
FGFC1 Exhibits Anti-Cancer Activity via Inhibiting NF-κB Signaling Pathway in EGFR-Mutant NSCLC Cells

 ,
, 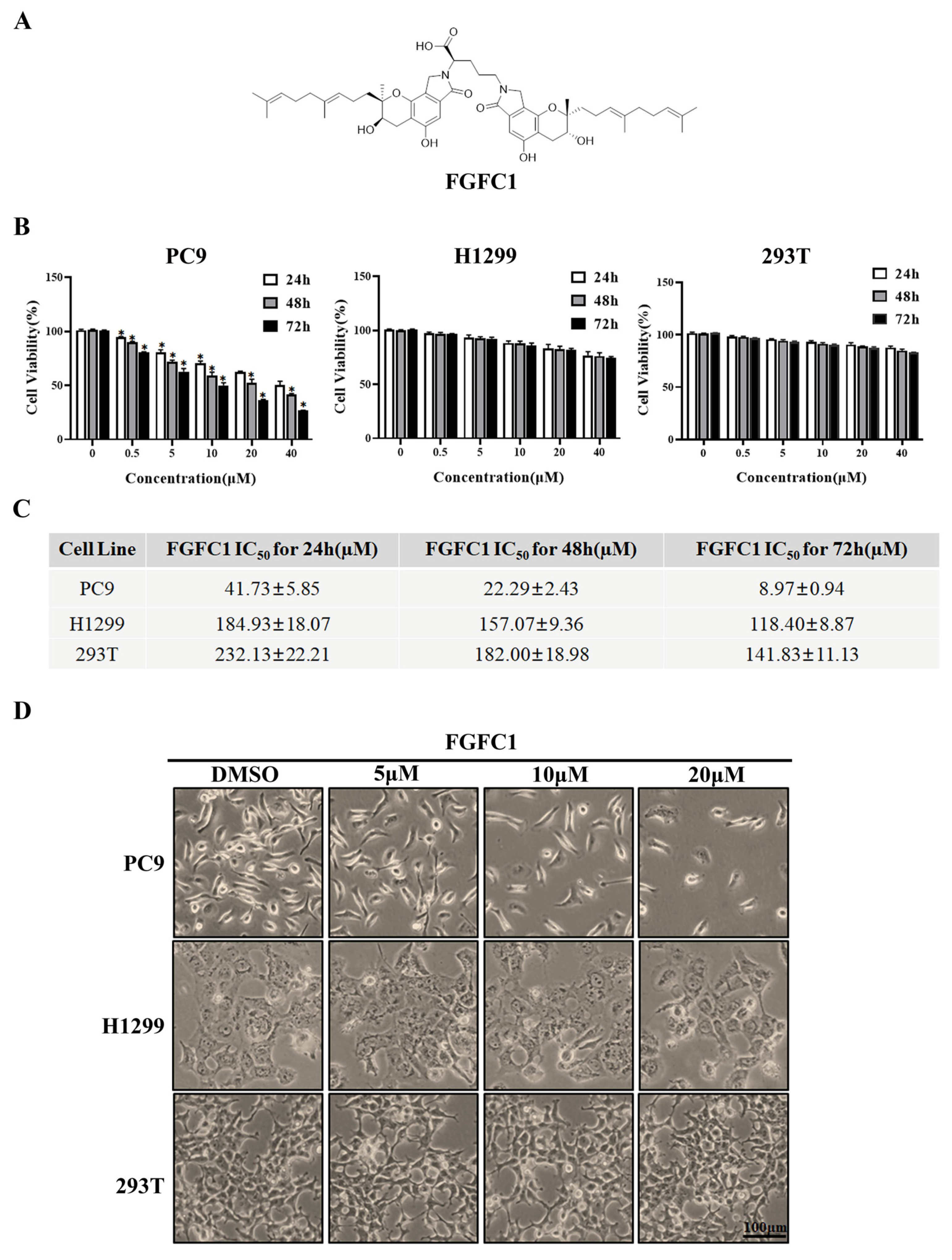{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. FGFC1 Significantly Inhibited PC9 Cells Viability In Vitro
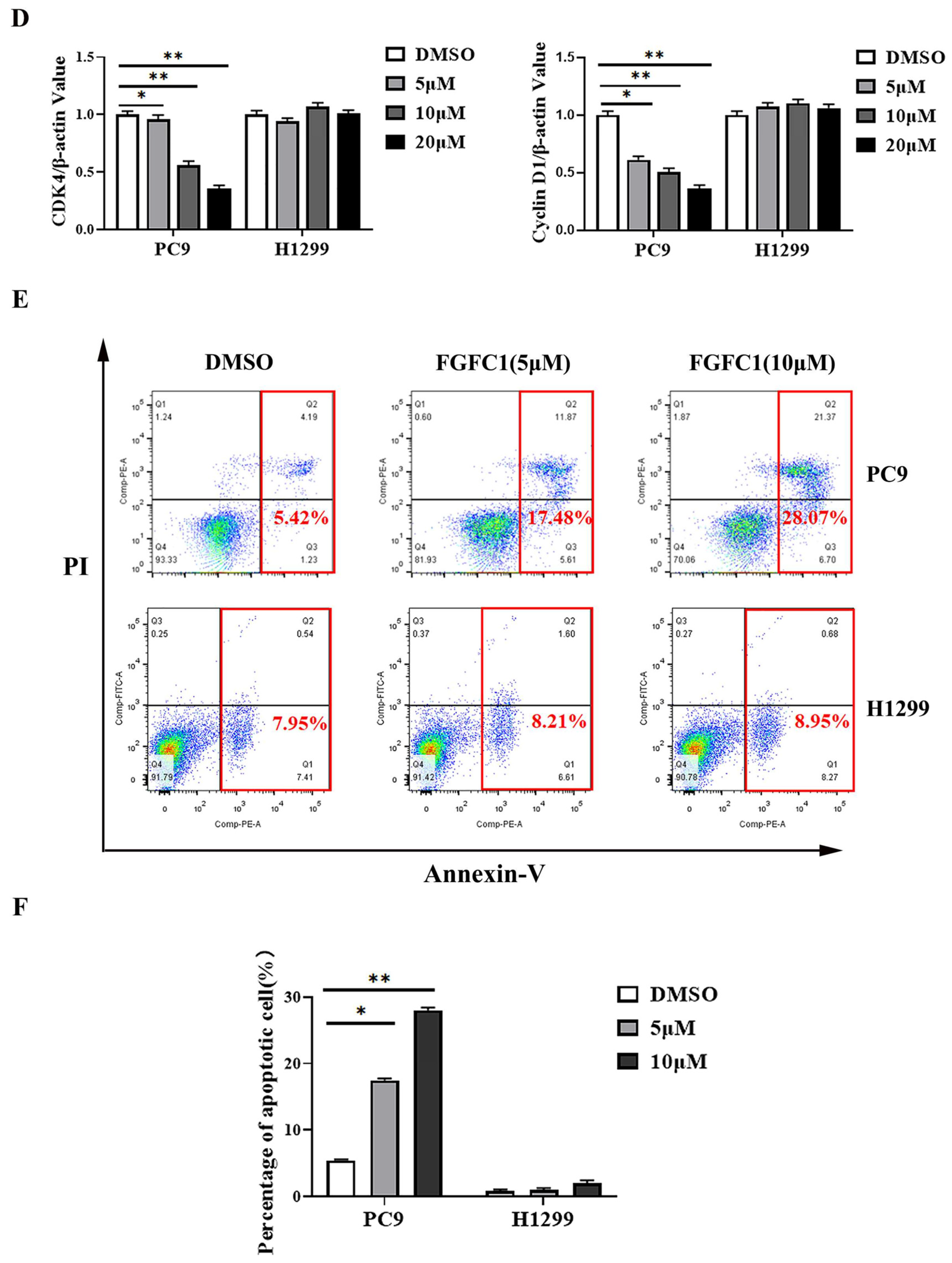
2.2. FGFC1 Induced G0/G1-Phase Arrest and Apoptosis in PC9 Cells
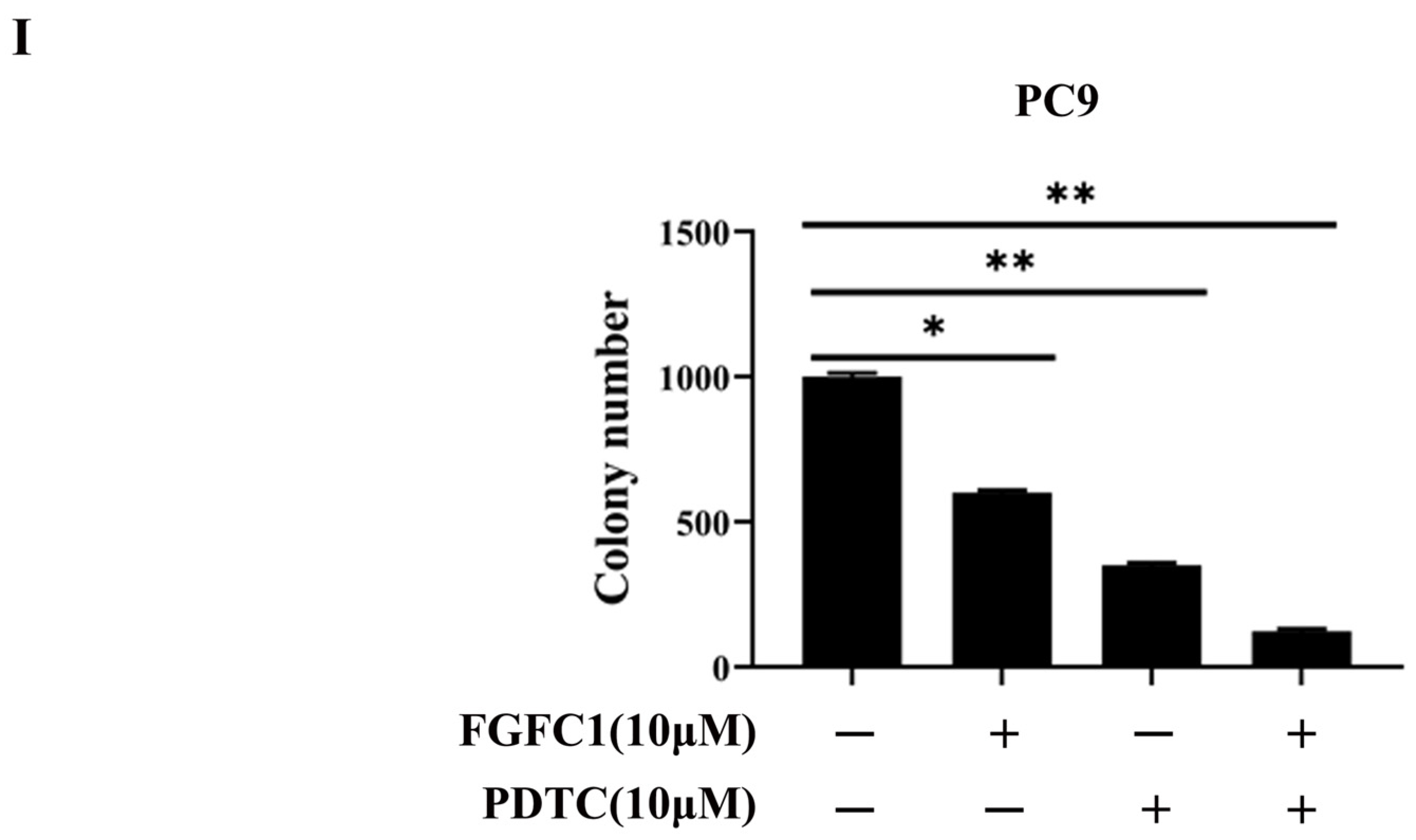
2.3. FGFC1 Affected NF-κB Signaling Activation in PC9 Cells
2.4. FGFC1 Inhibited NF-κB p-p65 Nuclear Translocation
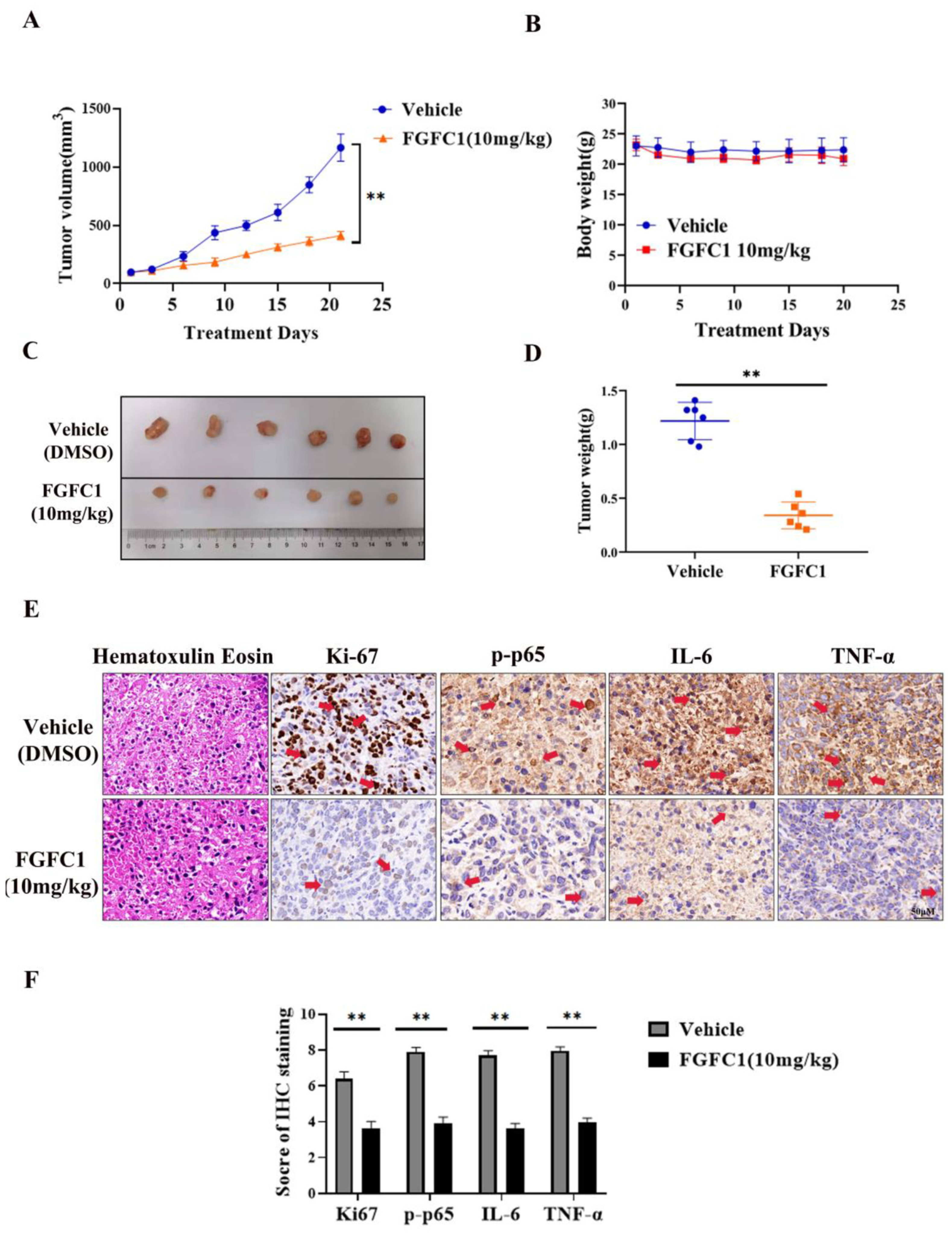
2.5. FGFC1 Inhibited Tumor Growth In Vivo
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Cell Lines and Cell Culture
4.3. Cell Proliferation Assay
4.4. Cell Morphology Observation
4.5. Colony Formation Assay
4.6. Cell Cycle Analysis
4.7. Apoptosis Assay
4.8. Transcriptome Analysis
4.9. Quantitative RT-PCR (qRT-PCR)
4.10. Western Blotting Analysis
4.11. Immunofluorescence Detection
4.12. In Vivo Antitumor Experiment
4.13. Immunohistochemistry
4.14. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Khanmohammadi, A.; Aghaie, A.; Vahedi, E.; Qazvini, A.; Ghanei, M.; Afkhami, A.; Hajian, A.; Bagheri, H. Electrochemical biosensors for the detection of lung cancer biomarkers: A review. Talanta 2020, 206, 120251. [Google Scholar] [CrossRef] [PubMed]
- Martinez, P.; Martinez-Marti, A.; Navarro, A.; Cedrés, S.; Felip, E. Molecular targeted therapy for early-stage non-small-cell lung cancer: Will it increase the cure rate? Lung Cancer 2014, 84, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Tsao, A.S.; Papadimitrakopoulou, V. The importance of molecular profiling in predicting response to epidermal growth factor receptor family inhibitors in non-small-cell lung cancer: Focus on clinical trial results. Clin. Lung Cancer 2013, 14, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Arriola, E.; García Gómez, R.; Diz, P.; Majem, M.; Martínez Aguillo, M. Clinical management and outcome of patients with advanced NSCLC carrying EGFR mutations in Spain. BMC Cancer 2018, 18, 106. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Tam, K.Y. Combination Strategies Using EGFR-TKi in NSCLC Therapy: Learning from the Gap between Pre-Clinical Results and Clinical Outcomes. Int. J. Biol. Sci. 2018, 14, 204–216. [Google Scholar] [CrossRef] [Green Version]
- Moya-Horno, I.; Viteri, S.; Karachaliou, N.; Rosell, R. Combination of immunotherapy with targeted therapies in advanced non-small cell lung cancer (NSCLC). Ther. Adv. Med. Oncol. 2018, 10, 1758834017745012. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Vong, C.T.; Chen, H.; Gao, Y.; Lyu, P.; Qiu, L.; Zhao, M.; Liu, Q.; Cheng, Z.; Zou, J.; et al. Naturally occurring anti-cancer compounds: Shining from Chinese herbal medicine. Chin. Med. 2019, 14, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roskoski, R., Jr. Cyclin-dependent protein serine/threonine kinase inhibitors as anticancer drugs. Pharmacol. Res. 2019, 139, 471–488. [Google Scholar] [CrossRef]
- Masuda, K.; Yasuda, N. The antibody against human CD25, ACT-1, recognizes canine T-lymphocytes in the G2/M and G0/G1 phases of the cell cycle during proliferation. J. Vet. Med. Sci. 2008, 70, 1285–1287. [Google Scholar] [CrossRef] [Green Version]
- Li, X.L.; Ma, R.H.; Ni, Z.J.; Thakur, K.; Cespedes-Acuña, C.L.; Wang, S.; Zhang, J.G.; Wei, Z.J. Dioscin inhibits human endometrial carcinoma proliferation via G0/G1 cell cycle arrest and mitochondrial-dependent signaling pathway. Food Chem. Toxicol. 2021, 148, 111941. [Google Scholar] [CrossRef]
- Singhal, S.; Vachani, A.; Antin-Ozerkis, D.; Kaiser, L.R.; Albelda, S.M. Prognostic implications of cell cycle, apoptosis, and angiogenesis biomarkers in non-small cell lung cancer: A review. Clin. Cancer Res. 2005, 11, 3974–3986. [Google Scholar] [CrossRef] [Green Version]
- De Falco, M.; De Luca, A. Cell cycle as a target of antineoplastic drugs. Curr. Pharm. Des. 2010, 16, 1417–1426. [Google Scholar] [CrossRef]
- Hao, X.L.; Han, F.; Zhang, N.; Chen, H.Q.; Jiang, X.; Yin, L.; Liu, W.B.; Wang, D.D.; Chen, J.P.; Cui, Z.H.; et al. TC2N, a novel oncogene, accelerates tumor progression by suppressing p53 signaling pathway in lung cancer. Cell Death Differ. 2019, 26, 1235–1250. [Google Scholar] [CrossRef]
- Guo, Y.; Tan, J.; Miao, Y.; Sun, Z.; Zhang, Q. Effects of Microvesicles on Cell Apoptosis under Hypoxia. Oxid. Med. Cell. Longev. 2019, 2019, 5972152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Gao, D.; Li, K.; Ye, S.; Liu, Q.; Peng, Y.; Lv, G.; Qiu, L.; Lin, J. Radiopharmacological evaluation of a caspase-3 responsive probe with optimized pharmacokinetics for PET imaging of tumor apoptosis. Org. Biomol. Chem. 2020, 18, 3512–3521. [Google Scholar] [CrossRef]
- Aoyagi Blue, Y.; Kusumi, J.; Satake, A. Copy number analyses of DNA repair genes reveal the role of poly(ADP-ribose) polymerase (PARP) in tree longevity. iScience 2021, 24, 102779. [Google Scholar] [CrossRef]
- Nalabothula, N.; Al-jumaily, T.; Eteleeb, A.M.; Flight, R.M.; Xiaorong, S.; Moseley, H.; Rouchka, E.C.; Fondufe-Mittendorf, Y.N. Genome-Wide Profiling of PARP1 Reveals an Interplay with Gene Regulatory Regions and DNA Methylation. PLoS ONE 2015, 10, e0135410. [Google Scholar] [CrossRef]
- Hoesel, B.; Schmid, J.A. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, Y.; Zhang, L.; Li, M.; Li, C.; Xue, C.; Huang, X.; Zhou, P. NF-κB downregulates Cbl-b through binding and suppressing Cbl-b promoter in T cell activation. J. Immunol. 2015, 194, 3778–3783. [Google Scholar] [CrossRef] [Green Version]
- Spiegelman, V.S.; Stavropoulos, P.; Latres, E.; Pagano, M.; Ronai, Z.; Slaga, T.J.; Fuchs, S.Y. Induction of beta-transducin repeat-containing protein by JNK signaling and its role in the activation of NF-κB. J. Biol. Chem. 2001, 276, 27152–27158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, T.; Liu, Z.; Shang, J.; Zhou, X.; Yu, S.; Tian, H.; Bao, Y. Polygonatum sibiricum polysaccharides play anti-cancer effect through TLR4-MAPK/NF-κB signaling pathways. Int. J. Biol. Macromol. 2018, 111, 813–821. [Google Scholar] [CrossRef]
- Wung, B.S.; Ni, C.W.; Wang, D.L. ICAM-1 induction by TNFalpha and IL-6 is mediated by distinct pathways via Rac in endothelial cells. J. Biomed. Sci. 2005, 12, 91–101. [Google Scholar] [CrossRef]
- Lepetsos, P.; Papavassiliou, K.A.; Papavassiliou, A.G. Redox and NF-κB signaling in osteoarthritis. Free Radic. Biol. Med. 2019, 132, 90–100. [Google Scholar] [CrossRef]
- Yang, Z.H.; Dang, Y.Q.; Ji, G. Role of epigenetics in transformation of inflammation into colorectal cancer. World J. Gastroenterol. 2019, 25, 2863–2877. [Google Scholar] [CrossRef]
- Pei, Z.; Li, H.; Guo, Y.; Jin, Y.; Lin, D. Sodium selenite inhibits the expression of VEGF, TGFbeta(1) and IL-6 induced by LPS in human PC3 cells via TLR4-NF-(K)B signaling blockage. Int. Immunopharmacol. 2010, 10, 50–56. [Google Scholar] [CrossRef]
- Florean, C.; Dicato, M.; Diederich, M. Immune-modulating and anti-inflammatory marine compounds against cancer. Semin. Cancer Biol. 2020, in press. [CrossRef]
- Wang, Y.; Chen, H.; Sheng, R.; Fu, Z.; Fan, J.; Wu, W.; Tu, Q.; Guo, R. Synthesis and Bioactivities of Marine Pyran-Isoindolone Derivatives as Potential Antithrombotic Agents. Mar. Drugs. 2021, 19, 218. [Google Scholar] [CrossRef]
- Shibata, K.; Hashimoto, T.; Nobe, K.; Hasumi, K.; Honda, K. A novel finding of a low-molecular-weight compound, SMTP-7, having thrombolytic and anti-inflammatory effects in cerebral infarction of mice. Naunyn-Schmied. Arch. Pharmacol. 2010, 382, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.; Duan, D.; Hong, S.; Zhou, Y.; Wang, F.; Wang, S.; Wu, W.; Bao, B. A Marine Fibrinolytic Compound FGFC1 Stimulating Enzymatic Kinetic Parameters of a Reciprocal Activation System Based on a Single Chain Urokinase-Type Plasminogen Activator and Plasminogen. Process Biochem. 2017, 68, 190–196. [Google Scholar] [CrossRef]
- Gao, C.; Shen, Q.; Tang, P.; Cao, Y.; Wu, W.J.M. In Vitro Study of the Fibrinolytic Activity via Single Chain Urokinase-Type Plasminogen Activator and Molecular Docking of FGFC1. Molecules 2021, 26, 1816. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Kimura, Y.; Ohata, H.; Hashimoto, T.; Shibata, K.; Hasumi, K.; Honda, K. Distinct effects of tissue-type plasminogen activator and SMTP-7 on cerebrovascular inflammation following thrombolytic reperfusion. Stroke 2011, 42, 1097–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, S.K.; Feng, J.W.; Chang, J. FGFC1 inhibits the proliferation and migration of non-small cell lung cancer cells via the PI3K/Akt/mTOR pathway. Chin. J. Biochem. Mol. Biol. 2021, 37, 9. [Google Scholar]
- Huang, M.H.; Lee, J.H.; Chang, Y.J.; Tsai, H.H.; Lin, Y.L.; Lin, A.M.; Yang, J.C. MEK inhibitors reverse resistance in epidermal growth factor receptor mutation lung cancer cells with acquired resistance to gefitinib. Mol. Oncol. 2013, 7, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Huang, H.P.; Wang, Y.; Jin, J.; Long, W.G.; Chen, K.; Zhao, X.H.; Chen, C.G.; Li, J. Curcumin overcome primary gefitinib resistance in non-small-cell lung cancer cells through inducing autophagy-related cell death. J. Exp. Clin. Cancer Res. 2019, 38, 254. [Google Scholar] [CrossRef] [Green Version]
- Piao, S.; Du, W.; Wei, Y.; Yang, Y.; Feng, X.; Bai, L. Protectin DX attenuates IL-1β-induced inflammation via the AMPK/NF-κB pathway in chondrocytes and ameliorates osteoarthritis progression in a rat model. Int. Immunopharmacol. 2020, 78, 106043. [Google Scholar] [CrossRef]
- Khan, H.; Ullah, H.; Castilho, P.; Gomila, A.S.; D’Onofrio, G.; Filosa, R.; Wang, F.; Nabavi, S.M.; Daglia, M.; Silva, A.S.; et al. Targeting NF-κB signaling pathway in cancer by dietary polyphenols. Crit. Rev. Food Sci. Nutr. 2020, 60, 2790–2800. [Google Scholar] [CrossRef]
- Tang, X.; Liu, D.; Shishodia, S.; Ozburn, N.; Behrens, C.; Lee, J.J.; Hong, W.K.; Aggarwal, B.B.; Wistuba, I.I. Nuclear factor-kappaB (NF-kappaB) is frequently expressed in lung cancer and preneoplastic lesions. Cancer 2006, 107, 2637–2646. [Google Scholar] [CrossRef]
- Xian, H.; Feng, W.; Zhang, J. Schizandrin A enhances the efficacy of gefitinib by suppressing IKKβ/NF-κB signaling in non-small cell lung cancer. Eur. J. Pharmacol. 2019, 855, 10–19. [Google Scholar] [CrossRef]
- Urruticoechea, A.; Smith, I.E.; Dowsett, M. Proliferation marker Ki-67 in early breast cancer. J. Clin. Oncol. 2005, 23, 7212–7220. [Google Scholar] [CrossRef]
- Gao, L.; Wang, Y.; Xu, Z.; Li, X.; Wu, J.; Liu, S.; Chu, P.; Sun, Z.; Sun, B.; Lin, Y.; et al. SZC017, a novel oleanolic acid derivative, induces apoptosis and autophagy in human breast cancer cells. Apoptosis 2015, 20, 1636–1650. [Google Scholar] [CrossRef] [PubMed]
- Chu, P.; Li, H.; Luo, R.; Ahsan, A.; Qaed, E. Oleanolic acid derivative SZC014 inhibit cell proliferation and induce apoptosis of human breast cancer cells in a ROS-dependent way. Neoplasma 2017, 64, 681–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knockaert, M.; Greengard, P.; Meijer, L. Pharmacological inhibitors of cyclin-dependent kinases. Trends Pharmacol. Sci. 2002, 23, 417–425. [Google Scholar] [CrossRef]
- Sun, F.; Li, N.; Tong, X.; Zeng, J.; He, S.; Gai, T.; Bai, Y.; Liu, L.; Lu, K.; Shen, J.; et al. Ara-c induces cell cycle G1/S arrest by inducing upregulation of the INK4 family gene or directly inhibiting the formation of the cell cycle-dependent complex CDK4/cyclin D1. Cell Cycle 2019, 18, 2293–2306. [Google Scholar] [CrossRef] [PubMed]
- Foster, D.A.; Yellen, P.; Xu, L.; Saqcena, M. Regulation of G1 Cell Cycle Progression: Distinguishing the Restriction Point from a Nutrient-Sensing Cell Growth Checkpoint(s). Genes Cancer 2010, 1, 1124–1131. [Google Scholar] [CrossRef]
- Wang, F.; Liu, Y.; Bi, Z. Pioglitazone inhibits growth of human retinoblastoma cells via regulation of NF-κB inflammation signals. J. Recept. Signal Transduct. Res. 2017, 37, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Fan, T.; Jiang, Y.; Zhang, X.; Wang, X. A small interfering RNA targeting NF-κB p65 alone or combined with 5-FU inhibits growth of esophageal squamous cell carcinoma in nude mice. Pathol. Res. Pract. 2012, 208, 32–38. [Google Scholar] [CrossRef]
- Lun, M.; Zhang, P.L.; Siegelmann-Danieli, N.; Blasick, T.M.; Brown, R.E. Intracellular inhibitory effects of Velcade correlate with morphoproteomic expression of phosphorylated-nuclear factor-κB and p53 in breast cancer cell lines. Ann. Clin. Lab. Sci. 2005, 35, 15–24. [Google Scholar]
- Chou, T.C. New mechanisms of antiplatelet activity of nifedipine, an L-type calcium channel blocker. BioMedicine 2014, 4, 24. [Google Scholar] [CrossRef]
- Huang, Y.; Ohta, Y.; Shang, J.; Morihara, R.; Nakano, Y. Antineuroinflammatory Effect of SMTP-7 in Ischemic Mice. J. Stroke Cerebrovasc. Dis. 2018, 27, 3084–3094. [Google Scholar] [CrossRef]
- Zerbini, L.F.; Wang, Y.; Czibere, A.; Correa, R.G.; Cho, J.Y.; Ijiri, K.; Wei, W.; Joseph, M.; Gu, X.; Grall, F.; et al. NF-kappa B-mediated repression of growth arrest- and DNA-damage-inducible proteins 45alpha and gamma is essential for cancer cell survival. Proc. Natl. Acad. Sci. USA 2004, 101, 13618–13623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paez, J.G.; Jänne, P.A.; Lee, J.C.; Tracy, S.; Greulich, H. EGFR mutations in lung cancer: Correlation with clinical response to gefitinib therapy. Science 2004, 304, 1497–1500. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Zhu, Y.; Zhou, Z.; Gumin, J.; Bengtsson, L.; Wu, W.; Songyang, Z.; Lang, F.F.; Lin, X. TMEM43/LUMA is a key signaling component mediating EGFR-induced NF-κB activation and tumor progression. Oncogene 2017, 36, 2813–2823. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, J.; Chen, Z.; Wang, L.; Wu, X.; Ou, M. Epidermal growth factor (EGF) triggers the malignancy of hemangioma cells via activation of NF-κB signals. Biomed. Pharmacother. 2016, 82, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, T.; Hu, M.; Zhang, Y.; Chen, H.; Xu, L. Metformin Overcomes Acquired Resistance to EGFR TKIs in EGFR-Mutant Lung Cancer via AMPK/ERK/NF-κB Signaling Pathway. Front. Oncol. 2020, 10, 1605. [Google Scholar] [CrossRef]
- Fukuoka, M.; Yoshioka, K.; Hohjoh, H. NF-κB activation is an early event of changes in gene regulation for acquiring drug resistance in human adenocarcinoma PC-9 cells. PLoS ONE 2018, 13, e0201796. [Google Scholar] [CrossRef]
- Bivona, T.G.; Hieronymus, H.; Parker, J.; Chang, K.; Taron, M.; Rosell, R.; Moonsamy, P.; Dahlman, K.; Miller, V.A.; Costa, C.; et al. FAS and NF-κB signalling modulate dependence of lung cancers on mutant EGFR. Nature 2011, 471, 523–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galvani, E.; Sun, J.; Leon, L.G.; Sciarrillo, R.; Narayan, R.S.; Sjin, R.T.; Lee, K.; Ohashi, K.; Heideman, D.A.; Alfieri, R.R.; et al. NF-κB drives acquired resistance to a novel mutant-selective EGFR inhibitor. Oncotarget 2015, 6, 42717–42732. [Google Scholar] [CrossRef] [Green Version]
- Maeda, M.; Murakami, Y.; Watari, K.; Kuwano, M.; Izumi, H.; Ono, M. CpG hypermethylation contributes to decreased expression of PTEN during acquired resistance to gefitinib in human lung cancer cell lines. Lung Cancer 2015, 87, 265–271. [Google Scholar] [CrossRef]
- Remmele, W.; Stegner, H.E. Recommendation for uniform definition of an immunoreactive score (IRS) for immunohistochemical estrogen receptor detection (ER-ICA) in breast cancer tissue. Pathologe 1987, 8, 138–140. [Google Scholar]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, J.; Li, S.; Zhang, B.; Duan, N.; Zhou, R.; Yan, S.; Elango, J.; Liu, N.; Wu, W. FGFC1 Exhibits Anti-Cancer Activity via Inhibiting NF-κB Signaling Pathway in EGFR-Mutant NSCLC Cells. Mar. Drugs 2022, 20, 76. https://doi.org/10.3390/md20010076
Feng J, Li S, Zhang B, Duan N, Zhou R, Yan S, Elango J, Liu N, Wu W. FGFC1 Exhibits Anti-Cancer Activity via Inhibiting NF-κB Signaling Pathway in EGFR-Mutant NSCLC Cells. Marine Drugs. 2022; 20(1):76. https://doi.org/10.3390/md20010076
Chicago/Turabian StyleFeng, Jingwen, Songlin Li, Bing Zhang, Namin Duan, Rui Zhou, Shike Yan, Jeevithan Elango, Ning Liu, and Wenhui Wu. 2022. "FGFC1 Exhibits Anti-Cancer Activity via Inhibiting NF-κB Signaling Pathway in EGFR-Mutant NSCLC Cells" Marine Drugs 20, no. 1: 76. https://doi.org/10.3390/md20010076