
Screening for Health-Promoting Fatty Acids in Ascidians and Seaweeds Grown under the Influence of Fish Farming Activities

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
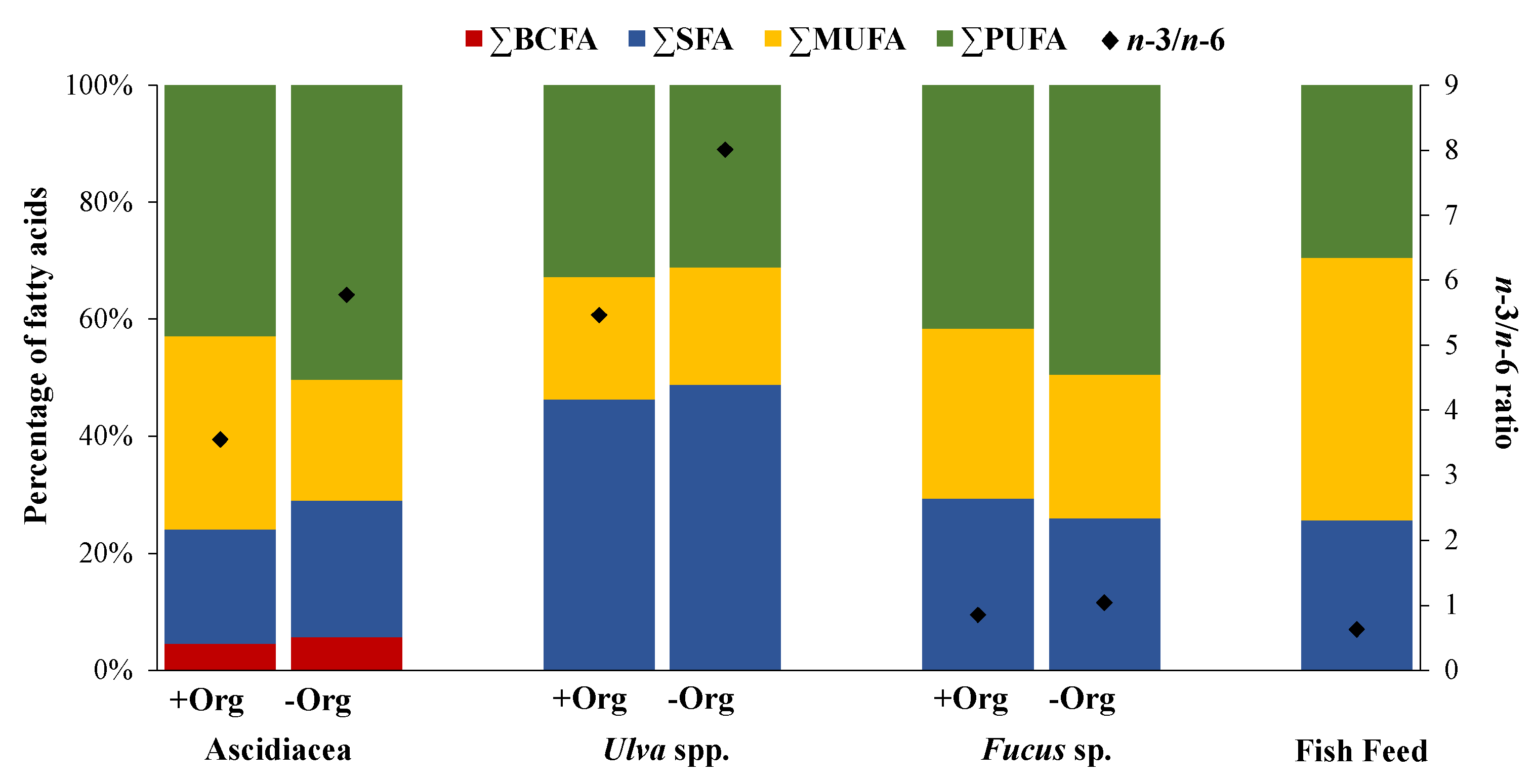
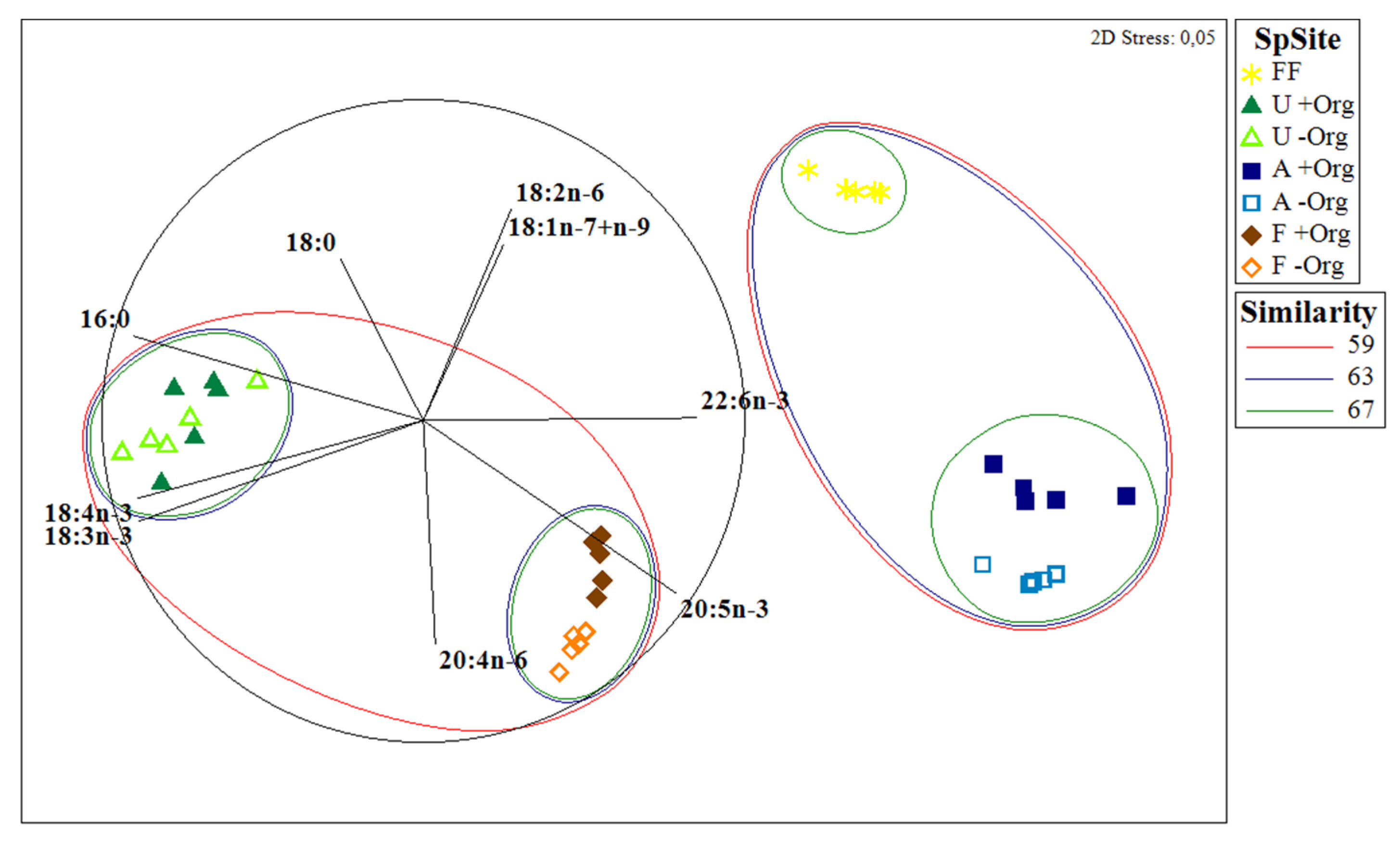
2. Results
2.1. Ascidiacea
2.2. Seaweeds
2.3. Fish Feed
3. Discussion
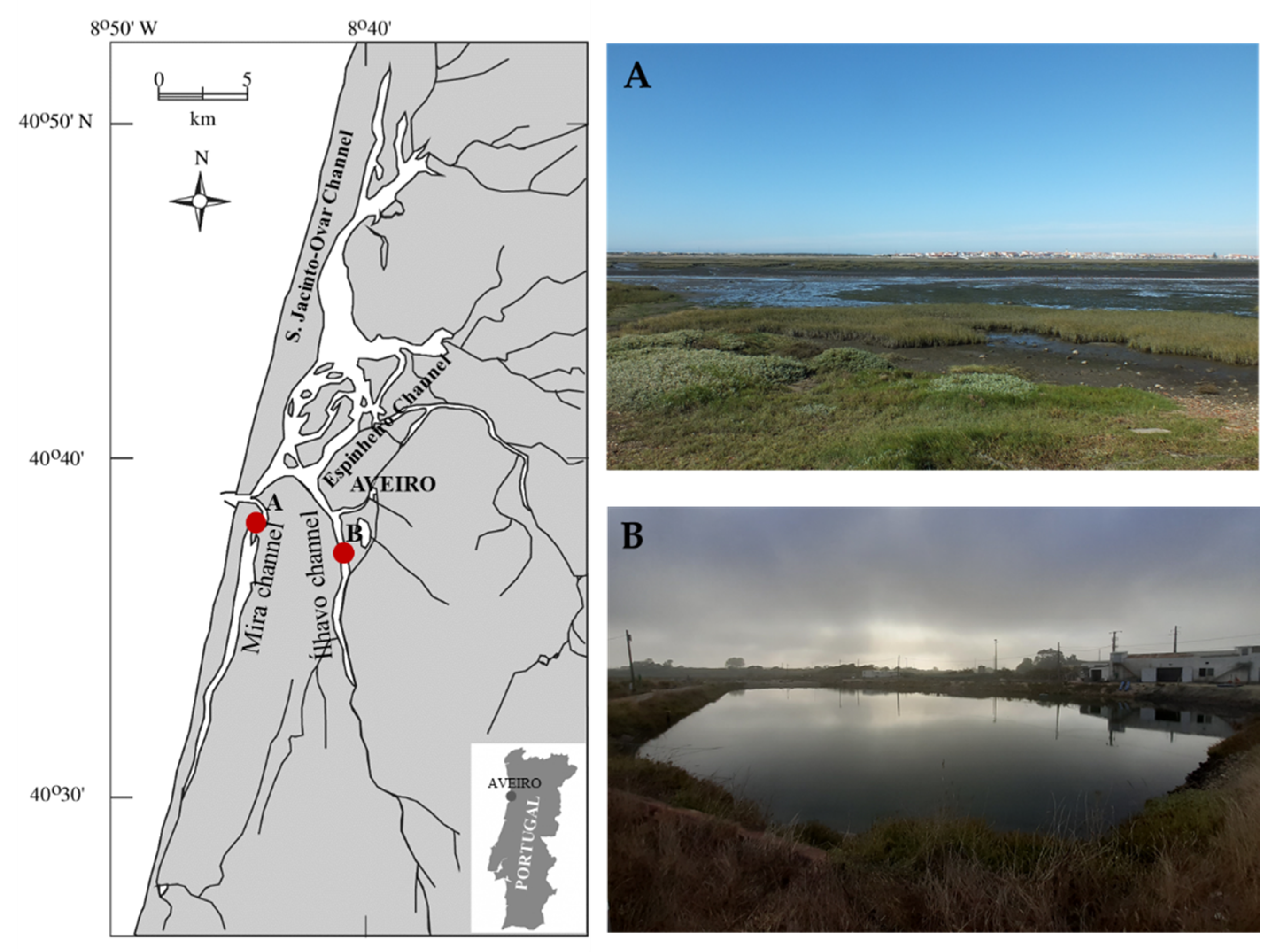
4. Materials and Methods
4.1. Study Areas
4.2. Sample Collection
4.2.1. Ascidiacea
4.2.2. Seaweeds
4.2.3. Fish Feed
4.3. Total Lipid Extraction
4.4. Fatty Acid Profiling
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Strobel, C.; Jahreis, G.; Kuhnt, K. Survey of n-3 and n-6 polyunsaturated fatty acids in fish and fish products. Lipids Health Dis. 2012, 11, 144. [Google Scholar] [CrossRef] [Green Version]
- Marques, B.; Lillebø, A.I.; Ricardo, F.; Nunes, C.; Coimbra, M.A.; Calado, R. Adding value to ragworms (Hediste diversicolor) through the bioremediation of a super-intensive marine fish farm. Aquac. Environ. Interact. 2018, 10, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Wang, M.; Lindström, M.E.; Li, J. Fatty acid and lipid profiles with emphasis on n-3 fatty acids and phospholipids from Ciona intestinalis. Lipids 2015, 50, 1009–1027. [Google Scholar] [CrossRef]
- Lopes, D.; Melo, T.; Rey, F.; Meneses, J.; Monteiro, F.L.; Helguero, L.A.; Abreu, M.H.; Lillebø, A.I.; Calado, R.; Domingues, M.R. Valuing bioactive lipids from green, red and brown macroalgae from aquaculture, to foster functionality and biotechnological applications. Molecules 2020, 25, 3883. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Candela, C.G.; López, L.M.B.; Kohen, V.L. Importance of a balanced omega 6/omega 3 ratio for the maintenance of health. Nutr. Recomm. Nutr. Hosp. 2011, 26, 323–329. [Google Scholar] [CrossRef]
- Swanson, D.; Block, R.; Mousa, S.A. Omega-3 fatty acids EPA and DHA: Health benefits throughout life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef]
- Béné, C.; Barange, M.; Subasinghe, R.; Pinstrup-Andersen, P.; Merino, G.; Hemre, G.I.; Williams, M. Feeding 9 billion by 2050—Putting fish back on the menu. Food Secur. 2015, 7, 261–274. [Google Scholar] [CrossRef] [Green Version]
- FAO. The State of World Fisheries and Aquaculture; FAO: Rome, Italy, 2020; ISBN 9789251326923. [Google Scholar]
- Abreu, M.H.; Pereira, R.; Sassi, J.F. Marine algae and the global food industry. In Marine Algae Biodiversity, Taxonomy, Environmental Assessment, and Biotechnology; CRC Press: Boca Raton, FL, USA, 2014; pp. 300–319. [Google Scholar] [CrossRef]
- Olsen, Y. Resources for fish feed in future mariculture. Aquac. Environ. Interact. 2011, 1, 187–200. [Google Scholar] [CrossRef] [Green Version]
- Engle, C.; Abramo, L.R.D.; Slater, M.J. Global Aquaculture 2050. J. World Aquac. Soc. 2017, 48, 3–6. [Google Scholar] [CrossRef] [Green Version]
- Alexander, K.A.; Angel, D.; Freeman, S.; Israel, D.; Johansen, J.; Kletou, D.; Meland, M.; Pecorino, D.; Rebours, C.; Rousou, M.; et al. Improving sustainability of aquaculture in Europe: Stakeholder dialogues on Integrated Multi-trophic Aquaculture (IMTA). Environ. Sci. Policy 2016, 55, 96–106. [Google Scholar] [CrossRef]
- Hasan, M.; Halwart, M. Fish as Feed Inputs for Aquaculture: Practices, Sustainability and Implications; FAO: Rome, Italy, 2009; ISBN 9789251064191. [Google Scholar]
- Custódio, M.; Villasante, S.; Calado, R.; Lillebø, A.I. Valuation of ecosystem services to promote sustainable aquaculture practices. Rev. Aquac. 2020, 12, 392–405. [Google Scholar] [CrossRef] [Green Version]
- Hodar, A.R.; Vasava, R.; Joshi, N.H. Fish meal and fish oil replacement for aquafeed formulation by using alternative sources: A review. J. Exp. Zool. India 2020, 23, 13–21. [Google Scholar]
- Apper-bossard, E.; Feneuil, A.; Wagner, A.; Respondek, F. Use of vital wheat gluten in aquaculture feeds. Aquat. Biosyst. 2013, 9, 21. [Google Scholar] [CrossRef] [Green Version]
- Metwalli, A.A.A. Effects of partial and total substitution of fish meal with corn gluten meal on growth performance, nutrients utilization and some blood constituents of the Nile tilapia Oreochromis niloticus. Egypt J. Aquat. Biol. Fish. 2013, 17, 91–100. [Google Scholar]
- Iqbal, M.; Yaqub, A.; Ayub, M. Partial and full substitution of fish meal and soybean meal by canola meal in diets for genetically improved farmed tilapia (O. niloticus): Growth performance, carcass composition, serum biochemistry, immune response, and intestine histology. J. Appl. Aquac. 2021, 1–26. [Google Scholar] [CrossRef]
- Soller, F.; Roy, L.A.; Davis, D.A. Replacement of fish oil in plant-based diets for Pacific white shrimp, Litopenaeus vannamei, by stearine fish oil and palm oil. Fundam. Stud. 2018, 50, 186–203. [Google Scholar] [CrossRef] [Green Version]
- Ayisi, C.L.; Zhao, J.; Wu, J. Replacement of fish oil with palm oil: Effects on growth performance, innate immune response, antioxidant capacity and disease resistance in Nile tilapia (Oreochromis niloticus). PLoS ONE 2018, 13, e0196100. [Google Scholar] [CrossRef] [Green Version]
- Makkar, H.P.; Tran, G.; Heuzé, V.; Giger-Reverdin, S.; Lessire, M.; Lebas, F.; Ankers, P. Seaweeds for livestock diets: A review. Anim. Feed Sci. Technol. 2016, 212, 1–17. [Google Scholar] [CrossRef]
- Glencross, B.D. Exploring the nutritional demand for essential fatty acids by aquaculture species. Rev. Aquac. 2009, 1, 71–124. [Google Scholar] [CrossRef]
- Broadhurst, C.L.; Wang, Y.; Crawford, M.A.; Cunnane, S.C.; Parkington, J.E.; Schmidt, W.F. Brain-specific lipids from marine, lacustrine, or terrestrial food resources: Potential impact on early African Homo sapiens. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2002, 131, 653–673. [Google Scholar] [CrossRef]
- Ju, B.; Jiang, A.; Xing, R.; Chen, L.; Teng, L. Optimization of conditions for an integrated multi- trophic aquaculture system consisting of sea cucumber Apostichopus japonicus and ascidian Styela clava. Aquac. Int. 2016, 25, 265–286. [Google Scholar] [CrossRef]
- Chatzoglou, E.; Kechagia, P.; Tsopelakos, A.; Miliou, H.; Slembrouck, J. Co-culture of Ulva sp. and Dicentrarchus labrax in Recirculating Aquaculture System: Effects on growth, retention of nutrients and fatty acid profile. Aquat. Living Resour. 2020, 33, 19. [Google Scholar] [CrossRef]
- Marques, B.; Calado, R.; Lillebø, A.I. New species for the biomitigation of a super-intensive marine fish farm effluent: Combined use of polychaete-assisted sand filters and halophyte aquaponics. Sci. Total Environ. 2017, 599–600, 1922–1928. [Google Scholar] [CrossRef]
- Jerónimo, D.; Lillebø, A.I.; Cremades, J.; Cartaxana, P.; Calado, R. Recovering wasted nutrients from shrimp farming through the combined culture of polychaetes and halophytes. Sci. Rep. 2021, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Cripps, S.J.; Bergheim, A. Solids management and removal for intensive land-based aquaculture production systems. Aquac. Eng. 2000, 22, 33–56. [Google Scholar] [CrossRef]
- Sprague, M.; Dick, J.R.; Tocher, D.R. Impact of sustainable feeds on omega-3 long-chain fatty acid levels in farmed Atlantic salmon. Sci. Rep. 2016, 6, 21892. [Google Scholar] [CrossRef] [Green Version]
- Aguado-Giménez, F.; Hernández, M.D.; Cerezo-Valverde, J.; Piedecausa, M.A.; García-García, B. Does flat oyster (Ostrea edulis) rearing improve under open-sea integrated multi-trophic conditions? Aquac. Int. 2014, 22, 447–467. [Google Scholar] [CrossRef]
- White, C.A.; Woodcock, S.H.; Bannister, R.J.; Nichols, P.D. Terrestrial fatty acids as tracers of finfish aquaculture waste in the marine environment. Rev. Aquac. 2017, 11, 133–148. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Jover, D.; Arechavala-Lopez, P.; Martinez-Rubio, L.; Tocher, D.R.; Bayle-Sempere, J.T.; Lopez-Jimenez, J.A.; Martinez-Lopez, F.; Sanchez-Jerez, P. Monitoring the influence of marine aquaculture on wild fish communities: Benefits and limitations of fatty acid profiles. Aquac. Environ. Interact. 2011, 2, 39–47. [Google Scholar] [CrossRef]
- Palanisamy, S.K.; Rajendran, N.M.; Marino, A. Natural products diversity of marine ascidians (Tunicates; Ascidiacea) and successful drugs in clinical development. Nat. Prod. Bioprospect. 2017, 7, 1–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monmai, C.; Go, S.H.; Shin, I.-S.; You, S.G.; Lee, H.; Kang, S.B.; Park, W.J. Immune-enhancement and anti-inflammatory activities of fatty acids extracted from Halocynthia aurantium tunic in RAW264.7 cells. Mar. Drugs 2018, 16, 309. [Google Scholar] [CrossRef] [Green Version]
- Dagorn, F.; Dumay, J.; Wielgosz-Collin, G.; Rabesaotra, V.; Viau, M.; Monniot, C.; Biard, J.-F.; Barnathan, G. Phospholipid distribution and phospholipid fatty acids of the tropical tunicates Eudistoma sp. and Leptoclinides uniorbis. Lipids 2010, 45, 253–261. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, J. Ascidian bioresources: Common and variant chemical compositions and exploitation strategy—examples of Halocynthia roretzi, Styela plicata, Ascidia sp. and Ciona intestinalis. Zeitschrift fur Naturforsch 2016, 71, 165–180. [Google Scholar] [CrossRef] [PubMed]
- Hassanzadeh, M. Unique marine organism: Identification of some methods for biomaterial production. Chem. Eng. Trans. 2014, 37, 385–390. [Google Scholar] [CrossRef]
- Choi, D.G.; Kim, J.; Yun, A.; Cho, S.H.; Jeong, H.S.; Lee, K.W.; Kim, H.S.; Kim, P.Y.; Ha, M.S. Dietary substitution effect of fishmeal with tunic meal of sea squirt, Halocynthia roretzi, Drasche, on growth and soft body composition of juvenile abalone, Haliotis discus, Reeve 1846. J. World Aquac. Soc. 2018, 49, 1095–1104. [Google Scholar] [CrossRef]
- Jang, B.; Kim, P.Y.; Kim, H.S.; Lee, K.W.; Kim, H.J.; Choi, D.G.; Cho, S.H.; Min, B.; Kim, K.; Han, H. Substitution effect of sea tangle (ST) (Laminaria japonica) with tunic of sea squirt (SS) (Halocynthia roretzi) in diet on growth and carcass composition of juvenile abalone (Haliotis discus, Reeve 1846). Aquac. Nutr. 2017, 24, 586–593. [Google Scholar] [CrossRef]
- Giangrande, A.; Pierri, C.; Arduini, D.; Borghese, J.; Licciano, M.; Trani, R.; Corriero, G.; Basile, G.; Cecere, E.; Petrocelli, A.; et al. An innovative IMTA system: Polychaetes, sponges and macroalgae co-cultured in a Southern Italian in-shore mariculture plant (Ionian Sea). J. Mar. Sci. Eng. 2020, 8, 733. [Google Scholar] [CrossRef]
- Vega, J.; Álvarez-Gómez, F.; Güenaga, L.; Figueroa, F.L.; Gómez-Pinchetti, J.L. Antioxidant activity of extracts from marine macroalgae, wild-collected and cultivated, in an integrated multi-trophic aquaculture system. Aquaculture 2020, 522, 735088. [Google Scholar] [CrossRef]
- Ashkenazi, D.Y.; Israel, A.; Abelson, A. A novel two-stage seaweed integrated multi-trophic aquaculture. Rev. Aquac. 2019, 11, 246–262. [Google Scholar] [CrossRef] [Green Version]
- Da Costa, E.; Domingues, P.; Melo, T.; Coelho, E.; Pereira, R.; Calado, R.; Abreu, M.H.; Domingues, M.R. Lipidomic signatures reveal seasonal shifts on the relative abundance of high-valued lipids from the brown algae Fucus vesiculosus. Mar. Drugs 2019, 17, 335. [Google Scholar] [CrossRef] [Green Version]
- Wan, A.H.L.; Davies, S.J.; Soler-Vila, A.; Fitzgerald, R.; Johnson, M.P. Macroalgae as a sustainable aquafeed ingredient. Rev. Aquac. 2019, 11, 458–492. [Google Scholar] [CrossRef]
- Pirian, K.; Jeliani, Z.Z.; Arman, M.; Sohrabipour, J.; Yousefzadi, M. Proximate analysis of selected macroalgal species from The Persian Gulf as a nutritional resource. Trop. Life Sci. Res. 2020, 31, 1–17. [Google Scholar] [CrossRef]
- Moreira, A.S.P.; da Costa, E.; Melo, T.; Sulpice, R.; Cardoso, S.M.; Pitarma, B.; Pereira, R.; Abreu, M.H.; Domingues, P.; Calado, R.; et al. Seasonal plasticity of the polar lipidome of Ulva rigida cultivated in a sustainable integrated multi-trophic aquaculture. Algal Res. 2020, 49, 101958. [Google Scholar] [CrossRef]
- Jeong, B.Y.; Ohshima, T.; Koizumi, C. Hydrocarbon chain distribution of ether phospholipids of the ascidian Halocynthia roretzi and the sea urchin Strongylocentrotus intermedius. Lipids 1996, 31, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Sri Kumaran, N.; Bragadeeswaran, S. Nutritional composition of the colonial ascidian Eudistoma viride and Didemnum psammathodes. Biosci. Biotechnol. Res. Asia 2014, 11, 331–338. [Google Scholar] [CrossRef]
- Maoufoud, S.; Abdelmjid, A.; Abboud, Y.; Tarik, A. Chemical composition of fatty acids and sterols from tunicates Cynthia savignyi, Cynthia squamulata and from the brown alga Cystoseira tamariscifolia. Phys. Chem. News 2009, 47, 115–119. [Google Scholar]
- Schmid, M.; Guihéneuf, F.; Stengel, D.B. Fatty acid contents and profiles of 16 macroalgae collected from the Irish Coast at two seasons. J. Appl. Phycol. 2014, 26, 451–463. [Google Scholar] [CrossRef]
- Kim, M.-K.; Dubacq, J.-P.; Thomas, J.-C.; Giraud, G. Seasonal variations of triacylglycerols and fatty acids in Fucus serratus. Phytochemistry 1996, 43, 49–55. [Google Scholar] [CrossRef]
- Morais, T.; Cotas, J.; Pacheco, D.; Pereira, L. Seaweeds compounds: An ecosustainable source of comestic ingredients? Cosmetics 2021, 8, 8. [Google Scholar] [CrossRef]
- Herbreteau, F.; Coiffard, L.J.M.; Derrien, A.; De Roeck-Holtzhauer, Y. The fatty acid composition of five species of macroalgae. Bot. Mar. 1997, 40, 25–28. [Google Scholar] [CrossRef]
- Silva, G.; Pereira, R.B.; Valentão, P.; Paula, B. Distinct fatty acid profile of ten brown macroalgae. Brazilian J. Pharmacogn. 2013, 23, 608–613. [Google Scholar] [CrossRef] [Green Version]
- Kendel, M.; Wielgosz-Collin, G.; Bertrand, S.; Roussakis, C.; Bourgougnon, N.B.; Bedoux, G. Lipid composition, fatty acids and sterols in the seaweeds Ulva armoricana, and Solieria chordalis from brittany (France): An analysis from nutritional, chemotaxonomic, and antiproliferative activity perspectives. Mar. Drugs 2015, 13, 5606–5628. [Google Scholar] [CrossRef] [PubMed]
- Pereira, H.; Barreira, L.; Figueiredo, F.; Custódio, L.; Vizetto-Duarte, C.; Polo, C.; Rešek, E.; Aschwin, E.; Varela, J. Polyunsaturated fatty acids of marine macroalgae: Potential for nutritional and pharmaceutical applications. Mar. Drugs 2012, 10, 1920–1935. [Google Scholar] [CrossRef] [Green Version]
- van Vliet, T.; Katan, B. Lower ratio of n-3 to n-6 fatty acids in cultured than in wild fish. Am. J. Clin. Nutr. 1990, 51, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, M.F.; Borum, J. Nutrient control of estuarine macroalgae: Growth strategy and the balance between nitrogen requirements and uptake. Mar. Ecol. Prog. Ser. 1997, 161, 155–163. [Google Scholar] [CrossRef]
- García-Alonso, J.; Müller, C.T.; Hardege, J.D. Influence of food regimes and seasonality on fatty acid composition in the ragworm. Aquat. Biol. 2008, 4, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Luis, O.J.; Passos, A.M. Seasonal changes in lipid content and composition of the polychaete Nereis (Hediste) diversicolor. Comp. Biochem. Physiol. Part B Biochem. 1995, 111, 579–586. [Google Scholar] [CrossRef]
- Prato, E.; Danieli, A.; Maffia, M.; Biandolino, F. Lipid contents and fatty acid compositions of Idotea baltica and Sphaeroma serratum (Crustacea: Isopoda) as indicators of food sources. Zool. Stud. 2012, 51, 38–50. [Google Scholar]
- Chetoui, I.; Rabeh, I.; Bejaoui, S.; Telahigue, K.; Ghribi, F.; El Cafsi, M. First seasonal investigation of the fatty acid composition in three organs of the Tunisian bivalve Mactra stultorum. Grasas y Aceites 2019, 70, 291. [Google Scholar] [CrossRef] [Green Version]
- Ezgeta-Balić, D.; Najdek, M.; Peharda, M.; Blažina, M. Seasonal fatty acid profile analysis to trace origin of food sources of four commercially important bivalves. Aquaculture 2012, 334–337, 89–100. [Google Scholar] [CrossRef]
- Zlatanos, S.; Laskaridis, K. Seasonal variation in the fatty acid composition of three Mediterranean fish—sardine (Sardina pilchardus), anchovy (Engraulis encrasicholus) and picarel (Spicara smaris). Food Chem. 2007, 103, 725–728. [Google Scholar] [CrossRef]
- Vieira, H.; Leal, M.C.; Calado, R. Fifty shades of blue: How blue biotechnology is shaping the bioeconomy. Trends Biotechnol. 2020, 38, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Melo, T.; Alves, E.; Azevedo, V.; Martins, A.S.; Neves, B.M.; Domingues, P.; Calado, R.; Abreu, M.H.; Domingues, M.R. Lipidomics as a new approach for the bioprospecting of marine macroalgae—Unraveling the polar lipid and fatty acid composition of Chondrus crispus. Algal Res. 2015, 8, 181–191. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Ascidiacea | Ulva spp. | Fucus sp. | Fish Feed | ||||
|---|---|---|---|---|---|---|---|
| +Org | −Org | +Org | −Org | +Org | −Org | ||
| 14:0 | 0.94 ± 0.17 | 1.45 ± 0.12 | 0.68 ± 0.22 | 0.64 ± 0.17 | 8.04 ± 0.64 | 8.47 ± 0.27 | 1.53 ± 0.35 |
| 16:0 | 11.50 ± 1.31 | 12.56 ± 0.67 | 37.74 ± 1.14 | 38.05 ± 1.86 | 16.17 ± 1.29 | 15.03 ± 0.62 | 17.25 ± 0.68 |
| 16:1n-9 | 5.78 ± 0.62 | 5.37 ± 0.29 | 3.33 ± 0.27 | 2.67 ± 0.27 | 0.25 ± 0.04 | 0.29 ± 0.03 | 3.62 ± 0.18 |
| 16:4n-3 | n.d | n.d | 5.18 ± 0.33 | 4.27 ± 0.67 | 0.59 ± 0.06 | 0.59 ± 0.05 | n.d |
| 18:0 | 4.87 ± 1.23 | 5.89 ± 0.53 | 6.58 ± 3.99 | 8.70 ± 2.29 | 4.34 ± 1.61 | 1.77 ± 0.16 | 6.51 ± 1.09 |
| 18:1n-7+n-9 | 20.27 ± 1.80 | 11.98 ± 0.95 ** | 15.23 ± 1.21 | 15.19 ± 1.22 | 26.50 ± 2.28 | 21.34 ± 1.51 * | 35.97 ± 0.43 |
| 18:2n-6 | 5.85 ± 1.62 | 2.26 ± 0.08 * | 4.41 ± 0.19 | 2.74 ± 0.41 | 6.82 ± 0.38 | 7.45 ± 0.21 * | 16.86 ± 0.19 |
| 18:3n-3 | 2.16 ± 0.22 | 2.38 ± 0.48 | 8.95 ± 0.70 | 7.85 ± 0.57 * | 6.96 ± 0.41 | 8.87 ± 0.51 ** | 2.85 ± 0.07 |
| 18:4n-3 | 1.54 ± 0.61 | 3.61 ± 0.69 | 9.72 ± 0.65 | 10.10 ± 0.72 | 3.70 ± 0.36 | 5.55 ± 0.62 | 0.62 ± 0.05 |
| 20:4n-6 | 2.43 ± 0.37 | 3.11 ± 0.27 | n.d | n.d | 14.08 ± 1.17 | 15.03 ± 0.22 | 0.47 ± 0.03 |
| 20:5n-3 | 17.77 ± 2.90 | 20.44 ± 1.00 | 0.61 ± 0.13 | 1.25 ± 1.14 | 7.66 ± 0.74 | 9.95 ± 0.39 ** | 2.13 ± 0.11 |
| 22:6n-3 | 8.75 ± 1.00 | 11.85 ± 1.01 ** | n.d | n.d | n.d | n.d | 4.59 ± 0.32 |
| ∑n-3 | 32.03 ± 3.62 | 40.07 ± 1.54 * | 27.35 ± 1.87 | 27.61 ± 2.30 | 19.16 ± 1.54 | 25.24 ± 1.42 ** | 11.43 ± 0.51 |
| ∑n-6 | 9.02 ± 1.25 | 6.94 ± 0.46 * | 5.00 ± 0.24 | 3.45 ± 0.44 ** | 22.42 ± 1.59 | 24.18 ± 0.07 * | 18.09 ± 0.23 |
| ∑n-3/∑n-6 | 3.66 ± 0.98 | 5.79 ± 0.37 * | 5.46 ± 0.25 | 8.04 ± 0.36 ** | 0.85 ± 0.03 | 1.04 ± 0.06 ** | 0.63 ± 0.03 |
| ∑SFA | 19.52 ± 2.36 | 22.39 ±1.00 * | 46.30 ± 3.35 | 48.78 ± 3.37 | 29.35 ± 3.48 | 26.02 ± 0.50 | 25.72 ± 1.42 |
| ∑MUFA | 32.99 ± 0.92 | 19.95 ± 1.39 ** | 20.88 ± 1.62 | 20.07 ± 1.66 | 29.07 ± 2.32 | 24.42 ± 1.48 * | 44.77 ± 0.81 |
| ∑PUFA | 42.81 ± 2.65 | 48.48 ± 1.80 * | 32.82 ± 1.94 | 31.19 ± 2.73 | 41.58 ± 3.08 | 49.43 ± 1.42 ** | 29.52 ± 0.64 |
| Permanova | ||
|---|---|---|
| +Org vs. −Org | ||
| Fatty Acids | Fatty Acids Classes | |
| Ascidiacea | 0.006 | 0.011 |
| Ulva spp. | 0.021 | 0.341 |
| Fucus sp. | 0.013 | 0.013 |
| Dissimilarity | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (A) | Ascidiacea | Ulva spp. | Fucus sp. | |||||||||
| +Org vs. −Org | +Org vs. −Org | +Org vs. −Org | ||||||||||
| 10.62% | 5.29% | 5.48% | ||||||||||
| −Org | +ORW | −ORW | Contrib% | +Org | −Org | Contrib% | +Org | −Org | Contrib% | |||
| vs. | 20:1n-9 | 2.21 | 1.03 | 15.81 | 18:0 | 2.48 | 2.93 | 23.19 | 18:0 | 2.05 | 1.33 | 22.78 |
| +Org | 18:1n-7+n-9 | 4.50 | 3.46 | 13.92 | 18:2n-6 | 2.10 | 1.65 | 14.23 | 18:1n-7+n-9 | 5.14 | 4.62 | 16.65 |
| 18:2n-6 | 2.40 | 1.50 | 11.94 | 22:5n-3 | 1.54 | 1.91 | 11.59 | 18:4n-3 | 1.92 | 2.35 | 13.56 | |
| 18:4n-3 | 1.22 | 1.89 | 8.98 | 20:5n-3 | 0.78 | 1.04 | 9.76 | |||||
| (B) | Ascidiacea | |||||||||||
| +Org vs. FF | −Org vs. FF | |||||||||||
| 31.06% | 36.35% | |||||||||||
| +Org | FF | Contrib% | −Org | FF | Contrib% | |||||||
| Org | 20:5n-3 | 4.21 | 1.46 | 13.91 | 20:5n-3 | 4.52 | 1.46 | 13.33 | ||||
| vs. | 18:2n-6 | 2.40 | 4.11 | 8.65 | 18:2n-6 | 1.50 | 4.11 | 11.34 | ||||
| FF | 20:4n-6 | 1.55 | 0 | 7.87 | 18:1n-7+n-9 | 3.46 | 6.00 | 11.06 | ||||
| 18:1n-7+n-9 | 4.5 | 6.00 | 7.58 | 18:4n-3 | 1.89 | 0 | 8.24 | |||||
| 22:1n-11 | 0 | 1.36 | 7.05 | 20:4n-6 | 1.76 | 0 | 7.68 | |||||
| 18:4n-3 | 1.22 | 0 | 6.18 | |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marques, L.; Domingues, M.R.; da Costa, E.; Abreu, M.H.; Lillebø, A.I.; Calado, R. Screening for Health-Promoting Fatty Acids in Ascidians and Seaweeds Grown under the Influence of Fish Farming Activities. Mar. Drugs 2021, 19, 469. https://doi.org/10.3390/md19080469
Marques L, Domingues MR, da Costa E, Abreu MH, Lillebø AI, Calado R. Screening for Health-Promoting Fatty Acids in Ascidians and Seaweeds Grown under the Influence of Fish Farming Activities. Marine Drugs. 2021; 19(8):469. https://doi.org/10.3390/md19080469
Chicago/Turabian StyleMarques, Luísa, Maria Rosário Domingues, Elisabete da Costa, Maria Helena Abreu, Ana Isabel Lillebø, and Ricardo Calado. 2021. "Screening for Health-Promoting Fatty Acids in Ascidians and Seaweeds Grown under the Influence of Fish Farming Activities" Marine Drugs 19, no. 8: 469. https://doi.org/10.3390/md19080469