
Exogenous Arachidonic Acid Affects Fucoxanthin Biosynthesis and Photoprotection in Phaeodactylum tricornutum

Abstract
:1. Introduction
2. Results
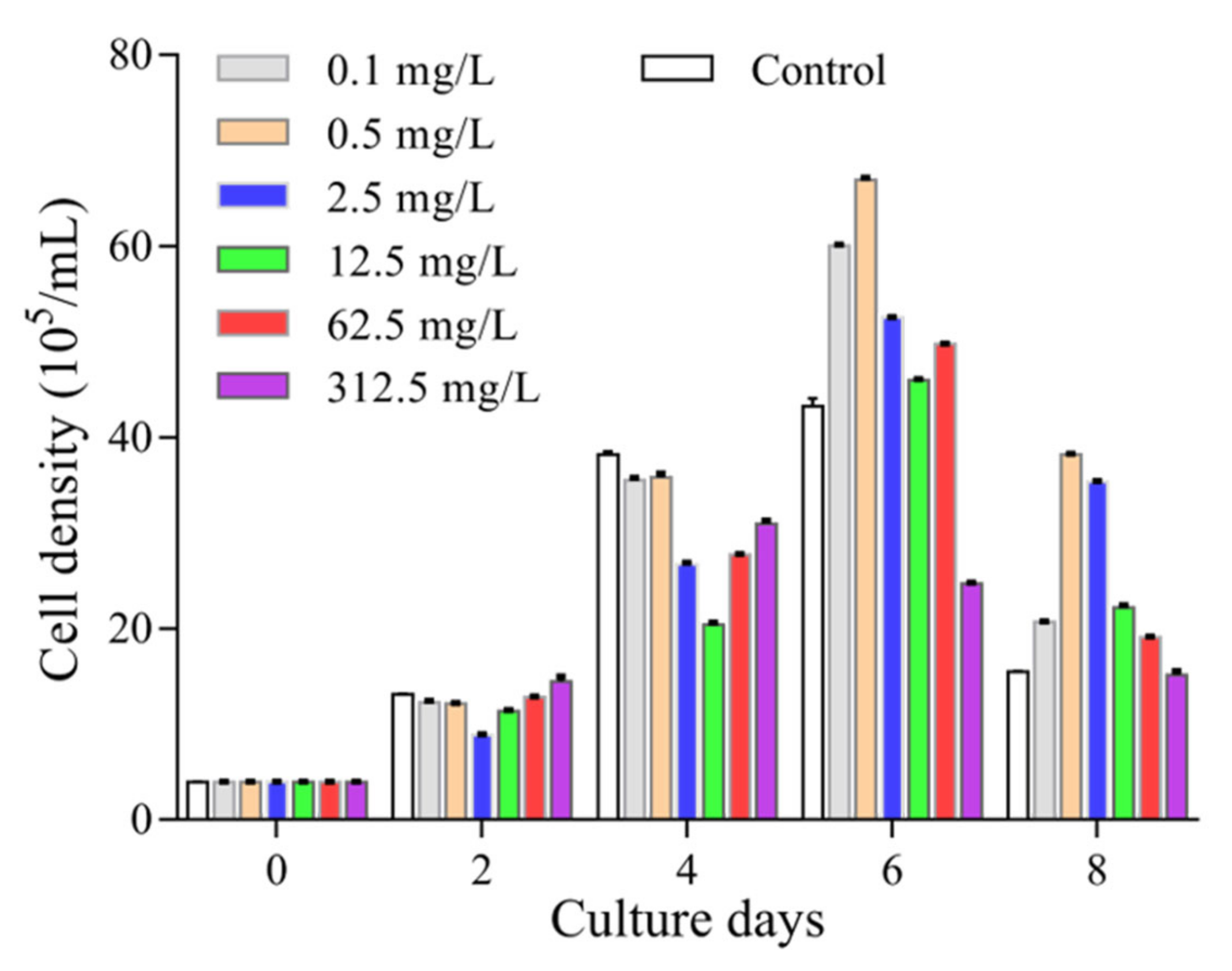
2.1. Arachidonic Acid Modulates the Growth of P. tricornutum
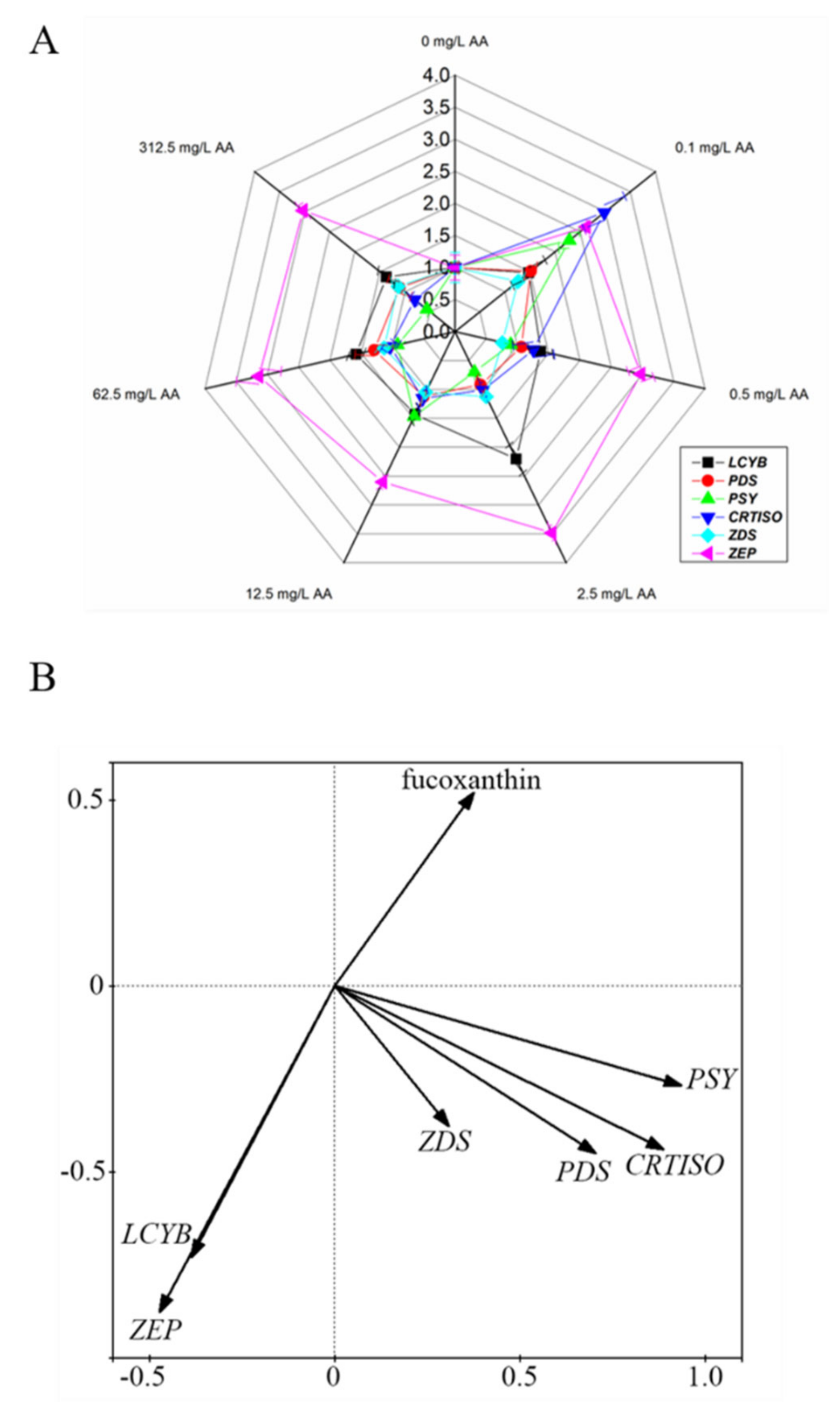
2.2. Low Level of AA Favors Fucoxanthin Production
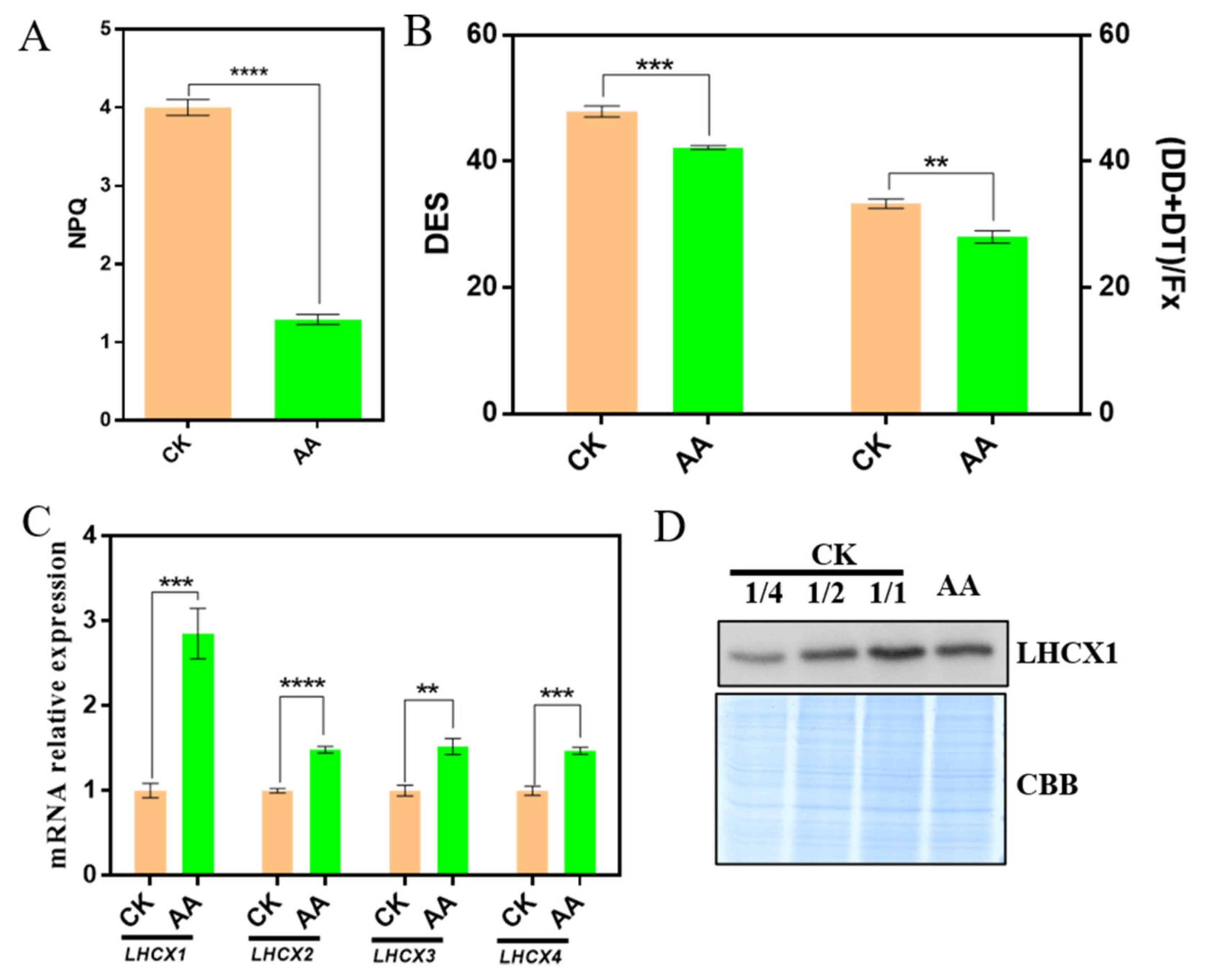
2.3. AA Affects the Expression of Carotenoid Metabolism Genes
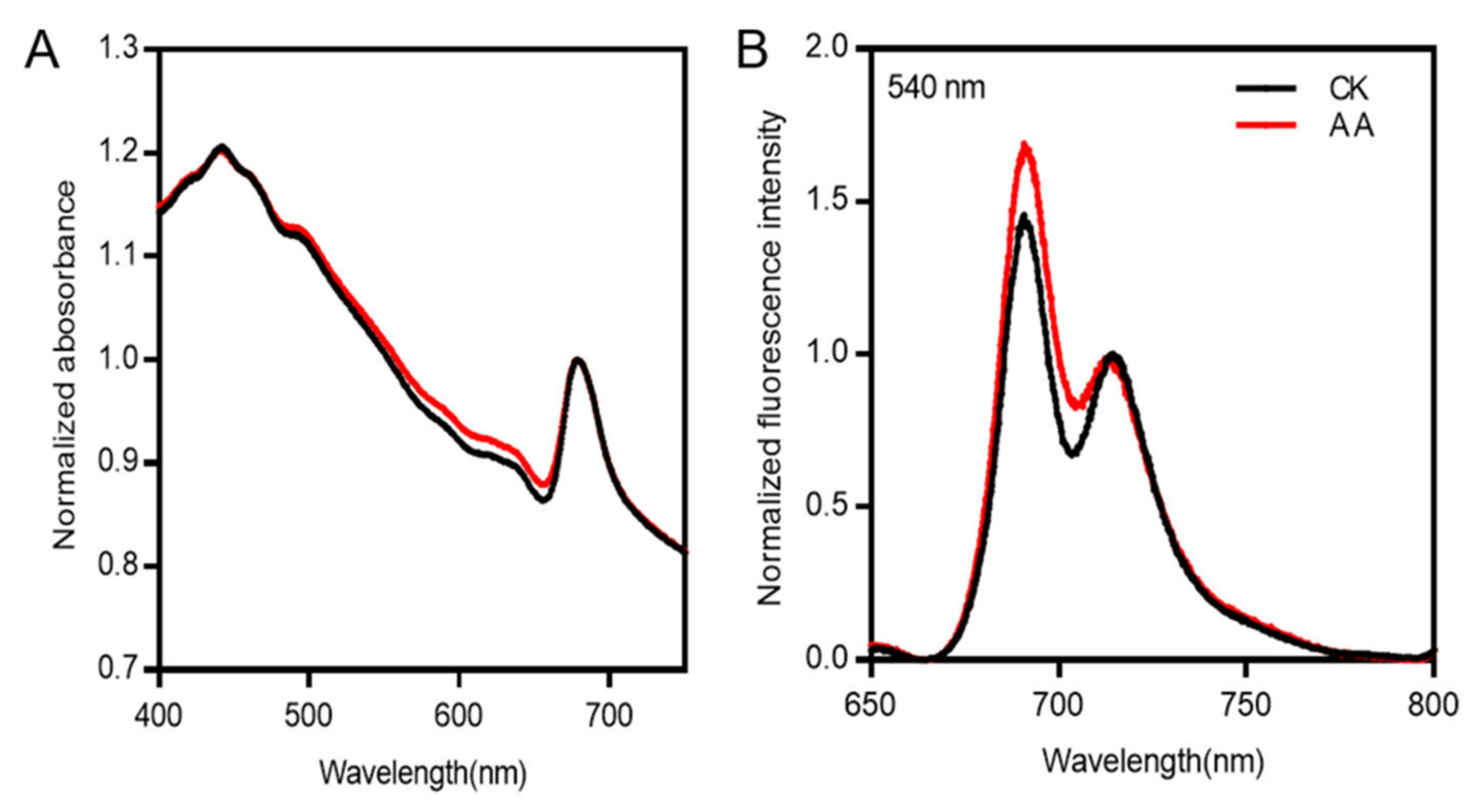
2.4. Energy Transfer Is Affected by AA Treatment in P. tricornutum
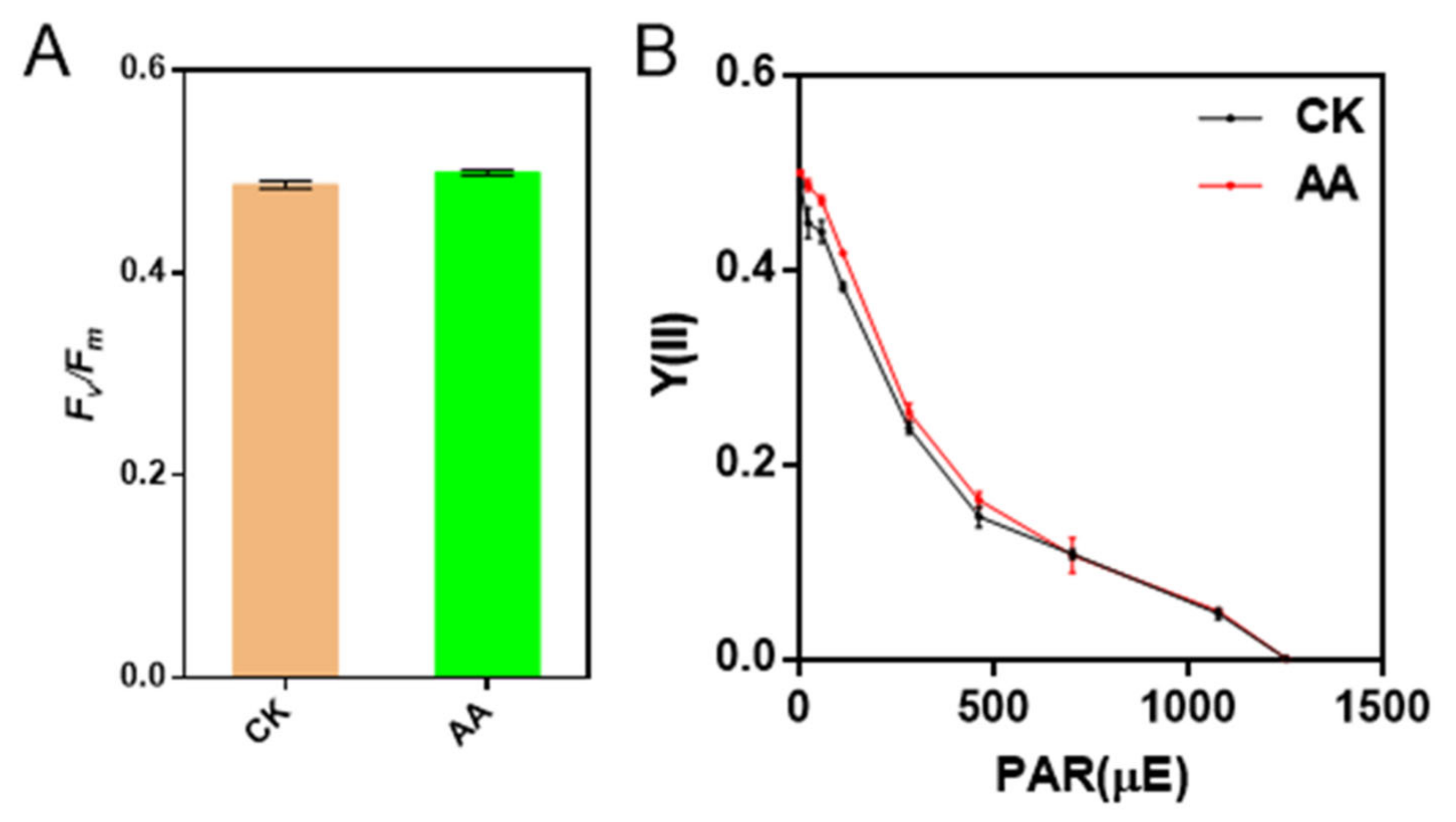
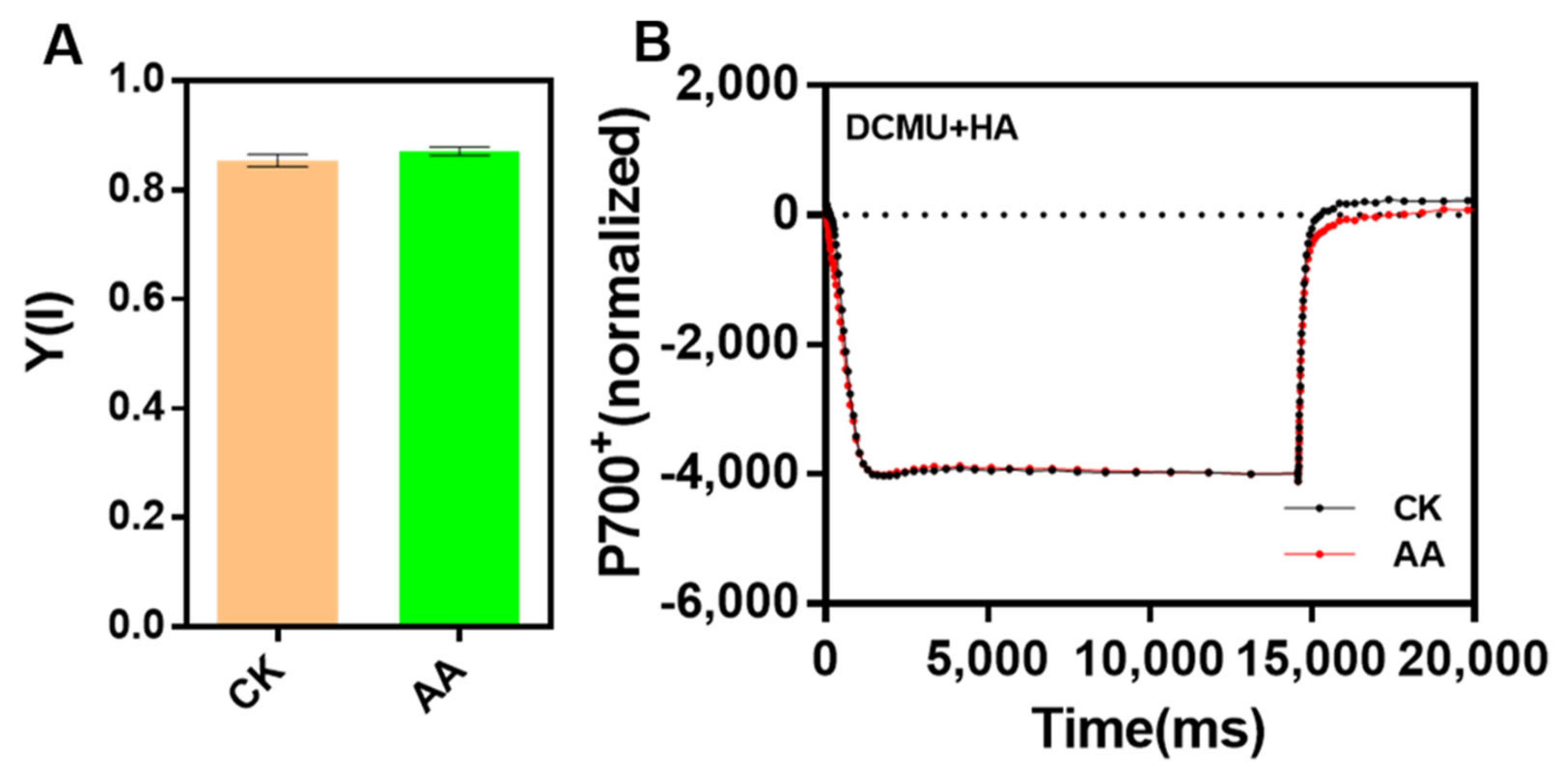
2.5. The Activities of PSII and PSI Are Not Affected by AA Treatment
2.6. Non-Photochemical Quenching Is Decreased under the Treatment of AA
2.7. Arachidonic Acid Treatment Induces Photoinhibition and Slows down Recovery of PSII Activity from Photoinhibition
3. Discussion
4. Materials and Methods
4.1. Algal Strain and Growth Conditions
4.2. Growth Measurement and Fucoxanthin Quantification
4.3. Molecular Manipulation
4.4. Gene Expression Analysis
4.5. Protein Extraction and Western Blot Analysis
4.6. Measurement of Chlorophyll Fluorescence
4.7. The Detections of PSI Activity, PSII Activity and Chl Fluorescence Emission Spectra
4.8. Photoinhibition Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cunningham, F.X.; Gantt, E. Genes and enzymes of carotenoid biosynthesis in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 557–583. [Google Scholar] [CrossRef]
- Kuczynska, P.; Jemiola-Rzeminska, M.; Strzalka, K. Photosynthetic pigments in diatoms. Mar. Drugs 2015, 13, 5847–5881. [Google Scholar] [CrossRef]
- Lu, S.; Li, L. Carotenoid metabolism: Biosynthesis, regulation, and beyond. J. Integr. Plant Biol. 2008, 50, 778–785. [Google Scholar] [CrossRef]
- Zhou, F.; Wang, C.Y.; Gutensohn, M.; Jiang, L.; Zhang, P.; Zhang, D.; Dudareva, N.; Lu, S. A recruiting protein of geranylgeranyl diphosphate synthase controls metabolic flux toward chlorophyll biosynthesis in rice. Proc. Natl. Acad. Sci. USA 2017, 114, 6866–6871. [Google Scholar] [CrossRef] [Green Version]
- Takaichi, S. Carotenoids in algae: Distributions, biosyntheses and functions. Mar. Drugs 2011, 9, 1101–1118. [Google Scholar] [CrossRef]
- Sun, T.; Yuan, H.; Cao, H.; Yazdani, M.; Tadmor, Y.; Li, L. Carotenoid metabolism in plants: The role of plastids. Mol. Plant 2018, 11, 58–74. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, F.X., Jr.; Gantt, E. Elucidation of the pathway to astaxanthin in the flowers of Adonis aestivalis. Plant Cell 2011, 23, 3055–3069. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, F.X., Jr.; Lee, H.; Gantt, E. Carotenoid biosynthesis in the primitive red alga Cyanidioschyzon merolae. Eukaryot. Cell 2007, 6, 533–545. [Google Scholar] [CrossRef] [Green Version]
- D’Orazio, N.; Gemello, E.; Gammone, M.A.; de Girolamo, M.; Ficoneri, C.; Riccioni, G. Fucoxantin: A treasure from the sea. Mar. Drugs 2012, 10, 604–616. [Google Scholar] [CrossRef] [Green Version]
- Caron, L.; Douady, D.; Quinet-Szely, M.; de Goer, S.; Berkaloff, C. Gene structure of a chlorophyll a/c-binding protein from a brown alga: Presence of an intron and phylogenetic implications. J. Mol. Evol. 1996, 43, 270–280. [Google Scholar] [CrossRef]
- Papagiannakis, E.; van Stokkum, I.H.M.; Fey, H.; Buchel, C.; van Grondelle, R. Spectroscopic characterization of the excitation energy transfer in the fucoxanthin-chlorophyll protein of diatoms. Photosynth. Res. 2005, 86, 241–250. [Google Scholar] [CrossRef]
- Veith, T.; Brauns, J.; Weisheit, W.; Mittag, M.; Buchel, C. Identification of a specific fucoxanthin-chlorophyll protein in the light harvesting complex of photosystem I in the diatom Cyclotella meneghiniana. Biochim. Biophys. Acta 2009, 1787, 905–912. [Google Scholar] [CrossRef] [Green Version]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Funayama, K.; Miyashita, K. Fucoxanthin from edible seaweed, Undaria pinnatifida, shows antiobesity effect through UCP1 expression in white adipose tissues. Biochem. Biophys. Res. Commun. 2005, 332, 392–397. [Google Scholar] [CrossRef]
- Zhao, D.; Kwon, S.H.; Chun, Y.S.; Gu, M.Y.; Yang, H.O. Anti-neuroinflammatory effects of fucoxanthin via inhibition of Akt/NF-kappaB and MAPKs/AP-1 pathways and activation of PKA/CREB pathway in lipopolysaccharide-activated BV-2 microglial cells. Neurochem. Res. 2017, 42, 667–677. [Google Scholar] [CrossRef]
- Jin, Y.; Qiu, S.; Shao, N.; Zheng, J. Fucoxanthin and tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) synergistically promotes apoptosis of human cervical cancer cells by targeting PI3K/Akt/NF-kappaB signaling pathway. Med. Sci. Monit. 2018, 24, 11–18. [Google Scholar] [CrossRef]
- Yu, R.X.; Yu, R.T.; Liu, Z. Inhibition of two gastric cancer cell lines induced by fucoxanthin involves downregulation of Mcl-1 and STAT3. Hum. Cell 2018, 31, 50–63. [Google Scholar] [CrossRef]
- Dambek, M.; Eilers, U.; Breitenbach, J.; Steiger, S.; Buchel, C.; Sandmann, G. Biosynthesis of fucoxanthin and diadinoxanthin and function of initial pathway genes in Phaeodactylum tricornutum. J. Exp. Bot. 2012, 63, 5607–5612. [Google Scholar] [CrossRef]
- Goss, R.; Lepetit, B. Biodiversity of NPQ. J. Plant Physiol. 2015, 172, 13–32. [Google Scholar] [CrossRef]
- Schuhmann, H.; Adamska, I. Deg proteases and their role in protein quality control and processing in different subcellular compartments of the plant cell. Physiol. Plant. 2012, 145, 224–234. [Google Scholar] [CrossRef]
- Kato, Y.; Hyodo, K.; Sakamoto, W. The photosystem II repair cycle requires FtsH turnover through the EngA GTPase. Plant Physiol. 2018, 178, 596–611. [Google Scholar] [CrossRef]
- Sun, K.M.; Gao, C.L.; Zhang, J.; Tang, X.X.; Wang, Z.L.; Zhang, X.L.; Li, Y. Rapid formation of antheraxanthin and zeaxanthin in seconds in microalgae and its relation to non-photochemical quenching. Photosynth. Res. 2020, 144, 317–326. [Google Scholar] [CrossRef]
- Taddei, L.; Stella, G.R.; Rogato, A.; Bailleul, B.; Fortunato, A.E.; Annunziata, R.; Sanges, R.; Thaler, M.; Lepetit, B.; Lavaud, J.; et al. Multisignal control of expression of the LHCX protein family in the marine diatom Phaeodactylum tricornutum. J. Exp. Bot. 2016, 67, 3939–3951. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Gao, Z.; Du, H.; Lin, B.; Yan, Y.; Li, G.; Guo, Y.; Fu, S.; Wei, G.; Wang, M.; et al. The effects of abscisic acid, salicylic acid and jasmonic acid on lipid accumulation in two freshwater Chlorella strains. J. Gen. Appl. Microbiol. 2018, 64, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.F.; Liao, Z.H.; Guo, B.H.; Sun, X.F.; Tang, K.X. Molecular cloning and expression profile analysis of Ginkgo biloba DXS gene encoding 1-deoxy-D-xylulose 5-phosphate synthase, the first committed enzyme of the 2-C-methyl-D-erythritol 4-phosphate pathway. Planta Med. 2006, 72, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.M.; Jung, Y.J.; Kwon, O.N.; Cha, K.H.; Um, B.H.; Chung, D.; Pan, C.H. A potential commercial source of fucoxanthin extracted from the microalga Phaeodactylum tricornutum. Appl. Biochem. Biotechnol. 2012, 166, 1843–1855. [Google Scholar] [CrossRef]
- Bowler, C.; Allen, A.E.; Badger, J.H.; Grimwood, J.; Jabbari, K.; Kuo, A.; Maheswari, U.; Martens, C.; Maumus, F.; Otillar, R.P.; et al. The Phaeodactylum genome reveals the evolutionary history of diatom genomes. Nature 2008, 456, 239–244. [Google Scholar] [CrossRef] [Green Version]
- Niu, Y.F.; Yang, Z.K.; Zhang, M.H.; Zhu, C.C.; Yang, W.D.; Liu, J.S.; Li, H.Y. Transformation of diatom Phaeodactylum tricornutum by electroporation and establishment of inducible selection marker. Biotechniques 2012, 52, 1–3. [Google Scholar] [CrossRef]
- Wang, L.J.; Fan, Y.; Parsons, R.L.; Hu, G.R.; Zhang, P.Y.; Li, F.L. A rapid method for the determination of fucoxanthin in diatom. Mar. Drugs 2018, 16, 33. [Google Scholar] [CrossRef] [Green Version]
- Wahadoszamen, M.; Ghazaryan, A.; Cingil, H.E.; Ara, A.M.; Buchel, C.; van Grondelle, R.; Berera, R. Stark fluorescence spectroscopy reveals two emitting sites in the dissipative state of FCP antennas. Biochim. Biophys. Acta 2014, 1837, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, M. Carotenoid biosynthesis in diatoms. Photosynth. Res. 2010, 106, 89–102. [Google Scholar] [CrossRef]
- Goss, R.; Ann Pinto, E.; Wilhelm, C.; Richter, M. The importance of a highly active and ΔpH-regulated diatoxanthin epoxidase for the regulation of the PSII antenna function in diadinoxanthin cycle containing algae. J. Plant Physiol. 2006, 163, 1008–1021. [Google Scholar] [CrossRef]
- Jahns, P.; Holzwarth, A.R. The role of the xanthophyll cycle and of lutein in photoprotection of photosystem II. Biochim. Biophys. Acta 2012, 1817, 182–193. [Google Scholar] [CrossRef] [Green Version]
- Alexandre, M.T.; Gundermann, K.; Pascal, A.A.; van Grondelle, R.; Buchel, C.; Robert, B. Probing the carotenoid content of intact Cyclotella cells by resonance Raman spectroscopy. Photosynth. Res. 2014, 119, 273–281. [Google Scholar] [CrossRef]
- Beer, A.; Juhas, M.; Buchel, C. Influence of different light intensities and different iron nutrition on the photosynthetic apparatus in the diatom Cyclotella Meneghiniana (bacillariophyceae). J. Phycol. 2011, 47, 1266–1273. [Google Scholar] [CrossRef]
- Yang, R.; Wei, D. Improving fucoxanthin production in mixotrophic culture of marine diatom Phaeodactylum tricornutum by LED light shift and nitrogen supplementation. Front. Bioeng. Biotechnol. 2020, 8, 820. [Google Scholar] [CrossRef]
- Erickson, E.; Wakao, S.; Niyogi, K.K. Light stress and photoprotection in Chlamydomonas reinhardtii. Plant J. 2015, 82, 449–465. [Google Scholar] [CrossRef]
- Dall’Osto, L.; Cazzaniga, S.; Havaux, M.; Bassi, R. Enhanced photoprotection by protein-bound vs free xanthophyll pools: A comparative analysis of chlorophyll b and xanthophyll biosynthesis mutants. Mol. Plant 2010, 3, 576–593. [Google Scholar] [CrossRef]
- Taddei, L.; Chukhutsina, V.U.; Lepetit, B.; Stella, G.R.; Bassi, R.; van Amerongen, H.; Bouly, J.P.; Jaubert, M.; Finazzi, G.; Falciatore, A. Dynamic changes between two LHCX-related energy quenching sites control diatom photoacclimation. Plant Physiol. 2018, 177, 953–965. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Yu, L.J.; Xu, C.; Tomizaki, T.; Zhao, S.; Umena, Y.; Chen, X.; Qin, X.; Xin, Y.; Suga, M.; et al. Structural basis for blue-green light harvesting and energy dissipation in diatoms. Science 2019, 363, eaav0365. [Google Scholar] [CrossRef]
- Miloslavina, Y.; Grouneva, I.; Lambrev, P.H.; Lepetit, B.; Goss, R.; Wilhelm, C.; Holzwarth, A.R. Ultrafast fluorescence study on the location and mechanism of non-photochemical quenching in diatoms. Biochim. Biophys. Acta 2009, 1787, 1189–1197. [Google Scholar] [CrossRef]
- Holzwarth, A.R.; Miloslavina, Y.; Nilkens, M.; Jahns, P. Identification of two quenching sites active in the regulation of photosynthetic light-harvesting studied by time-resolved fluorescence. Chem. Phys. Lett. 2009, 483, 262–267. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Fang, Y.; Jiang, Z.; Zhao, C.; Li, L.; Ranvilage, C.; Liu, S.; Wu, Y.; Huang, X. Efficient heat dissipation and cyclic electron flow confer daily air exposure tolerance in the intertidal seagrass Halophila beccarii Asch. Front. Plant Sci. 2020, 11, 571627. [Google Scholar] [CrossRef]
- Nguyen, K.; Vaughn, M.; Frymier, P.; Bruce, B.D. In vitro kinetics of P700+reduction of Thermosynechococcus elongatus trimeric Photosystem I complexes by recombinant cytochrome c6 using a Joliot-type LED spectrophotometer. Photosynth. Res. 2017, 131, 79–91. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AA Concentration (mg/L) | Content of Fucoxanthin [mg/g Dry Weight (DW)] | |
|---|---|---|
| Average | SD | |
| 0 | 0.805 a | ±0.190 |
| 0.1 | 1.099 a | ±0.205 |
| 0.5 | 0.865 a | ±0.033 |
| 2.5 | 0.775 a | ±0.156 |
| 12.5 | 0.797 a | ±0.275 |
| 62.5 | 0.774 a | ±0.095 |
| 312.5 | 0.271 b | ±0.003 |
| Gene | Best Hit Organism (Similarity) | Accession Number |
|---|---|---|
| LCYB | Fistulifera solaris (65%) | GAX21707.1 |
| PSY | Fistulifera solaris (70%) | GAX17065.1 |
| PDS | Chromochloris zofingiensis (60%) | ABR20877.1 |
| CRTISO | Fistulifera solaris (70%) | GAX26939.1 |
| ZDS | Fistulifera solaris (87%) | GAX15703.1 |
| ZEP | Fistulifera solaris (84%) | GAX11479.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, S.; Bin, S.; Wang, W.; Lu, S.; Yang, W. Exogenous Arachidonic Acid Affects Fucoxanthin Biosynthesis and Photoprotection in Phaeodactylum tricornutum. Mar. Drugs 2022, 20, 644. https://doi.org/10.3390/md20100644
Zhu S, Bin S, Wang W, Lu S, Yang W. Exogenous Arachidonic Acid Affects Fucoxanthin Biosynthesis and Photoprotection in Phaeodactylum tricornutum. Marine Drugs. 2022; 20(10):644. https://doi.org/10.3390/md20100644
Chicago/Turabian StyleZhu, Shuaiqi, Song Bin, Wenda Wang, Shan Lu, and Wenqiang Yang. 2022. "Exogenous Arachidonic Acid Affects Fucoxanthin Biosynthesis and Photoprotection in Phaeodactylum tricornutum" Marine Drugs 20, no. 10: 644. https://doi.org/10.3390/md20100644