubiolo, J.A.; Vale, C.; Martin, V.; Fuwa, H.; Sasaki, M.; Botana, L.M. Potassium currents inhibition by gambierol analogs prevents human T lymphocyte activation. Arch. Toxicol. 2015, 89, 1119–1134. [Google Scholar] [CrossRef] [PubMed]
- Kopljar, I.; Labro, A.J.; de Block, T.; Rainier, J.D.; Tytgat, J.; Snyders, D.J. The ladder-shaped polyether toxin gambierol anchors the gating machinery of Kv3.1 channels in the resting state. J. Gen. Physiol. 2013, 141, 359–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuypers, E.; Abdel-Mottaleb, Y.; Kopljar, I.; Rainier, J.D.; Raes, A.L.; Snyders, D.J.; Tytgat, J. Gambierol, a toxin produced by the dinoflagellate Gambierdiscus toxicus, is a potent blocker of voltage-gated potassium channels. Toxicon 2008, 51, 974–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boente-Juncal, A.; Vale, C.; Alfonso, A.; Botana, L.M. Synergistic Effect of Transient Receptor Potential Antagonist and Amiloride against Maitotoxin Induced Calcium Increase and Cytotoxicity in Human Neuronal Stem Cells. ACS Chem. Neurosci. 2018, 9, 2667–2678. [Google Scholar] [CrossRef] [PubMed]
- Boente-Juncal, A.; Alvarez, M.; Antelo, A.; Rodriguez, I.; Calabro, K.; Vale, C.; Thomas, O.P.; Botana, L.M. Structure Elucidation and Biological Evaluation of Maitotoxin-3, a Homologue of Gambierone, from Gambierdiscus belizeanus. Toxins 2019, 11, 79. [Google Scholar] [CrossRef] [Green Version]
- EFSA. Scientific Opinion on marine biotoxins in shellfish–Emerging toxins: Ciguatoxin group. EFSA J. 2010, 8, 1627. [Google Scholar]
- Solino, L.; Costa, P.R. Global impact of ciguatoxins and ciguatera fish poisoning on fish, fisheries and consumers. Environ. Res. 2020, 182, 109111. [Google Scholar] [CrossRef]
- Diaz-Asencio, L.; Clausing, R.J.; Vandersea, M.; Chamero-Lago, D.; Gomez-Batista, M.; Hernandez-Albernas, J.I.; Chomerat, N.; Rojas-Abrahantes, G.; Litaker, R.W.; Tester, P.; et al. Ciguatoxin Occurrence in Food-Web Components of a Cuban Coral Reef Ecosystem: Risk-Assessment Implications. Toxins 2019, 11, 722. [Google Scholar] [CrossRef] [Green Version]
- Roue, M.; Darius, H.T.; Ung, A.; Viallon, J.; Sibat, M.; Hess, P.; Amzil, Z.; Chinain, M. Tissue Distribution and Elimination of Ciguatoxins in Tridacna maxima (Tridacnidae, Bivalvia) Fed Gambierdiscus polynesiensis. Toxins 2018, 10, 189. [Google Scholar] [CrossRef] [Green Version]
- Vetter, I.; Touska, F.; Hess, A.; Hinsbey, R.; Sattler, S.; Lampert, A.; Sergejeva, M.; Sharov, A.; Collins, L.S.; Eberhardt, M.; et al. Ciguatoxins activate specific cold pain pathways to elicit burning pain from cooling. EMBO J. 2012, 31, 3795–3808. [Google Scholar] [CrossRef] [Green Version]
- Au, N.P.; Kumar, G.; Asthana, P.; Tin, C.; Mak, Y.L.; Chan, L.L.; Lam, P.K.; Ma, C.H. Ciguatoxin reduces regenerative capacity of axotomized peripheral neurons and delays functional recovery in pre-exposed mice after peripheral nerve injury. Sci. Rep. 2016, 6, 26809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, T.Y. Characteristic Features and Contributory Factors in Fatal Ciguatera Fish Poisoning—Implications for Prevention and Public Education. Am. J. Trop. Med. Hyg. 2016, 94, 704–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Cao, B.; Yang, X.; Wu, J.; Chan, L.L.; Li, Y. Chronic ciguatoxin poisoning causes emotional and cognitive dysfunctions in rats. Toxicol. Res. 2017, 6, 179–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatti, C.M.I.; Lonati, D.; Darius, H.T.; Zancan, A.; Roue, M.; Schicchi, A.; Locatelli, C.A.; Chinain, M. Tectus niloticus (Tegulidae, Gastropod) as a Novel Vector of Ciguatera Poisoning: Clinical Characterization and Follow-Up of a Mass Poisoning Event in Nuku Hiva Island (French Polynesia). Toxins 2018, 10, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leite, I.D.P.; Sdiri, K.; Taylor, A.; Viallon, J.; Gharbia, H.B.; Junior, L.L.M.; Swarzenski, P.; Oberhaensli, F.; Darius, H.T.; Chinain, M.; et al. Experimental Evidence of Ciguatoxin Accumulation and Depuration in Carnivorous Lionfish. Toxins 2021, 13, 564. [Google Scholar] [CrossRef]
- Rhodes, L.L.; Smith, K.F.; Murray, J.S.; Nishimura, T.; Finch, S.C. Ciguatera Fish Poisoning: The Risk from an Aotearoa/New Zealand Perspective. Toxins 2020, 12, 50. [Google Scholar] [CrossRef] [Green Version]
- Zingone, A.; Escalera, L.; Aligizaki, K.; Fernandez-Tejedor, M.; Ismael, A.; Montresor, M.; Mozetic, P.; Tas, S.; Totti, C. Toxic marine microalgae and noxious blooms in the Mediterranean Sea: A contribution to the Global HAB Status Report. Harmful Algae 2021, 102, 101843. [Google Scholar] [CrossRef]
- Tudo, A.; Gaiani, G.; Varela, M.R.; Tsumuraya, T.; Andree, K.B.; Fernandez-Tejedor, M.; Campas, M.; Diogene, J. Further advance of Gambierdiscus Species in the Canary Islands, with the First Report of Gambierdiscus belizeanus. Toxins 2020, 12, 692. [Google Scholar] [CrossRef]
- Navarro Quiroz, R.; Herrera-Usuga, J.C.; Osorio-Ospina, L.M.; Garcia-Pertuz, K.M.; Navarro Quiroz, E. Epidemiology and Toxicology of Ciguatera Poisoning in the Colombian Caribbean. Mar. Drugs 2020, 18, 504. [Google Scholar] [CrossRef]
- Murata, M.; Yasumoto, T. The structure elucidation and biological activities of high molecular weight algal toxins: Maitotoxin, prymnesins and zooxanthellatoxins. Nat. Prod. Rep. 2000, 17, 293–314. [Google Scholar] [CrossRef]
- Estevez, P.; Castro, D.; Leao-Martins, J.M.; Sibat, M.; Tudo, A.; Dickey, R.; Diogene, J.; Hess, P.; Gago-Martinez, A. Toxicity Screening of a Gambierdiscus australes Strain from the Western Mediterranean Sea and Identification of a Novel Maitotoxin Analogue. Mar. Drugs 2021, 19, 460. [Google Scholar] [CrossRef] [PubMed]
- Munday, R.; Murray, S.; Rhodes, L.L.; Larsson, M.E.; Harwood, D.T. Ciguatoxins and Maitotoxins in Extracts of Sixteen Gambierdiscus Isolates and One Fukuyoa Isolate from the South Pacific and Their Toxicity to Mice by Intraperitoneal and Oral Administration. Mar. Drugs 2017, 15, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Litaker, R.W.; Vandersea, M.W.; Faust, M.A.; Kibler, S.R.; Nau, A.W.; Holland, W.C.; Chinain, M.; Holmes, M.J.; Tester, P.A. Global distribution of ciguatera causing dinoflagellates in the genus Gambierdiscus. Toxicon 2010, 56, 711–730. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Li, X.H.; Li, Y.J. Glutamate in peripheral organs: Biology and pharmacology. Eur. J. Pharmacol. 2016, 784, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Hansen, K.B.; Wollmuth, L.P.; Bowie, D.; Furukawa, H.; Menniti, F.S.; Sobolevsky, A.I.; Swanson, G.T.; Swanger, S.A.; Greger, I.H.; Nakagawa, T.; et al. Structure, Function, and Pharmacology of Glutamate Receptor Ion Channels. Pharmacol. Rev. 2021, 73, 298–487. [Google Scholar] [CrossRef] [PubMed]
- Stonik, V.A.; Stonik, I.V. Marine Excitatory Amino Acids: Structure, Properties, Biosynthesis and Recent Approaches to Their Syntheses. Molecules 2020, 25, 3049. [Google Scholar] [CrossRef] [PubMed]
- Todd, E.C.D. Domoic Acid and Amnesic Shellfish Poisoning—A Review. J. Food Prot. 1993, 56, 69–83. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion of the Panel on Contaminants in the Food Chain on a request from the European Commission on marine biotoxins in shellfish—Domoic acid. EFSA J. 2009, 1181, 1–61. [Google Scholar]
- Saeed, A.F.; Awan, S.A.; Ling, S.; Wang, R.; Wang, S. Domoic acid: Attributes, exposure risks, innovative detection techniques and therapeutics. Algal Res. 2017, 24, 97–110. [Google Scholar] [CrossRef]
- Collingridge, G.L.; Olsen, R.W.; Peters, J.; Spedding, M. A nomenclature for ligand-gated ion channels. Neuropharmacology 2009, 56, 2–5. [Google Scholar] [CrossRef] [Green Version]
- Mollerud, S.; Frydenvang, K.; Pickering, D.S.; Kastrup, J.S. Lessons from crystal structures of kainate receptors. Neuropharmacology 2017, 112, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Clark, B.L.M.; Menard, F. Kainic Acid-Based Agonists of Glutamate Receptors: SAR Analysis and Guidelines for Analog Design. ACS Chem. Neurosci. 2019, 10, 4190–4198. [Google Scholar] [CrossRef] [PubMed]
- Nanao, M.H.; Green, T.; Stern-Bach, Y.; Heinemann, S.F.; Choe, S. Structure of the kainate receptor subunit GluR6 agonist-binding domain complexed with domoic acid. Proc. Natl. Acad. Sci. USA 2005, 102, 1708–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hald, H.; Naur, P.; Pickering, D.S.; Sprogoe, D.; Madsen, U.; Timmermann, D.B.; Ahring, P.K.; Liljefors, T.; Schousboe, A.; Egebjerg, J.; et al. Partial agonism and antagonism of the ionotropic glutamate receptor iGLuR5: Structures of the ligand-binding core in complex with domoic acid and 2-amino-3-[5-tert-butyl-3-(phosphonomethoxy)-4-isoxazolyl]propionic acid. J. Biol. Chem. 2007, 282, 25726–25736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco, J.; Morono, A.; Arevalo, F.; Correa, J.; Salgado, C.; Rossignoli, A.E.; Lamas, J.P. Twenty-Five Years of Domoic Acid Monitoring in Galicia (NW Spain): Spatial, Temporal and Interspecific Variations. Toxins 2021, 13, 756. [Google Scholar] [CrossRef]
- Perl, T.M.; Bedard, L.; Kosatsky, T.; Hockin, J.C.; Todd, E.C.; McNutt, L.A.; Remis, R.S. Amnesic shellfish poisoning: A new clinical syndrome due to domoic acid. Can. Dis. Wkly. Rep. 1990, 16 (Suppl. 1E), 7–8. [Google Scholar]
- Perl, T.M.; Bedard, L.; Kosatsky, T.; Hockin, J.C.; Todd, E.C.; Remis, R.S. An outbreak of toxic encephalopathy caused by eating mussels contaminated with domoic acid. N. Engl. J. Med. 1990, 322, 1775–1780. [Google Scholar] [CrossRef]
- Perl, T.M.; Teitelbaum, J.; Hockin, J.; Todd, E.C. Domoic acid toxicity. Panel discussion: Definition of the syndrome. Can. Dis. Wkly. Rep. 1990, 16 (Suppl. 1E), 41–45. [Google Scholar]
- Kimura, O.; Kotaki, Y.; Hamaue, N.; Haraguchi, K.; Endo, T. Transcellular transport of domoic acid across intestinal Caco-2 cell monolayers. Food Chem. Toxicol. 2011, 49, 2167–2171. [Google Scholar] [CrossRef]
- Munday, R.; Holland, P.T.; McNabb, P.; Selwood, A.I.; Rhodes, L.L. Comparative toxicity to mice of domoic acid and isodomoic acids A, B and C. Toxicon 2008, 52, 954–956. [Google Scholar] [CrossRef]
- Jing, J.; Petroff, R.; Shum, S.; Crouthamel, B.; Topletz, A.R.; Grant, K.S.; Burbacher, T.M.; Isoherranen, N. Toxicokinetics and Physiologically Based Pharmacokinetic Modeling of the Shellfish Toxin Domoic Acid in Nonhuman Primates. Drug Metab. Dispos. 2018, 46, 155–165. [Google Scholar] [CrossRef]
- Truelove, J.; Mueller, R.; Pulido, O.; Martin, L.; Fernie, S.; Iverson, F. 30-day oral toxicity study of domoic acid in cynomolgus monkeys: Lack of overt toxicity at doses approaching the acute toxic dose. Nat. Toxins 1997, 5, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, K.A.; Kendrick, P.S.; Ladiges, W.; Hiolski, E.M.; Ferriss, B.E.; Smith, D.R.; Marcinek, D.J. Chronic low-level exposure to the common seafood toxin domoic acid causes cognitive deficits in mice. Harmful Algae 2017, 64, 20–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petroff, R.; Hendrix, A.; Shum, S.; Grant, K.S.; Lefebvre, K.A.; Burbacher, T.M. Public health risks associated with chronic, low-level domoic acid exposure: A review of the evidence. Pharmacol. Ther. 2021, 227, 107865. [Google Scholar] [CrossRef] [PubMed]
- Doucette, T.A.; Tasker, R.A. Perinatal Domoic Acid as a Neuroteratogen. Curr. Top. Behav. Neurosci. 2016, 29, 87–110. [Google Scholar] [PubMed]
- Lefebvre, K.A.; Quakenbush, L.; Frame, E.; Huntington, K.B.; Sheffield, G.; Stimmelmayr, R.; Bryan, A.; Kendrick, P.; Ziel, H.; Goldstein, T.; et al. Prevalence of algal toxins in Alaskan marine mammals foraging in a changing arctic and subarctic environment. Harmful Algae 2016, 55, 13–24. [Google Scholar] [CrossRef] [Green Version]
- McKibben, S.M.; Peterson, W.; Wood, A.M.; Trainer, V.L.; Hunter, M.; White, A.E. Climatic regulation of the neurotoxin domoic acid. Proc. Natl. Acad. Sci. USA 2017, 114, 239–244. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Qu, P.; Fu, F.; Tennenbaum, N.; Tatters, A.O.; Hutchins, D.A. Understanding the blob bloom: Warming increases toxicity and abundance of the harmful bloom diatom Pseudo-nitzschia in California coastal waters. Harmful Algae 2017, 67, 36–43. [Google Scholar] [CrossRef]
- Goldstein, T.; Mazet, J.A.; Zabka, T.S.; Langlois, G.; Colegrove, K.M.; Silver, M.; Bargu, S.; Van Dolah, F.; Leighfield, T.; Conrad, P.A.; et al. Novel symptomatology and changing epidemiology of domoic acid toxicosis in California sea lions (Zalophus californianus): An increasing risk to marine mammal health. Proc. Biol. Sci. 2008, 275, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Takai, A.; Murata, M.; Torigoe, K.; Isobe, M.; Mieskes, G.; Yasumoto, T. Inhibitory effect of okadaic acid derivatives on protein phosphatases. A study on structure-affinity relationship. Biochem. J. 1992, 284 Pt 2, 539–544. [Google Scholar] [CrossRef]
- Prickett, T.D.; Brautigan, D.L. The alpha4 regulatory subunit exerts opposing allosteric effects on protein phosphatases PP6 and PP2A. J. Biol. Chem. 2006, 281, 30503–30511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Twiner, M.J.; Doucette, G.J.; Pang, Y.; Fang, C.; Forsyth, C.J.; Miles, C.O. Structure-Activity Relationship Studies Using Natural and Synthetic Okadaic Acid/Dinophysistoxin Toxins. Mar. Drugs 2016, 14, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maynes, J.T.; Bateman, K.S.; Cherney, M.M.; Das, A.K.; Luu, H.A.; Holmes, C.F.; James, M.N. Crystal structure of the tumor-promoter okadaic acid bound to protein phosphatase-1. J. Biol. Chem. 2001, 276, 44078–44082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, Y.; Xu, Y.; Chen, Y.; Jeffrey, P.D.; Chao, Y.; Lin, Z.; Li, Z.; Strack, S.; Stock, J.B.; Shi, Y. Structure of protein phosphatase 2A core enzyme bound to tumor-inducing toxins. Cell 2006, 127, 341–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huhn, J.; Jeffrey, P.D.; Larsen, K.; Rundberget, T.; Rise, F.; Cox, N.R.; Arcus, V.; Shi, Y.; Miles, C.O. A structural basis for the reduced toxicity of dinophysistoxin-2. Chem. Res. Toxicol. 2009, 22, 1782–1786. [Google Scholar] [CrossRef]
- Yadav, L.; Tamene, F.; Goos, H.; van Drogen, A.; Katainen, R.; Aebersold, R.; Gstaiger, M.; Varjosalo, M. Systematic Analysis of Human Protein Phosphatase Interactions and Dynamics. Cell Syst. 2017, 4, 430–444.e5. [Google Scholar] [CrossRef] [Green Version]
- Yasumoto, T.; Murata, M.; Oshima, Y.; Matsumoto, G.K.; Clardy, J. Diarrhetic Shellfish Poisoning. In Seafood Toxins; Ragelis, E.P., Ed.; American Chemical Society: Washington, DC, USA, 1984; Volume 262, pp. 207–214. [Google Scholar]
- Rao, M.C. Physiology of Electrolyte Transport in the Gut: Implications for Disease. Compr. Physiol. 2019, 9, 947–1023. [Google Scholar]
- Louzao, M.C.; Fernandez, D.A.; Abal, P.; Fraga, M.; Vilarino, N.; Vieytes, M.R.; Botana, L.M. Diarrhetic effect of okadaic acid could be related with its neuronal action: Changes in neuropeptide Y. Toxicol. Lett. 2015, 237, 151–160. [Google Scholar] [CrossRef]
- Louzao, M.C.; Costas, C.; Abal, P.; Suzuki, T.; Watanabe, R.; Vilarino, N.; Carrera, C.; Boente-Juncal, A.; Vale, C.; Vieytes, M.R.; et al. Serotonin involvement in okadaic acid-induced diarrhoea in vivo. Arch. Toxicol. 2021, 95, 2797–2813. [Google Scholar] [CrossRef]
- Hagbom, M.; De Faria, F.M.; Winberg, M.E.; Westerberg, S.; Nordgren, J.; Sharma, S.; Keita, A.V.; Loitto, V.; Magnusson, K.E.; Svensson, L. Neurotrophic Factors Protect the Intestinal Barrier from Rotavirus Insult in Mice. mBio 2020, 11, e02834-19. [Google Scholar] [CrossRef] [Green Version]
- Reale, O.; Bodi, D.; Huguet, A.; Fessard, V. Role of enteric glial cells in the toxicity of phycotoxins: Investigation with a tri-culture intestinal cell model. Toxicol. Lett. 2021, 351, 89–98. [Google Scholar] [CrossRef] [PubMed]
- McCarron, P.; Kilcoyne, J.; Hess, P. Effects of cooking and heat treatment on concentration and tissue distribution of okadaic acid and dinophysistoxin-2 in mussels (Mytilus edulis). Toxicon 2008, 51, 1081–1089. [Google Scholar] [CrossRef] [PubMed]
- Valdiglesias, V.; Prego-Faraldo, M.V.; Pasaro, E.; Mendez, J.; Laffon, B. Okadaic acid: More than a diarrheic toxin. Mar. Drugs 2013, 11, 4328–4349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, A.D.; Lewis, A.M.; Bradley, K.; Maskrey, B.H. Marine invertebrate interactions with Harmful Algal Blooms—Implications for One Health. J. Invertebr. Pathol. 2021, 186, 107555. [Google Scholar] [CrossRef]
- Campos, A.; Freitas, M.; de Almeida, A.M.; Martins, J.C.; Dominguez-Perez, D.; Osorio, H.; Vasconcelos, V.; Costa, P.R. OMICs Approaches in Diarrhetic Shellfish Toxins Research. Toxins 2020, 12, 493. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.Y.; Lin, L.; Gao, Y.; Hong, H.S.; Wang, D.Z. Quantitative proteomic analysis of okadaic acid treated mouse small intestines reveals differentially expressed proteins involved in diarrhetic shellfish poisoning. J. Proteom. 2012, 75, 2038–2052. [Google Scholar] [CrossRef]
- Lago, J.; Santaclara, F.; Vieites, J.M.; Cabado, A.G. Collapse of mitochondrial membrane potential and caspases activation are early events in okadaic acid-treated Caco-2 cells. Toxicon 2005, 46, 579–586. [Google Scholar] [CrossRef]
- Valdiglesias, V.; Mendez, J.; Pasaro, E.; Cemeli, E.; Anderson, D.; Laffon, B. Assessment of okadaic acid effects on cytotoxicity, DNA damage and DNA repair in human cells. Mutat. Res. 2010, 689, 74–79. [Google Scholar] [CrossRef]
- Jiao, Y.H.; Dou, M.; Wang, G.; Li, H.Y.; Liu, J.S.; Yang, X.; Yang, W.D. Exposure of okadaic acid alters the angiogenesis in developing chick embryos. Toxicon 2017, 133, 74–81. [Google Scholar] [CrossRef]
- Martin-Lopez, A.; Gallardo-Rodriguez, J.J.; Sanchez-Miron, A.; Garcia-Camacho, F.; Molina-Grima, E. Cytotoxicity of yessotoxin and okadaic acid in mouse T lymphocyte cell line EL-4. Toxicon 2012, 60, 1049–1056. [Google Scholar] [CrossRef]
- Oliveira, J.M.; da Cruz, E.S.C.B.; Muller, T.; Martins, T.S.; Cova, M.; da Cruz, E.S.O.A.B.; Henriques, A.G. Toward Neuroproteomics in Biological Psychiatry: A Systems Approach Unravels Okadaic Acid-Induced Alterations in the Neuronal Phosphoproteome. OMICS 2017, 21, 550–563. [Google Scholar] [CrossRef] [PubMed]
- Espina, B.; Louzao, M.C.; Cagide, E.; Alfonso, A.; Vieytes, M.R.; Yasumoto, T.; Botana, L.M. The methyl ester of okadaic acid is more potent than okadaic acid in disrupting the actin cytoskeleton and metabolism of primary cultured hepatocytes. Br. J. Pharmacol. 2010, 159, 337–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubiolo, J.A.; Lopez-Alonso, H.; Vega, F.V.; Vieytes, M.R.; Botana, L.M. Comparative study of toxicological and cell cycle effects of okadaic acid and dinophysistoxin-2 in primary rat hepatocytes. Life Sci. 2012, 90, 416–423. [Google Scholar] [CrossRef]
- Ferreiro, S.F.; Carrera, C.; Vilarino, N.; Louzao, M.C.; Santamarina, G.; Cantalapiedra, A.G.; Botana, L.M. Acute cardiotoxicity evaluation of the marine biotoxins OA, DTX-1 and YTX. Toxins 2015, 7, 1030–1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Campo, M.; Zhong, T.Y.; Tampe, R.; Garcia, L.; Lagos, N. Sublethal doses of dinophysistoxin-1 and okadaic acid stimulate secretion of inflammatory factors on innate immune cells: Negative health consequences. Toxicon 2017, 126, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Kamat, P.K.; Rai, S.; Nath, C. Okadaic acid induced neurotoxicity: An emerging tool to study Alzheimer’s disease pathology. Neurotoxicology 2013, 37, 163–172. [Google Scholar] [CrossRef]
- Jiao, Y.; Wang, G.; Li, D.; Li, H.; Liu, J.; Yang, X.; Yang, W. Okadaic Acid Exposure Induced Neural Tube Defects in Chicken (Gallus gallus) Embryos. Mar. Drugs 2021, 19, 322. [Google Scholar] [CrossRef]
- Jimenez-Carcamo, D.; Garcia, C.; Contreras, H.R. Toxins of Okadaic Acid-Group Increase Malignant Properties in Cells of Colon Cancer. Toxins 2020, 12, 179. [Google Scholar] [CrossRef] [Green Version]
- Manerio, E.; Rodas, V.L.; Costas, E.; Hernandez, J.M. Shellfish consumption: A major risk factor for colorectal cancer. Med. Hypotheses 2008, 70, 409–412. [Google Scholar] [CrossRef]
- Liu, Y.; Zheng, J.W.; Peng, X.C.; Li, H.Y.; Huang, L.; Li, D.W.; Liu, J.S.; Yang, W.D. Changes in colonic microbiotas in rat after long-term exposure to low dose of okadaic acid. Chemosphere 2020, 254, 126874. [Google Scholar] [CrossRef]
- Emery, H.; Traves, W.; Rowley, A.F.; Coates, C.J. The diarrhetic shellfish-poisoning toxin, okadaic acid, provokes gastropathy, dysbiosis and susceptibility to bacterial infection in a non-rodent bioassay, Galleria mellonella. Arch. Toxicol. 2021, 95, 3361–3376. [Google Scholar] [CrossRef] [PubMed]
- Litvak, Y.; Byndloss, M.X.; Baumler, A.J. Colonocyte metabolism shapes the gut microbiota. Science 2018, 362, eaat9076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Ota, H.; Yamasaki, M. Direct evidence of transformation of dinophysistoxin-1 to 7-O-acyl-dinophysistoxin-1 (dinophysistoxin-3) in the scallop Patinopecten yessoensis. Toxicon 1999, 37, 187–198. [Google Scholar] [CrossRef]
- Solino, L.; Sureda, F.X.; Diogene, J. Evaluation of okadaic acid, dinophysistoxin-1 and dinophysistoxin-2 toxicity on Neuro-2a, NG108-15 and MCF-7 cell lines. Toxicol Vitr. 2015, 29, 59–62. [Google Scholar] [CrossRef]
- Suzuki, H.; Okada, Y. Comparative toxicity of dinophysistoxin-1 and okadaic acid in mice. J. Vet. Med. Sci. 2018, 80, 616–619. [Google Scholar] [CrossRef] [Green Version]
- EFSA. Marine biotoxins in shellfish- okadaic acid and analogues, Scientific Opinion of the Panel on Contaminants in the Food chain. EFSA J. 2008, 589, 1–62. [Google Scholar]
- Ikehara, T.; Chikanishi, K.; Oshiro, N. Specification of the Okadaic Acid Equivalent for Okadaic Acid, Dinophysistoxin-1, and Dinophysistoxin-2 Based on Protein Phosphatase 2A Inhibition and Cytotoxicity Assays Using Neuro 2A Cell Line. J. Mar. Sci. Eng. 2021, 9, 1140. [Google Scholar] [CrossRef]
- O’Mahony, M. EU Regulatory Risk Management of Marine Biotoxins in the Marine Bivalve Mollusc Food-Chain. Toxins 2018, 10, 118. [Google Scholar] [CrossRef] [Green Version]
- Blanco, J.; Arevalo, F.; Correa, J.; Morono, A. Lipophilic Toxins in Galicia (NW Spain) between 2014 and 2017: Incidence on the Main Molluscan Species and Analysis of the Monitoring Efficiency. Toxins 2019, 11, 612. [Google Scholar] [CrossRef] [Green Version]
- Ozawa, M.; Uchida, H.; Watanabe, R.; Matsushima, R.; Oikawa, H.; Takahashi, K.; Iwataki, M.; Suzuki, T. Complex profiles of azaspiracid analogues in two culture strains of Azadinium poporum (Amphidomataceae, Dinophyceae) isolated from Japanese coastal waters determined by LC-MS/MS. Toxicon 2021, 199, 145–155. [Google Scholar] [CrossRef]
- Reale, O.; Huguet, A.; Fessard, V. Novel Insights on the Toxicity of Phycotoxins on the Gut through the Targeting of Enteric Glial Cells. Mar. Drugs 2019, 17, 429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boente-Juncal, A.; Raposo-Garcia, S.; Costas, C.; Louzao, M.C.; Vale, C.; Botana, L.M. Partial Blockade of Human Voltage-Dependent Sodium Channels by the Marine Toxins Azaspiracids. Chem. Res. Toxicol. 2020, 33, 2593–2604. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.; Heo, J.; Han, J.; Hong, H.D.; Jeon, H.; Hwang, H.J.; Hong, C.Y.; Kim, D.; Han, J.W.; Baek, K. Industrial Applications of Dinoflagellate Phycotoxins Based on Their Modes of Action: A Review. Toxins 2020, 12, 805. [Google Scholar] [CrossRef] [PubMed]
- Twiner, M.J.; Doucette, G.J.; Rasky, A.; Huang, X.P.; Roth, B.L.; Sanguinetti, M.C. Marine algal toxin azaspiracid is an open-state blocker of hERG potassium channels. Chem. Res. Toxicol. 2012, 25, 1975–1984. [Google Scholar] [CrossRef] [PubMed]
- Ferreiro, S.F.; Vilarino, N.; Carrera, C.; Louzao, M.C.; Cantalapiedra, A.G.; Santamarina, G.; Cifuentes, J.M.; Vieira, A.C.; Botana, L.M. Subacute Cardiovascular Toxicity of the Marine Phycotoxin Azaspiracid-1 in Rats. Toxicol. Sci. 2016, 151, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Pelin, M.; Kilcoyne, J.; Nulty, C.; Crain, S.; Hess, P.; Tubaro, A.; Sosa, S. Toxic equivalency factors (TEFs) after acute oral exposure of azaspiracid 1, -2 and -3 in mice. Toxicol. Lett. 2018, 282, 136–146. [Google Scholar] [CrossRef]
- Pelin, M.; Kilcoyne, J.; Florio, C.; Hess, P.; Tubaro, A.; Sosa, S. Azaspiracids Increase Mitochondrial Dehydrogenases Activity in Hepatocytes: Involvement of Potassium and Chloride Ions. Mar. Drugs 2019, 17, 276. [Google Scholar] [CrossRef] [Green Version]
- James, K.J.; Fidalgo Saez, M.J.; Furey, A.; Lehane, M. Azaspiracid poisoning, the food-borne illness associated with shellfish consumption. Food Addit. Contam. 2004, 21, 879–892. [Google Scholar] [CrossRef]
- Kellmann, R.; Schaffner, C.A.; Gronset, T.A.; Satake, M.; Ziegler, M.; Fladmark, K.E. Proteomic response of human neuroblastoma cells to azaspiracid-1. J. Proteom. 2009, 72, 695–707. [Google Scholar] [CrossRef]
- Cao, Z.; LePage, K.T.; Frederick, M.O.; Nicolaou, K.C.; Murray, T.F. Involvement of caspase activation in azaspiracid-induced neurotoxicity in neocortical neurons. Toxicol. Sci. 2010, 114, 323–334. [Google Scholar] [CrossRef]
- Mudge, E.M.; Miles, C.O.; Hardstaff, W.R.; McCarron, P. Fatty acid esters of azaspiracids identified in mussels (Mytilus edulis) using liquid chromatography-high resolution mass spectrometry. Toxicon X 2020, 8, 100059. [Google Scholar] [CrossRef] [PubMed]
- Kilcoyne, J.; McCarron, P.; Twiner, M.J.; Rise, F.; Hess, P.; Wilkins, A.L.; Miles, C.O. Identification of 21,22-Dehydroazaspiracids in Mussels (Mytilus edulis) and in Vitro Toxicity of Azaspiracid-26. J. Nat. Prod. 2018, 81, 885–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aasen, J.A.; Espenes, A.; Hess, P.; Aune, T. Sub-lethal dosing of azaspiracid-1 in female NMRI mice. Toxicon 2010, 56, 1419–1425. [Google Scholar] [CrossRef]
- Aune, T.; Espenes, A.; Aasen, J.A.; Quilliam, M.A.; Hess, P.; Larsen, S. Study of possible combined toxic effects of azaspiracid-1 and okadaic acid in mice via the oral route. Toxicon 2012, 60, 895–906. [Google Scholar] [CrossRef]
- Twiner, M.J.; Rehmann, N.; Hess, P.; Doucette, G.J. Azaspiracid shellfish poisoning: A review on the chemistry, ecology, and toxicology with an emphasis on human health impacts. Mar. Drugs 2008, 6, 39–72. [Google Scholar] [CrossRef]
- Ito, E.; Satake, M.; Ofuji, K.; Kurita, N.; McMahon, T.; James, K.; Yasumoto, T. Multiple organ damage caused by a new toxin azaspiracid, isolated from mussels produced in Ireland. Toxicon 2000, 38, 917–930. [Google Scholar] [CrossRef]
- Ito, E.; Satake, M.; Ofuji, K.; Higashi, M.; Harigaya, K.; McMahon, T.; Yasumoto, T. Chronic effects in mice caused by oral administration of sublethal doses of azaspiracid, a new marine toxin isolated from mussels. Toxicon 2002, 40, 193–203. [Google Scholar] [CrossRef]
- Chevallier, O.P.; Graham, S.F.; Alonso, E.; Duffy, C.; Silke, J.; Campbell, K.; Botana, L.M.; Elliott, C.T. New insights into the causes of human illness due to consumption of azaspiracid contaminated shellfish. Sci. Rep. 2015, 5, 9818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, Y.; Qiu, J.; Xie, T.; McCarron, P.; Li, A. Accumulation and transformation of azaspiracids in scallops (Chlamys farreri) and mussels (Mytilus galloprovincialis) fed with Azadinium poporum, and response of antioxidant enzymes. Toxicon 2018, 143, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, M.E.; Accoroni, S.; Mezzelani, M.; Lugarini, F.; Bacchiocchi, S.; Siracusa, M.; Tavoloni, T.; Piersanti, A.; Totti, C.; Regoli, F.; et al. Biological Effects of the Azaspiracid-Producing Dinoflagellate Azadinium dexteroporum in Mytilus galloprovincialis from the Mediterranean Sea. Mar. Drugs 2019, 17, 595. [Google Scholar] [CrossRef] [Green Version]
- Colman, J.R.; Twiner, M.J.; Hess, P.; McMahon, T.; Satake, M.; Yasumoto, T.; Doucette, G.J.; Ramsdell, J.S. Teratogenic effects of azaspiracid-1 identified by microinjection of Japanese medaka (Oryzias latipes) embryos. Toxicon 2005, 45, 881–890. [Google Scholar] [CrossRef]
- Allingham, J.S.; Miles, C.O.; Rayment, I. A structural basis for regulation of actin polymerization by pectenotoxins. J. Mol. Biol. 2007, 371, 959–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espina, B.; Louzao, M.C.; Ares, I.R.; Cagide, E.; Vieytes, M.R.; Vega, F.V.; Rubiolo, J.A.; Miles, C.O.; Suzuki, T.; Yasumoto, T.; et al. Cytoskeletal toxicity of pectenotoxins in hepatic cells. Br. J. Pharmacol. 2008, 155, 934–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgess, V.; Shaw, G. Pectenotoxins—An issue for public health: A review of their comparative toxicology and metabolism. Environ. Int. 2001, 27, 275–283. [Google Scholar] [CrossRef]
- Tubaro, A.; Dell’ovo, V.; Sosa, S.; Florio, C. Yessotoxins: A toxicological overview. Toxicon 2010, 56, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Alfonso, A.; Vieytes, M.R.; Botana, L.M. Yessotoxin, a Promising Therapeutic Tool. Mar. Drugs 2016, 14, 30. [Google Scholar] [CrossRef] [Green Version]
- Korsnes, M.S.; Espenes, A. Yessotoxin as an apoptotic inducer. Toxicon 2011, 57, 947–958. [Google Scholar] [CrossRef]
- Rubiolo, J.A.; Lopez-Alonso, H.; Martinez, P.; Millan, A.; Cagide, E.; Vieytes, M.R.; Vega, F.V.; Botana, L.M. Yessotoxin induces ER-stress followed by autophagic cell death in glioma cells mediated by mTOR and BNIP3. Cell Signal. 2014, 26, 419–432. [Google Scholar] [CrossRef]
- Tobio, A.; Alfonso, A.; Madera-Salcedo, I.; Botana, L.M.; Blank, U. Yessotoxin, a Marine Toxin, Exhibits Anti-Allergic and Anti-Tumoural Activities Inhibiting Melanoma Tumour Growth in a Preclinical Model. PLoS ONE 2016, 11, e0167572. [Google Scholar] [CrossRef]
- Alonso, E.; Vale, C.; Vieytes, M.R.; Botana, L.M. Translocation of PKC by yessotoxin in an in vitro model of Alzheimer’s disease with improvement of tau and beta-amyloid pathology. ACS Chem. Neurosci. 2013, 4, 1062–1070. [Google Scholar] [CrossRef] [Green Version]
- Franchini, A.; Malagoli, D.; Ottaviani, E. Targets and effects of yessotoxin, okadaic acid and palytoxin: A differential review. Mar. Drugs 2010, 8, 658–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aune, T.; Sorby, R.; Yasumoto, T.; Ramstad, H.; Landsverk, T. Comparison of oral and intraperitoneal toxicity of yessotoxin towards mice. Toxicon 2002, 40, 77–82. [Google Scholar] [CrossRef]
- EU. Commission Regulation (EU) No 786/2013 of 16 August 2013 amending Annex III to Regulation (EC) No 853/2004 of the European Parliament and of the Council as regards the permitted limits of yessotoxins in live bivalve molluscs. Off. J. Eur. Union 2013, L220, 14. [Google Scholar]
- Davidson, K.; Baker, C.; Higgins, C.; Higman, W.; Swan, S.; Veszelovszki, A.; Turner, A.D. Potential Threats Posed by New or Emerging Marine Biotoxins in UK Waters and Examination of Detection Methodologies Used for Their Control: Cyclic Imines. Mar. Drugs 2015, 13, 7087–7112. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific Opinion on marine biotoxins in shellfish—Cyclic imines (spirolides, gymnodimines, pinnatoxins and pteriatoxins). EFSA J. 2010, 8, 1628. [Google Scholar]
- Amzil, Z.; Sibat, M.; Royer, F.; Masson, N.; Abadie, E. Report on the first detection of pectenotoxin-2, spirolide-a and their derivatives in French shellfish. Mar. Drugs 2007, 5, 168–179. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, A.; Morrell, N.; Turner, A.D.; Maskrey, B.H. Method performance verification for the combined detection and quantitation of the marine neurotoxins cyclic imines and brevetoxin shellfish metabolites in mussels (Mytilus edulis) and oysters (Crassostrea gigas) by UHPLC-MS/MS. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2021, 1179, 122864. [Google Scholar] [CrossRef] [PubMed]
- Araoz, R.; Servent, D.; Molgo, J.; Iorga, B.I.; Fruchart-Gaillard, C.; Benoit, E.; Gu, Z.; Stivala, C.; Zakarian, A. Total synthesis of pinnatoxins A and G and revision of the mode of action of pinnatoxin A. J. Am. Chem. Soc. 2011, 133, 10499–10511. [Google Scholar] [CrossRef] [Green Version]
- Delcourt, N.; Lagrange, E.; Abadie, E.; Fessard, V.; Fremy, J.M.; Vernoux, J.P.; Peyrat, M.B.; Maignien, T.; Arnich, N.; Molgo, J.; et al. Pinnatoxins’ Deleterious Effects on Cholinergic Networks: From Experimental Models to Human Health. Mar. Drugs 2019, 17, 425. [Google Scholar] [CrossRef] [Green Version]
- Bourne, Y.; Sulzenbacher, G.; Radic, Z.; Araoz, R.; Reynaud, M.; Benoit, E.; Zakarian, A.; Servent, D.; Molgo, J.; Taylor, P.; et al. Marine Macrocyclic Imines, Pinnatoxins A and G: Structural Determinants and Functional Properties to Distinguish Neuronal alpha7 from Muscle alpha1(2)betagammadelta nAChRs. Structure 2015, 23, 1106–1115. [Google Scholar] [CrossRef] [Green Version]
- Sosa, S.; Pelin, M.; Cavion, F.; Herve, F.; Hess, P.; Tubaro, A. Acute Oral Toxicity of Pinnatoxin G in Mice. Toxins 2020, 12, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benoit, E.; Couesnon, A.; Lindovsky, J.; Iorga, B.I.; Araoz, R.; Servent, D.; Zakarian, A.; Molgo, J. Synthetic Pinnatoxins A and G Reversibly Block Mouse Skeletal Neuromuscular Transmission In Vivo and In Vitro. Mar. Drugs 2019, 17, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munday, R.; Quilliam, M.A.; LeBlanc, P.; Lewis, N.; Gallant, P.; Sperker, S.A.; Ewart, H.S.; MacKinnon, S.L. Investigations into the toxicology of spirolides, a group of marine phycotoxins. Toxins 2012, 4, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hess, P.; Abadie, E.; Herve, F.; Berteaux, T.; Sechet, V.; Araoz, R.; Molgo, J.; Zakarian, A.; Sibat, M.; Rundberget, T.; et al. Pinnatoxin G is responsible for atypical toxicity in mussels (Mytilus galloprovincialis) and clams (Venerupis decussata) from Ingril, a French Mediterranean lagoon. Toxicon 2013, 75, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Altares, M.; Casanova, A.; Bane, V.; Diogene, J.; Furey, A.; de la Iglesia, P. Confirmation of pinnatoxins and spirolides in shellfish and passive samplers from Catalonia (Spain) by liquid chromatography coupled with triple quadrupole and high-resolution hybrid tandem mass spectrometry. Mar. Drugs 2014, 12, 3706–3732. [Google Scholar] [CrossRef]
- Arnich, N.; Abadie, E.; Delcourt, N.; Fessard, V.; Fremy, J.M.; Hort, V.; Lagrange, E.; Maignien, T.; Molgo, J.; Peyrat, M.B.; et al. Health risk assessment related to pinnatoxins in French shellfish. Toxicon 2020, 180, 1–10. [Google Scholar] [CrossRef]
- Servent, D.; Malgorn, C.; Bernes, M.; Gil, S.; Simasotchi, C.; Herard, A.S.; Delzescaux, T.; Thai, R.; Barbe, P.; Keck, M.; et al. First evidence that emerging pinnatoxin-G, a contaminant of shellfish, reaches the brain and crosses the placental barrier. Sci. Total Environ. 2021, 790, 148125. [Google Scholar] [CrossRef]
- ANSES. Opinion of the French Agency for Food, Environmental and Occupational Health & Safety on the Assessment of the Health Risks Associated with Pinnatoxins in Shellfish; ANSES: Maisons-Alfort, France, 2019; pp. 1–23. [Google Scholar]
- Patocka, J.; Nepovimova, E.; Wu, Q.; Kuca, K. Palytoxin congeners. Arch. Toxicol. 2018, 92, 143–156. [Google Scholar] [CrossRef]
- Rossini, G.P.; Bigiani, A. Palytoxin action on the Na(+),K(+)-ATPase and the disruption of ion equilibria in biological systems. Toxicon 2011, 57, 429–439. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Benedettini, G.; Onorari, M.; Serena, F.; Battocchi, C.; et al. First finding of Ostreopsis cf. ovata toxins in marine aerosols. Environ. Sci. Technol. 2014, 48, 3532–3540. [Google Scholar] [CrossRef]
- Tichadou, L.; Glaizal, M.; Armengaud, A.; Grossel, H.; Lemee, R.; Kantin, R.; Lasalle, J.L.; Drouet, G.; Rambaud, L.; Malfait, P.; et al. Health impact of unicellular algae of the Ostreopsis genus blooms in the Mediterranean Sea: Experience of the French Mediterranean coast surveillance network from 2006 to 2009. Clin. Toxicol. 2010, 48, 839–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelin, M.; Forino, M.; Brovedani, V.; Tartaglione, L.; Dell’Aversano, C.; Pistocchi, R.; Poli, M.; Sosa, S.; Florio, C.; Ciminiello, P.; et al. Ovatoxin-a, A Palytoxin Analogue Isolated from Ostreopsis cf. ovata Fukuyo: Cytotoxic Activity and ELISA Detection. Environ. Sci. Technol. 2016, 50, 1544–1551. [Google Scholar] [CrossRef] [PubMed]
- Ramos, V.; Vasconcelos, V. Palytoxin and analogs: Biological and ecological effects. Mar. Drugs 2010, 8, 2021–2037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tartaglione, L.; Dell’Aversano, C.; Mazzeo, A.; Forino, M.; Wieringa, A.; Ciminiello, P. Determination of Palytoxins in Soft Coral and Seawater from a Home Aquarium. Comparison between Palythoa- and Ostreopsis-Related Inhalatory Poisonings. Environ. Sci. Technol. 2016, 50, 1023–1030. [Google Scholar] [CrossRef] [Green Version]
- Nordt, S.P.; Wu, J.; Zahller, S.; Clark, R.F.; Cantrell, F.L. Palytoxin poisoning after dermal contact with zoanthid coral. J. Emerg. Med. 2011, 40, 397–399. [Google Scholar] [CrossRef]
- Vale, C.; Nicolaou, K.C.; Frederick, M.O.; Gomez-Limia, B.; Alfonso, A.; Vieytes, M.R.; Botana, L.M. Effects of azaspiracid-1, a potent cytotoxic agent, on primary neuronal cultures. A structure-activity relationship study. J. Med. Chem. 2007, 50, 356–363. [Google Scholar] [CrossRef]
- Valverde, I.; Lago, J.; Vieites, J.M.; Cabado, A.G. In vitro approaches to evaluate palytoxin-induced toxicity and cell death in intestinal cells. J. Appl. Toxicol. 2008, 28, 294–302. [Google Scholar] [CrossRef]
- EFSA. Scientific opinion on marine biotoxins in shellfish-palytoxin group. EFSA J. 2009, 1393, 1–38. [Google Scholar]
- Aligizaki, K.; Katikou, P.; Milandri, A.; Diogene, J. Occurrence of palytoxin-group toxins in seafood and future strategies to complement the present state of the art. Toxicon 2011, 57, 390–399. [Google Scholar] [CrossRef]
- Sosa, S.; Del Favero, G.; De Bortoli, M.; Vita, F.; Soranzo, M.R.; Beltramo, D.; Ardizzone, M.; Tubaro, A. Palytoxin toxicity after acute oral administration in mice. Toxicol. Lett. 2009, 191, 253–259. [Google Scholar] [CrossRef]
- Del Favero, G.; Sosa, S.; Poli, M.; Tubaro, A.; Sbaizero, O.; Lorenzon, P. In vivo and in vitro effects of 42-hydroxy-palytoxin on mouse skeletal muscle: Structural and functional impairment. Toxicol. Lett. 2014, 225, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Del Favero, G.; Beltramo, D.; Sciancalepore, M.; Lorenzon, P.; Coslovich, T.; Poli, M.; Testai, E.; Sosa, S.; Tubaro, A. Toxicity of palytoxin after repeated oral exposure in mice and in vitro effects on cardiomyocytes. Toxicon 2013, 75, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, H.; Suganuma, M.; Nakayasu, M.; Hakii, H.; Horiuchi, T.; Takayama, S.; Sugimura, T. Palytoxin is a non-12-O-tetradecanoylphorbol-13-acetate type tumor promoter in two-stage mouse skin carcinogenesis. Carcinogenesis 1986, 7, 707–710. [Google Scholar] [CrossRef] [PubMed]
- Wieringa, A.; Bertholee, D.; Ter Horst, P.; van den Brand, I.; Haringman, J.; Ciminiello, P. Respiratory impairment in four patients associated with exposure to palytoxin containing coral. Clin. Toxicol. 2014, 52, 150–151. [Google Scholar] [CrossRef]
- Bire, R.; Trotereau, S.; Lemee, R.; Oregioni, D.; Delpont, C.; Krys, S.; Guerin, T. Hunt for Palytoxins in a Wide Variety of Marine Organisms Harvested in 2010 on the French Mediterranean Coast. Mar. Drugs 2015, 13, 5425–5446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, B.; Huang, Y.; Baumann, K.; Fry, B.G.; Shi, Q. From Marine Venoms to Drugs: Efficiently Supported by a Combination of Transcriptomics and Proteomics. Mar. Drugs 2017, 15, 103. [Google Scholar] [CrossRef] [PubMed]
- Lindequist, U. Marine-Derived Pharmaceuticals—Challenges and Opportunities. Biomol. Ther. 2016, 24, 561–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catterall, W.A.; Goldin, A.L.; Waxman, S.G. International Union of Pharmacology. XLVII. Nomenclature and structure-function relationships of voltage-gated sodium channels. Pharmacol. Rev. 2005, 57, 397–409. [Google Scholar] [CrossRef]
- Alvarez, P.; Levine, J.D. Antihyperalgesic effect of tetrodotoxin in rat models of persistent muscle pain. Neuroscience 2015, 311, 499–507. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.J.; Lue, J.H.; Lin, L.H.; Huang, C.T.; Chiang, R.P.; Chen, C.L.; Tsai, Y.J. Effects of pre-emptive drug treatment on astrocyte activation in the cuneate nucleus following rat median nerve injury. Pain 2010, 148, 158–166. [Google Scholar] [CrossRef]
- Lyu, Y.S.; Park, S.K.; Chung, K.; Chung, J.M. Low dose of tetrodotoxin reduces neuropathic pain behaviors in an animal model. Brain Res. 2000, 871, 98–103. [Google Scholar] [CrossRef]
- Marcil, J.; Walczak, J.S.; Guindon, J.; Ngoc, A.H.; Lu, S.; Beaulieu, P. Antinociceptive effects of tetrodotoxin (TTX) in rodents. Br. J. Anaesth. 2006, 96, 761–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldlust, S.A.; Kavoosi, M.; Nezzer, J.; Kavoosi, M.; Korz, W.; Deck, K. Tetrodotoxin for Chemotherapy-Induced Neuropathic Pain: A Randomized, Double-Blind, Placebo-Controlled, Parallel-Dose Finding Trial. Toxins 2021, 13, 235. [Google Scholar] [CrossRef] [PubMed]
- Alguacil, L.F.; Perez-Garcia, C.; Salas, E.; Gonzalez-Martin, C.; Castillo, C.; Polanco, M.J.; Herradon, G.; Morales, L. Subcutaneous tetrodotoxin and inflammatory pain. Br. J. Anaesth. 2008, 100, 729–730. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.; Strong, J.A.; Meij, J.T.A.; Zhang, J.M.; Yu, L. Neuropathic pain: Early spontaneous afferent activity is the trigger. Pain 2005, 116, 243–256. [Google Scholar] [CrossRef]
- Hagen, N.A.; Cantin, L.; Constant, J.; Haller, T.; Blaise, G.; Ong-Lam, M.; du Souich, P.; Korz, W.; Lapointe, B. Tetrodotoxin for Moderate to Severe Cancer-Related Pain: A Multicentre, Randomized, Double-Blind, Placebo-Controlled, Parallel-Design Trial. Pain Res. Manag. 2017, 2017, 7212713. [Google Scholar] [CrossRef]
- Hagen, N.A.; Fisher, K.M.; Lapointe, B.; du Souich, P.; Chary, S.; Moulin, D.; Sellers, E.; Ngoc, A.H.; Canadian Tetrodotoxin Study Group. An open-label, multi-dose efficacy and safety study of intramuscular tetrodotoxin in patients with severe cancer-related pain. J. Pain Symptom Manag. 2007, 34, 171–182. [Google Scholar] [CrossRef]
- Hagen, N.A.; Lapointe, B.; Ong-Lam, M.; Dubuc, B.; Walde, D.; Gagnon, B.; Love, R.; Goel, R.; Hawley, P.; Ngoc, A.H.; et al. A multicentre open-label safety and efficacy study of tetrodotoxin for cancer pain. Curr. Oncol. 2011, 18, e109–e116. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Liu, A.; Santamaria, C.M.; Shomorony, A.; Ji, T.; Wei, T.; Gordon, A.; Elofsson, H.; Mehta, M.; Yang, R.; et al. Polymer-tetrodotoxin conjugates to induce prolonged duration local anesthesia with minimal toxicity. Nat. Commun. 2019, 10, 2566. [Google Scholar] [CrossRef]
- Adams, H.J.; Blair, M.R., Jr.; Takman, B.H. The local anesthetic activity of saxitoxin alone and with vasoconstrictor and local anesthetic agents. Arch. Int. Pharmacodyn. Ther. 1976, 224, 275–282. [Google Scholar]
- Berde, C.B.; Athiraman, U.; Yahalom, B.; Zurakowski, D.; Corfas, G.; Bognet, C. Tetrodotoxin-bupivacaine-epinephrine combinations for prolonged local anesthesia. Mar. Drugs 2011, 9, 2717–2728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Santamaria, C.M.; Wei, T.; Zhao, C.; Ji, T.; Yang, T.; Shomorony, A.; Wang, B.Y.; Kohane, D.S. Hollow Silica Nanoparticles Penetrate the Peripheral Nerve and Enhance the Nerve Blockade from Tetrodotoxin. Nano Lett. 2018, 18, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Lago, J.; Rodriguez, L.P.; Blanco, L.; Vieites, J.M.; Cabado, A.G. Tetrodotoxin, an Extremely Potent Marine Neurotoxin: Distribution, Toxicity, Origin and Therapeutical Uses. Mar. Drugs 2015, 13, 6384–6406. [Google Scholar] [CrossRef] [PubMed]
- Kohane, D.S.; Lu, N.T.; Gokgol-Kline, A.C.; Shubina, M.; Kuang, Y.; Hall, S.; Strichartz, G.R.; Berde, C.B. The local anesthetic properties and toxicity of saxitonin homologues for rat sciatic nerve block in vivo. Reg. Anesth. Pain Med. 2000, 25, 52–59. [Google Scholar] [CrossRef]
- Rodriguez-Navarro, A.J.; Lagos, M.; Figueroa, C.; Garcia, C.; Recabal, P.; Silva, P.; Iglesias, V.; Lagos, N. Potentiation of local anesthetic activity of neosaxitoxin with bupivacaine or epinephrine: Development of a long-acting pain blocker. Neurotox. Res. 2009, 16, 408–415. [Google Scholar] [CrossRef]
- Rodriguez-Navarro, A.J.; Lagos, N.; Lagos, M.; Braghetto, I.; Csendes, A.; Hamilton, J.; Figueroa, C.; Truan, D.; Garcia, C.; Rojas, A.; et al. Neosaxitoxin as a local anesthetic: Preliminary observations from a first human trial. Anesthesiology 2007, 106, 339–345. [Google Scholar] [CrossRef]
- Berde, C.; Kohane, D.S. Combined Formulations of Neosaxitoxin for Prolonged Local Anesthesia. Spain Patent ES 2,727,292 T3, 17 March 2014. [Google Scholar]
- Kamat, P.K.; Rai, S.; Swarnkar, S.; Shukla, R.; Nath, C. Molecular and cellular mechanism of okadaic acid (OKA)-induced neurotoxicity: A novel tool for Alzheimer’s disease therapeutic application. Mol. Neurobiol. 2014, 50, 852–865. [Google Scholar] [CrossRef]
- Medina, M.; Avila, J.; Villanueva, N. Use of okadaic acid to identify relevant phosphoepitopes in pathology: A focus on neurodegeneration. Mar. Drugs 2013, 11, 1656–1668. [Google Scholar] [CrossRef] [Green Version]
- Nagai, H.; Satake, M.; Yasumoto, T. Antimicrobial activities of polyether compounds of dinoflagellate origins. J. Appl. Phycol. 1990, 2, 305–308. [Google Scholar] [CrossRef]
- Botana, L.M.; Alonso, E.; Vale, C. Use of Yessotoxin and Analogs and Derivatives thereof for Treating and/or Preventing Neurodegenerative Diseases Linked to Tau and Beta Amyloid. Patent WO 2,011,095,668 A1, 8 August 2011. [Google Scholar]
- Botana, L.M.; Alfonso, A.; Rodriguez-Vieytes, M.; Loza Garcia, M.I. Therapeutic use of Yessotoxin as Human Tumor Cell Growth Inhibitor. Patent EP 1,875,906 B1, 29 June 2011. [Google Scholar]
- Fernandez-Araujo, A.; Sanchez, J.A.; Alfonso, A.; Vieytes, M.R.; Botana, L.M. Different toxic effects of YTX in tumor K-562 and lymphoblastoid cell lines. Front. Pharmacol. 2015, 6, 124. [Google Scholar] [CrossRef] [Green Version]
- Korsnes, M.S.; Korsnes, R. Lifetime Distributions from Tracking Individual BC3H1 Cells Subjected to Yessotoxin. Front. Bioeng. Biotechnol. 2015, 3, 166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Haro, L.; Schmitt, C.; Sinno-Tellier, S.; Paret, N.; Boels, D.; Le Roux, G.; Langrand, J.; Delcourt, N.; Labadie, M.; Simon, N. Ciguatera fish poisoning in France: Experience of the French Poison Control Centre Network from 2012 to 2019. Clin. Toxicol. 2021, 59, 252–255. [Google Scholar] [CrossRef] [PubMed]
- Legrand, A.M.; Bagnis, R. Effects of highly purified maitotoxin extracted from dinoflagellate Gambierdiscus toxicus on action potential of isolated rat heart. J. Mol. Cell. Cardiol. 1984, 16, 663–666. [Google Scholar] [CrossRef]
- Murata, M.; Naoki, H.; Iwashita, T.; Matsunaga, S.; Sasaki, M.; Yokoyama, A.; Yasumoto, T. Structure of maitotoxin. J. Am. Chem. Soc. 1993, 115, 2060–2062. [Google Scholar] [CrossRef]
- Reyes, J.G.; Sánchez-Cárdenas, C.; Acevedo-Castillo, W.; Leyton, P.L.-G.; Ignacio Felix, R.; Gandini, M.A.; Treviño, M.B.; Treviño, C.L. Maitotoxin: An Enigmatic Toxic Molecule with Useful Applications in the Biomedical Sciences. In Seafood and Freshwater Toxins, 3rd ed.; Botana, L.M., Ed.; Taylor and Francis Group: Boca Raton, FL, USA, 2014; pp. 677–694. [Google Scholar]
- Yokoyama, A.; Murata, M.; Oshima, Y.; Iwashita, T.; Yasumoto, T. Some chemical properties of maitotoxin, a putative calcium channel agonist isolated from a marine dinoflagellate. J. Biochem. 1988, 104, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Taglialatela, M.; Amoroso, S.; Yasumoto, T.; Di Renzo, G.; Annunziato, L. Maitotoxin and Bay-K-8644: Two putative calcium channel activators with different effects on endogenous dopamine release from tuberoinfundibular neurons. Brain Res. 1986, 381, 356–358. [Google Scholar] [CrossRef]
- Estacion, M.; Schilling, W.P. Maitotoxin-induced membrane blebbing and cell death in bovine aortic endothelial cells. BMC Physiol. 2001, 1, 2. [Google Scholar] [CrossRef]
- Chavez, J.C.; de Blas, G.A.; de la Vega-Beltran, J.L.; Nishigaki, T.; Chirinos, M.; Gonzalez-Gonzalez, M.E.; Larrea, F.; Solis, A.; Darszon, A.; Trevino, C.L. The opening of maitotoxin-sensitive calcium channels induces the acrosome reaction in human spermatozoa: Differences from the zona pellucida. Asian J. Androl. 2011, 13, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Leech, C.A.; Habener, J.F. Insulinotropic glucagon-like peptide-1-mediated activation of non-selective cation currents in insulinoma cells is mimicked by maitotoxin. J. Biol. Chem. 1997, 272, 17987–17993. [Google Scholar] [CrossRef] [Green Version]
- Soergel, D.G.; Gusovsky, F.; Yasumoto, T.; Daly, J.W. Stimulatory effects of maitotoxin on insulin release in insulinoma HIT cells: Role of calcium uptake and phosphoinositide breakdown. J. Pharmacol. Exp. Ther. 1990, 255, 1360–1365. [Google Scholar]
- Alonso, E.; Otero, P.; Vale, C.; Alfonso, A.; Antelo, A.; Gimenez-Llort, L.; Chabaud, L.; Guillou, C.; Botana, L.M. Benefit of 13-desmethyl spirolide C treatment in triple transgenic mouse model of Alzheimer disease: Beta-amyloid and neuronal markers improvement. Curr. Alzheimer Res. 2013, 10, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Alonso, E.; Vale, C.; Vieytes, M.R.; Laferla, F.M.; Gimenez-Llort, L.; Botana, L.M. 13-Desmethyl spirolide-C is neuroprotective and reduces intracellular Abeta and hyperphosphorylated tau in vitro. Neurochem. Int. 2011, 59, 1056–1065. [Google Scholar] [CrossRef] [PubMed]
- Boente-Juncal, A.; Mendez, A.G.; Vale, C.; Vieytes, M.R.; Botana, L.M. In Vitro Effects of Chronic Spirolide Treatment on Human Neuronal Stem Cell Differentiation and Cholinergic System Development. ACS Chem. Neurosci. 2018, 9, 1441–1452. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Wang, X.; Shi, C. Volume-regulated anion channel as a novel cancer therapeutic target. Int. J. Biol. Macromol. 2020, 159, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Beekwilder, J.P.; O’Leary, M.E.; van den Broek, L.P.; van Kempen, G.T.; Ypey, D.L.; van den Berg, R.J. Kv1.1 channels of dorsal root ganglion neurons are inhibited by n-butyl-p-aminobenzoate, a promising anesthetic for the treatment of chronic pain. J. Pharmacol. Exp. Ther. 2003, 304, 531–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Judge, S.I.; Bever, C.T., Jr. Potassium channel blockers in multiple sclerosis: Neuronal Kv channels and effects of symptomatic treatment. Pharmacol. Ther. 2006, 111, 224–259. [Google Scholar] [CrossRef]
- Varga, Z.; Hajdu, P.; Panyi, G. Ion channels in T lymphocytes: An update on facts, mechanisms and therapeutic targeting in autoimmune diseases. Immunol. Lett. 2010, 130, 19–25. [Google Scholar] [CrossRef]
- Wulff, H.; Beeton, C.; Chandy, K.G. Potassium channels as therapeutic targets for autoimmune disorders. Curr. Opin. Drug Discov. Devel. 2003, 6, 640–647. [Google Scholar]
- Beeton, C.; Wulff, H.; Standifer, N.E.; Azam, P.; Mullen, K.M.; Pennington, M.W.; Kolski-Andreaco, A.; Wei, E.; Grino, A.; Counts, D.R.; et al. Kv1.3 channels are a therapeutic target for T cell-mediated autoimmune diseases. Proc. Natl. Acad. Sci. USA 2006, 103, 17414–17419. [Google Scholar] [CrossRef] [Green Version]
- Shieh, C.C.; Coghlan, M.; Sullivan, J.P.; Gopalakrishnan, M. Potassium channels: Molecular defects, diseases, and therapeutic opportunities. Pharmacol. Rev. 2000, 52, 557–594. [Google Scholar]
- Alonso, E.; Fuwa, H.; Vale, C.; Suga, Y.; Goto, T.; Konno, Y.; Sasaki, M.; LaFerla, F.M.; Vieytes, M.R.; Giménez-Llort, L.; et al. Design and synthesis of skeletal analogues of gambierol: Attenuation of amyloid-β and tau pathology with voltage-gated potassium channel and N-methyl-D-aspartate receptor implications. J. Am. Chem. Soc. 2012, 134, 7467–7479. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, A.; Ishii, D.N.; Backx, P.H.; Pulford, B.E.; Birks, B.R.; Tamkun, M.M. Altered K(+) channel gene expression in diabetic rat ventricle: Isoform switching between Kv4.2 and Kv1.4. Am. J. Physiol. Heart Circ. Physiol. 2001, 281, H1800-7. [Google Scholar] [CrossRef] [PubMed]
- Bertaso, F.; Sharpe, C.C.; Hendry, B.M.; James, A.F. Expression of voltage-gated K+ channels in human atrium. Basic Res. Cardiol. 2002, 97, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Dom, I.; Bire, R.; Hort, V.; Lavison-Bompard, G.; Nicolas, M.; Guerin, T. Extended Targeted and Non-Targeted Strategies for the Analysis of Marine Toxins in Mussels and Oysters by (LC-HRMS). Toxins 2018, 10, 375. [Google Scholar] [CrossRef] [Green Version]
- AOAC Official Method 991. 26-1999, Domoic acid in mussels, liquid chromatographic method. In Official Methods of Analysis of AOAC International; AOAC International: Gaithersburg, MD, USA, 2000. [Google Scholar]
- AOAC. AOAC Official method 2005.06. Quantitative determination of Paralytic Shellfish Poisoning Toxins in shellfish using pre-chromatographic oxidation and liquid chromatography with fluorescence detection. In Official Methods of Analysis of AOAC International, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2006. [Google Scholar]
- AOAC. AOAC Official Method 2011.02. Paralytic Shellfish Toxins in Mussels, Clams, Oysters, and Scallops. Post-Column Oxidation (PCOX) Method. In Official Methods of Analysis of AOAC International, 19th ed.; AOAC International: Gaithersburg, MD, USA, 2012; Available online: http://www.aesan.gob.es/AECOSAN/docs/documentos/laboratorios/LNRBM/ARCHIVO2EU-Harmonised-SOP-LIPO-LCMSMS_Version5.pdf (accessed on 4 March 2022).
- Rey, V.; Botana, A.M.; Alvarez, M.; Antelo, A.; Botana, L.M. Liquid Chromatography with a Fluorimetric Detection Method for Analysis of Paralytic Shellfish Toxins and Tetrodotoxin Based on a Porous Graphitic Carbon Column. Toxins 2016, 8, 196. [Google Scholar] [CrossRef]
- CRLMB. EU-Harmonised Standard Operating Procedure for Determination of Lipophilic Marine Biotoxins in Molluscs by LC-MS/MS. 2015. Available online: http://aesan.msssi.gob.es/en/CRLMB/web/procedimientos_crlmb/crlmb_standard_operating_procedures.shtml (accessed on 4 March 2022).
- UNE-EN 16204:2012; Foodstuffs—Determination of Lipophilic Algal Toxins (Okadaic Acid Group Toxins, Yessotoxins, Azaspiracids, Pectenotoxins) in Shellfish and Shellfish Products by LC-MS/MS. European Committee of Standardization: Brussels, Belgium, 2012.
- Jansson, D.; Astot, C. Analysis of paralytic shellfish toxins, potential chemical threat agents, in food using hydrophilic interaction liquid chromatography-mass spectrometry. J. Chromatogr. A 2015, 1417, 41–48. [Google Scholar] [CrossRef]
- Thomas, K.M.; Beach, D.G.; Reeves, K.L.; Gibbs, R.S.; Kerrin, E.S.; McCarron, P.; Quilliam, M.A. Hydrophilic interaction liquid chromatography-tandem mass spectrometry for quantitation of paralytic shellfish toxins: Validation and application to reference materials. Anal. Bioanal. Chem. 2017, 409, 5675–5687. [Google Scholar] [CrossRef]
- Turner, A.D.; McNabb, P.S.; Harwood, D.T.; Selwood, A.I.; Boundy, M.J. Single-Laboratory Validation of a Multitoxin Ultra-Performance LC-Hydrophilic Interaction LC-MS/MS Method for Quantitation of Paralytic Shellfish Toxins in Bivalve Shellfish. J. AOAC Int. 2015, 98, 609–621. [Google Scholar] [CrossRef]
- Rey, V.; Botana, A.M.; Antelo, A.; Alvarez, M.; Botana, L.M. Rapid analysis of paralytic shellfish toxins and tetrodotoxins by liquid chromatography-tandem mass spectrometry using a porous graphitic carbon column. Food Chem. 2018, 269, 166–172. [Google Scholar] [CrossRef]
- CRLMB. Standard Operating Procedure for Determination of Domoic Acid (ASP Toxins) in Molluscs by UPLC-MS. 2010. Available online: http://aesan.msssi.gob.es/en/https://www.aesan.gob.es/en/CRLMB/web/public_documents/seccion/crlmb_standard_operating_procedures.htm/web/procedimientos_crlmb/crlmb_standard_operating_procedures.shtml (accessed on 4 March 2022).
- Rodriguez, I.; Alfonso, A.; Gonzalez-Jartin, J.M.; Vieytes, M.R.; Botana, L.M. A single run UPLC-MS/MS method for detection of all EU-regulated marine toxins. Talanta 2018, 189, 622–628. [Google Scholar] [CrossRef]
- Beach, D.G.; Kerrin, E.S.; Thomas, K.; Quilliam, M.A.; McCarron, P. Capillary electrophoresis-tandem mass spectrometry for multiclass analysis of polar marine toxins. Anal. Bioanal. Chem. 2018, 410, 5405–5420. [Google Scholar] [CrossRef] [PubMed]
- McNabb, P.; Selwood, A.I.; Holland, P.T.; Aasen, J.; Aune, T.; Eaglesham, G.; Hess, P.; Igarishi, M.; Quilliam, M.; Slattery, D.; et al. Multiresidue method for determination of algal toxins in shellfish: Single-laboratory validation and interlaboratory study. J. AOAC Int. 2005, 88, 761–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otero, P.; Miguens, N.; Rodriguez, I.; Botana, L.M. LC-MS/MS Analysis of the Emerging Toxin Pinnatoxin-G and High Levels of Esterified OA Group Toxins in Galician Commercial Mussels. Toxins 2019, 11, 394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estevez, P.; Sibat, M.; Leao-Martins, J.M.; Reis Costa, P.; Gago-Martinez, A.; Hess, P. Liquid Chromatography Coupled to High-Resolution Mass Spectrometry for the Confirmation of Caribbean Ciguatoxin-1 as the Main Toxin Responsible for Ciguatera Poisoning Caused by Fish from European Atlantic Coasts. Toxins 2020, 12, 267. [Google Scholar] [CrossRef] [Green Version]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Forino, M.; Tartaglione, L. Liquid chromatography-high-resolution mass spectrometry for palytoxins in mussels. Anal. Bioanal. Chem. 2015, 407, 1463–1473. [Google Scholar] [CrossRef]
- Estevez, P.; Castro, D.; Manuel Leao, J.; Yasumoto, T.; Dickey, R.; Gago-Martinez, A. Implementation of liquid chromatography tandem mass spectrometry for the analysis of ciguatera fish poisoning in contaminated fish samples from Atlantic coasts. Food Chem. 2019, 280, 8–14. [Google Scholar] [CrossRef]
- Pasinszki, T.; Lako, J.; Dennis, T.E. Advances in Detecting Ciguatoxins in Fish. Toxins 2020, 12, 494. [Google Scholar] [CrossRef]
- Wang, L.; Shi, X.; Zhao, Q.; Sun, A.; Li, D.; Zhao, J. Determination of lipophilic marine toxins in fresh and processed shellfish using modified QuEChERS and ultra-high-performance liquid chromatography-tandem mass spectrometry. Food Chem. 2019, 272, 427–433. [Google Scholar] [CrossRef]
- Rubies, A.; Munoz, E.; Gibert, D.; Cortes-Francisco, N.; Granados, M.; Caixach, J.; Centrich, F. New method for the analysis of lipophilic marine biotoxins in fresh and canned bivalves by liquid chromatography coupled to high resolution mass spectrometry: A quick, easy, cheap, efficient, rugged, safe approach. J. Chromatogr. A 2015, 1386, 62–73. [Google Scholar] [CrossRef]
- Kuklenyik, Z.; Ye, X.; Needham, L.L.; Calafat, A.M. Automated solid-phase extraction approaches for large scale biomonitoring studies. J. Chromatogr. Sci. 2009, 47, 12–18. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Chen, J.; He, X.; Hao, S.; Wang, Y.; Zheng, X.; Wang, B. Simple determination of six groups of lipophilic marine algal toxins in seawater by automated on-line solid phase extraction coupled to liquid chromatography-tandem mass spectrometry. Chemosphere 2021, 262, 128374. [Google Scholar] [CrossRef] [PubMed]
- Frederick, M.O.; De Lamo Marin, S.; Janda, K.D.; Nicolaou, K.C.; Dickerson, T.J. Monoclonal antibodies with orthogonal azaspiracid epitopes. Chembiochem 2009, 10, 1625–1629. [Google Scholar] [CrossRef] [PubMed]
- Garthwaite, I.; Ross, K.M.; Miles, C.O.; Briggs, L.R.; Towers, N.R.; Borrell, T.; Busby, P. Integrated enzyme-linked immunosorbent assay screening system for amnesic, neurotoxic, diarrhetic, and paralytic shellfish poisoning toxins found in New Zealand. J. AOAC Int. 2001, 84, 1643–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleivdal, H.; Kristiansen, S.I.; Nilsen, M.V.; Goksoyr, A.; Briggs, L.; Holland, P.; McNabb, P. Determination of domoic acid toxins in shellfish by biosense ASP ELISA—A direct competitive enzyme-linked immunosorbent assay: Collaborative study. J. AOAC Int. 2007, 90, 1011–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreuzer, M.P.; O’Sullivan, C.K.; Guilbault, G.G. Development of an ultrasensitive immunoassay for rapid measurement of okadaic acid and its isomers. Anal. Chem. 1999, 71, 4198–4202. [Google Scholar] [CrossRef]
- Bignami, G.S.; Raybould, T.J.; Sachinvala, N.D.; Grothaus, P.G.; Simpson, S.B.; Lazo, C.B.; Byrnes, J.B.; Moore, R.E.; Vann, D.C. Monoclonal antibody-based enzyme-linked immunoassays for the measurement of palytoxin in biological samples. Toxicon 1992, 30, 687–700. [Google Scholar] [CrossRef]
- Hokama, Y.; Abad, M.A.; Kimura, L.H. A rapid enzyme-immunoassay for the detection of ciguatoxin in contaminated fish tissues. Toxicon 1983, 21, 817–824. [Google Scholar] [CrossRef]
- Campas, M.; Leonardo, S.; Oshiro, N.; Kuniyoshi, K.; Tsumuraya, T.; Hirama, M.; Diogene, J. A smartphone-controlled amperometric immunosensor for the detection of Pacific ciguatoxins in fish. Food Chem. 2021, 374, 131687. [Google Scholar] [CrossRef]
- McNamee, S.E.; Elliott, C.T.; Greer, B.; Lochhead, M.; Campbell, K. Development of a planar waveguide microarray for the monitoring and early detection of five harmful algal toxins in water and cultures. Environ. Sci. Technol. 2014, 48, 13340–13349. [Google Scholar] [CrossRef]
- Vilarino, N.; Louzao, M.C.; Fraga, M.; Rodriguez, L.P.; Botana, L.M. Innovative detection methods for aquatic algal toxins and their presence in the food chain. Anal. Bioanal. Chem. 2013, 405, 7719–7732. [Google Scholar] [CrossRef]
- Jawaid, W.; Meneely, J.; Campbell, K.; Hooper, M.; Melville, K.; Holmes, S.; Rice, J.; Elliott, C. Development and validation of the first high performance-lateral flow immunoassay (HP-LFIA) for the rapid screening of domoic acid from shellfish extracts. Talanta 2013, 116, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Jawaid, W.; Campbell, K.; Melville, K.; Holmes, S.J.; Rice, J.; Elliott, C.T. Development and Validation of a Novel Lateral Flow Immunoassay (LFIA) for the Rapid Screening of Paralytic Shellfish Toxins (PSTs) from Shellfish Extracts. Anal. Chem. 2015, 87, 5324–5332. [Google Scholar] [CrossRef]
- Jawaid, W.; Meneely, J.P.; Campbell, K.; Melville, K.; Holmes, S.J.; Rice, J.; Elliott, C.T. Development and Validation of a Lateral Flow Immunoassay for the Rapid Screening of Okadaic Acid and All Dinophysis Toxins from Shellfish Extracts. J. Agric. Food Chem. 2015, 63, 8574–8583. [Google Scholar] [CrossRef] [PubMed]
- Anfossi, L.; Baggiani, C.; Giovannoli, C.; D’Arco, G.; Giraudi, G. Lateral-flow immunoassays for mycotoxins and phycotoxins: A review. Anal. Bioanal Chem. 2013, 405, 467–480. [Google Scholar] [CrossRef] [PubMed]
- Desmet, C.; Blum, L.J.; Marquette, C.A. High-throughput multiplexed competitive immunoassay for pollutants sensing in water. Anal. Chem. 2012, 84, 10267–10276. [Google Scholar] [CrossRef] [PubMed]
- Marquette, C.A.; Blum, L.J. Chemiluminescent enzyme immunoassays: A review of bioanalytical applications. Bioanalysis 2009, 1, 1259–1269. [Google Scholar] [CrossRef] [PubMed]
- Szkola, A.; Campbell, K.; Elliott, C.T.; Niessner, R.; Seidel, M. Automated, high performance, flow-through chemiluminescence microarray for the multiplexed detection of phycotoxins. Anal. Chim. Acta 2013, 787, 211–218. [Google Scholar] [CrossRef]
- Fraga, M.; Vilarino, N.; Louzao, M.C.; Rodriguez, P.; Campbell, K.; Elliott, C.T.; Botana, L.M. Multidetection of paralytic, diarrheic, and amnesic shellfish toxins by an inhibition immunoassay using a microsphere-flow cytometry system. Anal. Chem. 2013, 85, 7794–7802. [Google Scholar] [CrossRef]
- Stewart, L.D.; Elliott, C.T.; Walker, A.D.; Curran, R.M.; Connolly, L. Development of a monoclonal antibody binding okadaic acid and dinophysistoxins-1, -2 in proportion to their toxicity equivalence factors. Toxicon 2009, 54, 491–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AOAC. AOAC Official Method 2011.27. Paralytic shellfish toxins (PSTs) in shellfish, receptor binding assay. In Official Methods of Analysis of AOAC International, 19th ed.; AOAC International: Gaithersburg, MD, USA, 2012; Available online: http://www.eoma.aoac.org (accessed on 4 March 2022).
- Vieytes, M.R.; Cabado, A.G.; Alfonso, A.; Louzao, M.C.; Botana, A.M.; Botana, L.M. Solid-phase radioreceptor assay for paralytic shellfish toxins. Anal. Biochem. 1993, 211, 87–93. [Google Scholar] [CrossRef]
- McCall, J.R.; Jacocks, H.M.; Niven, S.C.; Poli, M.A.; Baden, D.G.; Bourdelais, A.J. Development and utilization of a fluorescence-based receptor-binding assay for the site 5 voltage-sensitive sodium channel ligands brevetoxin and ciguatoxin. J. AOAC Int. 2014, 97, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, L.P.; Vilarino, N.; Molgo, J.; Araoz, R.; Antelo, A.; Vieytes, M.R.; Botana, L.M. Solid-phase receptor-based assay for the detection of cyclic imines by chemiluminescence, fluorescence, or colorimetry. Anal. Chem. 2011, 83, 5857–5863. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, L.P.; Vilarino, N.; Molgo, J.; Araoz, R.; Botana, L.M. High-throughput receptor-based assay for the detection of spirolides by chemiluminescence. Toxicon 2013, 75, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Fonfria, E.S.; Vilarino, N.; Espina, B.; Louzao, M.C.; Alvarez, M.; Molgo, J.; Araoz, R.; Botana, L.M. Feasibility of gymnodimine and 13-desmethyl C spirolide detection by fluorescence polarization using a receptor-based assay in shellfish matrixes. Anal. Chim. Acta 2010, 657, 75–82. [Google Scholar] [CrossRef]
- Araoz, R.; Ramos, S.; Pelissier, F.; Guerineau, V.; Benoit, E.; Vilarino, N.; Botana, L.M.; Zakarian, A.; Molgo, J. Coupling the Torpedo microplate-receptor binding assay with mass spectrometry to detect cyclic imine neurotoxins. Anal. Chem. 2012, 84, 10445–10453. [Google Scholar] [CrossRef] [Green Version]
- Rubio, F.; Kamp, L.; Carpino, J.; Faltin, E.; Loftin, K.; Molgo, J.; Araoz, R. Colorimetric microtiter plate receptor-binding assay for the detection of freshwater and marine neurotoxins targeting the nicotinic acetylcholine receptors. Toxicon 2014, 91, 45–56. [Google Scholar] [CrossRef]
- Vieytes, M.R.; Fontal, O.I.; Leira, F.; Baptista de Sousa, J.M.; Botana, L.M. A fluorescent microplate assay for diarrheic shellfish toxins. Anal. Biochem. 1997, 248, 258–264. [Google Scholar] [CrossRef]
- Smienk, H.G.; Calvo, D.; Razquin, P.; Dominguez, E.; Mata, L. Single laboratory validation of a ready-to-use phosphatase inhibition assay for detection of okadaic acid toxins. Toxins 2012, 4, 339–352. [Google Scholar] [CrossRef]
- Campas, M.; de la Iglesia, P.; Le Berre, M.; Kane, M.; Diogene, J.; Marty, J.L. Enzymatic recycling-based amperometric immunosensor for the ultrasensitive detection of okadaic acid in shellfish. Biosens. Bioelectron. 2008, 24, 716–722. [Google Scholar] [CrossRef]
- Molinero-Abad, B.; Perez, L.; Izquierdo, D.; Escudero, I.; Arcos-Martinez, M.J. Sensor system based on flexible screen-printed electrodes for electrochemical detection of okadaic acid in seawater. Talanta 2019, 192, 347–352. [Google Scholar] [CrossRef]
- Zhao, L.; Huang, Y.; Dong, Y.; Han, X.; Wang, S.; Liang, X. Aptamers and Aptasensors for Highly Specific Recognition and Sensitive Detection of Marine Biotoxins: Recent Advances and Perspectives. Toxins 2018, 10, 427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eissa, S.; Ng, A.; Siaj, M.; Tavares, A.C.; Zourob, M. Selection and identification of DNA aptamers against okadaic acid for biosensing application. Anal. Chem. 2013, 85, 11794–11801. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Duan, N.; Zhang, H.; Wang, Z. Simultaneous detection of microcysin-LR and okadaic acid using a dual fluorescence resonance energy transfer aptasensor. Anal. Bioanal. Chem. 2015, 407, 1303–1312. [Google Scholar] [CrossRef] [PubMed]
- Alfaro, K.; Bustos, P.; O’Sullivan, C.; Conejeros, P. Facile and Cost-Effective Detection of Saxitoxin Exploiting Aptamer Structural Switching. Food Technol. Biotechnol. 2015, 53, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.J.; Park, J.H.; Lee, B.; Kim, M.G. Label-Free Direct Detection of Saxitoxin Based on a Localized Surface Plasmon Resonance Aptasensor. Toxins 2019, 11, 274. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Hu, B.; Zheng, X.; Cao, Y.; Liu, D.; Sun, M.; Jiao, B.; Wang, L. Gonyautoxin 1/4 aptamers with high-affinity and high-specificity: From efficient selection to aptasensor application. Biosens. Bioelectron. 2016, 79, 938–944. [Google Scholar] [CrossRef]
- Gao, S.; Zheng, X.; Wu, J. A biolayer interferometry-based enzyme-linked aptamer sorbent assay for real-time and highly sensitive detection of PDGF-BB. Biosens. Bioelectron. 2018, 102, 57–62. [Google Scholar] [CrossRef]
- Gu, H.; Duan, N.; Xia, Y.; Hun, X.; Wang, H.; Wang, Z. Magnetic Separation-Based Multiple SELEX for Effectively Selecting Aptamers against Saxitoxin, Domoic Acid, and Tetrodotoxin. J. Agric. Food Chem. 2018, 66, 9801–9809. [Google Scholar] [CrossRef]
- Caglayan, M.O.; Ustundag, Z. Saxitoxin aptasensor based on attenuated internal reflection ellipsometry for seafood. Toxicon 2020, 187, 255–261. [Google Scholar] [CrossRef]
- Ullah, N.; Chen, W.; Noureen, B.; Tian, Y.; Du, L.; Wu, C.; Ma, J. An Electrochemical Ti3C2Tx Aptasensor for Sensitive and Label-Free Detection of Marine Biological Toxins. Sensors 2021, 21, 4938. [Google Scholar] [CrossRef]
- Pan, Y.; Wan, Z.; Zhong, L.; Li, X.; Wu, Q.; Wang, J.; Wang, P. Label-free okadaic acid detection using growth of gold nanoparticles in sensor gaps as a conductive tag. Biomed. Microdevices 2017, 19, 33. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Duan, N.; Wu, S.; Hao, L.; Xia, Y.; Ma, X.; Wang, Z. Graphene oxide-assisted non-immobilized SELEX of okdaic acid aptamer and the analytical application of aptasensor. Sci. Rep. 2016, 6, 21665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramalingam, S.; Chand, R.; Singh, C.B.; Singh, A. Phosphorene-gold nanocomposite based microfluidic aptasensor for the detection of okadaic acid. Biosens. Bioelectron. 2019, 135, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, Y.; Wu, P.; Wang, W.; Cheng, Y.; Huang, L.; Bai, J.; Peng, Y.; Ning, B.; Gao, Z.; et al. Development of a highly sensitive detection method for TTX based on a magnetic bead-aptamer competition system under triple cycle amplification. Anal. Chim. Acta 2020, 1119, 18–24. [Google Scholar] [CrossRef]
- Gao, S.; Zheng, X.; Hu, B.; Sun, M.; Wu, J.; Jiao, B.; Wang, L. Enzyme-linked, aptamer-based, competitive biolayer interferometry biosensor for palytoxin. Biosens. Bioelectron. 2017, 89 Pt 2, 952–958. [Google Scholar] [CrossRef]
- Kepp, O.; Galluzzi, L.; Lipinski, M.; Yuan, J.; Kroemer, G. Cell death assays for drug discovery. Nat. Rev. Drug Discov. 2011, 10, 221–237. [Google Scholar] [CrossRef]
- An, W.F.; Tolliday, N. Cell-based assays for high-throughput screening. Mol. Biotechnol. 2010, 45, 180–186. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Caillaud, A.; Eixarch, H.; de la Iglesia, P.; Rodriguez, M.; Dominguez, L.; Andree, K.B.; Diogene, J. Towards the standardisation of the neuroblastoma (neuro-2a) cell-based assay for ciguatoxin-like toxicity detection in fish: Application to fish caught in the Canary Islands. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2012, 29, 1000–1010. [Google Scholar] [CrossRef]
- Espina, B.; Cagide, E.; Louzao, M.C.; Fernandez, M.M.; Vieytes, M.R.; Katikou, P.; Villar, A.; Jaen, D.; Maman, L.; Botana, L.M. Specific and dynamic detection of palytoxins by in vitro microplate assay with human neuroblastoma cells. Biosci. Rep. 2009, 29, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Ledreux, A.; Serandour, A.L.; Morin, B.; Derick, S.; Lanceleur, R.; Hamlaoui, S.; Furger, C.; Bire, R.; Krys, S.; Fessard, V.; et al. Collaborative study for the detection of toxic compounds in shellfish extracts using cell-based assays. Part II: Application to shellfish extracts spiked with lipophilic marine toxins. Anal. Bioanal. Chem. 2012, 403, 1995–2007. [Google Scholar] [CrossRef] [PubMed]
- Canete, E.; Diogene, J. Comparative study of the use of neuroblastoma cells (Neuro-2a) and neuroblastomaxglioma hybrid cells (NG108-15) for the toxic effect quantification of marine toxins. Toxicon 2008, 52, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Bodero, M.; Bovee, T.F.H.; Wang, S.; Hoogenboom, R.; Klijnstra, M.D.; Portier, L.; Hendriksen, P.J.M.; Gerssen, A. Screening for the presence of lipophilic marine biotoxins in shellfish samples using the neuro-2a bioassay. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2018, 35, 351–365. [Google Scholar] [CrossRef] [PubMed]
- Bignami, G.S. A rapid and sensitive hemolysis neutralization assay for palytoxin. Toxicon 1993, 31, 817–820. [Google Scholar] [CrossRef]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerford, J.M.; Wekell, M.M. Tetrazolium-based cell bioassay for neurotoxins active on voltage-sensitive sodium channels: Semiautomated assay for saxitoxins, brevetoxins, and ciguatoxins. Anal. Biochem. 1993, 214, 190–194. [Google Scholar] [CrossRef]
- Castro, D.; Manger, R.; Vilarino, O.; Gago-Martinez, A. Evaluation of Matrix Issues in the Applicability of the Neuro-2a Cell Based Assay on the Detection of CTX in Fish Samples. Toxins 2020, 12, 308. [Google Scholar] [CrossRef]
- Wang, Q.; Fang, J.; Cao, D.; Li, H.; Su, K.; Hu, N.; Wang, P. An improved functional assay for rapid detection of marine toxins, saxitoxin and brevetoxin using a portable cardiomyocyte-based potential biosensor. Biosens. Bioelectron. 2015, 72, 10–17. [Google Scholar] [CrossRef]
- Zou, L.; Wu, C.; Wang, Q.; Zhou, J.; Su, K.; Li, H.; Hu, N.; Wang, P. An improved sensitive assay for the detection of PSP toxins with neuroblastoma cell-based impedance biosensor. Biosens. Bioelectron. 2015, 67, 458–464. [Google Scholar] [CrossRef]
- Louzao, M.C.; Vieytes, M.R.; Baptista de Sousa, J.M.; Leira, F.; Botana, L.M. A fluorimetric method based on changes in membrane potential for screening paralytic shellfish toxins in mussels. Anal. Biochem. 2001, 289, 246–250. [Google Scholar] [CrossRef]
- Manger, R.; Woodle, D.; Berger, A.; Dickey, R.W.; Jester, E.; Yasumoto, T.; Lewis, R.; Hawryluk, T.; Hungerford, J. Flow cytometric-membrane potential detection of sodium channel active marine toxins: Application to ciguatoxins in fish muscle and feasibility of automating saxitoxin detection. J. AOAC Int. 2014, 97, 299–306. [Google Scholar] [CrossRef]
- Rivera, A.A.; Aballay-Gonzalez, A.; Goncalves, A.T.; Tarifeno, E.; Ulloa, V.; Gallardo, J.J.; Astuya-Villalon, A. Search for potential biomarkers for saxitoxin detection. Toxicol Vitr. 2021, 72, 105092. [Google Scholar] [CrossRef] [PubMed]
- Bodero, M.; Bovee, T.F.H.; Portier, L.; Hendriksen, P.J.M. Detection and profiling of diarrheic marine biotoxins in shellfish by mRNA analysis of exposed Caco-2 cells using qRT-PCR and multiplex magnetic bead-based assays. ALTEX 2019, 36, 203–214. [Google Scholar] [PubMed]
- Rodrigues, S.M.; Vale, P.; Chaveca, T.; Laires, A.; Rueff, J.; Oliveira, N.G. Naturally contaminated shellfish samples: Quantification of diarrhetic shellfish poisoning toxins in unhydrolysed and hydrolysed extracts and cytotoxicity assessment. J. Appl. Toxicol. 2010, 30, 699–707. [Google Scholar] [CrossRef] [PubMed]
- Serandour, A.L.; Ledreux, A.; Morin, B.; Derick, S.; Augier, E.; Lanceleur, R.; Hamlaoui, S.; Moukha, S.; Furger, C.; Bire, R.; et al. Collaborative study for the detection of toxic compounds in shellfish extracts using cell-based assays. Part I: Screening strategy and pre-validation study with lipophilic marine toxins. Anal. Bioanal. Chem. 2012, 403, 1983–1993. [Google Scholar] [CrossRef]
- Beani, L.; Bianchi, C.; Guerrini, F.; Marani, L.; Pistocchi, R.; Tomasini, M.C.; Ceredi, A.; Milandri, A.; Poletti, R.; Boni, L. High sensitivity bioassay of paralytic (PSP) and amnesic (ASP) algal toxins based on the fluorimetric detection of [Ca(2+)](i) in rat cortical primary cultures. Toxicon 2000, 38, 1283–1297. [Google Scholar] [CrossRef]
- Dechraoui, M.Y.; Tiedeken, J.A.; Persad, R.; Wang, Z.; Granade, H.R.; Dickey, R.W.; Ramsdell, J.S. Use of two detection methods to discriminate ciguatoxins from brevetoxins: Application to great barracuda from Florida Keys. Toxicon 2005, 46, 261–270. [Google Scholar] [CrossRef]
- EU. Commission Implementing Regulation (EU) 2019/627 of 15 March 2019 laying down uniform practical arrangements for the performance of official controls on products of animal origin intended for human consumption in accordance with Regulation (EU) 2017/625 of the European Parliament and of the Council and amending Commission Regulation (EC) No 2074/2005 as regards official controls. Off. J. Eur. Union 2019, L131, 51–100. [Google Scholar]
- EU. Commission Regulation 2017/1980 of 31 October 2017 amending Annex III to Regulation (EC) No 2074/2005 as regards paralytic shellfish poison (PSP) detection method. Off. J. Eur. Union 2017, L285, 8–9. [Google Scholar]
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [Green Version]
- Wells, M.L.; Trainer, V.L.; Smayda, T.J.; Karlson, B.S.; Trick, C.G.; Kudela, R.M.; Ishikawa, A.; Bernard, S.; Wulff, A.; Anderson, D.M.; et al. Harmful algal blooms and climate change: Learning from the past and present to forecast the future. Harmful Algae 2015, 49, 68–93. [Google Scholar] [CrossRef] [Green Version]
- Comiso, J.C. Variability and trends of globla sea ice cover and sea level: Effects on physicochemical parameters. In Climate Change and Marine and Freshwater Toxins, 2nd ed.; Botana, L.M., Louzao, M.C., Vilariño, N., Eds.; De Gruyter: Berlin, Germany, 2021; pp. 1–34. [Google Scholar]
- McCarthy, G.D.; Haigh, I.D.; Hirschi, J.J.; Grist, J.P.; Smeed, D.A. Ocean impact on decadal Atlantic climate variability revealed by sea-level observations. Nature 2015, 521, 508–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, S.; John, U.; Savela, H.; Kremp, A. Alexandrium spp.: Genetic and ecological factors influencing saxitoxin production and proliferation. In Climate Change and Marine and Freshwater Toxins, 2nd ed.; Botana, L.M., Louzao, M.C., Vilariño, N., Eds.; De Gruyter: Berlin, Germany, 2021; pp. 133–166. [Google Scholar]
- Aligizaki, K.; Katikou, P.; Nikolaidis, G.; Panou, A. First episode of shellfish contamination by palytoxin-like compounds from Ostreopsis species (Aegean Sea, Greece). Toxicon 2008, 51, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Ciminiello, P.; Dell’Aversano, C.; Iacovo, E.D.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Battocchi, C.; Crinelli, R.; Carloni, E.; Magnani, M.; et al. Unique toxin profile of a Mediterranean Ostreopsis cf. ovata strain: HR LC-MS(n) characterization of ovatoxin-f, a new palytoxin congener. Chem. Res. Toxicol. 2012, 25, 1243–1252. [Google Scholar] [CrossRef] [PubMed]
- Tartaglione, L.; Dello Iacovo, E.; Mazzeo, A.; Casabianca, S.; Ciminiello, P.; Penna, A.; Dell’Aversano, C. Variability in Toxin Profiles of the Mediterranean Ostreopsis cf. ovata and in Structural Features of the Produced Ovatoxins. Environ. Sci. Technol. 2017, 51, 13920–13928. [Google Scholar]
- Santos, M.; Oliveira, P.B.; Moita, M.T.; David, H.; Caeiro, M.F.; Zingone, A.; Amorim, A.; Silva, A. Ocurrence of Ostreopsis in two temperate coastal bays (SW iberia): Insights from the plankton. Harmful Algae 2019, 86, 20–36. [Google Scholar] [CrossRef]
- Nunez, D.; Matute, P.; Garcia, A.; Garcia, P.; Abadia, N. Outbreak of ciguatera food poisoning by consumption of amberjack (Seriola spp.) in the Canary Islands, May 2012. Euro Surveill. 2012, 17, pii=20188. [Google Scholar] [CrossRef]
- Laza-Martinez, A.; David, H.; Riobo, P.; Miguel, I.; Orive, E. Characterization of a Strain of Fukuyoa paulensis (Dinophyceae) from the Western Mediterranean Sea. J. Eukaryot. Microbiol. 2016, 63, 481–497. [Google Scholar] [CrossRef] [Green Version]
- Fraga, S.; Rodriguez, F. Genus Gambierdiscus in the Canary Islands (NE Atlantic Ocean) with description of Gambierdiscus silvae sp. nov., a new potentially toxic epiphytic benthic dinoflagellate. Protist 2014, 165, 839–853. [Google Scholar] [CrossRef]
- Katikou, P.; Georgantelis, D.; Sinouris, N.; Petsi, A.; Fotaras, T. First report on toxicity assessment of the Lessepsian migrant pufferfish Lagocephalus sceleratus (Gmelin, 1789) from European waters (Aegean Sea, Greece). Toxicon 2009, 54, 50–55. [Google Scholar] [CrossRef]
- Kosker, A.R.; Ozogul, F.; Ayas, D.; Durmus, M.; Ucar, Y.; Regenstein, J.M.; Ozogul, Y. Tetrodotoxin levels of three pufferfish species (Lagocephalus sp.) caught in the North-Eastern Mediterranean sea. Chemosphere 2019, 219, 95–99. [Google Scholar] [CrossRef]
- Gerssen, A.; Bovee, T.H.F.; Klijnstra, M.D.; Poelman, M.; Portier, L.; Hoogenboom, R. First Report on the Occurrence of Tetrodotoxins in Bivalve Mollusks in The Netherlands. Toxins 2018, 10, 450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leao, J.M.; Lozano-Leon, A.; Giraldez, J.; Vilarino, O.; Gago-Martinez, A. Preliminary Results on the Evaluation of the Occurrence of Tetrodotoxin Associated to Marine Vibrio spp. in Bivalves from the Galician Rias (Northwest of Spain). Mar. Drugs 2018, 16, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, M.; Rodriguez, I.; Barreiro, A.; Kaufmann, M.; Neto, A.I.; Hassouani, M.; Sabour, B.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. Tetrodotoxins Occurrence in Non-Traditional Vectors of the North Atlantic Waters (Portuguese Maritime Territory, and Morocco Coast). Toxins 2019, 11, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]








| Toxin | TEF EFSA [124] | TEFs Based on In Vitro Assay: Nav1.6 Current Peak [59] | TEFs Based on In Vitro Assay: Nav1.6 Shift Activation Potential [59] |
|---|---|---|---|
| P-CTX-1 | 1 | 1 | 1 |
| P-CTX-2 | 0.3 | ||
| P-CTX-3 | 0.3 | 0.28 | 0.28 |
| P-CTX-3C | 0.2 | ||
| 2,3-dihydroxy P-CTX-3C | 0.1 | ||
| 51-hydroxy P-CTX-3C | 1 | ||
| P-CTX-4A | 0.1 | 0.0016 | 0.1 |
| P-CTX-4B | 0.05 | ||
| C-CTX-1 | 0.1 | ||
| C-CTX-2 | 0.3 |
| Toxin | TEF EFSA [205] | TEF FAO [70] | TEFs Based on Oral Gavage [39,40] | TEFs Based on In Vitro IC50 for PP2A [206] | TEFs Based on PP2A Inhibition by FAO [70] |
|---|---|---|---|---|---|
| OA | 1 | 1 | 1 | 1 | 1 |
| DTX1 | 1 | 1 | 1.5 | 1.6 | 1.6 |
| DTX2 | 0.6 | 0.5 | 0.3 | 0.3 | 0.5 |
| Toxin | TEF EFSA [19] | TEF FAO [70] | TEFs Based on Oral Gavage [215] |
|---|---|---|---|
| AZA1 | 1 | 1 | 1 |
| AZA2 | 1.8 | 0.7 | 0.7 |
| AZA3 | 1.4 | 0.5 | 0.5 |
| AZA6 | 0.7 |
| Old Facts | Current Facts | |
|---|---|---|
| Mechanism of Action | ||
| OA | PP inhibition, diarrhea caused by disruption of gap-junctions | PP inhibition in combination with neurological pathways |
| AZAs | Main target hERG | Remains unknown |
| CTX | Sodium current peak amplitude | Sodium current peak amplitude and activation shift to more negative potential |
| TEFs | ||
| OA | OA ≈ DTX1 | DTX1 > OA |
| STX | STX ≈ NeoSTX ≈ dcSTX | NeoSTX > STX > dcSTX |
| Toxicity | ||
| Maitotoxins | Most potent toxins | No CFP cases reported, nor oral toxicity detected |
| PTXs | Causative DSP agent | Not causative of DSP, no human poisoning reported |
| TTX | Minimum lethal dose for humans 2 mg | EFSA proposed ARfD of 0.25 μg/kg bw |
| European legislation | ||
| PTXs | Regulated as part of OA group of toxins | Not legislated anymore |
| YTX | Legal limit 1 mg YTX equivalents/kg shellfish | Legal limit 3.75 mg YTX equivalents/kg shellfish |
| TTX | Emerging toxins responsible for human poisoning | Detected in seafood products more frequently, non-regulated |
| CTX | ||
| PLTX | ||
| CIs | Emerging toxins | |
| Toxin | Aptamer Name | Aptamer Sequence | Sensor Technology | LOD (ng/mL) | Seafood Samples | Reference |
|---|---|---|---|---|---|---|
| STX | M-30f | TTG AGG GTC GCA TCC CGT GGA AAC AGG TTC ATTG | Biolayer interferometry | 0.5 ng/mL | Shellfish | [388] |
| STX | APTSTX | GGT ATT GAG GGT CGC ATC CCG TGG AAA CAT GTT CAT TGG GCG CAC TCC GCT TTC TGT AGA TGG CTC TAA CTC TCC TCT | Fluorescence | 7.5 ng/mL | - | [385] |
| STX | - | TTT TTT AGG GAA GAG AAG GAC ATA TGA TGG CAC AAG GCC CAT CAA TCG GTA TAC GGG TTG ACT AGT ACA TGA CCA CTT GA | Localized surface plasmon resonance | 2.5 ng/mL | Mussel | [386] |
| STX | STX-41 | ATA GGA GTC ACG ACG ACC AGC TTT TTA CAA AAT TCT CTT TTT ACC TAT ATT ATG AAC AGA TAT GTG CGT CTA CCT CTT GA | Fluorescence | 0.4 ng/mL | Clam | [389] |
| STX | APTSTX | GGT ATT GAG GGT CGC ATC CCG TGG AAA CAT GTT CAT TGG GCG CAC TCC GCT TTC TGT AGA TGG CTC TAA CTC TCC TCT | Attenuated Internal reflection spectroscopic ellipsometry | 10 pg/mL | Shrimp | [390] |
| STX | APTSTX | GGT ATT GAG GGT CGC ATC CCG TGG AAA CAT GTT CAT TGG GCG CAC TCC GCT TTC TGT AGA TGG CTC TAA CTC TCC TCT | Electrochemistry | 9 pg/mL | Mussel | [391] |
| DA | DA-06 | ATA GGA GTC ACG ACG ACC AGA AAA ATA ATT TAA ATT TTC TAC CCA ATG CTT TTC GCA TAA TAT GTG CGT CTA CCT CTT GA | Fluorescence | 0.45 ng/mL | Clam | [389] |
| GTX1/4 | GO18-T-d | AAC CTT TGG TCG GGC AAG GTA GGT T | Biolayer interferometry | 50 pg/mL | Shellfish | [387] |
| OA | OA34 | GGT CAC CAA CAA CAG GGA GCG CTA CGC GAA GGG TCA ATG TGA CGT CAT GCG GAT GTG TGG | Electrochemistry | 70 pg/mL | shellfish | [383] |
| OA | OA34 | GGT CAC CAA CAA CAG GGA GCG CTA CGC GAA GGG TCA ATG TGA CGT CAT GCG GAT GTG TGG | Electrochemistry | 1 ng/mL | - | [392] |
| OA | OA34 | GGT CAC CAA CAA CAG GGA GCG CTA CGC GAA GGG TCA ATG TGA CGT CAT GCG GAT GTG TGG | Fluorescence | 50 pg/mL | shrimp | [384] |
| OA | OA27-1 | TGT CGA GGG AGA CGC GCA GTC GCT ACC ACC T | Colorimetric (enzyme-linked aptamer assay) | 10 pg/mL | Clam | [393] |
| OA | OA-Apt | GGT CAC CAA CAA CAG GGA GCG CTA CGC GAA GGG TCA ATG TGA CGT CAT GCG GAT GTG TGG | Colorimetric (enzyme-linked aptamer assay) | 6.4 pg/mL | Mussel | [394] |
| Tetrodotoxin | TTX-07 | ATA GGA GTC ACG ACG ACC AGT CAA ATT TTC GTC TAC TCA ATC TTT CTG TCT TAT CTA TGT GCG TCT ACC TCT TGA | Fluorescence | 0.26 pg/mL | Clam, shellfish | [395] |
| Palytoxin | PTX-13 | GGA GGT GGT GGG GAC TTT GCT TGT ACT GGG CGC CCG GTT GAA | Biolayer interferometry | 0.04 pg/mL | Mussel, clam, scallop | [396] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Louzao, M.C.; Vilariño, N.; Vale, C.; Costas, C.; Cao, A.; Raposo-Garcia, S.; Vieytes, M.R.; Botana, L.M. Current Trends and New Challenges in Marine Phycotoxins. Mar. Drugs 2022, 20, 198. https://doi.org/10.3390/md20030198
Louzao MC, Vilariño N, Vale C, Costas C, Cao A, Raposo-Garcia S, Vieytes MR, Botana LM. Current Trends and New Challenges in Marine Phycotoxins. Marine Drugs. 2022; 20(3):198. https://doi.org/10.3390/md20030198
Chicago/Turabian StyleLouzao, Maria Carmen, Natalia Vilariño, Carmen Vale, Celia Costas, Alejandro Cao, Sandra Raposo-Garcia, Mercedes R. Vieytes, and Luis M. Botana. 2022. "Current Trends and New Challenges in Marine Phycotoxins" Marine Drugs 20, no. 3: 198. https://doi.org/10.3390/md20030198