
Optimized Degradation and Inhibition of α-glucosidase Activity by Gracilaria lemaneiformis Polysaccharide and Its Production In Vitro

 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Optimized Degradation of Gracilaria Lemaneiformis Polysaccharide
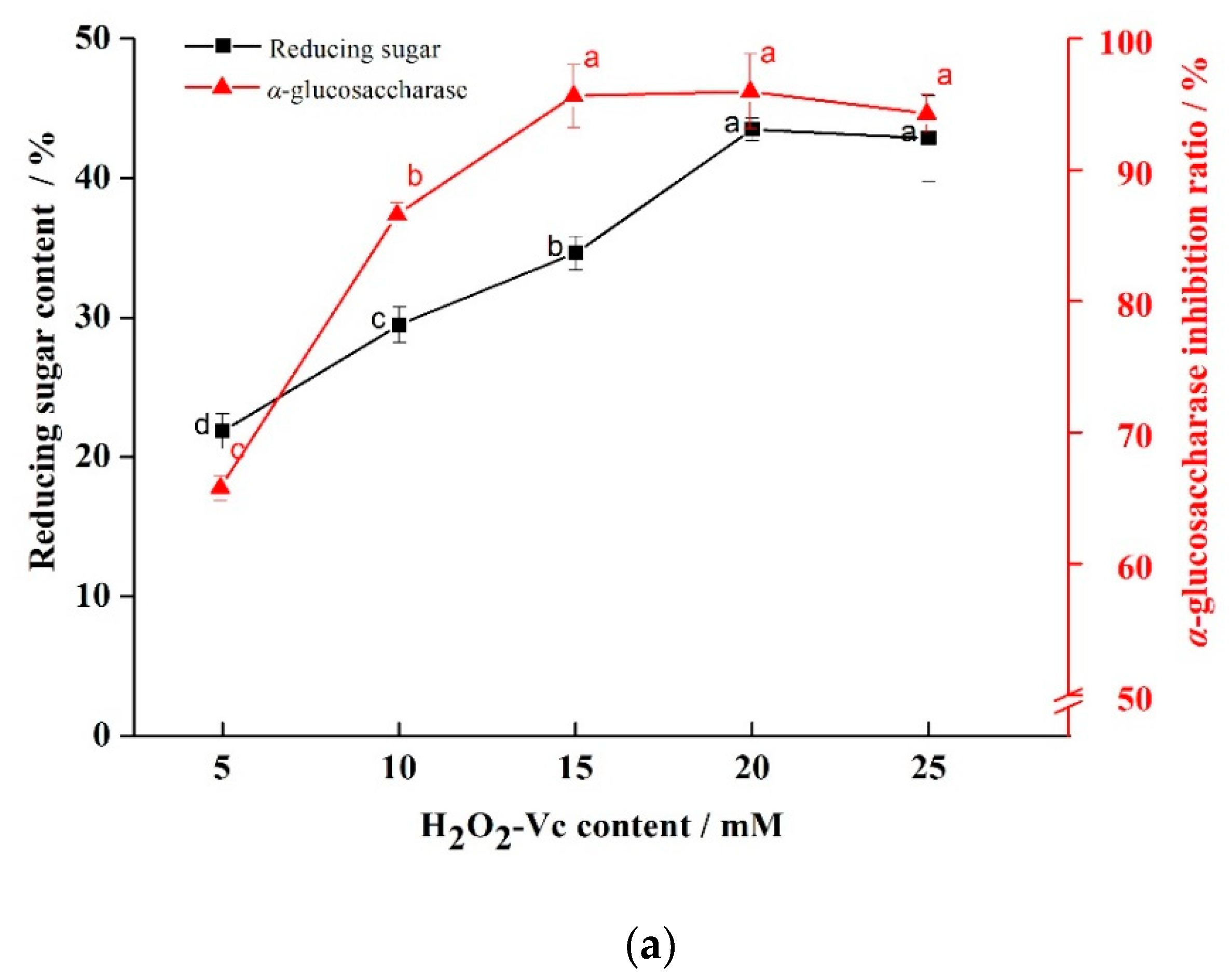
2.1.1. Results of Single Factor Experiment
2.1.2. Results of Response Surface Experiment
Establishment of Regression Model
Regression Model Analysis
Analysis of Model Interaction Items
Response Surface Optimization and Validation
2.2. The Content of Total Sugar, Reducing Sugar and Protein
2.3. Monosaccharide Composition
2.4. Molecular Weight
2.5. UV-Visible Spectroscopy
2.6. I2-KI Test
2.7. Congo Red Test
2.8. Scanning Electron Microscope Analysis
2.9. The Inhibition Effect on α-glucosidase
3. Discussion
4. Materials and Methods
4.1. Materials and Chemicals
4.2. Preparation of GLP
4.3. Degradation of GLP with H2O2-Vc
4.3.1. Single-Factor Experiment
4.3.2. Response Surface Analysis
4.3.3. Preparation of Degradation Products from GLP
4.4. Analysis of Chemical Characterizatics
4.4.1. Determination of Total Sugar, Reducing Sugar and Protein Content
4.4.2. Determination of Monosaccharide Composition
4.4.3. Determination of Molecular Weight
4.4.4. UV-Visible Spectroscopy
4.4.5. I2-KI Test
4.4.6. Congo Red Test
4.4.7. Scanning Electron Microscope Analysis (SEM)
4.5. The Inhibition Effect on α-Glucosidase
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yu, Y.-Y.; Chen, W.-D.; Liu, Y.-J.; Niu, J.; Chen, M.; Tian, L.-X. Effect of different dietary levels of Gracilaria lemaneiformis dry power on growth performance, hematological parameters and intestinal structure of juvenile Pacific white shrimp (Litopenaeus vannamei). Aquaculture 2016, 450, 356–362. [Google Scholar] [CrossRef]
- Long, X.; Hu, X.; Liu, S.; Pan, C.; Chen, S.; Li, L.; Qi, B.; Yang, X. Insights on preparation, structure and activities of Gracilaria lemaneiformis polysaccharide. Food Chem. X 2021, 12, 100153. [Google Scholar] [CrossRef]
- Fisheries Administration of Ministry of Agriculture and Rural Affairs. China Fishery Statistical Yearbook; China Agriculture Press: Beijing, China, 2020. [Google Scholar]
- Li, X.; Huang, S.; Chen, X.; Xu, Q.; Ma, Y.; You, L.; Kulikouskaya, V.; Xiao, J.; Piao, J. Structural characteristic of a sulfated polysaccharide from Gracilaria Lemaneiformis and its lipid metabolism regulation effect. Food Funct. 2020, 11, 10876–10885. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Duan, M.; Liu, Y.; Luo, T.; Ma, N.; Song, S.; Ai, C. The beneficial effects of Gracilaria lemaneiformis polysaccharides on obesity and the gut microbiota in high fat diet-fed mice. J. Funct. Foods 2018, 46, 48–56. [Google Scholar] [CrossRef]
- Fan, Y.; Wang, W.; Song, W.; Chen, H.; Teng, A.; Liu, A. Partial characterization and anti-tumor activity of an acidic polysaccharide from Gracilaria lemaneiformis. Carbohydr. Polym. 2012, 88, 1313–1318. [Google Scholar] [CrossRef]
- Chen, M.-Z.; Xie, H.-G.; Yang, L.-W.; Liao, Z.-H.; Yu, J. In vitro anti-influenza virus activities of sulfated polysaccharide fractions from Gracilaria lemaneiformis. Virol. Sin. 2010, 25, 341–351. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Z.; Zhou, H.; Sun, X.; Chen, X.; Xu, N. The anti-aging effects of Gracilaria lemaneiformis polysaccharide in Caenorhabditis elegans. Int. J. Biol. Macromol. 2019, 140, 600–604. [Google Scholar] [CrossRef]
- Kang, Y.; Wang, Z.-J.; Xie, D.; Sun, X.; Yang, W.; Zhao, X.; Xu, N. Characterization and Potential Antitumor Activity of Polysaccharide from Gracilariopsis lemaneiformis. Mar. Drugs 2017, 15, 100. [Google Scholar] [CrossRef]
- Chen, S.; Liu, H.; Yang, X.; Li, L.; Qi, B.; Hu, X.; Ma, H.; Li, C.; Pan, C. Degradation of sulphated polysaccharides from Grateloupia livida and antioxidant activity of the degraded components. Int. J. Biol. Macromol. 2020, 156, 660–668. [Google Scholar] [CrossRef]
- Gong, Y.; Ma, Y.; Cheung, P.C.-K.; You, L.; Liao, L.; Pedisić, S.; Kulikouskaya, V. Structural characteristics and anti-inflammatory activity of UV/H2O2-treated algal sulfated polysaccharide from Gracilaria lemaneiformis. Food Chem. Toxicol. 2021, 152, 112157. [Google Scholar] [CrossRef]
- Zhang, X.; Aweya, J.J.; Huang, Z.-X.; Kang, Z.-Y.; Bai, Z.-H.; Li, K.-H.; He, X.-T.; Liu, Y.; Chen, X.-Q.; Cheong, K.-L. In vitro fermentation of Gracilaria lemaneiformis sulfated polysaccharides and its agaro-oligosaccharides by human fecal inocula and its impact on microbiota. Carbohydr. Polym. 2020, 234, 115894. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.-Q.; Su, B.-M.; Xie, J.-S.; Li, R.-K.; Yang, J.; Lin, J.; Ye, X.-Y. Preparation of bioactive neoagaroligosaccharides through hydrolysis of Gracilaria lemaneiformis agar: A comparative study. Food Chem. 2018, 240, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Liu, H.; Hou, Y.; Chan, Z.; Di, W.; Li, L.; Zeng, R. Preparation, characterization and alcoholic liver injury protective effects of algal oligosaccharides from Gracilaria lemaneiformis. Food Res. Int. 2017, 100, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhang, Y.; Shu, Z.; Liu, M.; Zeng, R.; Wang, Y.; Liu, H.; Cao, M.; Su, W.; Liu, G. Sulfated oligosaccharide of Gracilaria lemaneiformis protect against food allergic response in mice by up-regulating immunosuppression. Carbohydr. Polym. 2020, 230, 115567. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Pan, C.; Yang, X.; Chen, S.; Qi, B.; Huang, H. Degradation of Codium cylindricum polysaccharides by H2O2-Vc-ultrasonic and H2O2-Fe2+-ultrasonic treatment: Structural characterization and antioxidant activity. Int. J. Biol. Macromol. 2021, 182, 129–135. [Google Scholar] [CrossRef]
- Shen, X.; Liu, Z.; Li, J.; Wu, D.; Zhu, M.; Yan, L.; Mao, G.; Ye, X.; Linhardt, R.J.; Chen, S. Development of low molecular weight heparin by H2O2/ascorbic acid with ultrasonic power and its anti-metastasis property. Int. J. Biol. Macromol. 2019, 133, 101–109. [Google Scholar] [CrossRef]
- Shokri, Z.; Seidi, F.; Karami, S.; Li, C.; Saeb, M.R.; Xiao, H. Laccase Immobilization onto Natural Polysaccharides for Biosensing and Biodegradation. Carbohydr. Polym. 2021, 262, 117963. [Google Scholar] [CrossRef]
- Chen, H.; Xiao, Q.; Weng, H.; Zhang, Y.; Yang, Q.; Xiao, A. Extraction of sulfated agar from Gracilaria lemaneiformis using hydrogen peroxide-assisted enzymatic method. Carbohydr. Polym. 2020, 232, 115790. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, X.; Sun-Waterhouse, D.; Zhu, B.; You, L.; Hileuskaya, K. Polysaccharides from Sargassum fusiforme after UV/H2O2 degradation effectively ameliorate dextran sulfate sodium-induced colitis. Food Funct. 2021, 12, 11747–11759. [Google Scholar] [CrossRef]
- Li, J.; Li, S.; Liu, S.; Wei, C.; Yan, L.; Ding, T.; Linhardt, R.J.; Liu, D.; Ye, X.; Chen, S. Pectic oligosaccharides hydrolyzed from citrus canning processing water by Fenton reaction and their antiproliferation potentials. Int. J. Biol. Macromol. 2019, 124, 1025–1032. [Google Scholar] [CrossRef]
- Nasab, S.B.; Homaei, A.; Pletschke, B.I.; Salinas-Salazar, C.; Castillo-Zacarias, C.; Parra-Saldívar, R. Marine resources effective in controlling and treating diabetes and its associated complications. Process. Biochem. 2020, 92, 313–342. [Google Scholar] [CrossRef]
- Lim, J.; Ferruzzi, M.G.; Hamaker, B.R. Structural requirements of flavonoids for the selective inhibition of α-amylase versus α-glucosidase. Food Chem. 2022, 370, 130981. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Zhang, Y.; Sun-Waterhouse, D.; You, L.; Fu, X. Advantages of the polysaccharides from Gracilaria lemaneiformis over metformin in antidiabetic effects on streptozotocin-induced diabetic mice. RSC Adv. 2017, 7, 9141–9151. [Google Scholar] [CrossRef] [Green Version]
- Liao, X.; Yang, L.; Chen, M.; Yu, J.; Zhang, S.; Ju, Y. The hypoglycemic effect of a polysaccharide (GLP) from Gracilaria lemaneiformis and its degradation products in diabetic mice. Food Funct. 2015, 6, 2542–2549. [Google Scholar] [CrossRef]
- Zheng, Q.; Jia, R.-B.; Ou, Z.-R.; Li, Z.-R.; Zhao, M.; Luo, D.; Lin, L. Comparative study on the structural characterization and α-glucosidase inhibitory activity of polysaccharide fractions extracted from Sargassum fusiforme at different pH conditions. Int. J. Biol. Macromol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Lu, M.; Wang, S. Amylase-assisted extraction and antioxidant activity of polysaccharides from Gracilaria lemaneiformis. 3 Biotech 2017, 7, 341. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.-M.; Yang, Y.; Maleki, S.J.; Alcocer, M.; Xu, S.-S.; Shi, C.-L.; Cao, M.-J.; Liu, G.-M. Anti-Food Allergic Activity of Sulfated Polysaccharide from Gracilaria lemaneiformis is Dependent on Immunosuppression and Inhibition of p38 MAPK. J. Agric. Food Chem. 2016, 64, 4536–4544. [Google Scholar] [CrossRef]
- Ren, Y.; Zheng, G.; You, L.; Wen, L.; Li, C.; Fu, X.; Zhou, L. Structural characterization and macrophage immunomodulatory activity of a polysaccharide isolated from Gracilaria lemaneiformis. J. Funct. Foods 2017, 33, 286–296. [Google Scholar] [CrossRef]
- Lv, Q.-Q.; Cao, J.-J.; Liu, R.; Chen, H.-Q. Structural characterization, α-amylase and α-glucosidase inhibitory activities of polysaccharides from wheat bran. Food Chem. 2021, 341, 128218. [Google Scholar] [CrossRef]
- Fu, M.; Shen, W.; Gao, W.; Namujia, L.; Yang, X.; Cao, J.; Sun, L. Essential moieties of myricetins, quercetins and catechins for binding and inhibitory activity against α-Glucosidase. Bioorganic Chem. 2021, 115, 105235. [Google Scholar] [CrossRef]
- Cao, X.; Xia, Y.; Liu, D.; He, Y.; Mu, T.; Huo, Y.; Liu, J. Inhibitory effects of Lentinus edodes mycelia polysaccharide on α-glucosidase, glycation activity and high glucose-induced cell damage. Carbohydr. Polym. 2020, 246, 116659. [Google Scholar] [CrossRef]
- Zhao, M.; Bai, J.; Bu, X.; Yin, Y.; Wang, L.; Yang, Y.; Xu, Y. Characterization of selenized polysaccharides from Ribes nigrum L. and its inhibitory effects on α-amylase and α-glucosidase. Carbohydr. Polym. 2021, 259, 117729. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, Y.; Huang, D.; Chen, S.; Xia, Y.; Zhu, S. The inhibitory mechanism of chlorogenic acid and its acylated derivatives on α-amylase and α-glucosidase. Food Chem. 2022, 372, 131334. [Google Scholar] [CrossRef] [PubMed]
- Bharathi, D.; Siddlingeshwar, B.; Krishna, R.H.; Kirilova, E.M.; Divakar, D.D.; Alkheraif, A.A. Interaction of CuO and ZnO nanoparticles with 3-N-(N′-methylacetamidino) benzanthrone: A temperature dependent fluorescence quenching study. Inorg. Chem. Commun. 2021, 134, 109069. [Google Scholar] [CrossRef]
- Zhang, S.; Hong, H.; Zhang, H.; Chen, Z. Investigation of anti-aging mechanism of multi-dimensional nanomaterials modified asphalt by FTIR, NMR and GPC. Constr. Build. Mater. 2021, 305, 124809. [Google Scholar] [CrossRef]
- Pattanayak, S.; Chakraborty, S.; Biswas, S.; Chattopadhyay, D.; Chakraborty, M. Degradation of Methyl Parathion, a common pesticide and fluorescence quenching of Rhodamine B, a carcinogen using β-d glucan stabilized gold nanoparticles. J. Saudi Chem. Soc. 2018, 22, 937–948. [Google Scholar] [CrossRef]
- Wang, L.; Li, L.; Gao, J.; Huang, J.; Yang, Y.; Xu, Y.; Liu, S.; Yu, W. Characterization, antioxidant and immunomodulatory effects of selenized polysaccharides from dandelion roots. Carbohydr. Polym. 2021, 260, 117796. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Level | ||
|---|---|---|---|
| −1 | 0 | 1 | |
| A H2O2-Vc concentration/mM | 15 | 20 | 25 |
| B Time/h | 0.3 | 0.5 | 0.7 |
| C Temperature/°C | 40 | 50 | 60 |
| Code | A:H2O2-Vc Concentration/mM | B: Time/h | C:Temperature/°C | Y:Reducing Sugar Content/% |
|---|---|---|---|---|
| 1 | 0 | 0 | 0 | 41.6564 |
| 2 | 0 | 0 | 0 | 40.6566 |
| 3 | 1 | 0 | −1 | 32.9644 |
| 4 | 1 | 0 | 1 | 35.5119 |
| 5 | 0 | 1 | 1 | 39.2347 |
| 6 | 0 | 0 | 0 | 40.6399 |
| 7 | −1 | 0 | 1 | 40.1668 |
| 8 | 0 | 0 | 0 | 41.0565 |
| 9 | 0 | −1 | −1 | 34.0254 |
| 10 | 0 | −1 | 1 | 38.7351 |
| 11 | −1 | 1 | 0 | 36.8824 |
| 12 | 1 | 1 | 0 | 36.1679 |
| 13 | −1 | 0 | −1 | 34.2537 |
| 14 | 1 | −1 | 0 | 32.7301 |
| 15 | 0 | 0 | 0 | 40.0567 |
| 16 | −1 | −1 | 0 | 36.8499 |
| 17 | 0 | 1 | −1 | 36.5345 |
| Source | Sun of Squares | df | Mean Square | F Value | p Value | Significance |
|---|---|---|---|---|---|---|
| Model | 139.59 | 9 | 15.51 | 63.28 | <0.0001 | ** |
| A-H2O2-Vc concentration | 14.52 | 1 | 14.52 | 59.25 | 0.0001 | ** |
| B-Time | 5.25 | 1 | 5.25 | 21.41 | 0.0024 | ** |
| C-Temperature | 31.48 | 1 | 31.48 | 128.46 | <0.0001 | ** |
| AB | 2.90 | 1 | 2.90 | 11.83 | 0.0108 | * |
| AC | 2.83 | 1 | 2.83 | 11.55 | 0.0115 | * |
| BC | 1.01 | 1 | 1.01 | 4.12 | 0.0820 | |
| A2 | 45.35 | 1 | 45.35 | 185.04 | <0.0001 | ** |
| B2 | 14.78 | 1 | 14.78 | 60.31 | 0.0001 | ** |
| C2 | 13.75 | 1 | 13.75 | 56.10 | 0.0001 | ** |
| Residual | 1.72 | 7 | 0.25 | |||
| Lack of Fit | 0.32 | 3 | 0.11 | 0.30 | 0.8223 | |
| Pure Error | 1.40 | 4 | 0.35 | |||
| Cor Total | 141.31 | 16 |
| Indexes | GLP | GLP-HV | GLP-H | GLP-V | |
|---|---|---|---|---|---|
| Total sugar | Content (%) | 98.77 ± 5.94 a | 98.43 ± 3.21 a | 97.28 ± 2.63 a | 97.5 ± 1.56 a |
| Standard curve | y = 2.4623x + 0.0516, R2 = 0.9939 | ||||
| Reducing sugar | Content (%) | 2.47 ± 0.03 c | 46.92 ± 2.38 b | 1.95 ± 0.15 c | 50.2 ± 1.00 a |
| Standard curve | y = 0.8185x − 0.0258, R2 = 0.9999 | ||||
| Protein | Content (%) | ND | ND | ND | ND |
| Standard curve | y = 0.6288x + 0.5582, R2 = 0.9902 | ||||
| Molecular Weight (Da) | GLP | GLP-HV | GLP-H | GLP-V |
|---|---|---|---|---|
| Number average molecular weight(Mn) (Da) | 10,250 | 6695 | 17,508 | 21,224 |
| Weight average molecular weight(Mw) (Da) | 1,478,524 | 16,245 | 1,329,838 | 1,000,630 |
| Mw/Mn | 144.24 | 2.43 | 75.96 | 47.15 |
| [S] (mg/mL) | V max (mg/mL·min−1) | Km (mg/mL) | Inhibition Type |
|---|---|---|---|
| 0 | 0.0059 | 2.409 | The mix of competitive and non-competitive |
| 1 | 0.0030 | 4.164 | |
| 5 | 0.0029 | 7.839 |
| [Q] (mg/mL) | F | F0/F | Ksv (L/mol) | Kq (L/mol/s) | Ka (L/mol) | n |
|---|---|---|---|---|---|---|
| 0 | 419 | 1 | 0.07566 | 7.566 × 106 | 9.5082 | 0.8137 |
| 0.5 | 399 | 1.0518 | ||||
| 1 | 371 | 1.1309 | ||||
| 2.5 | 346 | 1.2107 | ||||
| 5 | 263 | 1.5930 | ||||
| 10 | 251 | 1.6690 | ||||
| equation of curve | F0/F= −1.23796∗e−[Q]/13.21797 + 2.24913, R2 = 0.99148 | lg((F0 − F)/F) = 0.8137∗lg[Q]−0.9781, R2 = 0.9788 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Long, X.; Hu, X.; Zhou, S.; Xiang, H.; Chen, S.; Li, L.; Liu, S.; Yang, X. Optimized Degradation and Inhibition of α-glucosidase Activity by Gracilaria lemaneiformis Polysaccharide and Its Production In Vitro. Mar. Drugs 2022, 20, 13. https://doi.org/10.3390/md20010013
Long X, Hu X, Zhou S, Xiang H, Chen S, Li L, Liu S, Yang X. Optimized Degradation and Inhibition of α-glucosidase Activity by Gracilaria lemaneiformis Polysaccharide and Its Production In Vitro. Marine Drugs. 2022; 20(1):13. https://doi.org/10.3390/md20010013
Chicago/Turabian StyleLong, Xiaoshan, Xiao Hu, Shaobo Zhou, Huan Xiang, Shengjun Chen, Laihao Li, Shucheng Liu, and Xianqing Yang. 2022. "Optimized Degradation and Inhibition of α-glucosidase Activity by Gracilaria lemaneiformis Polysaccharide and Its Production In Vitro" Marine Drugs 20, no. 1: 13. https://doi.org/10.3390/md20010013