
Anti-Allergic Effect of 3,4-Dihydroxybenzaldehyde Isolated from Polysiphonia morrowii in IgE/BSA-Stimulated Mast Cells and a Passive Cutaneous Anaphylaxis Mouse Model

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
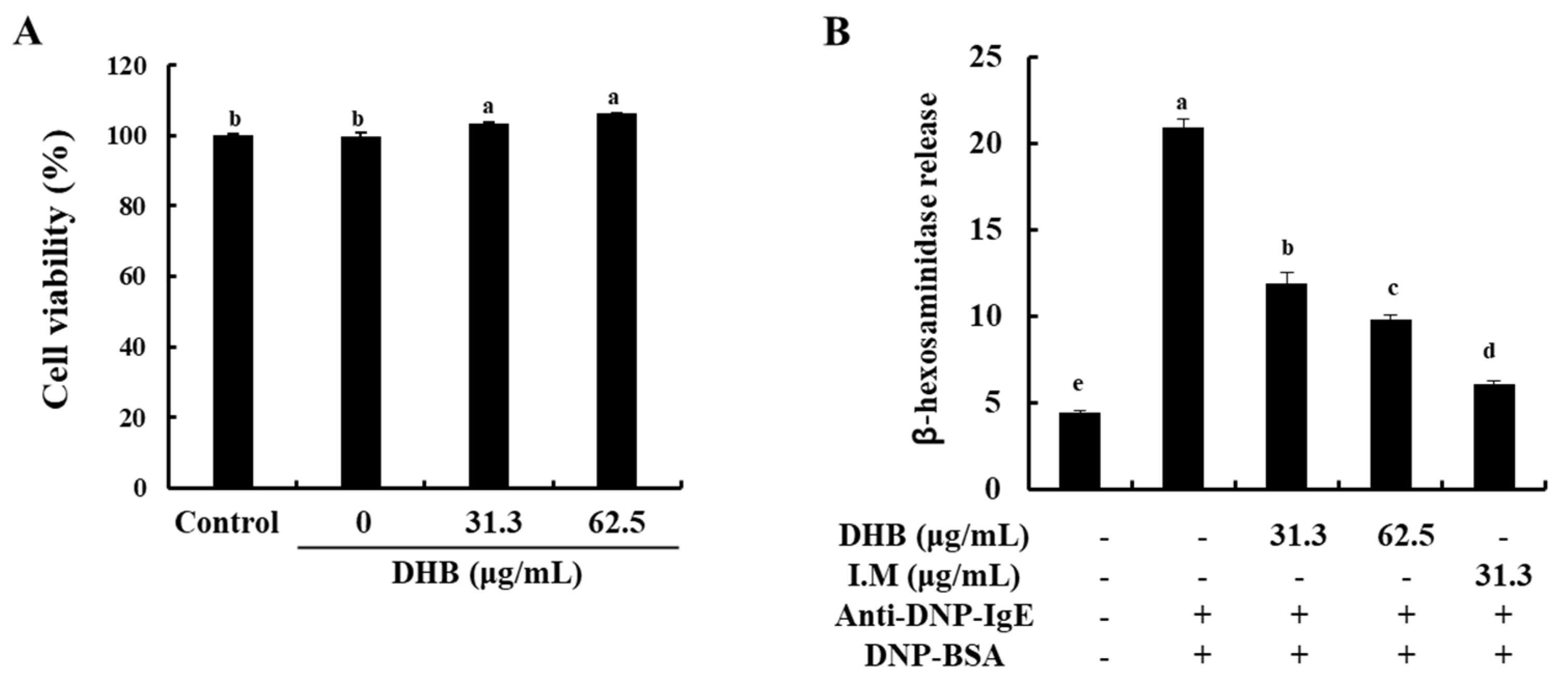
2.1. DHB Effectively Decreased β-Hexosaminidase Release without Cytotoxicity in BMCMCs
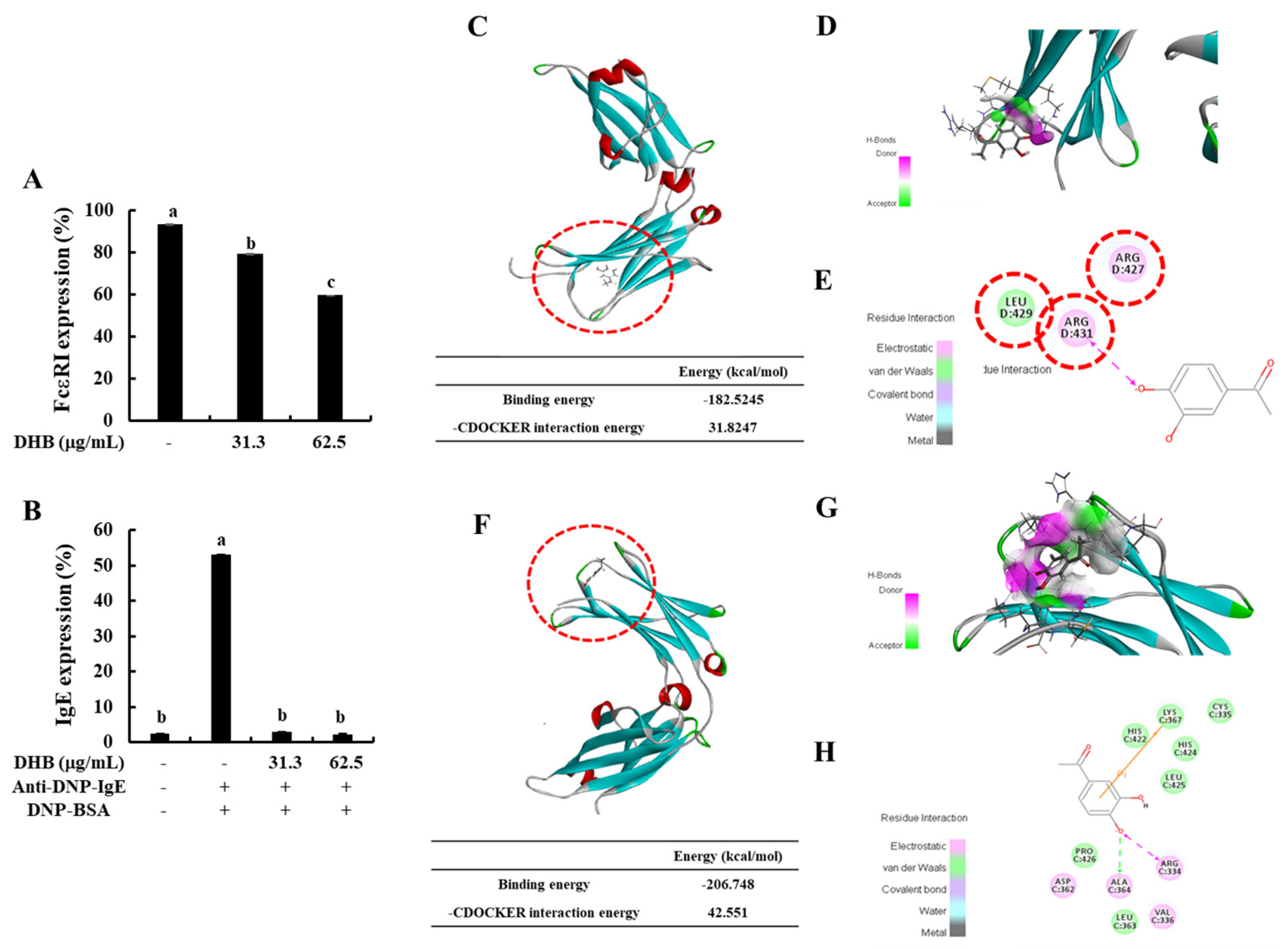
2.2. DHB Inhibited the Expression of FcεRI and the Binding of IgE to FcεRI
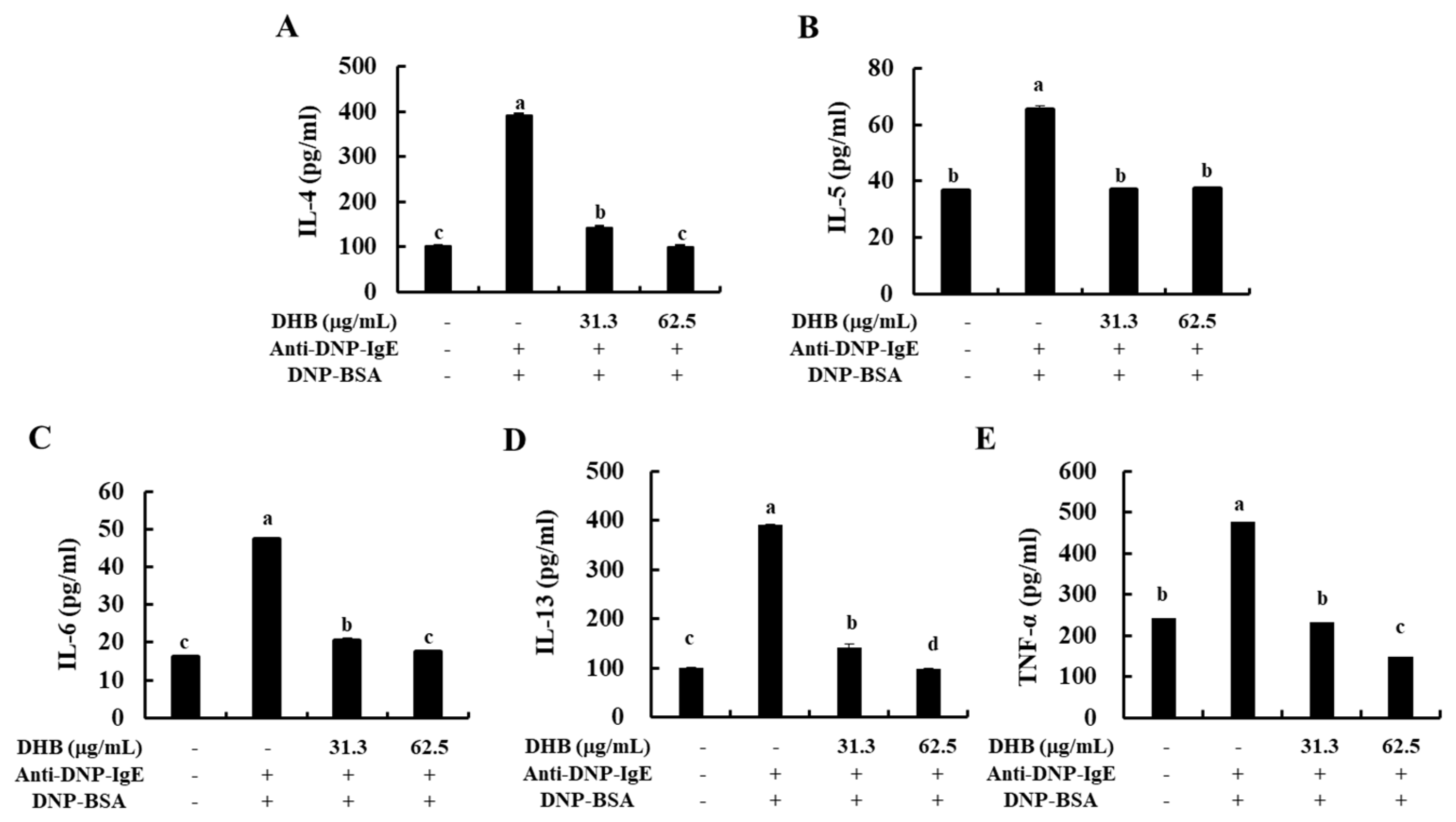
2.3. DHB Reduced the Secretion of Allergic Cytokines in the IgE/BSA-Stimulated BMCMCs
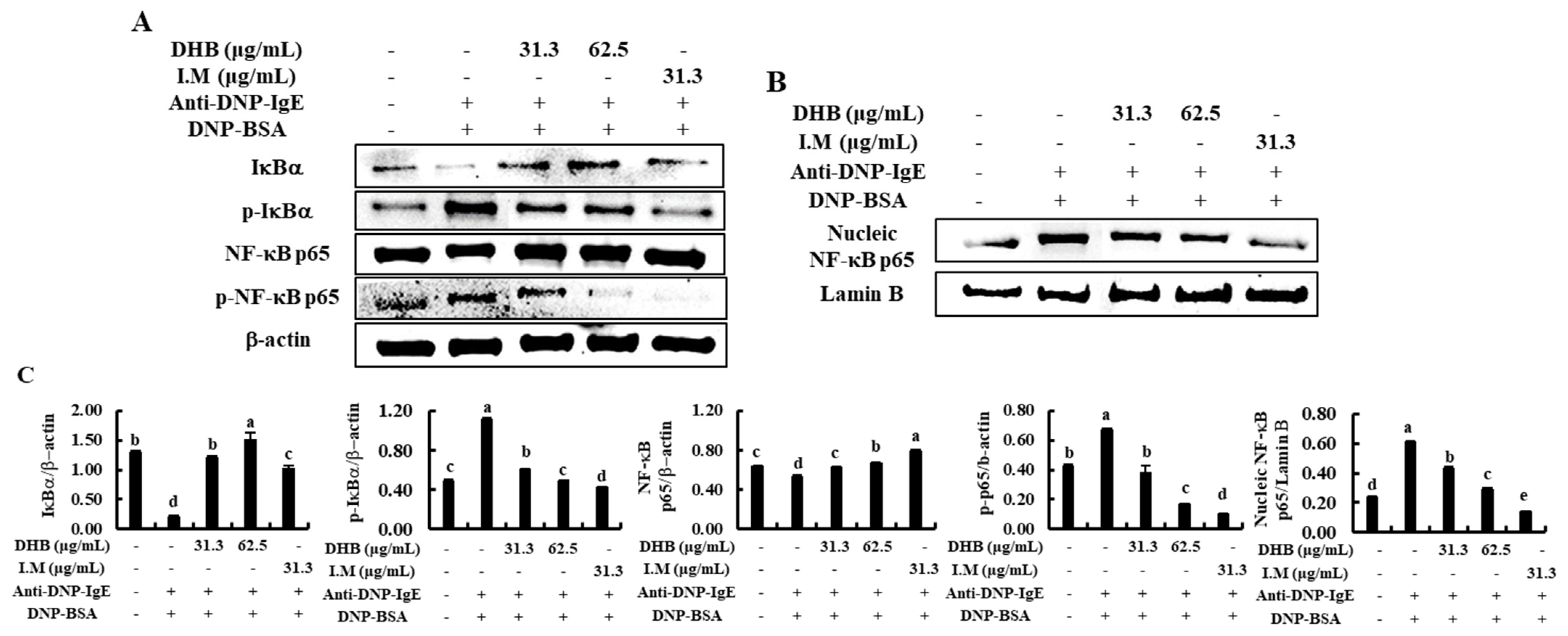
2.4. DHB Inhibited the Activation of NF-κB Signaling Pathway in the IgE/BSA-Stimulated BMCMCs
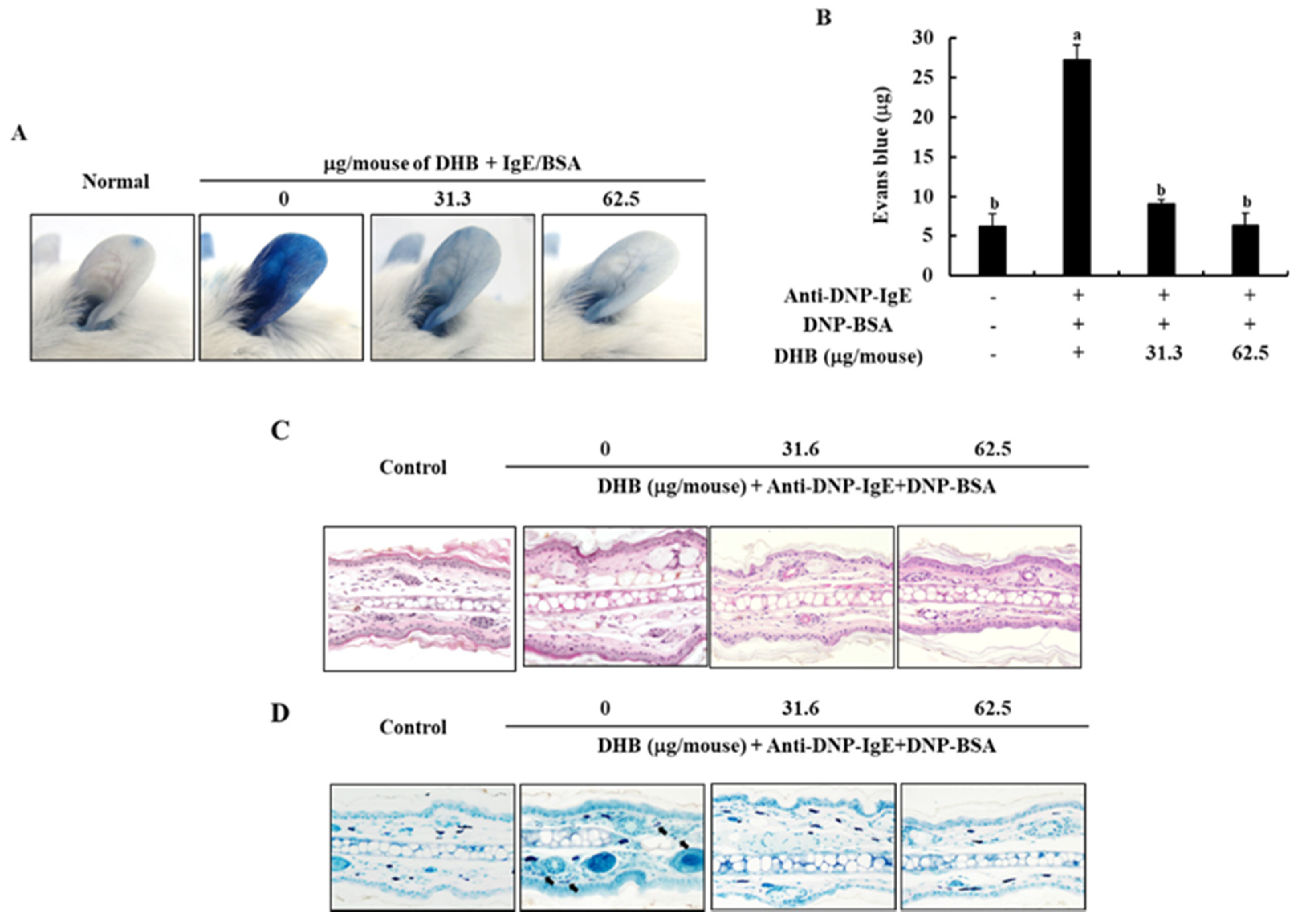
2.5. DHB Improves the IgE/BSA Stimulation-Induced the PCA Reaction in Mice
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Purification of DHB from P. morrowii
4.3. Preparation of BMCMCs
4.4. Measurement of Cytotoxicity
4.5. Measurement of β-Hexosaminidase Release
4.6. RT-PCR Analysis
4.7. Measurement of Cytokines Levels
4.8. Flow Cytometric Analysis of Mast Cells Expressing FcεRI and FcεRI-IgE (IgE Binding)
4.9. Western Blot Analysis
4.10. In Silico Docking of DHB with IgE and FcεRI
4.11. PCA Test
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Min, K.-D.; Yi, S.-J.; Kim, H.-C.; Leem, J.-H.; Kwon, H.-J.; Hong, S.; Kim, K.S.; Kim, S.-Y. Association between exposure to traffic-related air pollution and pediatric allergic diseases based on modeled air pollution concentrations and traffic measures in Seoul, Korea: A comparative analysis. Environ. Health 2020, 19, 6. [Google Scholar] [CrossRef] [Green Version]
- Dhakal, H.; Lee, S.; Choi, J.K.; Kwon, T.K.; Khang, D.; Kim, S.-H. Inhibitory effects of orientin in mast cell-mediated allergic inflammation. Pharmacol. Rep. 2020, 72, 1002–1010. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.-T.; Goodarzi, H.; Chen, H.-Y. IgE, Mast Cells, and Eosinophils in Atopic Dermatitis. Clin. Rev. Allergy Immunol. 2011, 41, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Kalesnikoff, J.; Huber, M.; Lam, V.; Damen, J.E.; Zhang, J.; Siraganian, R.P.; Krystal, G. Monomeric IgE Stimulates Signaling Pathways in Mast Cells that Lead to Cytokine Production and Cell Survival. Immunity 2001, 14, 801–811. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.-J.; Choi, Y.-A.; Lee, S.; Choi, J.K.; Kim, Y.-Y.; Kim, E.-N.; Jeong, G.-S.; Shin, T.-Y.; Jang, Y.H.; Kim, S.-H. Prunus serrulata var. spontanea inhibits mast cell activation and mast cell-mediated anaphylaxis. J. Ethnopharmacol. 2019, 250, 112484. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Piao, H.; Jiang, J.; Jin, G.; Zheng, M.; Yang, J.; Jin, X.; Sun, T.; Choi, Y.H.; Li, L.; et al. Polydatin inhibits mast cell-mediated allergic inflammation by targeting PI3K/Akt, MAPK, NF-κB and Nrf2/HO−1 pathways. Sci. Rep. 2017, 7, 11895. [Google Scholar] [CrossRef] [PubMed]
- Ahn, G.; Amagai, Y.; Matsuda, A.; Kang, S.-M.; Lee, W.; Jung, K.; Oida, K.; Jang, H.; Ishizaka, S.; Matsuda, K.; et al. Dieckol, a phlorotannin ofEcklonia cava, suppresses IgE-mediated mast cell activation and passive cutaneous anaphylactic reaction. Exp. Dermatol. 2015, 24, 968–970. [Google Scholar] [CrossRef] [Green Version]
- Wee, J.H.; Zhang, Y.-L.; Rhee, C.-S.; Kim, D.-Y. Inhibition of allergic response by intranasal selective NF-κB decoy oligodeoxynucleotides in a murine model of allergic rhinitis. Allergy Asthma Immunol. Res. 2017, 9, 61. [Google Scholar] [CrossRef] [Green Version]
- Han, E.J.; Fernando, I.P.S.; Kim, E.-A.; Kim, J.; Jung, K.; Kim, S.-Y.; Cha, S.-H.; Kim, K.-N.; Heo, S.-J.; Ahn, G. 5-Bromo-3,4-dihydroxybenzaldehyde from Polysiphonia morrowii attenuate IgE/BSA-stimulated mast cell activation and passive cutaneous anaphylaxis in mice. Biochem. Pharmacol. 2020, 178, 114087. [Google Scholar] [CrossRef]
- Han, E.J.; Kim, H.-S.; Sanjeewa, K.; Herath, K.; Jeon, Y.-J.; Jee, Y.; Lee, J.; Kim, T.; Shim, S.-Y.; Ahn, G. Eckol from Ecklonia cava Suppresses Immunoglobulin E-mediated Mast Cell Activation and Passive Cutaneous Anaphylaxis in Mice (Running Title: Anti-Allergic Activity of Ecklonia cava). Nutrients 2020, 12, 1361. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, B.; Kim, H.K.; Kim, E.-Y.; Kim, J.-H.; Min, J.-H.; Kim, S.; Sohn, Y.; Jung, H.-S. Peimine inhibits the production of proinflammatory cytokines through regulation of the phosphorylation of NF-κB and MAPKs in HMC-1 Cells. Pharmacogn. Mag. 2017, 13, S359. [Google Scholar] [PubMed] [Green Version]
- Han, E.J.; Shin, E.-J.; Kim, M.J.; Heo, S.-J.; Kim, E.-A.; Ahn, G. Anti-allergic Effect of Methanol Extract from Polysiphonia morrowii Harveyin IgE/BSA-stimulated Bone Marrow-derived Cultured Mast Cells anda Passive Cutaneous Anaphylaxis Mouse Model. J. Chitin Chitosan 2019, 24, 99–104. [Google Scholar] [CrossRef]
- Ko, E.-Y.; Heo, S.-J.; Cho, S.-H.; Lee, W.; Kim, S.-Y.; Yang, H.-W.; Ahn, G.; Cha, S.-H.; Kwon, S.-H.; Jeong, M.S.; et al. 3-Bromo-5-(ethoxymethyl)-1, 2-benzenediol inhibits LPS-induced pro-inflammatory responses by preventing ROS production and downregulating NF-κB in vitro and in a zebrafish model. Int. Immunopharmacol. 2019, 67, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.-H.; Heo, S.-J.; Yang, H.-W.; Ko, E.-Y.; Jung, M.S.; Cha, S.-H.; Ahn, G.; Jeon, Y.-J.; Kim, K.-N. Protective Effect of 3-Bromo-4,5-Dihydroxybenzaldehyde from Polysiphonia morrowii Harvey against Hydrogen Peroxide-Induced Oxidative Stress In Vitro and In Vivo. J. Microbiol. Biotechnol. 2019, 29, 1193–1203. [Google Scholar] [CrossRef]
- Ko, S.-C.; Ding, Y.; Kim, J.; Ye, B.-R.; Kim, E.-A.; Jung, W.-K.; Heo, S.-J.; Lee, S.-H. Bromophenol (5-bromo-3,4-dihydroxybenzaldehyde) isolated from red alga Polysiphonia morrowii inhibits adipogenesis by regulating expression of adipogenic transcription factors and AMP-activated protein kinase activation in 3T3-L1 adipocytes. Phytother. Res. 2018, 33, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.-J.; Han, S.-C.; Kang, H.-J.; Ko, G.; Yoon, W.-J.; Kang, H.-K.; Yoo, E.-S. Anti-Inflammatory Effect of 3-Bromo-4,5-Dihydroxybenzaldehyde, a Component of Polysiphonia morrowii, In Vivo and In Vitro. Toxicol. Res. 2017, 33, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-Y.; Kim, S.R.; Oh, M.-J.; Jung, S.-J.; Kang, S.Y. In Vitro antiviral activity of red alga, Polysiphonia morrowii extract and its bromophenols against fish pathogenic infectious hematopoietic necrosis virus and infectious pancreatic necrosis virus. J. Microbiol. 2011, 49, 102–106. [Google Scholar] [CrossRef]
- Choi, Y.K.; Ye, B.-R.; Kim, E.-A.; Kim, J.; Kim, M.-S.; Lee, W.W.; Ahn, G.-N.; Kang, N.; Jung, W.-K.; Heo, S.-J. Bis (3-bromo-4,5-dihydroxybenzyl) ether, a novel bromophenol from the marine red alga Polysiphonia morrowii that suppresses LPS-induced inflammatory response by inhibiting ROS-mediated ERK signaling pathway in RAW 264.7 macrophages. Biomed. Pharmacother. 2018, 103, 1170–1177. [Google Scholar] [CrossRef]
- Kang, S.Y.; Lee, S.-Y.; Choi, J.-H.; Jung, S.-J. In vitro Anti-bacterial and Anti-scuticociliate Activities of Extract and Bromophenols of the Marine Red Alga Polysiphonia morrowii with Structure-activity Relationships. Korean J. Fish. Aquat. Sci. 2014, 47, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Kono, R.; Nakamura, M.; Nomura, S.; Kitano, N.; Kagiya, T.; Okuno, Y.; Inada, K.-I.; Tokuda, A.; Utsunomiya, H.; Ueno, M. Biological and epidemiological evidence of anti-allergic effects of traditional Japanese food ume (Prunus mume). Sci. Rep. 2018, 8, 11638. [Google Scholar] [CrossRef]
- Fukuishi, N.; Murakami, S.; Ohno, A.; Yamanaka, N.; Matsui, N.; Fukutsuji, K.; Yamada, S.; Itoh, K.; Akagi, M. Does β-Hexosaminidase Function Only as a Degranulation Indicator in Mast Cells? The Primary Role of β-Hexosaminidase in Mast Cell Granules. J. Immunol. 2014, 193, 1886–1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garman, S.C.; Wurzburg, B.A.; Tarchevskaya, S.S.; Kinet, J.-P.; Jardetzky, T.S. Structure of the Fc fragment of human IgE bound to its high-affinity receptor FcεRIα. Nature 2000, 406, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Štěrbová, D.; Matějíček, D.; Vlček, J.; Kubáň, V. Combined microwave-assisted isolation and solid-phase purification procedures prior to the chromatographic determination of phenolic compounds in plant materials. Anal. Chim. Acta 2004, 513, 435–444. [Google Scholar] [CrossRef]
- Mlcek, J.; Jurikova, T.; Skrovankova, S.; Sochor, J. Quercetin and Its Anti-Allergic Immune Response. Molecules 2016, 21, 623. [Google Scholar] [CrossRef] [PubMed]
- Mahdi, J.; Al-Musayeib, N.; Mahdi, E.; Pepper, C. Pharmacological importance of simple phenolic compounds on inflammation, cell proliferation and apoptosis with a special reference to β-D-salicin and hydroxybenzoic acid. Eur. J. Inflamm. 2013, 11, 327–336. [Google Scholar] [CrossRef]
- Tan, J.W.; Israf, D.A.; Harith, H.H.; Hashim, N.F.M.; Ng, C.H.; Shaari, K.; Tham, C.L. Anti-allergic activity of 2,4,6-trihydroxy-3-geranylacetophenone (tHGA) via attenuation of IgE-mediated mast cell activation and inhibition of passive systemic anaphylaxis. Toxicol. Appl. Pharmacol. 2017, 319, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Zhang, Y.; Ding, X.-R.; Chen, S.-H.; Yang, J.; Wang, X.-J.; Jia, G.-L.; Chen, H.-S.; Bo, X.-C.; Wang, S.-Q. Protocatechuic aldehyde inhibits hepatitis B virus replication both in vitro and in vivo. Antivir. Res. 2007, 74, 59–64. [Google Scholar] [CrossRef]
- Guo, R.H.; Park, J.U.; Jo, S.J.; Ahn, J.H.; Park, J.H.; Yang, J.Y.; Lee, S.S.; Park, M.J.; Kim, Y.R. Anti-allergic Inflammatory Effects of the Essential Oil From Fruits of Zanthoxylum coreanum Nakai. Front. Pharmacol. 2018, 9, 1441. [Google Scholar] [CrossRef]
- Vo, T.-S.; Ngo, D.-H.; Kim, S.-K. Marine algae as a potential pharmaceutical source for anti-allergic therapeutics. Process Biochem. 2012, 47, 386–394. [Google Scholar] [CrossRef]
- Williams, C.M.; Galli, S.J. The diverse potential effector and immunoregulatory roles of mast cells in allergic disease. J. Allergy Clin. Immunol. 2000, 105, 847–859. [Google Scholar] [CrossRef]
- Gessner, A.; Mohrs, K.; Mohrs, M. Mast Cells, Basophils, and Eosinophils Acquire Constitutive IL-4 and IL-13 Transcripts during Lineage Differentiation That Are Sufficient for Rapid Cytokine Production. J. Immunol. 2005, 174, 1063–1072. [Google Scholar] [CrossRef] [Green Version]
- McLeod, J.J.; Baker, B.; Ryan, J.J. Mast cell production and response to IL-4 and IL-13. Cytokine 2015, 75, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Robinson, D.S. Th-2 cytokines in allergic disease. Br. Med. Bull. 2000, 56, 956–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- A Neveu, W.; Allard, J.L.; Raymond, D.M.; Bourassa, L.M.; Burns, S.M.; Bunn, J.Y.; Irvin, C.G.; Kaminsky, D.A.; Rincon, M. Elevation of IL-6 in the allergic asthmatic airway is independent of inflammation but associates with loss of central airway function. Respir. Res. 2010, 11, 28. [Google Scholar] [CrossRef] [Green Version]
- Debnath, T.; Lee, Y.M.; Lim, J.H.; Lim, B.O. Anti-allergic and anti-atopic dermatitis effects of Gardenia Fructus extract. Food Agric. Immunol. 2018, 29, 665–674. [Google Scholar] [CrossRef]
- Sohn, E.-H.; Jang, S.-A.; Joo, H.; Park, S.; Kang, S.-C.; Lee, C.-H.; Kim, S.-Y. Anti-allergic and anti-inflammatory effects of butanol extract from Arctium Lappa L. Clin. Mol. Allergy 2011, 9, 4. [Google Scholar] [CrossRef] [Green Version]
- Syafni, N.; Putra, D.P.; Arbain, D. 3,4-dihydroxybenzoic acid and 3,4-dihydroxybenzaldehyde from the fern Trichomanes Chinense L.; Isolation, antimicrobial and antioxidant Properties. Indones. J. Chem. 2012, 12, 273–278. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.; Tanaka, A.; Fujita, H.; Matsuda, A.; Oida, K.; Karasawa, K.; Okamoto, N.; Ohmori, K.; Jee, Y.; Shin, T.; et al. Peroxisome proliferator–activated receptor γ–mediated suppression of dendritic cell function prevents the onset of atopic dermatitis in NC/Tnd mice. J. Allergy Clin. Immunol. 2011, 127, 420–429.e6. [Google Scholar] [CrossRef]
- Kim, D.K.; Kim, H.S.; Kim, A.-R.; Kim, J.H.; Kim, B.; Noh, G.; Kim, H.S.; Beaven, M.A.; Kim, Y.M.; Choi, W.S. DJ-1 regulates mast cell activation and IgE-mediated allergic responses. J. Allergy Clin. Immunol. 2012, 131, 1653–1662.e1. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, E.-A.; Han, E.-J.; Kim, J.; Fernando, I.P.S.; Oh, J.-Y.; Kim, K.-N.; Ahn, G.; Heo, S.-J. Anti-Allergic Effect of 3,4-Dihydroxybenzaldehyde Isolated from Polysiphonia morrowii in IgE/BSA-Stimulated Mast Cells and a Passive Cutaneous Anaphylaxis Mouse Model. Mar. Drugs 2022, 20, 133. https://doi.org/10.3390/md20020133
Kim E-A, Han E-J, Kim J, Fernando IPS, Oh J-Y, Kim K-N, Ahn G, Heo S-J. Anti-Allergic Effect of 3,4-Dihydroxybenzaldehyde Isolated from Polysiphonia morrowii in IgE/BSA-Stimulated Mast Cells and a Passive Cutaneous Anaphylaxis Mouse Model. Marine Drugs. 2022; 20(2):133. https://doi.org/10.3390/md20020133
Chicago/Turabian StyleKim, Eun-A, Eui-Jeong Han, Junseong Kim, Ilekuttige Priyan Shanura Fernando, Jae-Young Oh, Kil-Nam Kim, Ginnae Ahn, and Soo-Jin Heo. 2022. "Anti-Allergic Effect of 3,4-Dihydroxybenzaldehyde Isolated from Polysiphonia morrowii in IgE/BSA-Stimulated Mast Cells and a Passive Cutaneous Anaphylaxis Mouse Model" Marine Drugs 20, no. 2: 133. https://doi.org/10.3390/md20020133