
Anti-Inflammatory and Antioxidant Properties of Malapterurus electricus Skin Fish Methanolic Extract in Arthritic Rats: Therapeutic and Protective Effects
 , , , , , , ,
, , , , , , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Effects of Fish Extract on Ankle Swelling in MSU Crystal-Gouty Arthritis Rats
2.2. Effects of Fish Extract on Inflammatory Markers TNF-α, IL-1β, and NF-𝜅B in Synovial Tissues Homogenate
2.3. Effects of Fish Extract on Oxidative Stress and Antioxidant Markers in Synovial Tissues Homogenate
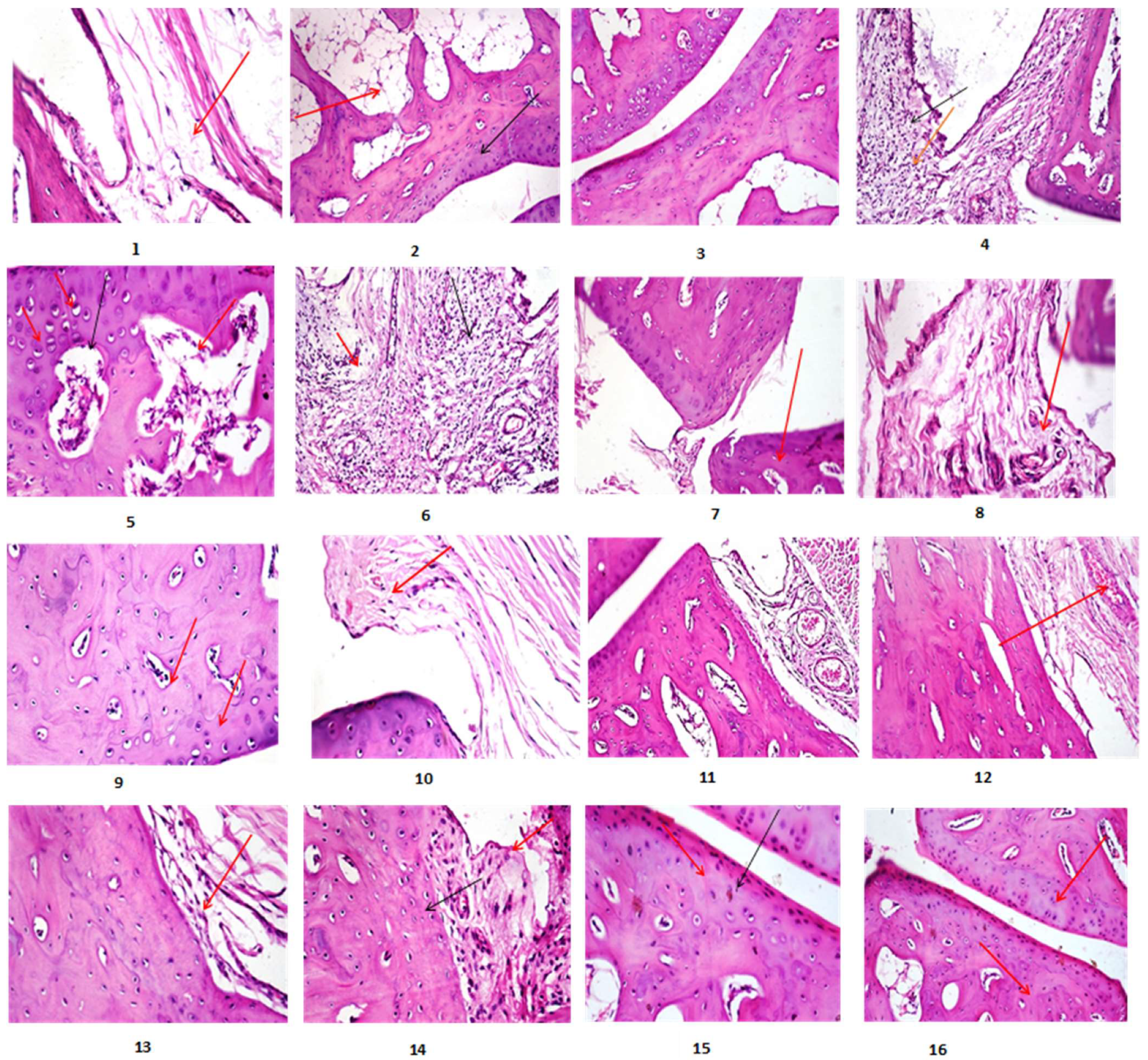
2.4. Histopathological Parameters
2.4.1. Histopathological Findings
2.4.2. Lesion Score (Scoring of Histopathological Alterations of All Treated Groups)
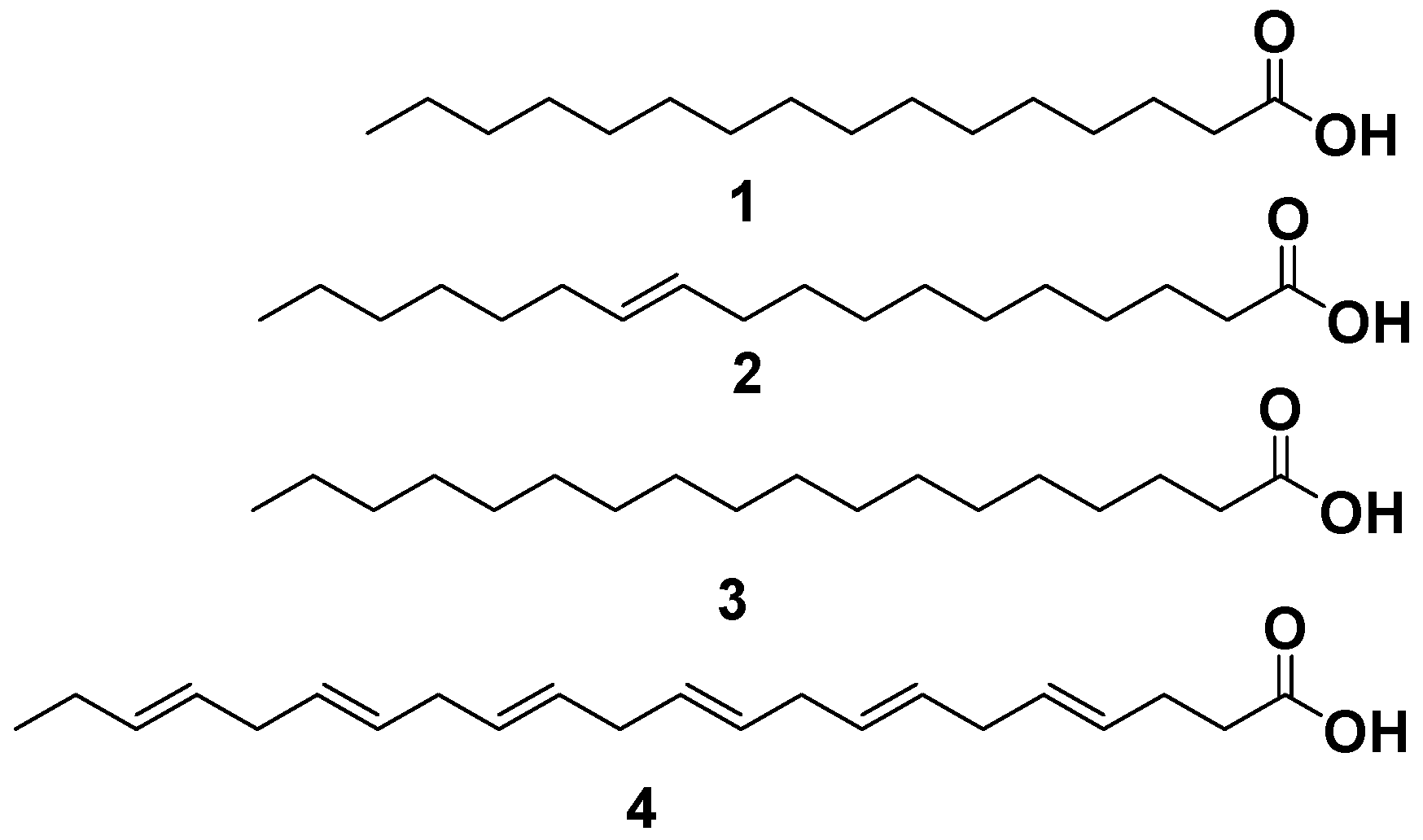
2.5. Phyto-Chemical Investigation of Malapterurus electricus Crude Extract
2.5.1. GC-MS Analysis
2.5.2. Physicochemical Investigation of Malapterurus electricus Crude Extract
2.5.3. Amino Acid Content
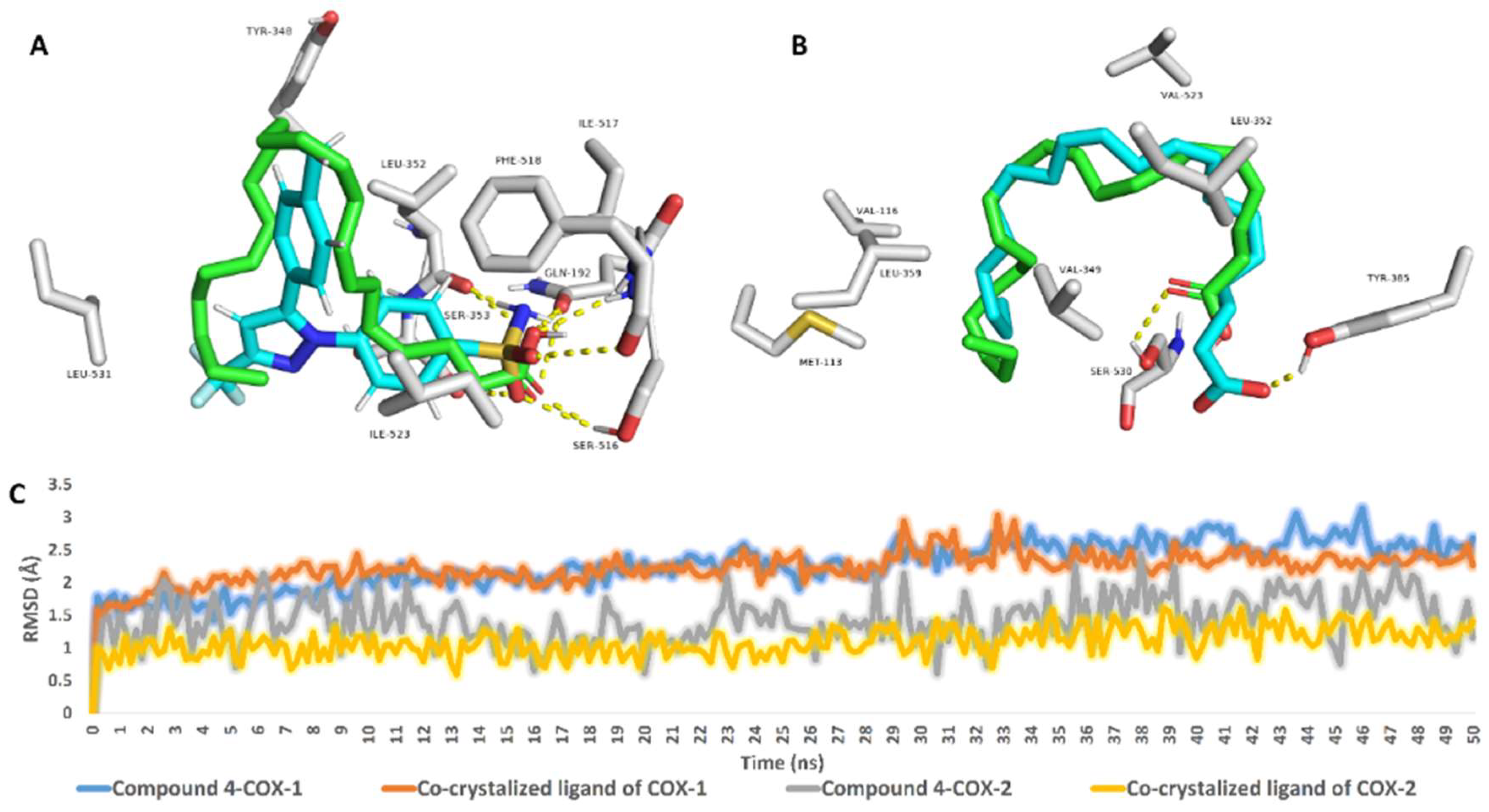
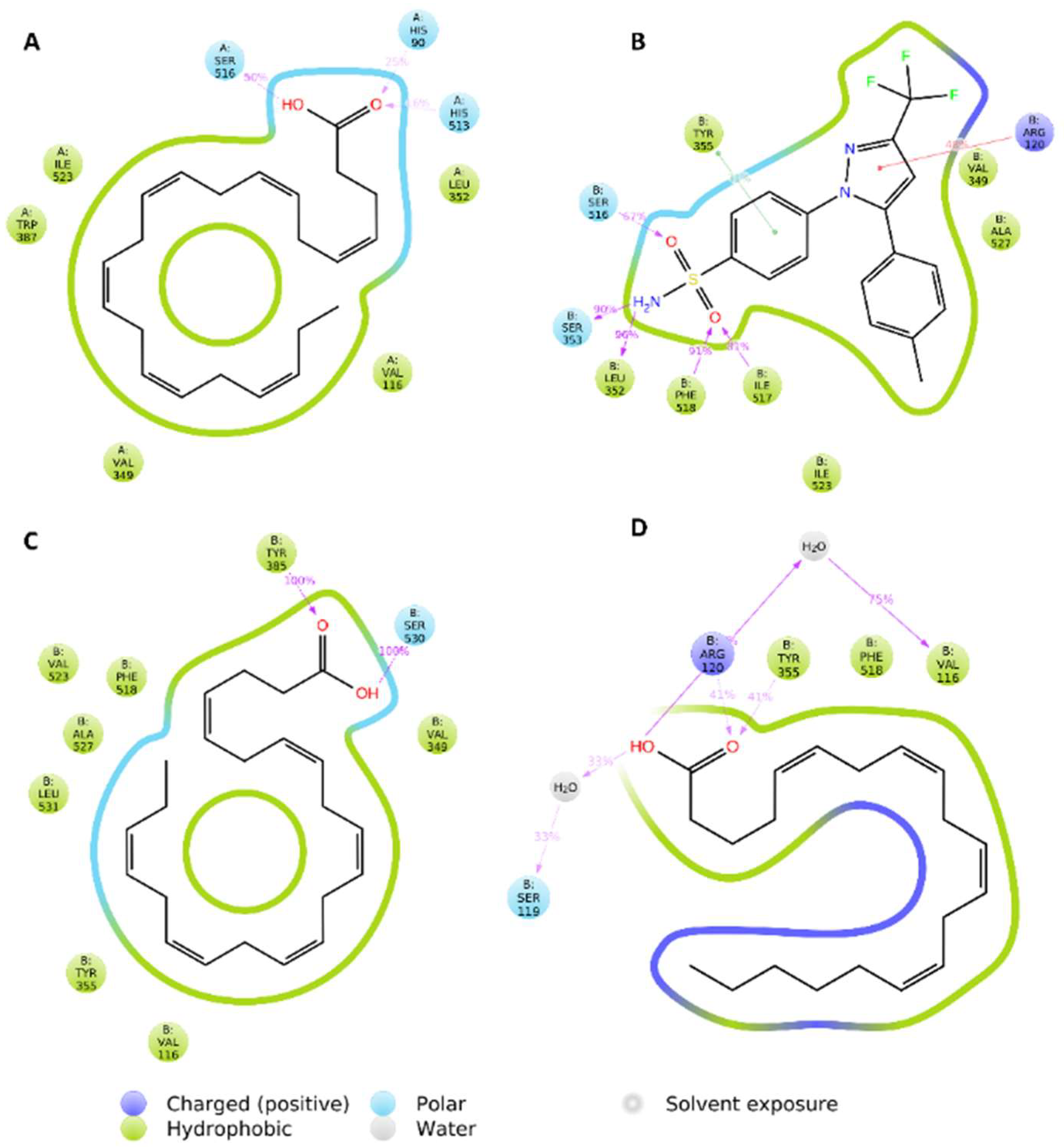
2.6. Molecular Modeling Study
3. Discussion
4. Materials and Methods
4.1. Fish Collection
4.2. Chemicals and Reagents
4.3. NMR Spectral Analyses
4.4. Sample Preparation and Extraction
4.5. Animal Preparation
4.6. MSU-Crystal Synthesis
4.7. Gouty-Arthritis-Animal Model
4.8. Biochemical Assays
4.9. Blood Sampling
4.10. Histopathological Examination
4.11. Histopathological Lesion Scoring
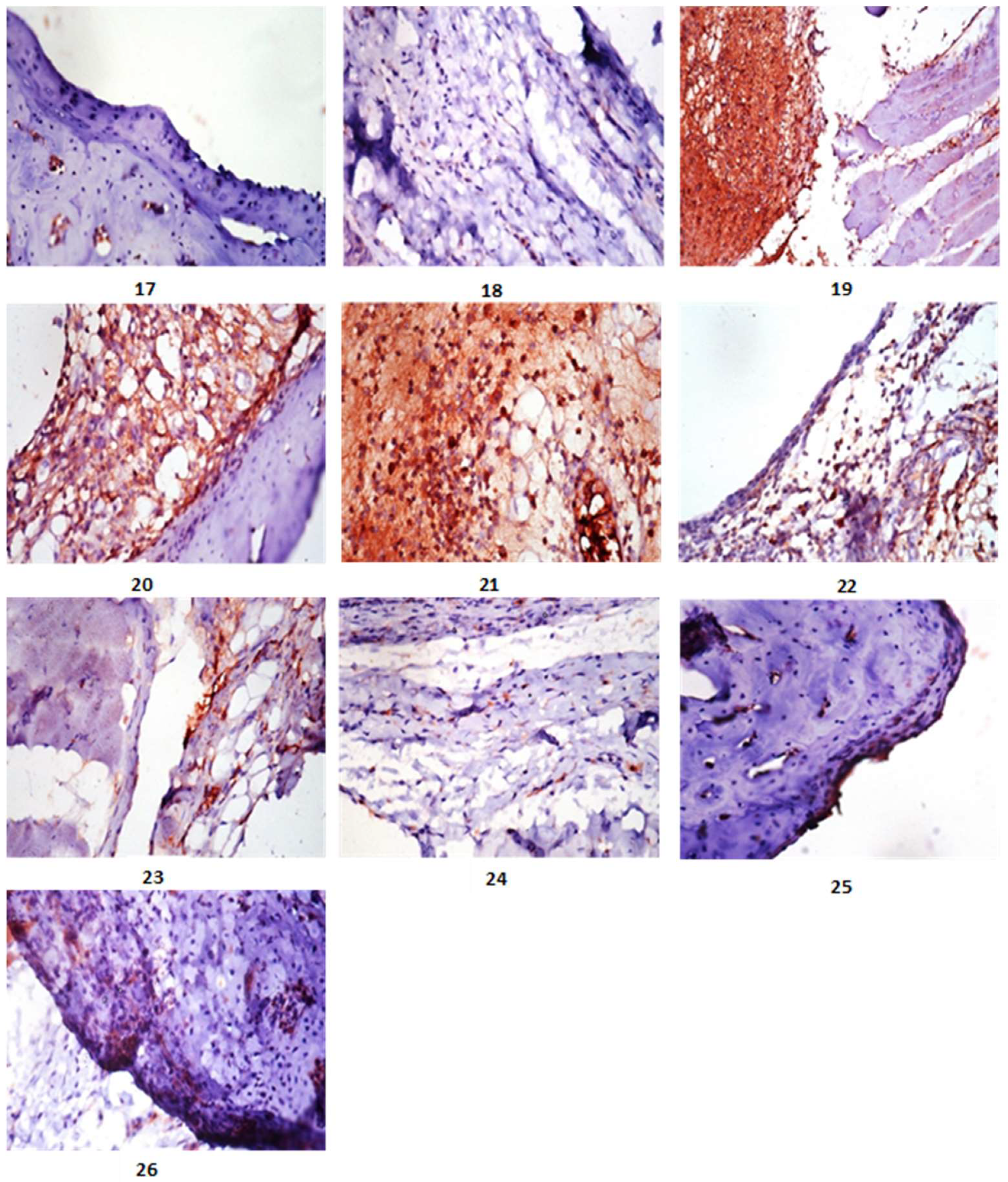
4.12. Immunohistochemistry
4.13. Evaluation of iNOS Immunostaining
4.14. Preparation of Fatty Acids Methyl Esters
4.15. GC-MS Analysis of Fatty Acids Methyl Esters
4.16. Isolation and Purification of Compounds
4.17. Amino Acid Analysis
4.17.1. Device Specification
4.17.2. Standard Preparation
4.17.3. Sample Preparation
4.17.4. Instrument Parameters
4.18. Molecular Modeling Study
4.18.1. Ligand Preparation
4.18.2. Receptor Preparation
4.18.3. Structural Docking
4.18.4. Molecular Dynamics Simulation
4.18.5. Absolute Binding Free Energy Calculations
4.19. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Li, W.-Y.; Yang, F.; Chen, J.-H.; Ren, G.-F. β-Caryophyllene Ameliorates MSU-Induced Gouty Arthritis and Inflammation Through Inhibiting NLRP3 and NF-κB Signal Pathway: In Silico and In Vivo. Front. Pharmacol. 2021, 12, 651305. [Google Scholar] [CrossRef] [PubMed]
- Kou, Y.-Y.; Li, Y.-F.; Xu, M.; Li, W.-Y.; Yang, M.; Li, R.-L. Effects of RuPeng15 Powder (RPP15) on Monosodium Urate Crystal-Induced Gouty Arthritis in Rats. Evid.-Based Complement. Altern. Med. 2015, 2015, 527019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Lin, Z.; Zhang, B.; Jiang, Z.; Guo, F.; Yang, T. Cichorium intybus L. Extract Suppresses Experimental Gout by Inhibiting the NF-κB and NLRP3 Signaling Pathways. Int. J. Mol. Sci. 2019, 20, 4921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Y.; Li, J.; Zhu, H.; Liu, J.; Dong, S.; Li, L.; Qu, L.; Chen, H.; Song, X.; Lan, X. Deletion mutation within the goat PPP3CA gene identified by GWAS significantly affects litter size. Reprod. Fertil. Dev. 2021, 33, 476–483. [Google Scholar] [CrossRef]
- Zhou, M.; Li, S.; Song, L.; Hu, Q.; Liu, W. 4-(2-(4-chlorophenyl)-1-((4-chlorophenyl)amino)ethyl)benzene-1, 3-diol is a potential agent for gout therapy as a dual inhibitor of XOD and NLRP3. Phytomedicine 2018, 42, 9–17. [Google Scholar] [CrossRef]
- Liu, W.-H.; Shi, L.-S.; Chung, M.-C.; Chang, T.-C.; Lee, S.-Y. Antcamphin M Inhibits TLR4-Mediated Inflammatory Responses by Upregulating the Nrf2/HO-1 Pathway and Suppressing the NLRP3 Inflammasome Pathway in Macrophages. Am. J. Chin. Med. 2019, 47, 1611–1626. [Google Scholar] [CrossRef]
- Qaseem, A.; McLean, R.M.; Starkey, M.; Forciea, M.A.; for the Clinical Guidelines Committee of the American College of Physicians. Diagnosis of Acute Gout: A Clinical Practice Guideline from the American College of Physicians. Ann. Intern. Med. 2016, 166, 52–57. [Google Scholar] [CrossRef] [Green Version]
- Alsenani, F.; Ashour, A.M.; Alzubaidi, M.A.; Azmy, A.F.; Hetta, M.H.; Abu-Baih, D.H.; Elrehany, M.A.; Zayed, A.; Sayed, A.M.; Abdelmohsen, U.R.; et al. Wound Healing Metabolites from Peters’ Elephant-Nose Fish Oil: An In Vivo Investigation Supported by In Vitro and In Silico Studies. Mar. Drugs 2021, 19, 605. [Google Scholar] [CrossRef]
- Cheung, P.C.K.; Mehta, B.M. Handbook of Food Chemistry; Springer: Cham, Switzerland, 2015; Volume 11. [Google Scholar]
- Fournier, V.; Destaillats, F.; Juanéda, P.; Dionisi, F.; Lambelet, P.; Sébédio, J.; Berdeaux, O. Thermal degradation of long-chain polyunsaturated fatty acids during deodorization of fish oil. Eur. J. Lipid Sci. Technol. 2006, 108, 33–42. [Google Scholar] [CrossRef]
- Teitelbaum, J.E.; Walker, W.A. Review: The role of omega 3 fatty acids in intestinal inflammation. J. Nutr. Biochem. 2001, 12, 21–32. [Google Scholar] [CrossRef]
- Vancassel, S. Oméga 3 et neurotransmission cérébrale. Oléagineux Corps Gras Lipides 2004, 11, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Al-Khudairy, L.; Hartley, L.; Clar, C.; Flowers, N.; Hooper, L.; Rees, K. Omega 6 fatty acids for the primary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2015, 11, CD011094. [Google Scholar] [CrossRef] [Green Version]
- Simopoulos, A.P. The Importance of the Omega-6/Omega-3 Fatty Acid Ratio in Cardiovascular Disease and Other Chronic Diseases. Exp. Biol. Med. 2008, 233, 674–688. [Google Scholar] [CrossRef] [PubMed]
- Skelton, P.; Cambray, J.; Lombard, A.; Benn, G. Patterns of distribution and conservation status of freshwater fishes in South Africa. S. Afr. J. Zoöl. 1995, 30, 71–81. [Google Scholar] [CrossRef]
- Lissmann, H.W. On the Function and Evolution of Electric Organs in Fish. J. Exp. Biol. 1958, 35, 156–191. [Google Scholar] [CrossRef]
- Bau, M.; Dulski, P.; Möller, P. Yttrium and holmium in South Pacific seawater: Vertical distribution and possible fractionation mechanisms. Chem. Erde 1995, 55, 955. [Google Scholar]
- Volknandt, W.; Zimmermann, H. Acetylcholine, ATP, and Proteoglycan Are Common to Synaptic Vesicles Isolated from the Electric Organs of Electric Eel and Electric Catfish as Well as from Rat Diaphragm. J. Neurochem. 1986, 47, 1449–1462. [Google Scholar] [CrossRef]
- Janetzko, A.; Zimmermann, H.; Volknandt, W. The electromotor system of the electric catfish (Malapterurus electricus): A fine-structural analysis. Cell Tissue Res. 1987, 247, 613–624. [Google Scholar] [CrossRef]
- Silva, J.R.; Burger, B.; Kühl, C.M.C.; Candreva, T.; Dos Anjos, M.B.P.; Rodrigues, H.G. Wound Healing and Omega-6 Fatty Acids: From Inflammation to Repair. Mediat. Inflamm. 2018, 2018, 2503950. [Google Scholar] [CrossRef] [Green Version]
- Chalamaiah, M.; Kumar, B.D.; Hemalatha, R.; Jyothirmayi, T. Fish protein hydrolysates: Proximate composition, amino acid composition, antioxidant activities and applications: A review. Food Chem. 2012, 135, 3020–3038. [Google Scholar] [CrossRef]
- Darlington, L.G.; Stone, T.W. Antioxidants and fatty acids in the amelioration of rheumatoid arthritis and related disorders. Br. J. Nutr. 2001, 85, 251–269. [Google Scholar] [CrossRef] [PubMed]
- Navarini, L.; Afeltra, A.; Afflitto, G.G.; Margiotta, D.P.E. Polyunsaturated fatty acids: Any role in rheumatoid arthritis? Lipids Health Dis. 2017, 16, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostoglou-Athanassiou, I.; Athanassiou, L.; Athanassiou, P. The Effect of Omega-3 Fatty Acids on Rheumatoid Arthritis. Mediterr. J. Rheumatol. 2020, 31, 190–194. [Google Scholar] [CrossRef]
- Iniguez, M.A.; Pablos, J.L.; Carreira, P.E.; Cabre, F.; Gomez-Reino, J.J. Detection of COX-1 and COX-2 isoforms in synovial fluid cells from inflammatory joint diseases. Br. J. Rheumatol. 1998, 37, 773–778. [Google Scholar] [CrossRef] [Green Version]
- Nakata, K.; Hanai, T.; Take, Y.; Osada, T.; Tsuchiya, T.; Shima, D.; Fujimoto, Y. Disease-modifying effects of COX-2 selective inhibitors and non-selective NSAIDs in osteoarthritis: A systematic review. Osteoarthr. Cartil. 2018, 26, 1263–1273. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.-H.; Chung, C.-H.; Lee, C.-H.; Hsieh, C.-H.; Hung, Y.-J.; Lin, F.-H.; Tsao, C.-H.; Hsieh, P.-S.; Chien, W.-C. Combination COX-2 inhibitor and metformin attenuate rate of joint replacement in osteoarthritis with diabetes: A nationwide, retrospective, matched-cohort study in Taiwan. PLoS ONE 2018, 13, e0191242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngo, S.T.; Tam, N.M.; Pham, M.Q.; Nguyen, T.H. Benchmark of Popular Free Energy Approaches Revealing the Inhibitors Binding to SARS-CoV-2 Mpro. J. Chem. Inf. Model. 2021, 61, 2302–2312. [Google Scholar] [CrossRef]
- Getting, S.J.; Christian, H.C.; Flower, R.J.; Perretti, M. Activation of melanocortin type 3 receptor as a molecular mechanism for adrenocorticotropic hormone efficacy in gouty arthritis. Arthritis Care Res. 2002, 46, 2765–2775. [Google Scholar] [CrossRef]
- Majumdar, K.N.; Banerjee, A.; Ratha, J.; Mandal, M.; Sarkar, R.N.; Das Saha, K. Leishmanial lipid suppresses tumor necrosis factor α, interleukin-1β, and nitric oxide production by adherent synovial fluid mononuclear cells in rheumatoid arthritis patients and induces apoptosis through the mitochondrial-mediated pathway. Arthritis Care Res. 2008, 58, 696–706. [Google Scholar] [CrossRef]
- Si, Y.; Zhang, Y.; Zhao, J.; Guo, S.; Zhai, L.; Yao, S.; Sang, H.; Yang, N.; Song, G.; Gu, J.; et al. Niacin Inhibits Vascular Inflammation via Downregulating Nuclear Transcription Factor-κB Signaling Pathway. Mediat. Inflamm. 2014, 2014, 263786. [Google Scholar] [CrossRef] [Green Version]
- Park, J.-E.; Yeom, Z.; Park, K.-T.; Han, E.H.; Yu, H.J.; Kang, H.S.; Lim, Y.-H. Hypouricemic Effect of Ethanol Extract of Aster glehni Leaves in Potassium Oxonate-Induced Hyperuricemic Rats. Clin. Nutr. Res. 2018, 7, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Hameed, B.J.; Ramadhan, U.H. Xanthine oxidase inhibitory, antihyperuricemic, anti-inflammatory, antinociceptive activity of α-lipoic acid in gouty arthritis model. Asian J. Pharm. Clin. Res. 2018, 11, 483–487. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.R.; Fröhlich, J.K.; Oliveira, S.M.; Cabreira, T.N.; Rossato, M.F.; Trevisan, G.; Froeder, A.L.; Bochi, G.V.; Moresco, R.N.; Athayde, M.L.; et al. The antinociceptive and anti-inflammatory effects of the crude extract of Jatropha isabellei in a rat gout model. J. Ethnopharmacol. 2013, 145, 205–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.; Xie, Y.; Sui, F.; Liu, C.; Du, X.; Liu, C.; Feng, X.; Jiang, D. Zisheng Shenqi decoction ameliorates monosodium urate crystal-induced gouty arthritis in rats through anti-inflammatory and anti-oxidative effects. Mol. Med. Rep. 2016, 14, 2589–2597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.-J.; Lee, C.-H.; Wang, C.-C.; Salter, D.M.; Lee, H.-S. Pycnogenol attenuates the inflammatory and nitrosative stress on joint inflammation induced by urate crystals. Free Radic. Biol. Med. 2011, 52, 765–774. [Google Scholar] [CrossRef]
- Kurz, B.; Steinhagen, J.; Schünke, M. Articular chondrocytes and synoviocytes in a co-culture system: Influence on reactive oxygen species-induced cytotoxicity and lipid peroxidation. Cell Tissue Res. 1999, 296, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, E. Inflammation, oxidative stress and lipids: The risk triad for atherosclerosis in gout. Rheumatology 2010, 49, 1229–1238. [Google Scholar] [CrossRef] [Green Version]
- So, A.; Thorens, B. Uric acid transport and disease. J. Clin. Investig. 2010, 120, 1791–1799. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-P.; Huang, S.-Y.; Lin, Y.-Y.; Wang, H.-M.; Jean, Y.-H.; Wu, S.-F.; Duh, C.-Y.; Wen, Z.-H. Soft Coral-Derived Lemnalol Alleviates Monosodium Urate-Induced Gouty Arthritis in Rats by Inhibiting Leukocyte Infiltration and iNOS, COX-2 and c-Fos Protein Expression. Mar. Drugs 2013, 11, 99–113. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Hsieh, M.-S.; Ho, H.-C.; Liu, Y.-H.; Chou, D.-T.; Tsai, S.-H. Stimulation of inducible nitric oxide synthase by monosodium urate crystals in macrophages and expression of iNOS in gouty arthritis. Nitric Oxide 2004, 11, 228–236. [Google Scholar] [CrossRef]
- McInnes, I.B.; Schett, G. The pathogenesis of rheumatoid arthritis. N. Engl. J. Med. 2011, 365, 2205–2219. [Google Scholar] [CrossRef] [PubMed]
- Nimse, S.B.; Pal, D. Free radicals, natural antioxidants, and their reaction mechanisms. RSC Adv. 2015, 5, 27986–28006. [Google Scholar] [CrossRef] [Green Version]
- Pal, R.; Chaudhary, M.J.; Tiwari, P.C.; Babu, S.; Pant, K. Protective role of theophylline and their interaction with nitric oxide (NO) in adjuvant-induced rheumatoid arthritis in rats. Int. Immunopharmacol. 2015, 29, 854–862. [Google Scholar] [CrossRef] [PubMed]
- Bahadori, B.; Uitz, E.; Thonhofer, R.; Trummer, M.; Pestemer-Lach, I.; McCarty, M.; Krejs, G.J. ω-3 Fatty Acids Infusions as Adjuvant Therapy in Rheumatoid Arthritis. J. Parenter. Enter. Nutr. 2010, 34, 151–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruggiero, C.; Lattanzio, F.; Lauretani, F.; Gasperini, B.; Andres-Lacueva, C.; Cherubini, A. Ω-3 Polyunsaturated Fatty Acids and Immune-Mediated Diseases: Inflammatory Bowel Disease and Rheumatoid Arthritis. Curr. Pharm. Des. 2009, 15, 4135–4148. [Google Scholar] [CrossRef]
- Kokkinos, P.P.; Shaye, R.; Alam, B.S.; Alam, S.Q. Dietary lipids, prostaglandin E2 levels, and tooth movement in alveolar bone of rats. Calcif. Tissue Res. 1993, 53, 333–337. [Google Scholar] [CrossRef]
- Serhan, C.N.; Arita, M.; Hong, S.; Gotlinger, K. Resolvins, docosatrienes, and neuroprotectins, novel omega-3-derived mediators, and their endogenous aspirin-triggered epimers. Lipids 2004, 39, 1125–1132. [Google Scholar] [CrossRef]
- Herrera, B.; Ohira, T.; Gao, L.; Omori, K.; Yang, R.; Zhu, M.; Muscara, M.; Serhan, C.N.; Van Dyke, T.E.; Gyurko, R. An endogenous regulator of inflammation, resolvin E1, modulates osteoclast differentiation and bone resorption. J. Cereb. Blood Flow Metab. 2008, 155, 1214–1223. [Google Scholar] [CrossRef]
- Poulsen, R.C.; Gotlinger, K.H.; Serhan, C.N.; Kruger, M.C. Identification of inflammatory and proresolving lipid mediators in bone marrow and their lipidomic profiles with ovariectomy and omega-3 intake. Am. J. Hematol. 2008, 83, 437–445. [Google Scholar] [CrossRef]
- James, M.J.; Gibson, R.A.; Cleland, L.G. Dietary polyunsaturated fatty acids and inflammatory mediator production. Am. J. Clin. Nutr. 2000, 71, 343s–348s. [Google Scholar] [CrossRef] [Green Version]
- Saxena, R.N.; Pendse, V.K.; Khanna, N.K. Anti-inflammatory and analgesic properties of four amino-acids. Indian J. Physiol. Pharmacol. 1984, 28, 299–305. [Google Scholar] [PubMed]
- Lee, J.H.; Park, E.; Jin, H.J.; Lee, Y.; Choi, S.J.; Lee, G.W.; Chang, P.-S.; Paik, H.-D. Anti-inflammatory and anti-genotoxic activity of branched chain amino acids (BCAA) in lipopolysaccharide (LPS) stimulated RAW 264.7 macrophages. Food Sci. Biotechnol. 2017, 26, 1371–1377. [Google Scholar] [CrossRef]
- Alhagamhmad, M.H.; Day, A.S.; Lemberg, D.; Leach, S.T. Exploring and Enhancing the Anti-Inflammatory Properties of Polymeric Formula. J. Parenter. Enter. Nutr. 2016, 41, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Rapin, J.R.; Wiernsperger, N. Possible links between intestinal permeablity and food processing: A potential therapeutic niche for glutamine. Clinics 2010, 65, 635–643. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, S.; Ichiyama, T.; Sonaka, I.; Ohsaki, A.; Hirano, R.; Haneda, Y.; Fukano, R.; Hara, M.; Furukawa, S. Amino acids exhibit anti-inflammatory effects in human monocytic leukemia cell line, THP-1 cells. Inflamm. Res. 2011, 60, 1013–1019. [Google Scholar] [CrossRef] [PubMed]
- Davies, N.W.; Guillemin, G.; Brew, B.J. Tryptophan, Neurodegeneration and HIV-Associated Neurocognitive Disorder. Int. J. Tryptophan Res. 2010, 3, IJTR.S4321. [Google Scholar] [CrossRef] [Green Version]
- Érces, D.; Varga, G.; Fazekas, B.; Kovács, T.; Tőkés, T.; Tiszlavicz, L.; Fülöp, F.; Vécsei, L.; Boros, M.; Kaszaki, J. N-methyl-d-aspartate receptor antagonist therapy suppresses colon motility and inflammatory activation six days after the onset of experimental colitis in rats. Eur. J. Pharmacol. 2012, 691, 225–234. [Google Scholar] [CrossRef]
- He, F.; Wu, C.; Li, P.; Li, N.; Zhang, D.; Zhu, Q.; Ren, W.; Peng, Y. Functions and Signaling Pathways of Amino Acids in Intestinal Inflammation. BioMed Res. Int. 2018, 2018, 9171905. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, X.; Hu, C.-A.A. Therapeutic Potential of Amino Acids in Inflammatory Bowel Disease. Nutrients 2017, 9, 920. [Google Scholar] [CrossRef] [Green Version]
- Olaosebikan, B.; Raji, A. Field Guide to Nigerian Freshwater Fishes; Federal College of Freshwater Fisheries Technology: New Bussa, Nigeria, 1998. [Google Scholar]
- AOAC. Official Methods of Analysis, 18th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2006. [Google Scholar]
- Jiang, Y.; You, X.-Y.; Fu, K.-L.; Yin, W.-L. Effects of Extract fromMangifera indicaLeaf on Monosodium Urate Crystal-Induced Gouty Arthritis in Rats. Evid.-Based Complement. Altern. Med. 2012, 2012, 967573. [Google Scholar] [CrossRef] [Green Version]
- Hidayat, R.; Reagan, M.; Hayati, L. Tempuyung Leaves (Sonchus arvensis) Ameliorates Monosodium Urate Crystal-Induced Gouty Arthritis in Rats through Anti-Inflammatory Effects. Open Access Maced. J. Med. Sci. 2020, 8, 220–224. [Google Scholar] [CrossRef]
- Estevão-Silva, C.F.; Ritter, A.M.V.; Ames, F.Q.; da Rocha, B.A.; Comar, J.F.; Wisniewski-Rebecca, E.S.; Caparroz-Assef, S.M.; Cuman, R.K.N.; Hernandes, L.; Bersani-Amado, C.A. Fish oil as effective supplement preventing inflammatory and histopathological alterations in adjuvant-induced arthritis in rats. Res. Soc. Dev. 2021, 10, e22610414046. [Google Scholar] [CrossRef]
- Mendes, J.; Guerois, R.; Serrano, L. Energy estimation in protein design. Curr. Opin. Struct. Biol. 2002, 12, 441–446. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Hsiao, G.; Lin, Y.-H.; Lin, C.-H.; Chou, D.-S.; Lin, W.-C.; Sheu, J.-R. The Protective Effects of PMC against Chronic Carbon Tetrachloride-Induced Hepatotoxicity In Vivo. Biol. Pharm. Bull. 2001, 24, 1271–1276. [Google Scholar] [CrossRef] [Green Version]
- Fridovich, I. Superoxide dismutase: An adaptation to a pragmatic gase. J. Biol. Chem. 1989, 264, 7762–7764. [Google Scholar] [CrossRef]
- Sinha, A. Catalase-An extra ordinary enzyme. Science 1972, 210, 71–82. [Google Scholar]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques; Elsevier Health Sciences: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Korany, R.M.; Ahmed, K.S.; Halawany, H.A.; Ahmed, K.A. Effect of long-term arsenic exposure on female Albino rats with special reference to the protective role of Spirulina platensis. Explor. Anim. Med. Res. 2019, 9, 125–136. [Google Scholar]
- Madkour, D.A.; Ahmed, M.M.; Orabi, S.H.; Sayed, S.M.; Korany, R.M.S.; Khalifa, H.K. Nigella sativa oil protects against emamectin benzoate-Induced neurotoxicity in rats. Environ. Toxicol. 2021, 36, 1521–1535. [Google Scholar] [CrossRef]
- Saleh, N.; Allam, T.; Korany, R.M.S.; Abdelfattah, A.M.; Omran, A.M.; Eldaim, M.A.A.; Hassan, A.M.; El-Borai, N.B. Protective and Therapeutic Efficacy of Hesperidin versus Cisplatin against Ehrlich Ascites Carcinoma-Induced Renal Damage in Mice. Pharmaceuticals 2022, 15, 294. [Google Scholar] [CrossRef] [PubMed]
- Al-Warhi, T.; Zahran, E.M.; Selim, S.; Al-Sanea, M.M.; Ghoneim, M.M.; Maher, S.A.; Mostafa, Y.A.; Alsenani, F.; Elrehany, M.A.; Almuhayawi, M.S.; et al. Antioxidant and Wound Healing Potential of Vitis vinifera Seeds Supported by Phytochemical Characterization and Docking Studies. Antioxidants 2022, 11, 881. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- Eastman, P.; Friedrichs, M.S.; Chodera, J.D.; Radmer, R.J.; Bruns, C.M.; Ku, J.P.; Beauchamp, K.A.; Lane, T.J.; Wang, L.-P.; Shukla, D.; et al. OpenMM 4: A Reusable, Extensible, Hardware Independent Library for High Performance Molecular Simulation. J. Chem. Theory Comput. 2013, 9, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Dallakyan, S.; Olson, A.J. Chemical Biology; Hempel, J.E., Williams, C.H., Hong, C.C., Eds.; Springer: New York, NY, USA, 2015; pp. 243–250. [Google Scholar] [CrossRef]
- Seeliger, D.; De Groot, B.L. Ligand docking and binding site analysis with PyMOL and Autodock/Vina. J. Comput. Mol. Des. 2010, 24, 417–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowers, K.J.; Chow, D.E.; Xu, H.; Dror, R.O.; Eastwood, M.P.; Gregersen, B.A.; Klepeis, J.L.; Kolossvary, I.; Moraes, M.A.; Sacerdoti, F.D.; et al. Scalable Algorithms for Molecular Dynamics Simulations on Commodity Clusters. In Proceedings of the SC’06: 2006 ACM/IEEE Conference on Supercomputing, Tampa, FL, USA, 11–17 November 2006; IEEE: New York, NY, USA, 2006; p. 43. [Google Scholar]
- Desmond Molecular Dynamics System, Release S.3; DE Shaw Research: New York, NY, USA, 2017.
- Maestro, Version 9.0; Schrodinger LLC: New York, NY, USA, 2009.
- Maestro-Desmond Interoperability Tools; Schrödinger: New York, NY, USA, 2017.
- Phillips, J.C.; Braun, R.; Wang, W.; Gumbart, J.; Tajkhorshid, E.; Villa, E.; Chipot, C.; Skeel, R.D.; Kalé, L.; Schulten, K. Scalable molecular dynamics with NAMD. J. Comput. Chem. 2005, 26, 1781–1802. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Oshima, H.; Zhang, H.; Kern, N.R.; Re, S.; Lee, J.; Roux, B.; Sugita, Y.; Jiang, W.; Im, W. CHARMM-GUI Free Energy Calculator for Absolute and Relative Ligand Solvation and Binding Free Energy Simulations. J. Chem. Theory Comput. 2020, 16, 7207–7218. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Uric Acid (mg/dL) | Ankle Swelling (mm) Post 24 h | Ankle Swelling (mm) Post 7 Says |
|---|---|---|---|
| Control Group | 3.00 ± 0.23 d | 0.50 ± 0.03 a | 0.50 ± 0.02 a |
| Msu Group % Change | 10.90 ± 0.30 a 263.33 | 3.55 ± 0.12 b 610.00 | 6.55 ± 0.56 b 1210 |
| Msu + Fe % Reduction | 5.00 ± 2.20 c 54.13 | 1.32 ± 0.44 c 62.82 | 1.90 ± 0.9 c 70.99 |
| Msu treated with Fe % Reduction | 4.00 ± 0.90 b 63.30 | 1.95 ± 0.66 d 45.10 | 1.89 ± 0.11 c 71.15 |
| Msu treated Standard Drug % Reduction | 3.50 ± 0.60 c 67.89 | 1.00 ± 0.43 e 71.83 | 1.80 ± 0.20 c 72.52 |
| TNF-α (pg/mL) | IL-1β (pg/mL) | NF-𝜅B (pg/mL) | |
|---|---|---|---|
| Control Group | 89.90 ± 9.00 a | 17.55 ± 1.40 a | 0.033 ± 0.003 a |
| Msu Group % Change | 700.00 ± 30.00 b 678.64 | 190.00 ± 19.00 b 982.62 | 0.10 ± 0.03 b 203.03 |
| Msu + FE % Reduction | 340.00 ± 16.90 c 51.43 | 89.00 ± 11.00 c 53.16 | 0.050 ± 0.02 c 50.00 |
| Msu treated Fe % Reduction | 200.00 ± 10.00 d 71.43 | 50.00 ± 3.00 d 73.68 | 0.040 ± 0.02 d 60.00 |
| Msu treated Standard Drug % Reduction | 190.00 ± 8.10 d 72.86 | 45.00 ± 2.90 d 76.32 | 0.037 ± 0.03 d 63.00 |
| Groups | GSH Reductase (mmol/min/g Tissue) | Catalase (µmol/min/g Tissue) | GSH (mg/g Tissue) | MDA (µmol/g Tissue) | SOD (µmol/g Tissue) |
|---|---|---|---|---|---|
| Control Group | 0.08 + 0.01 a | 0.09 + 0.02 a | 90.00 + 6.00 a | 4.10 + 0.18 a | 255.00 + 22.0 a |
| MSU Group % Change | 0.04 + 0.02 b 50.00 | 0.04 + 0.04 b 55.56 | 33.34 + 2.11 b 62.95 | 11.20 + 1.00 b 173.17 | 105.0 + 9.10 b 58.82 |
| MSU + FE % Reduction | 0.055 + 0.02 c 37.50 | 0.070 + 0.03 c 75.00 | 57.00 + 4.10 d 70.96 | 7.80 + 0.90 c 30.36 | 168.0 + 9.15 c 60.00 |
| MSU Treated FE % Reduction | 0.079 + 0.03 a 97.50 | 0.080 + 0.05 a 100.00 | 82.00 + 4.66 a 145.95 | 5.14 + 0.59 a 54.11 | 190.00 + 10.0 c 80.95 |
| MSU Treated Standard Drug % Reduction | 0.064+ ±0.04 d 60.00 | 0.080 + 0.02 a 100.00 | 80.00 + 3.50 a 139.95 | 5.48 + 0.66 a 51.10 | 188.10 + 10.00 c 79.14 |
| Lesions | G1 | G2 | G3 | G4 | G5 |
|---|---|---|---|---|---|
| Irregularity and deformities of articular surface | 0 | 3 | 0 | 1 | 1 |
| Necrosis of chondrocytes | 0 | 3 | 1 | 1 | 1 |
| Synovial membrane infiltrated with inflammatory cells | 0 | 3 | 1 | 1 | 1 |
| Widening of trabecular space | 0 | 3 | 0 | 1 | 1 |
| Groups | G1 | G2 | G3 | G4 | G5 |
|---|---|---|---|---|---|
| Affected Area | |||||
| Articular surface | 0 | 41.10 ± 1.50 b | 18.00 ± 0.88 a | 28.00 ± 3.00 c | 31.00 ± 1.80 c |
| % Reduction to arthritic rats | - | - | 56.20 | 31.87 | 24.57 |
| Synovial membrane | 0 | 60.15 ± 3.00 b | 19.00 ± 1.00 a | 31.00 ± 1.00 c | 34.00 ± 2.00 c |
| % Reduction to arthritic rats | - | - | 68.41 | 48.46 | 43.47 |
| No. | Identified Compound | C:D | Type | Area % | RT | RI |
|---|---|---|---|---|---|---|
| Palmitoleic acid | C16:1 (9) | MUFA | 1.70 | 27.81 | 919 | |
| 1 | Palmitic acid | C16:0 | SFA | 34.66 * | 28.68 | 931 |
| 2 | Vaccenic acid | C18:1 (11) | MUFA | 24.52 * | 32.64 | 931 |
| 9-Octadecenoic acid | C18:1 (9) | MUFA | 11.66 | 32.75 | 946 | |
| 3 | Stearic acid | C18:0 | SFA | 14.63 | 33.24 | 935 |
| Arachidonic acid | C20:4 (5,8,11,14) | PUFA | 3.41 | 35.83 | 913 | |
| 5,8,11,14,17-Eicosapentaenoic acid | C20:5 (5,8,11,14,17) | PUFA | 1.33 | 35.94 | 905 | |
| 4 | 4,7,10,13,16,19-Docosahexaenoic acid | C22:6 (4,7,10,13,16,19) | PUFA | 6.00 | 39.78 | 938 |
| SFA | 49.29% | |||||
| MUFA | 37.88% | |||||
| PUFA | 10.74% | |||||
| Total | 97.91% | |||||
| Identified Compound | RT | Amount (mg/100 mg) |
|---|---|---|
| Aspartic Acid | 7.699 | 0.142 |
| Threonine | 9.816 | 0.144 |
| Serine | 10.549 | 0.103 |
| Glutamic | 11.915 | 0.138 |
| Proline | 13.933 | 0.289 |
| Glycine | 17.824 | 0.813 |
| Alanine | 19.056 | 1.645 * |
| Cystine | 21.264 | 0.034 |
| Valine | 21.915 | 0.206 |
| Methionine | 23.803 | 0.019 |
| Isoleucine | 25.915 | 0.108 |
| Leucine | 27.171 | 0.247 |
| Tyrosine | 30.315 | 0.014 |
| Phenylalanine | 31.389 | 0.124 |
| Histidine | 35.128 | 0.142 |
| Lysine | 39.381 | 0.242 |
| Arginine | 43.056 | 0.173 |
| Total AA | 4.584 |
| Fatty Acid | Docking Score | ΔGbinding | ||
|---|---|---|---|---|
| COX-1 | COX-2 | COX-1 | COX-2 | |
| 1 | −7.41 | −7.56 | −3.23 | −3.57 |
| 2 | −7.51 | −7.34 | −5.38 | −6.34 |
| 3 | −7.39 | −7.44 | −3.86 | −3.92 |
| 4 | −8.46 | −8.75 | −8.23 | −12.45 |
| CEL * | −12.56 | −9.45 | −10.43 | −8.43 |
| Arachidonic acid ** | −11.54 | −10.53 | −9.42 | −9.54 |
| Buffer A | Buffer B | Column Regeneration Solution | Sample Dilution Buffer | |
|---|---|---|---|---|
| pH Value | 3.45 | 10.85 | 2.20 | |
| Normality | 0.12 | 0.20 | 0.50 | 0.12 |
| Tri-sodium citrate dihydrate | 11.8 g | 19.6 g | 11.8 g | |
| NaOH | 3.1 g | 20.0 g | ||
| Citric acid | 6.0 g | 6.0 g | ||
| Boric acid | 5.0 g | |||
| Methanol | 65 mL | |||
| Thiodiglycol | 14 mL | |||
| Hydrochloric acid 32% | 6.5 mL | 12 mL | ||
| EDTA | 0.2 g | |||
| Phenol | 0.5 g | 2.0 g | ||
| Final volume | 1 L | 1 L | 1 L | 1 L |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elmaidomy, A.H.; Mohamed, E.M.; Aly, H.F.; Younis, E.A.; Shams, S.G.E.; Altemani, F.H.; Alzubaidi, M.A.; Almaghrabi, M.; Harbi, A.A.; Alsenani, F.; et al. Anti-Inflammatory and Antioxidant Properties of Malapterurus electricus Skin Fish Methanolic Extract in Arthritic Rats: Therapeutic and Protective Effects. Mar. Drugs 2022, 20, 639. https://doi.org/10.3390/md20100639
Elmaidomy AH, Mohamed EM, Aly HF, Younis EA, Shams SGE, Altemani FH, Alzubaidi MA, Almaghrabi M, Harbi AA, Alsenani F, et al. Anti-Inflammatory and Antioxidant Properties of Malapterurus electricus Skin Fish Methanolic Extract in Arthritic Rats: Therapeutic and Protective Effects. Marine Drugs. 2022; 20(10):639. https://doi.org/10.3390/md20100639
Chicago/Turabian StyleElmaidomy, Abeer H., Esraa M. Mohamed, Hanan F. Aly, Eman A. Younis, Shams Gamal Eldin Shams, Faisal H. Altemani, Mubarak A. Alzubaidi, Mohammed Almaghrabi, Adnan Al Harbi, Faisal Alsenani, and et al. 2022. "Anti-Inflammatory and Antioxidant Properties of Malapterurus electricus Skin Fish Methanolic Extract in Arthritic Rats: Therapeutic and Protective Effects" Marine Drugs 20, no. 10: 639. https://doi.org/10.3390/md20100639