
Micrococcin P1 and P2 from Epibiotic Bacteria Associated with Isolates of Moorea producens from Kenya


Abstract
:1. Introduction
2. Results and Discussion
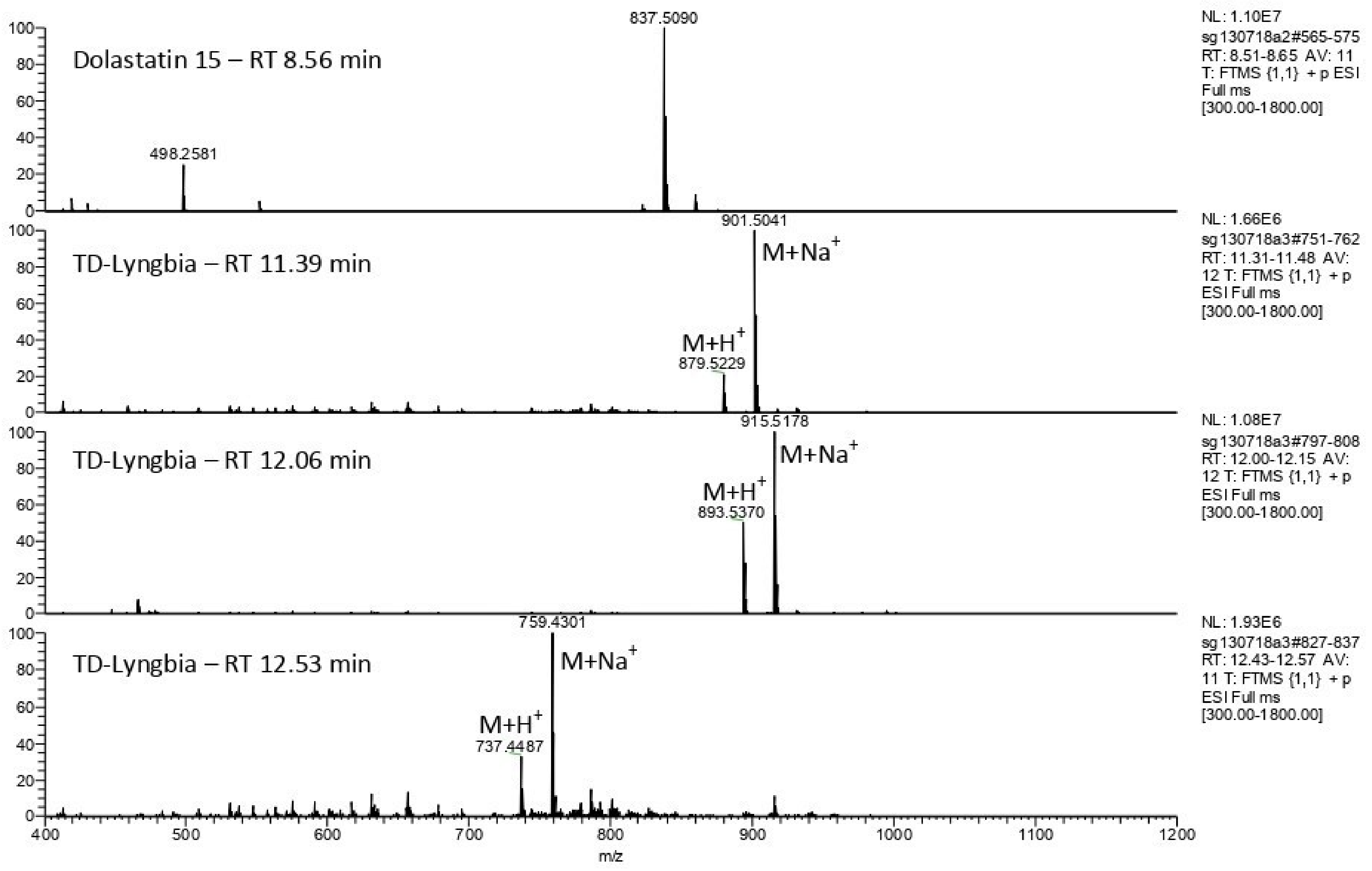
2.1. Detection of Homodolastatin 16, Dolastatin 16, and Antanapeptin A in Kenyan Isolates of a Filamentous Cyanobacterium, Moorea producens
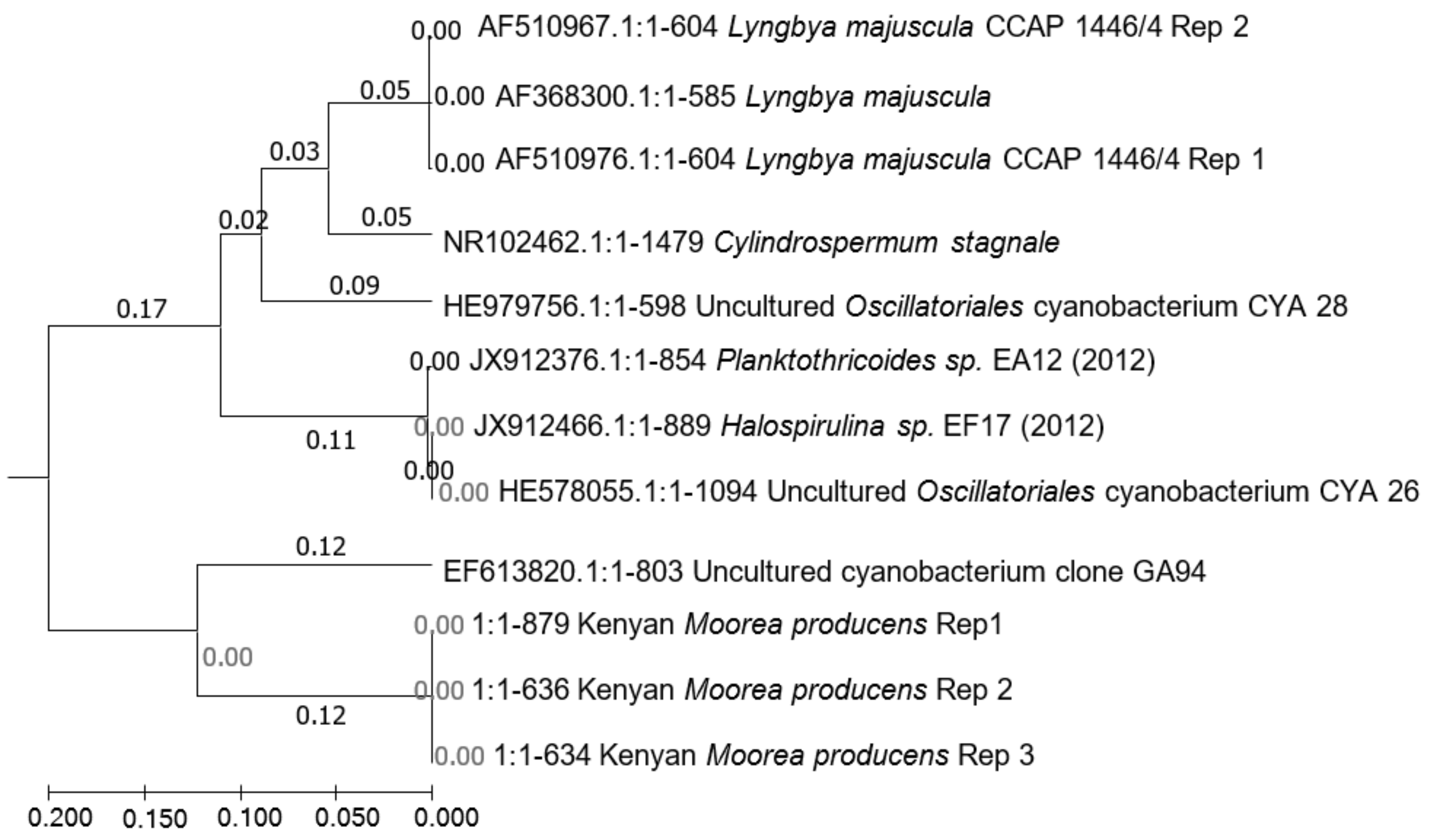
2.2. Phylogenetic Divergence of M. producens from L. majuscula
2.3. Bacterial Isolates from M. producens’ Filaments
2.4. Isolation of Bacteria from M. producens and Their Antimicrobial Activities
2.4.1. Isolation and Identification of Bacteria from M. producens’ Sheath
2.4.2. Isolation of Brightly Colored Bacteria from M. producens’ Sheath during Neap Tide
2.4.3. Antimicrobial Activities of Organic Extracts of Bacterial Isolates
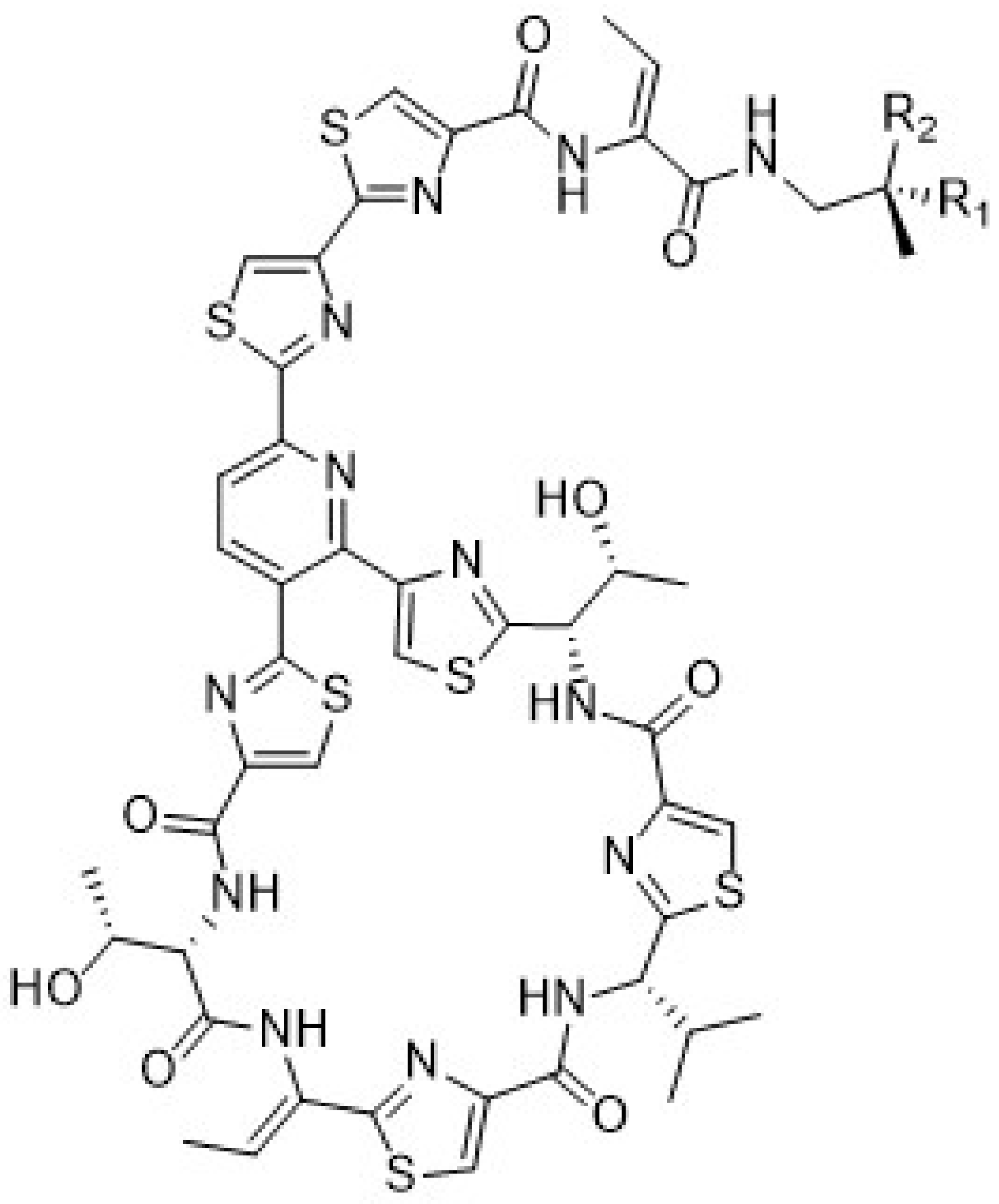
2.5. Isolation of Micrococcin P1 and Micrococcin P2 and Biological Activities
2.5.1. Isolation of Micrococcin P1 (1) and Micrococcin P2 (2)
2.5.2. Biological Activity of Micrococcin P1 against Staphylococcus aureus
3. Experimental
3.1. Study Area and Specimen Preparation
3.2. Detection of Homodolastatin 16, Dolastatin 16, and Antanapeptin A in Kenyan Isolates of a Filamentous Cyanobacterium, Moorea producens
3.3. CuSO4·5H2O Assisted Differential Genomic DNA (gDNA) Extraction of M. producens
3.4. Isolation of Bacteria from M. producens’ Filaments
3.5. The 16S rDNA Identification of Bacterial Isolates
3.6. Antibacterial and Antifungal Assays
3.7. Isolation of Micrococcin P1 (1) and Micrococcin P2 (2)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- WHO Report on Surveillance of Antibiotic Consumption 2016–2018 Early Implementation; WHO: Geneva, Switzerland, 2018.
- Demain, A.L.; Sanchez, S. Microbial drug discovery: 80 years of progress. J. Antibiot. 2009, 62, 5–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchings, M.I.; Truman, A.W.; Wilkinson, B. Antibiotics: Past, present and future. Curr. Opin. Microbiol. 2019, 51, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Ball, S.; Bessell, C.J.; Mortimer, A. The Production of Polyenic Antibiotics by Soil Streptomycetes. J. Gen. Microbiol. 1957, 17, 96–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, S.; Adachi, K.; Matsuo, Y.; Nukina, M.; Shizuri, Y. Salinisporamycin, a novel metabolite from Salinispora arenicola. J. Antibiot. 2009, 62, 519–526, Erratum in J. Antibiot. 2009, 62, 537. [Google Scholar] [CrossRef]
- Jensen, P.R.; Moore, B.S.; Fenical, W. The marine actinomycete genus Salinispora: A model organism for secondary metabolite discovery. Nat. Prod. Rep. 2015, 32, 738–751. [Google Scholar] [CrossRef] [Green Version]
- Takizawa, M.; Colwell, R.R.; Hill, R. Isolation and Diversity of Actinomycetes in the Chesapeake Bay. Appl. Environ. Microbiol. 1993, 59, 997–1002. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, T.; Müller, S.; Nadmid, S.; Garcia, R.; Müller, R. Microsclerodermins from terrestrial myxobacteria: An intriguing biosynthesis likely connected to a sponge symbiont. J. Am. Chem. Soc. 2013, 135, 16904–16911. [Google Scholar] [CrossRef]
- Gemperlein, K.; Zaburannyi, N.; Garcia, R.; La Clair, J.J.; Müller, R. Metabolic and Biosynthetic Diversity in Marine Myxobacteria. Mar. Drugs 2018, 16, 314. [Google Scholar] [CrossRef] [Green Version]
- Nijland, R.; Hall, M.; Burgess, J.G. Dispersal of Biofilms by Secreted, Matrix Degrading, Bacterial DNase. PLoS ONE 2010, 5, e15668. [Google Scholar] [CrossRef] [Green Version]
- Tomescu, A.M.; Honegger, R.; Rothwell, G.W. Earliest fossil record of bacterial-cyanobacterial mat consortia: The early Silurian Passage Creek biota (440 Ma, Virginia, USA). Geobiology 2008, 6, 120–124. [Google Scholar] [CrossRef]
- Praveen Kumar, R.; Vijayan, D.; Leo Antony, M.; Muthu Kumar, C.; Thajuddin, N. Phylogenetic diversity of cultivable bacteria associated with filamentous non-hetrocystous marine cyanobacteria. J. Algal Biomass Util. 2009, 1, 86–101. [Google Scholar]
- Engene, N.; Rottacker, E.C.; Kaštovský, J.; Byrum, T.; Choi, H.; Ellisman, M.H.; Komárek, J.; Gerwick, W.H. Moorea producens gen. nov., sp. nov. and Moorea bouillonii comb. nov., tropical marine cyanobacteria rich in bioactive secondary metabolites. Int. J. Syst. Evol. Microbiol. 2012, 62, 1171–1178. [Google Scholar] [CrossRef]
- Omarova, E.O.; Zenova, G.M.; Orleanskii, V.K.; Lobakova, E.S. Experimental associations of cyanobacteria and actinomycetes. Mosc. Univ. Biol. Sci. Bull. 2007, 62, 1–6. [Google Scholar] [CrossRef]
- Gil-Turnes, M.S. Antimicrobial Metabolites Produced by Epibiotic Bacteria: Their Role in Microbial Competition and Host Defense. Ph.D. Thesis, University of California at San Diego, San Diego, CA, USA, 1988. [Google Scholar]
- Orjala, J.; Gerwick, W.H. Two quinoline alkaloids from the caribbean cyanobacterium Lyngbya majuscula. Phytochemistry 1997, 45, 1087–1090. [Google Scholar] [CrossRef]
- Cummings, S.L.; Barbé, D.; Leao, T.F.; Korobeynikov, A.; Engene, N.; Glukhov, E.; Gerwick, W.H.; Gerwick, L. A novel uncultured heterotrophic bacterial associate of the cyanobacterium Moorea producens JHB. BMC Microbiol. 2016, 16, 198. [Google Scholar] [CrossRef] [Green Version]
- Harr, K.E.; Szabo, N.J.; Cichra, M.; Phlips, E.J. Debromoaplysiatoxin in Lyngbya-dominated mats on manatees (Trichechus manatus latirostris) in the Florida King’s Bay ecosystem. Toxicon 2008, 52, 385–388. [Google Scholar] [CrossRef]
- Dzeha, T.; Nyiro, C.; Kardasopoulos, D.; Mburu, D.; Mwafaida, J.; Hall, M.J.; Burgess, J.G. UV Resistance of bacteria from the Kenyan Marine cyanobacterium Moorea producens. MicrobiologyOpen 2019, 8, e00697. [Google Scholar] [CrossRef]
- Tronholm, A.; Engene, N. Moorena gen. nov., a valid name for “Moorea Engene & al.” nom. inval. (Oscillatoriaceae, Cyanobacteria). Not. Algarum 2019, 122, 1–2. [Google Scholar]
- Davies-Coleman, M.T.; Dzeha, T.M.; Gray, C.A.; Hess, S.; Pannell, L.K.; Hendricks, D.T.; Arendse, C.E. Isolation of Homodolastatin 16, a New Cyclic Depsipeptide from a Kenyan Collection of Lyngbya majuscula. J. Nat. Prod. 2003, 66, 712–715. [Google Scholar] [CrossRef]
- Engene, N.; Gerwick, W.H. Intra-genomic 16S rRNA gene heterogeneity in cyanobacterial genomes. Fottea 2011, 11, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Engene, N.; Gunasekera, S.P.; Gerwick, W.H.; Paul, V.J. Phylogenetic inferences reveal a large extent of novel biodiversity in chemically rich tropical marine cyanobacteria. Appl. Environ. Microbiol. 2013, 79, 1882–1888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, A.C.; Monroe, E.A.; Podell, S.; Hess, W.R.; Klages, S.; Esquenazi, E.; Niessen, S.; Hoover, H.; Rothmann, M.; Lasken, R.S.; et al. Genomic insights into the physiology and ecology of the marine filamentous cyanobacterium Lyngbya majuscula. Proc. Natl. Acad. Sci. USA 2011, 108, 8815–8820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lasken, R.S. Single-cell genomic sequencing using Multiple Displacement Amplification. Curr. Opin. Microbiol. 2007, 10, 510–516. [Google Scholar] [CrossRef] [PubMed]
- Vaara, T.; Vaara, M.; Niemela, S. Two improved methods for obtaining axenic cultures of cyanobacteria. Appl. Environ. Microbiol. 1979, 38, 1011–1014. [Google Scholar] [CrossRef] [Green Version]
- Grass, G.; Rensing, C.; Solioz, M. Metallic Copper as an Antimicrobial Surface. Appl. Environ. Microbiol. 2011, 77, 1541–1547. [Google Scholar] [CrossRef] [Green Version]
- Dollwet, H.H.A.; Sorenson, J.R.J. Historic uses of copper compounds in medicine. Trace Elem. Med. 1985, 2, 80–87. [Google Scholar]
- Nubel, U.; Garcia-Pichel, F.; Muyzer, G. PCR primers to amplify 16S rRNA genes from cyanobacteria. Appl. Environ. Microbiol. 1997, 63, 3327–3332. [Google Scholar] [CrossRef] [Green Version]
- Whitton, B.; Durham University, Durham, UK. Personal communication, 2013.
- Rainey, F.A.; Ward-Rainey, N.; Kroppenstedt, R.M.; Stackebrandt, E. The genus Nocardiopsis represents a phylogenetically coherent taxon and a distinct actinomy-cete lineage: Proposal of Nocardiopsaceae fam. nov. Int. J. Syst. Bacteriol. 1996, 46, 1088–1092. [Google Scholar] [CrossRef]
- Lechevalier, M.P.; De Bievre, C.; Lechevalier, H. Chemotaxonomy of aerobic Actinomycetes: Phospholipid composition. Biochem. Syst. Ecol. 1977, 5, 249–260. [Google Scholar] [CrossRef]
- Harker, M.; Hirschberg, J.; Oren, A. Paracoccus marcusii sp. nov., an orange Gram-negative coccus. Int. J. Syst. Bacteriol. 1998, 48, 543–548. [Google Scholar] [CrossRef]
- Su, T.L. Micrococcin, an antibacterial substance formed by a strain of Micrococcus. Br. J. Exp. Pathol. 1948, 29, 473–481. [Google Scholar]
- Bycroft, B.W.; Gowland, M.S. The structures of the highly modified peptide antibiotics micrococcin P1 and P2. J. Chem. Soc. Chem. Commun. 1978, 1978, 256–258. [Google Scholar] [CrossRef]
- Lefranc, D.; Ciufolini, M.A. Total Synthesis and Stereochemical Assignment of Micrococcin P1. Angew. Chem. Int. Ed. 2009, 48, 4198–4201. [Google Scholar] [CrossRef]
- Walsh, C.T.; Acker, M.G.; Bowers, A.A. Thiazolyl peptide antibiotic biosynthesis: A cascade of post-translational modifications on ribosomal nascent proteins. J. Biol. Chem. 2010, 285, 27525–27531. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Park, K.-H.; Lee, J.; Oh, E.; Park, C.; Kang, E.; Lee, J.; Kang, H. A New Thiopeptide Antibiotic, micrococcin P3, from a marine-derived Strain of the bacterium Bacillus stratosphericus. Molecules 2020, 25, 4383. [Google Scholar] [CrossRef]
- Kranjec, C.; Ovchinnikov, K.V.; Grønseth, T.; Ebineshan, K.; Srikantam, A.; Diep, D.B. A bacteriocin-based antimicrobial formulation to effectively disrupt the cell viability of methicillin-resistant Staphylococcus aureus (MRSA) biofilms. NPJ Biofilms Microbiomes 2020, 6, 1–13. [Google Scholar] [CrossRef]
- Jansen, R.; Sood, S.; Huch, V.; Kunze, B.; Stadler, M.; Müller, R. Pyrronazols, Metabolites from the Myxobacteria Nannocystis pusilla and N. exedens, Are Unusual Chlorinated Pyrone-Oxazole-Pyrroles. J. Nat. Prod. 2014, 77, 320–326. [Google Scholar] [CrossRef]
- Okanya, P.W.; Mohr, K.I.; Gerth, K.; Jansen, R.; Müller, R. Marinoquinolines A–F, Pyrroloquinolines from Ohtaekwangia kribbensis (Bacteroidetes). J. Nat. Prod. 2011, 74, 603–608. [Google Scholar] [CrossRef]
- Bader, C.D.; Neuber, M.; Panter, F.; Krug, D.; Müller, R. Supercritical Fluid Extraction Enhances Discovery of Secondary Metabolites from Myxobacteria. Anal. Chem. 2020, 92, 15403–15411. [Google Scholar] [CrossRef]
- Morin, N.; Vallaeys, T.; Hendrickx, L.; Natalie, L.; Wilmotte, A. An efficient DNA isolation protocol for filamentous cyanobacteria of the genus Arthrospira. J. Microbiol. Methods 2010, 80, 148–154. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F.; Bragard, C.; Mahillon, J. Overview of the Antimicrobial Compounds Produced by Members of the Bacillus subtilis Group. Front. Microbiol. 2019, 10, 302. [Google Scholar] [CrossRef] [Green Version]
- Charousová, I.; Steinmetz, H.; Medo, J.; Javoreková, S.; Wink, J. Characterization of Antimycins—Producing Streptomycete Strain VY46 Isolated from Slovak Soil. Braz. Arch. Biol. Technol. 2016, 59, e16160274. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Strain | % Similarity of 16S Sequence | Taxon | |
|---|---|---|---|---|
| Shewanella algae | KC660130 | SHALG-01 | 99 | γ-proteobacteria |
| Shewanella algae | KC660131 | SHALG-02 | 99 | γ-proteobacteria |
| Marinobacterium stanieri | KC660132 | MARIS-01 | 99 | γ-proteobacteria |
| Acinetobacter johnsonii | KC660133 | ACJ-01 | 99 | γ-proteobacteria |
| Marinobacterium stanieri | KC660134 | MARIS-02 | 99 | γ-proteobacteria |
| Staphylococcus saprophyticus | KC660135 | STAPRO | 99 | Firmicutes |
| Pseudomonas stutzeri | KC660136 | PST-01 | 99 | γ-proteobacteria |
| Enterobacter cloacae | KC660137 | ENTCLO | 99 | γ-proteobacteria |
| Cellulosimicrobium cellulans | KC660138 | CCL-01 | 99 | Actinobacteria |
| Cellulosimicrobium cellulans | KC660139 | CCL-02 | 99 | Actinobacteria |
| Pseudomonas pseudoalcaligenes | KC660140 | PPS | 99 | γ-proteobacteria |
| Pseudomonas putida | KC660141 | PPT | 99 | γ-proteobacteria |
| Bacillus aereus | ND | ND | 99 | Firmicutes |
| Bacillus licheniformis | KC660142 | BLC-01 | 99 | Firmicutes |
| Bacillus licheniformi | KC660143 | BLC-02 | 99 | Firmicutes |
| Bacillus subtilis | KC660144 | BS-00 | 99 | Firmicutes |
| Pseudomonas stutzeri | KC660145 | PST-02 | 99 | γ-proteobacteria |
| Enterobacter cancerogenus | ND | ND | 99 | γ-proteobacteria |
| Klebsiella oxytoca | ND | ND | 99 | γ-proteobacteria |
| Yokenella regensburgei | ND | ND | 99 | γ-proteobacteria |
| Ochrobactrumanthropi | ND | ND | 99 | α-proteobacteria |
| Pseudomonas stutzeri | ND | ND | 99 | γ-proteobacteria |
| Pseudoalteromonas carrageenovora | ND | ND | 99 | γ-proteobacteria |
| Bacterial Strains | Closest Match in GenBank (Accession Number) | % Similarity of 16S Sequence | Antimicrobial Activity | Level of Inhibition |
|---|---|---|---|---|
| TD1, TD15 | Bacillus marislavi JC556 (LS974830.1) | 100 | M. luteus, B. subtilis | Active |
| TD2, TD26 | Bacillus marislavi LQ1 (MG025780.1) | 100 | B. subtilis | Minor activity |
| TD3, TD6 | Bacillus safensis 6-11 (MK205159.1) | 100 | B. subtilis | Minor activity |
| TD10 | Bacillus safensis 6-11 (MK210556.1) | 100 | Not Done | |
| TD11 | Bacillus safensis D11 (KX068630.1) | 100 | B. subtilis, P. anomalis | Minor activity |
| TD23, 25 | Bacillus safensis D11 MK337676.1 | 100 | M. luteus, B. subtilis | Minor activity |
| TD4 | Bacillus aryabhattai PYMW (MK346120.1) | 100 | B. subtilis, P. anomalis | Minor activity |
| TD8 | Bacillus aryabhattai P6 (MK346850.1) | 100 | B. subtilis | Minor activity |
| TD5, TD22 | Bacillus licheniformis PB3 (CP025226.1) | 99 | E. coli, B. subtilis | Moderately active |
| TD7 | Bacillus subtilis MJ01 | 100 | E. coli, M. luteus | Very active |
| TD12 | Bacillus subtilis TBS-CBE-BS01 (MK346244.1) | 100 | M. luteus | Minor activity |
| TD9 | Sequence failed | Not done | B. subtilis | Minor activity |
| TD13 | Arthrobacter sp. ABCH 95.B (KY327809.1) | 100 | NA | No activity |
| TD14 | Bacillus flexus 00F26 MH542283.1 | Not done | NA | No activity |
| Molecular Ion | Expected m/z (a.m.u) | Observed m/z (a.m.u) | |
|---|---|---|---|
| 1 | M + H | 1144.21732 | 1144.21930 |
| 2 | M − H2O + H | 1126.20676 | 1126.20946 |
| 3 | M − CO | 1116.22241 | 1116.22442 |
| 4 | M − 2H2O + H | 1108.19619 | 1108.19865 |
| 5 | M − CO − H2O | 1098.21184 | 1098.21380 |
| 6 | M − CO − 2H2O | 1080.20128 | 1080.20333 |
| 7 | b13 + H2O | 1051.13834 | 1051.14040 |
| 8 | a13 | 1041.15399 | 1041.15222 |
| 9 | b13 − 2H2O | 1033.12778 | 1033.12980 |
| 10 | b2 + H2O | 1025.15908 | 1025.16113 |
| 11 | a13 − H2O | 1023.14343 | 1023.14492 |
| 12 | b2 − 2H2O | 1007.14851 | 1007.15051 |
| 13 | a13 − 2H2O | 1005.13286 | 1005.13482 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dzeha, T.; Hall, M.J.; Burgess, J.G. Micrococcin P1 and P2 from Epibiotic Bacteria Associated with Isolates of Moorea producens from Kenya. Mar. Drugs 2022, 20, 128. https://doi.org/10.3390/md20020128
Dzeha T, Hall MJ, Burgess JG. Micrococcin P1 and P2 from Epibiotic Bacteria Associated with Isolates of Moorea producens from Kenya. Marine Drugs. 2022; 20(2):128. https://doi.org/10.3390/md20020128
Chicago/Turabian StyleDzeha, Thomas, Michael John Hall, and James Grant Burgess. 2022. "Micrococcin P1 and P2 from Epibiotic Bacteria Associated with Isolates of Moorea producens from Kenya" Marine Drugs 20, no. 2: 128. https://doi.org/10.3390/md20020128