
Pharmacological Activities of Sulfated Fucose-Rich Polysaccharides after Oral Administration: Perspectives for the Development of New Carbohydrate-Based Drugs

Abstract
:1. Introduction

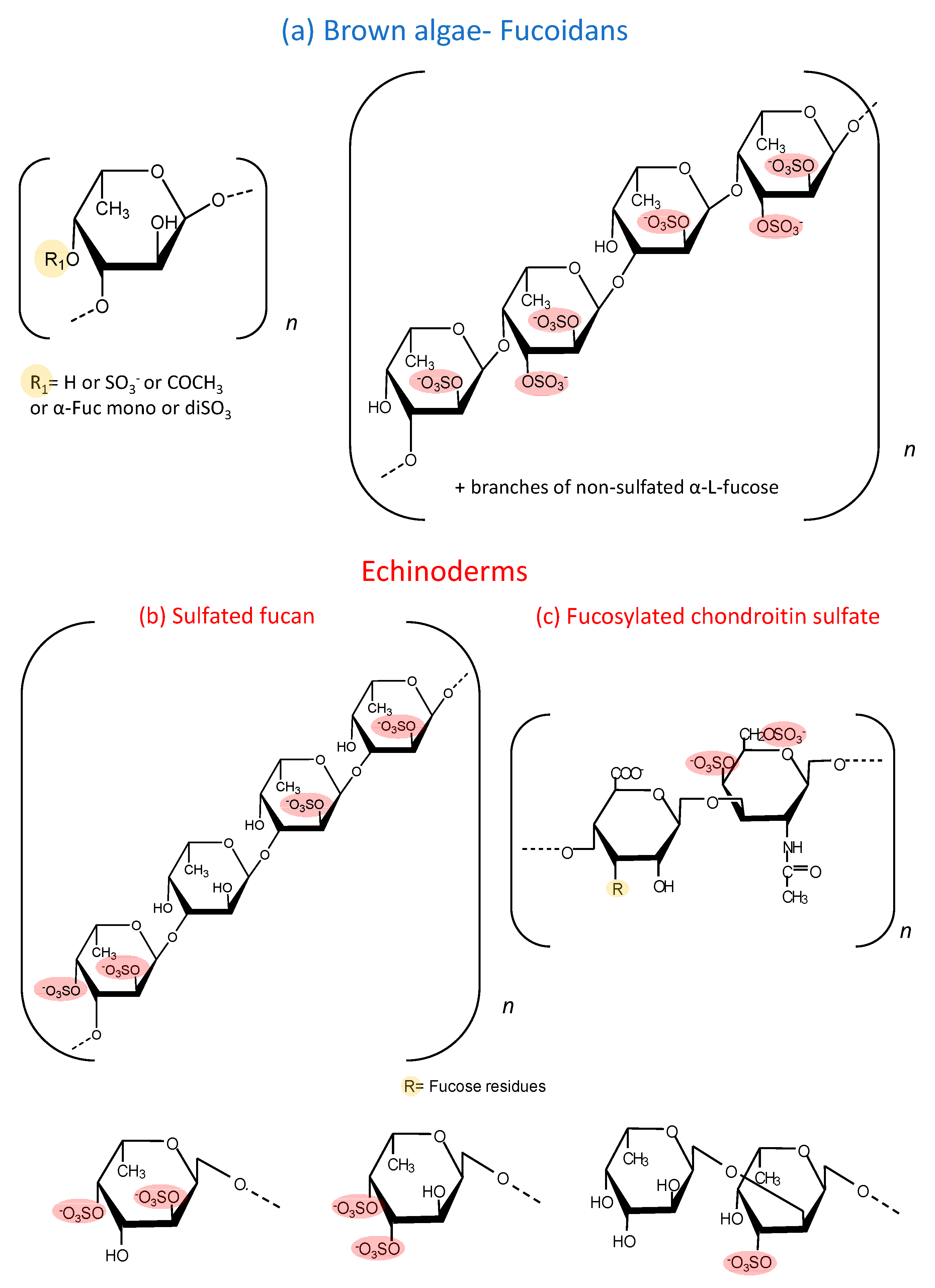{kind=link}
{kind=link}
| Species | Structure | Sugar and Sulfate Content | Mw (kDa) | Ref |
|---|---|---|---|---|
| A. nodosum | 1→3)-α-l-Fucp and a few (1→4)-α-l-Fucp with →3-α-l-(2 and/or 4 Fucp) | The carbohydrate and sulfate content of the fraction A3 were 74.7% and 12.0%, respectively. | 97.52 | [22] |
| S. henslowianum | →3)-α-l-Fucp(2 SO3−)-(1→3)-α-l-Fucp (4 SO3−)-(1→ | Fucose and glucose as main sugars. Sulfate content: 25.20%. | ND | [23] |
| F. evanescens | →3)-α-l-Fucp(2 SO3−)-(1→4)-α-l-Fucp(2 SO3−)-(1→ | Fucose, sulfate and acetyl groups at a molar ratio of 1:1.23:0.36 and trace amounts of galactose and xylose. | 10–100 | [24] |
| F. vesiculosus | →3)-α-l-Fucp(2 SO3−)-(1→4)-α-l-Fucp(2,3-SO3−)-(1→3) | 55.9% of carbohydrates, 27.0% of sulfate residues and 5.7% of uronic acid. Carbohydrates were represented mainly by fucose (38%), galactose (3.5%), xylose (2.7%). | 20.7 | [25] |
| C. okamuranus | →3)-α-l-Fucp (SO3−)-(1→3)-α-l-Fucp(4 SO3−)-(1→ | The glucuronic acid residues are linked to the C-2 positions of the fucose residues, which are not substituted by a sulfate group. Sulfate content ~15%. | 92.1 | [26] |
| U. pinnatifida | →3)-α-l-Fucp(2 SO3−)-(1→4)-α-l-Fucp(2,3-di SO3−)-(1→3) | This sulphated galactofucan is composed of: galactose 44.6% and fucose 50.9%. Xylose (4.2%), mannose (0.3%). Sulfate content 15%. A significant number of O-acetyl groups. | 378 | [27] |
| S. japonica | →3)-α-l-Fucp(2 SO3−)-(1→4)-α-l-Fucp(2,3-di SO3−)-(1→3) | 79.49% of fucose and 16.76% of galactose. Sulfate content ~30.72%. | 30 | [28] |
| Mozuku (High molecular weight fraction) | ND | Sulfate content: 13%. | 240 | [29] |
| L. japonica | →3)-α-l-Fucp(4 SO3−)-(1→ | 46.5% fucoxanthin, 8.01% lipids and 45.4% carbohydrates of mostly cellulose. Sulfate content: 13%. | 300 | [30] |
| Species | Proportions of the Branching Sulfated Fucose Units | Mw (kDa) | Ref |
|---|---|---|---|
| P. graeffei | 81.6% α-Fuc-4SO4, 18.4% α-Fuc-2,4diSO4 | 49 kDa | [31] |
| I. badionotus | 4.1% α-Fuc-4SO4, 95.9% α-Fuc-2,4diSO4 | 70.4 kDa | [32] |
| L. grisea | ~27% α-Fuc-2,4diSO4; ~20% α-Fuc3,4diSO4 and ~53% disaccharides composed of α-Fuc1→2-α-Fuc-3SO4→ | 40 kDa | [20] |
| C. frondosa | The chemical composition contained mainly glucuronic acid, galactosamine and fucose in the molar ratio of 1:1.50:1.16, with 30.07% sulfate content. | 14.76 kDa | [33] |
2. Antidislipidemic Effect
| Polysaccharide | Dosage Regimen and Species | Major Observations and Mechanism Proposed | Ref. |
|---|---|---|---|
| Fucoidan from A. nodosum | 100 mg/kg/day, 4 weeks, Mice | Improvement of reverse cholesterol transport and bile acid synthesis related genes expression. Reduction of plasma TC (~23.2%) and triglyceride (~48.7%) levels. | [22] |
| Fucoidan from A. nodosum | 50 and 100 mg/kg/day, 8 weeks, ApoE−/− mice | Reduction of hepatotoxicity induced by high-fat diet; increased plasma LPL activity, apoA1 level and protein expression of PPARα/β (∼2-fold), improved fatty acid oxidation and TG lowering (∼24.5%). | [37] |
| Fucoidan from S. henslowianum | 100 mg/kg/day, 4 weeks, Obese mice | Decreased cholesterol and LDL levels by ~23% and 18%, respectively. | [23] |
| Glycosaminoglycans from M. scabra | 5, 10, 20 and 50 mg/kg, 6 weeks, Rats | Inhibition of HMG-CoA reductase and/or increased lipoprotein lipase activity and metabolism of cholesterol. | [36] |
| FucCS and sulfated fucan from P. graeffei and from I. badionotus | 40 mg/kg, 8 days, Rats on high-fat diet | Hypolipidemic activity of sulfated polysaccharides is determined by the molecular dynamics of the sulfated polysaccharide. | [38] |
3. Anticancer Effect
| Polysaccharide | Dosage Regimen and Species | Major Observations and Mechanism Proposed | Ref. |
|---|---|---|---|
| Fucoidan from F. evanescens | 1–50 mg/ kg, 3 times/week/ up to 21 days, Rats | Inhibition of lymphokine-activated killer T-cell-originated protein kinase (TOPK) (64% at 400 µg/mL) and EGF-downstream signaling. ↓ Tumor growth 72% at 50 mg/kg. | [24] |
| Fucoidan from F. vesiculosus | 150 mg/kg/body weight, 2 weeks, Athymic mice | Enhancement of the cytotoxic activity of splenic NK cells (~2.3 fold). | [41] |
| Fucoidan from F. vesiculosus | 100 mg/kg, 21 days starting on the seventh day pos tumor implantation, Mice | Induces G0/G1 cell cycle arrest (2–10%) and caspase-dependent apoptosis. | [42] |
| Fucoidan from F. vesiculosus | 144 mg/kg, 26 days, Mice | Reduction of Transforming Growth Factor Receptor (TGFR) levels (↓ ~50%) and its downstream signaling pathways. Enhancement of TGFR degradation. | [43] |
| Fucoidan from F. vesiculosus | 20 mg/kg, 28 days, Athymic mice | Inhibition of angiogenesis by decreasing mRNA expression level of angiogenesis related markers (↓ ~70%) and gene promoters. | [44] |
4. Immunomodulatory Effect
| Immunomodulatory Effect | Polysaccharide | Dosage Regimen and Species | Major Observations and Mechanism Proposed | Ref. |
|---|---|---|---|---|
| Antifibrotic effect | Fucoidan from C. okamuranus | Free access to drinking water containing 2% low (28.8 kDa) or high (41.4 kDa) MW fractions, 12 weeks, Rats | ↓ TGF-β1 mRNA expression and the levels of chemokine ligand CXCL12 in the liver (~3 fold). | [26] |
| Hepatoprotection | Fucoidan from F. vesiculosus | 30 or 60 mg/kg, 7 days, Mice | ↓ expression of liver TGF-β1(~40%) and COX-2, ↑ antioxidant pathways. | [45] |
| Hepatoprotection | Fucoidan from F. vesiculosus | 100 mg/kg, 4 weeks, Rats on high-fat diet | ↓TNF-α, IL-1β and MMP-2 mRNA expressions (~50–70%). Prevention of the increase in serum lipids and glucose levels induced by HFD. | [46] |
| Nephroprotection | Fucoidan from F. vesiculosus | 50 and 75 mg/kg, 13 weeks, Rats | Decreased levels of collagen IV, NF-κB, TGF-β1 and fibronectin in the renal cortex and in the glomerular mesangial cells. | [47] |
| Anti-arthritic and antioxidant effects | Fucoidan from U. pinnatifida | 50 or 150 mg/kg, 25 days, Rats | Downregulation of COX-2 and other inflammatory mediators (68% inhibition of in vivo inflammation). | [27] |
| Immunostimulatory effects | Fucoidan from U. pinnatifida | 300 mg daily,20 weeks, Human | Higher immunogenicity of influenza trivalent vaccine than control group and increase of natural killer cell activity. | [50] |
| Suppression of allergic symptoms | Fucoidan from S. japonica | 100–400 μg/day,4 days, Rats | Prevention of the interaction of IgE and mast cells via an increase in galectin-9 mRNA expression (↑ ~50%) in intestinal epithelial cells. | [28] |
| Anti-inflammatory effect | FucCS from I. badionotus | 80 m/kg, 7 days, Rats | Downregulation of NF-kB and downstream genes such as COX-2 and TNF-α and a benefic effect on gut microbiota. | [49] |
| Anti-inflammatory effect | Sulfated fucan from A. molpadioides with varying degrees of polymerization (10–500 kDa) | 50 mg/kg, 26 days, Mice | Regulation of IFN-γ/IL-4 ratio (0.53 to 0.70) and Th1/Th2 response, IL-6 and IL-10 levels, enhanced IgA protein expression levels (~35%) in intestinal mucosa. | [48] |
5. Effects on Diabetes
| Polysaccharide | Dosage Regimen and Species | Major Observations and Mechanism Proposed | Ref. |
|---|---|---|---|
| Fucoidan from F. vesiculosus | 75 m/kg, 13 weeks, Rats | Reduced islet atrophy, fibrosis and inflammation mediated by cAMP signaling pathway. Inhibition of dipeptidyl peptidase-IV. | [25,52] |
| High molecular weight fucoidan from Mozuku (C. okamuranus) | 1620 mg, 12 weeks, Human | Alterations in GLP-1 (from 6.42 ± 3.52 to 4.93 ± 1.88 pmol/L) and hemoglobin A1c levels (from 6.73 ± 1.00 to 6.59 ± 1.00). | [29] |
| Fucoidan extract from Laminaria ssp. | 500 mg, 3 months, Human | Decrease in diastolic blood pressure and LDL-C (↓13%) with increase in insulin levels (↑ 30%), HOMA β-cell, and HOMA IR. | [29,55] |
| FucCS from C. frondosa | 20 or 80 mg/kg, 19 weeks, Mice | ↑insulin-stimulated phosphorylation of PI3K and PKB; ↑GLUT4 translocation ↑ glycogen synthesis-related gene expression; ↓ liver ER stress markers, ROS, TNF-α and other inflammatory markers levels in serum and liver; ↓ inflammatory signaling pathways in the liver. | [33,53,54] |
| Sulfated polysaccharides from 10 sea cucumber species | 200 or 400 mg/kg, 8 weeks, Rats | ↓TNF-α, ↑ antioxidant enzymes; ↑ glucose metabolism related gene signaling pathway. | [32] |
6. Thrombosis and Hemostasis
| Polysaccharide | Dosage Regimen and Species | Major Observations and Mechanism Proposed | Ref. |
|---|---|---|---|
| Low molecular weight fucoidan (Mw7.6 kDa) from L. japonica | 400 and 800 mg/kg 30 days, Rats | ↑ TFPI (4.5 to 110.2 U/mL) and 6-keto-PGF1α levels (32.8 to 50.4 U/mL). ↑ Fibrinolysis (tPA and PAI-1 levels) ↓Thromboxane A2 levels. | [68] |
| Fucoidan from L. japonica | 400 mg for 5 weeks to humans | ↑ 6-keto-PGF1a (44 to 113 ng/L) ↑ fibrinolysis. | [30] |
| Fucoidans from F. vesiculosus and L. japonica | 5–20 mg/kg, Twice daily in a multiweek escalation dose, Dogs | Procoagulant effect, Inhibition of TFPI activity. | [71] |
| Native and gastro-resistant tablets of fucCS from L. grisea | 5–50 mg/kg, Single dose or 5 days, Rats | Serpin-independent anticoagulant effect by inhibiting the formation of factor Xa and/or IIa through the procoagulants tenase and prothrombinase complexes. Antithrombotic effects at 50 mg/kg: ~85% vs. 55% inhibition of the venous and arterial thrombus weight, respectively. | [20,66,67] |
| Gastro-resistant tablets containing FucCS oligomers (6 to 18 saccharide units, Mw 3,4 kDa) from P. graeffei | 10 or 50 mg/kg Single dose, Rats | Anticoagulant and antithrombotic effects (82% of venous thrombosis inhibition at 50 mg/kg). | [70] |
7. Clinical Trials
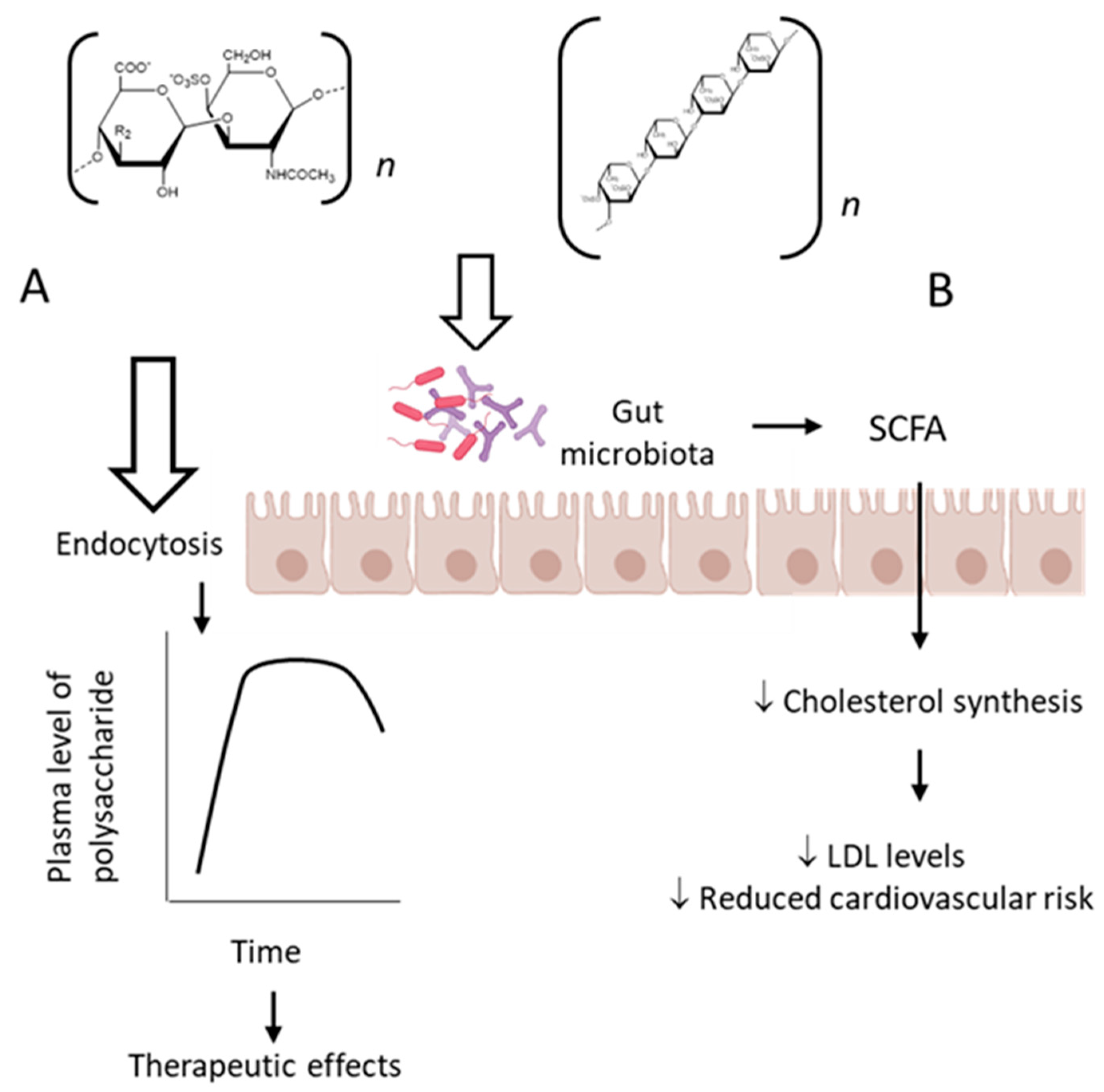
8. Future Perspectives: Pharmacokinetics Studies and Prebiotic Effects
9. Conclusions
Funding
Conflicts of Interest
References
- Springer, G.F.; Wurzel, H.A.; Mcneal, G.M.; Ansell, N.J.; Doughty, M.F. Isolation of Anticoagulant Fractions from Crude Fucoidin. Proc. Soc. Exp. Biol. Med. 1957, 94, 404–409. [Google Scholar] [CrossRef]
- Pomin, V.H. NMR Structural Determination of Unique Invertebrate Glycosaminoglycans Endowed with Medical Properties. Carbohydr. Res. 2015, 413, 41–50. [Google Scholar] [CrossRef]
- Soares, P.A.G.; Queiroz, I.N.L.; Pomin, V.H. NMR Structural Biology of Sulfated Glycans. J. Biomol. Struct. Dyn. 2017, 35, 1069–1084. [Google Scholar] [CrossRef]
- Fitton, J.H.; Stringer, D.N.; Karpiniec, S.S. Therapies from Fucoidan: An Update. Mar. Drugs 2015, 13, 5920–5946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mourão, P.A.S.; Bastos, I.G. Highly Acidic Glycans from Sea Cucumbers: Isolation and Fractionation of Fucose-rich Sulfated Polysaccharides from the Body Wall of Ludwigothurea Grisea. Eur. J. Biochem. 1987, 166, 639–645. [Google Scholar] [CrossRef]
- Mourão, P.A.S.; Perlin, A.S. Structural Features of Sulfated Glycans from the Tunic of Styela Plicata (Chordata-Tunicata): A Unique Occurrence of L-galactose in Sulfated Polysaccharides. Eur. J. Biochem. 1987, 166, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.A.; Mulloy, B.; Mourão, P.A.S. Structural Diversity among Sulfated A-L-galactans from Ascidians (Tunicates): Studies on the Species Ciona Intestinalis and Herdmania Monus. Eur. J. Biochem. 1992, 204, 669–677. [Google Scholar] [CrossRef]
- Pereira, M.S.; Mulloy, B.; Mourão, P.A.S. Structure and Anticoagulant Activity of Sulfated Fucans. Comparison between the Regular, Repetitive, and Linear Fucans from Echinoderms with the More Heterogeneous and Branched Polymers from Brown Algae. J. Biol. Chem. 1999, 274, 7656–7667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilela-Silva, A.C.E.S.; Alves, A.P.; Valente, A.P.; Vacquier, V.D.; Mourão, P.A.S. Structure of the Sulfated α-L-Fucan from the Egg Jelly Coat of the Sea Urchin Strongylocentrotus Franciscanus: Patterns of Preferential 2-O- and 4-O-Sulfation Determine Sperm Cell Recognition. Glycobiology 1999, 9, 927–933. [Google Scholar] [CrossRef] [Green Version]
- Alves, A.P.; Mulloy, B.; Diniz, J.A.; Mourão, P.A.S. Sulfated Polysaccharides from the Egg Jelly Layer Are Species-Specific Inducers of Acrosomal Reaction in Sperms of Sea Urchins. J. Biol. Chem. 1997, 272, 6965–6971. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, A.C.; Vieira, R.P.; Mourão, P.A.S.; Mulloy, B. A Sulfated α-l-Fucan from Sea Cucumber. Carbohydr. Res. 1994, 255, 225–240. [Google Scholar] [CrossRef]
- Pacheco, R.G.; Vicente, C.P.; Zancan, P.; Mourão, P.A.S. Different Antithrombotic Mechanisms among Glycosaminoglycans Revealed with a New Fucosylated Chondroitin Sulfate from an Echinoderm. Blood Coagul. Fibrinolysis 2000, 11, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Jiao, G.; Yu, G.; Zhang, J.; Ewart, H.S. Chemical Structures and Bioactivities of Sulfated Polysaccharides from Marine Algae. Mar. Drugs 2011, 9, 196–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, M.S.; Melo, F.R.; Mourão, P.A.S. Is There a Correlation between Structure and Anticoagulant Action of Sulfated Galactans and Sulfated Fucans? Glycobiology 2002, 12, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Mourão, P.A.S. Perspective on the Use of Sulfated Polysaccharides from Marine Organisms as a Source of New Antithrombotic Drugs. Mar. Drugs 2015, 13, 2770–2784. [Google Scholar] [CrossRef] [Green Version]
- Pomin, V.H. Holothurian Fucosylated Chondroitin Sulfate. Mar. Drugs 2014, 12, 232–254. [Google Scholar] [CrossRef] [Green Version]
- Borsig, L.; Wang, L.; Cavalcante, M.C.M.; Cardilo-Reis, L.; Ferreira, P.L.; Mourão, P.A.S.; Esko, J.D.; Pavão, M.S.G. Selectin Blocking Activity of a Fucosylated Chondroitin Sulfate Glycosaminoglycan from Sea Cucumber: Effect on Tumor Metastasis and Neutrophil Recruitment. J. Biol. Chem. 2007, 282, 14984–14991. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.E.; Kim, H.; Seo, C.; Park, T.; Lee, K.B.; Yoo, S.Y.; Hong, S.C.; Kim, J.T.; Lee, J. Marine Polysaccharides: Therapeutic Efficacy and Biomedical Applications. Arch. Pharmacal Res. 2017, 40, 1006–1020. [Google Scholar] [CrossRef]
- Wang, Y.; Xing, M.; Cao, Q.; Ji, A.; Liang, H.; Song, S. Biological Activities of Fucoidan and the Factors Mediating Its Therapeutic Effects: A Review of Recent Studies. Mar. Drugs 2019, 17, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca, R.J.C.; Mourão, P.A.S. Fucosylated Chondroitin Sulfate as a New Oral Antithrombotic Agent. Thromb. Haemost. 2006, 96, 822–829. [Google Scholar] [CrossRef]
- Richards, C.; Williams, N.A.; Fitton, J.H.; Stringer, D.N.; Karpiniec, S.S.; Park, A.Y. Oral Fucoidan Attenuates Lung Pathology and Clinical Signs in a Severe Influenza a Mouse Model. Mar. Drugs 2020, 18, 246. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Yin, J.; Wang, Y.; Wang, J.; Xia, B.; Li, T.; Yang, X.; Hu, S.; Ji, C.; Guo, S. The Fucoidan A3 from the Seaweed Ascophyllum Nodosum Enhances RCT-Related Genes Expression in Hyperlipidemic C57BL/6J Mice. Int. J. Biol. Macromol. 2019, 134, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Cuong, H.D.; Thuy, T.T.T.; Huong, T.T.; Ly, B.M.; Van, T.T.T. Structure and Hypolipidaemic Activity of Fucoidan Extracted from Brown Seaweed Sargassum Henslowianum. Nat. Prod. Res. 2015, 29, 411–415. [Google Scholar] [CrossRef]
- Vishchuk, O.S.; Sun, H.; Wang, Z.; Ermakova, S.P.; Xiao, J.J.; Lu, T.; Xue, P.P.; Zvyagintseva, T.N.; Xiong, H.; Shao, C.; et al. PDZ-Binding Kinase/T-LAK Cell-Originated Protein Kinase Is a Target of the Fucoidan from Brown Alga Fucus Evanescens in the Prevention of EGF-Induced Neoplastic Cell Transformation and Colon Cancer Growth. Oncotarget 2016, 7, 18763–18773. [Google Scholar] [CrossRef] [Green Version]
- Pozharitskaya, O.N.; Obluchinskaya, E.D.; Shikov, A.N. Mechanisms of Bioactivities of Fucoidan from the Brown Seaweed Fucus Vesiculosus L. Of the Barents Sea. Mar. Drugs 2020, 18, 275. [Google Scholar] [CrossRef]
- Nakazato, K.; Takada, H.; Iha, M.; Nagamine, T. Attenuation of N-Nitrosodiethylamine-Induced Liver Fibrosis by High-Molecular-Weight Fucoidan Derived from Cladosiphon Okamuranus. J. Gastroenterol. Hepatol. 2010, 25, 1692–1701. [Google Scholar] [CrossRef]
- Phull, A.R.; Majid, M.; Haq, I.U.; Khan, M.R.; Kim, S.J. In Vitro and in Vivo Evaluation of Anti-Arthritic, Antioxidant Efficacy of Fucoidan from Undaria Pinnatifida (Harvey) Suringar. Int. J. Biol. Macromol. 2017, 97, 468–480. [Google Scholar] [CrossRef]
- Tanino, Y.; Hashimoto, T.; Ojima, T.; Mizuno, M. F-Fucoidan from Saccharina Japonica Is a Novel Inducer of Galectin-9 and Exhibits Anti-Allergic Activity. J. Clin. Biochem. Nutr. 2016, 59, 25–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakai, C.; Abe, S.; Kouzuki, M.; Shimohiro, H.; Ota, Y.; Sakinada, H.; Takeuchi, T.; Okura, T.; Kasagi, T.; Hanaki, K. A Randomized Placebo-Controlled Trial of an Oral Preparation of High Molecular Weight Fucoidan in Patients with Type 2 Diabetes with Evaluation of Taste Sensitivity. Yonago Acta Med. 2019, 62, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Ren, R.; Azuma, Y.; Ojima, T.; Hashimoto, T.; Mizuno, M.; Nishitani, Y.; Yoshida, M.; Azuma, T.; Kanazawa, K. Modulation of Platelet Aggregation-Related Eicosanoid Production by Dietary F-Fucoidan from Brown Alga Laminaria Japonica in Human Subjects. Br. J. Nutr. 2013, 110, 880–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, C.Y.; Liao, N.B.; Zhang, Y.; Ye, X.Q.; Li, S.; Hu, Y.Q.; Liu, D.H.; Linhardt, R.J.; Wang, X.; Chen, S.G. In Vitro Fermentation Behaviors of Fucosylated Chondroitin Sulfate from Pearsonothuria Graeffei by Human Gut Microflora. Int. J. Biol. Macromol. 2017, 102, 1195–1201. [Google Scholar] [CrossRef]
- Zhu, Q.; Lin, L.; Zhao, M. Sulfated Fucan/Fucosylated Chondroitin Sulfate-Dominated Polysaccharide Fraction from Low-Edible-Value Sea Cucumber Ameliorates Type 2 Diabetes in Rats: New Prospects for Sea Cucumber Polysaccharide Based-Hypoglycemic Functional Food. Int. J. Biol. Macromol. 2020, 159, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Jiang, W.; Hu, S.; Song, W.; Ji, L.; Wang, Y.; Cai, L. Fucosylated Chondroitin Sulphate from Cusumaria Frondosa Mitigates Hepatic Endoplasmic Reticulum Stress and Inflammation in Insulin Resistant Mice. Food Funct. 2015, 6, 1547–1556. [Google Scholar] [CrossRef] [PubMed]
- Soppert, J.; Lehrke, M.; Marx, N.; Jankowski, J.; Noels, H. Lipoproteins and Lipids in Cardiovascular Disease: From Mechanistic Insights to Therapeutic Targeting. Adv. Drug Deliv. Rev. 2020, 159, 4–33. [Google Scholar] [CrossRef]
- Penson, P.E.; Banach, M. The Role of Nutraceuticals in the Optimization of Lipid-Lowering Therapy in High-Risk Patients with Dyslipidaemia. Curr. Atheroscler. Rep. 2020, 22, 1–9. [Google Scholar] [CrossRef]
- Liu, H.H.; Ko, W.C.; Hu, M.L. Hypolipidemic Effect of Glycosaminoglycans from the Sea Cucumber Metriatyla Scabra in Rats Fed a Cholesterol-Supplemented Diet. J. Agric. Food Chem. 2002, 50, 3602–3606. [Google Scholar] [CrossRef]
- Yin, J.; Wang, J.; Li, F.; Yang, Z.; Yang, X.; Sun, W.; Xia, B.; Li, T.; Song, W.; Guo, S. The Fucoidan from the Brown Seaweed: Ascophyllum Nodosum Ameliorates Atherosclerosis in Apolipoprotein E-Deficient Mice. Food Funct. 2019, 10, 5124–5139. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, J.; Zhi, Z.; Wei, C.; Wang, W.; Ding, T.; Ye, X.; Hu, Y.; Linhardt, R.J.; Chen, S. Macromolecular Properties and Hypolipidemic Effects of Four Sulfated Polysaccharides from Sea Cucumbers. Carbohydr. Polym. 2017, 173, 330–337. [Google Scholar] [CrossRef]
- Kumar, M.; Nagpal, R.; Hemalatha, R.; Verma, V.; Kumar, A.; Singh, S.; Marotta, F.; Jain, S.; Yadav, H. Targeted Cancer Therapies: The Future of Cancer Treatment. Acta Biomed. 2012, 83, 220–233. [Google Scholar] [PubMed]
- Rejhová, A.; Opattová, A.; Čumová, A.; Slíva, D.; Vodička, P. Natural Compounds and Combination Therapy in Colorectal Cancer Treatment. Eur. J. Med. Chem. 2018, 144, 582–594. [Google Scholar] [CrossRef]
- Atashrazm, F.; Lowenthal, R.M.; Woods, G.M.; Holloway, A.F.; Karpiniec, S.S.; Dickinson, J.L. Fucoidan Suppresses the Growth of Human Acute Promyelocytic Leukemia Cells In Vitro and In Vivo. J. Cell. Physiol. 2016, 231, 688–697. [Google Scholar] [CrossRef]
- Yang, G.; Zhang, Q.; Kong, Y.; Xie, B.; Gao, M.; Tao, Y.; Xu, H.; Zhan, F.; Dai, B.; Shi, J.; et al. Antitumor Activity of Fucoidan against Diffuse Large B Cell Lymphoma in Vitro and in Vivo. Acta Biochim. Sinica 2015, 47, 925–931. [Google Scholar] [CrossRef] [Green Version]
- Hsu, H.Y.; Lin, T.Y.; Wu, Y.C.; Tsao, S.M.; Hwang, P.A.; Shih, Y.W.; Hsu, J. Fucoidan Inhibition of Lung Cancer in Vivo and in Vitro: Role of the Smurf2-Dependent Ubiquitin Proteasome Pathway in TGFβ Receptor Degradation. Oncotarget 2014, 5, 7870–7885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rui, X.; Pan, H.F.; Shao, S.L.; Xu, X.M. Anti-Tumor and Anti-Angiogenic Effects of Fucoidan on Prostate Cancer: Possible JAK-STAT3 Pathway. BMC Complement. Altern. Med. 2017, 17, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.D.; Lee, S.R.; Kim, T.; Jang, S.A.; Kang, S.C.; Koo, H.J.; Sohn, E.; Bak, J.P.; Namkoong, S.; Kim, H.K.; et al. Fucoidan from Fucus Vesiculosus Protects against Alcohol-Induced Liver Damage by Modulating Inflammatory Mediators in Mice and Hepg2 Cells. Mar. Drugs 2015, 13, 1051–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heeba, G.H.; Morsy, M.A. Fucoidan Ameliorates Steatohepatitis and Insulin Resistance by Suppressing Oxidative Stress and Inflammatory Cytokines in Experimental Non-Alcoholic Fatty Liver Disease. Environ. Toxicol. Pharmacol. 2015, 40, 907–914. [Google Scholar] [CrossRef]
- Wang, Y.; Nie, M.; Lu, Y.; Wang, R.; Li, J.; Yang, B.; Xia, M.; Zhang, H.; Li, X. Fucoidan Exerts Protective Effects against Diabetic Nephropathy Related to Spontaneous Diabetes through the NF-ΚB Signaling Pathway in Vivo and in Vitro. Int. J. Mol. Med. 2015, 35, 1067–1073. [Google Scholar] [CrossRef] [Green Version]
- Zuo, T.; Li, X.; Chang, Y.; Duan, G.; Yu, L.; Zheng, R.; Xue, C.; Tang, Q. Dietary Fucoidan of Acaudina Molpadioides and Its Enzymatically Degraded Fragments Could Prevent Intestinal Mucositis Induced by Chemotherapy in Mice. Food Funct. 2015, 6, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Olivera-Castillo, L.; Grant, G.; Kantún-Moreno, N.; Barrera-Pérez, H.A.; Montero, J.; Olvera-Novoa, M.A.; Carrillo-Cocom, L.M.; Acevedo, J.J.; Puerto-Castillo, C.; Solís, V.M.; et al. A Glycosaminoglycan-Rich Fraction from Sea Cucumber Isostichopus Badionotus Has Potent Anti-Inflammatory Properties in Vitro and in Vivo. Nutrients 2020, 12, 1698. [Google Scholar] [CrossRef] [PubMed]
- Negishi, H.; Mori, M.; Mori, H.; Yamori, Y. Supplementation of Elderly Japanese Men and Women with Fucoidan from Seaweed Increases Immune Responses to Seasonal Influenza Vaccination. J. Nutr. 2013, 143, 1794–1798. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Sultan, S. Oral Hypoglycemic Medications; StatPearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Jiang, X.; Yu, J.; Ma, Z.; Zhang, H.; Xie, F. Effects of Fucoidan on Insulin Stimulation and Pancreatic Protection via the CAMP Signaling Pathway in Vivo and in Vitro. Mol. Med. Rep. 2015, 12, 4501–4507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, S.; Chang, Y.; He, M.; Wang, J.; Wang, Y.; Xue, C. Fucosylated Chondroitin Sulfate from Sea Cucumber Improves Insulin Sensitivity via Activation of PI3K/PKB Pathway. J. Food Sci. 2014, 79, H1424–H1429. [Google Scholar] [CrossRef]
- Hu, S.; Chang, Y.; Wang, J.; Xue, C.; Li, Z.; Wang, Y. Fucosylated Chondroitin Sulfate from Sea Cucumber in Combination with Rosiglitazone Improved Glucose Metabolism in the Liver of the Insulin-Resistant Mice. Biosci. Biotechnol. Biochem. 2013, 77, 2263–2268. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Corona, D.M.; Martínez-Abundis, E.; González-Ortiz, M. Effect of Fucoidan Administration on Insulin Secretion and Insulin Resistance in Overweight or Obese Adults. J. Med. Food 2014, 17, 830–832. [Google Scholar] [CrossRef] [PubMed]
- Mulloy, B.; Hogwood, J.; Gray, E.; Lever, R.; Page, C.P. Pharmacology of Heparin and Related Drugs. Pharmacol. Rev. 2015, 68, 76–141. [Google Scholar] [CrossRef]
- Bleeding Associated with Antithrombotic—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/6160618/ (accessed on 13 June 2021).
- Heparin-Induced Thrombocytopenia: Molecular Pathogenesis—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/10605736/ (accessed on 13 June 2021).
- Nutescu, E.A.; Dager, W.E.; Kalus, J.S.; Lewin, J.J.; Cipolle, M.D. Management of Bleeding and Reversal Strategies for Oral Anticoagulants: Clinical Practice Considerations. Am. J. Health-Syst. Pharm. 2013, 70, 1914–1929. [Google Scholar] [CrossRef] [PubMed]
- Mourão, P.A.S.; Pereira, M.S. Searching for Alternatives to Heparin: Sulfated Fucans from Marine Invertebrates. Trends Cardiovasc. Med. 1999, 9, 225–232. [Google Scholar] [CrossRef]
- Mourao, P. Use of Sulfated Fucans as Anticoagulant and Antithrombotic Agents: Future Perspectives. Curr. Pharm. Des. 2005, 10, 967–981. [Google Scholar] [CrossRef]
- Mourao, P.A.S.; Pereira, M.S.; Pavo, M.S.G.; Mulloy, B.; Tollefsen, D.M.; Mowinckel, M.C.; Abildgaard, U. Structure and Anticoagulant Activity of a Fucosylated Chondroitin Sulfate from Echinoderm. Sulfated Fucose Branches on the Polysaccharide Account for Its High Anticoagulant Action. J. Biol. Chem. 1996, 271, 23973–23984. [Google Scholar] [CrossRef] [Green Version]
- Thorlacius, H.; Vollmar, B.; Seyfert, U.T.; Vestweber, D.; Menger, M. The Polysaccharide Fucoidan Inhibits Microvascular Thrombus Formation Independently from P- and L-Selectin Function in Vivo. Eur. J. Clin. Investig. 2000, 30, 804–810. [Google Scholar] [CrossRef]
- Zancan, P.; Mourão, P.A. Venous and Arterial Thrombosis in Rat Models. Blood Coagul. Fibrinolysis 2004, 15, 45–54. [Google Scholar] [CrossRef]
- Fonseca, R.J.C.; Santos, G.R.C.; Mourão, P.A.S. Effects of Polysaccharides Enriched in 2,4-Disulfated Fucose Units on Coagulation, Thrombosis and Bleeding: Practical and Conceptual Implications. Thromb. Haemost. 2009, 102, 829–836. [Google Scholar] [CrossRef]
- Glauser, B.F.; Pereira, M.S.; Monteiro, R.Q.; Mourão, P.A.S. Serpin-Independent Anticoagulant Activity of a Fucosylated Chondroitin Sulfate. Thromb. Haemost. 2008, 100, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, R.J.C.; Sucupira, I.D.; Oliveira, S.N.M.C.G.; Santos, G.R.C.; Mourão, P.A.S. Improved Anticoagulant Effect of Fucosylated Chondroitin Sulfate Orally Administered as Gastro-Resistant Tablets. Thromb. Haemost. 2017, 117, 662–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Guo, F.; Hu, J.; Zhang, L.; Xue, C.; Zhang, Z.; Li, B. Antithrombotic Activity of Oral Administered Low Molecular Weight Fucoidan from Laminaria Japonica. Thromb. Res. 2016, 144, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, R.J.C.; Oliveira, S.N.M.C.G.; Pomin, V.H.; Mecawi, A.S.; Araujo, I.G.; Mourão, P.A.S. Effects of Oversulfated and Fucosylated Chondroitin Sulfates on Coagulation: Challenges for the Study of Anticoagulant Polysaccharides. Thromb. Haemost. 2010, 103, 994–1004. [Google Scholar] [CrossRef]
- Yan, L.; Zhu, M.; Wang, D.; Tao, W.; Liu, D.; Zhang, F.; Linhardt, R.J.; Ye, X.; Chen, S. Oral Administration of Fucosylated Chondroitin Sulfate Oligomers in Gastro-Resistant Microcapsules Exhibits a Safe Antithrombotic Activity. Thromb. Haemost. 2021, 121, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Lillicrap, D.; Labelle, A.; Knappe, S.; Keller, T.; Burnett, E.; Powell, S.; Johnson, K.W. Efficacy and Safety of a New-Class Hemostatic Drug Candidate, AV513, in Dogs with Hemophilia A. Blood 2008, 111, 672–679. [Google Scholar] [CrossRef]
- Improved Coagulation in Bleeding Disorders by Non-Anticoagulant Sulfated Polysaccharides (NASP)—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/16543964/ (accessed on 13 June 2021).
- Fonseca, R.J.C.; Oliveira, S.N.M.C.G.; Melo, F.R.; Pereira, M.G.; Benevides, N.M.B.; Mourão, P.A.S. Slight Differences in Sulfation of Algal Galactans Account for Differences in Their Anticoagulant and Venous Antithrombotic Activities. Thromb. Haemost. 2008, 99, 539–545. [Google Scholar] [CrossRef]
- Takahashi, H.; Kawaguchi, M.; Kitamura, K.; Narumiya, S.; Kawamura, M.; Tengan, I.; Nishimoto, S.; Hanamure, Y.; Majima, Y.; Tsubura, S.; et al. An Exploratory Study on the Anti-Inflammatory Effects of Fucoidan in Relation to Quality of Life in Advanced Cancer Patients. Integr. Cancer Ther. 2018, 17, 282–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagamine, T.; Kadena, K.; Tomori, M.; Nakajima, K.; Iha, M. Activation of NK Cells in Male Cancer Survivors by Fucoidan Extracted from Cladosiphon Okamuranus. Mol. Clin. Oncol. 2020, 12, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Hsu, H.; Hwang, P. Clinical Applications of Fucoidan in Translational Medicine for Adjuvant Cancer Therapy. Clin. Transl. Med. 2019, 8, 15. [Google Scholar] [CrossRef] [Green Version]
- Ikeguchi, M.; Yamamoto, M.; Arai, Y.; Maeta, Y.; Ashida, K.; Katano, K.; Miki, Y.; Kimura, T. Fucoidan Reduces the Toxicities of Chemotherapy for Patients with Unresectable Advanced or Recurrent Colorectal Cancer. Oncol. Lett. 2011, 2, 319–322. [Google Scholar] [CrossRef] [PubMed]
- Myers, S.P.; Mulder, A.M.; Baker, D.G.; Robinson, S.R.; Rolfe, M.I.; Brooks, L.; Fitton, J.H. Effects of Fucoidan from Fucus Vesiculosus in Reducing Symptoms of Osteoarthritis: A Randomized Placebo-Controlled Trial. Biol. Targets Ther. 2016, 10, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Imanari, T.; Washio, Y.; Huang, Y.; Toyoda, H.; Suzuki, A.; Toida, T. Oral Absorption and Clearance of Partially Depolymerized Fucosyl Chondroitin Sulfate from Sea Cucumber. Thromb. Res. 1999, 93, 129–135. [Google Scholar] [CrossRef]
- Irhimeh, M.R.; Fitton, J.H.; Lowenthal, R.M.; Kongtawelert, P. A Quantitative Method to Detect Fucoidan in Human Plasma Using a Novel Antibody. Methods Find. Exp. Clin. Pharmacol. 2005, 27, 705–710. [Google Scholar] [CrossRef]
- Tokita, Y.; Hirayama, M.; Nakajima, K.; Tamaki, K.; Iha, M.; Nagamine, T. Detection of Fucoidan in Urine after Oral Intake of Traditional Japanese Seaweed, Okinawa Mozuku (Cladosiphon Okamuranus Tokida). J. Nutr. Sci. Vitaminol. 2017, 63, 419–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagamine, T.; Nakazato, K.; Tomioka, S.; Iha, M.; Nakajima, K. Intestinal Absorption of Fucoidan Extracted from the Brown Seaweed, Cladosiphon Okamuranus. Mar. Drugs 2015, 13, 48–64. [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Shikov, A.N.; Faustova, N.M.; Obluchinskaya, E.D.; Kosman, V.M.; Vuorela, H.; Makarov, V.G. Pharmacokinetic and Tissue Distribution of Fucoidan from Fucus Vesiculosus after Oral Administration to Rats. Mar. Drugs 2018, 16, 132. [Google Scholar] [CrossRef] [Green Version]
- Shikov, A.N.; Flisyuk, E.V.; Obluchinskaya, E.D.; Pozharitskaya, O.N. Pharmacokinetics of Marine-Derived Drugs. Mar. Drugs 2020, 18, 557. [Google Scholar] [CrossRef]
- Conte, A.; Volpi, N.; Palmieri, L.; Bahous, I.; Ronca, G. Biochemical and Pharmacokinetic Aspects of Oral Treatment with Chondroitin Sulfate. Arzneim. Forsch. Drug Res. 1995, 45, 918–925. [Google Scholar]
- Zhang, T.; Wu, S.; Ai, C.; Wen, C.; Liu, Z.; Wang, L.; Jiang, L.; Shen, P.; Zhang, G.; Song, S. Galactofucan from Laminaria Japonica Is Not Degraded by the Human Digestive System but Inhibits Pancreatic Lipase and Modifies the Intestinal Microbiota. Int. J. Biol. Macromol. 2021, 166, 611–620. [Google Scholar] [CrossRef]
- Mueller, M.; Ganesh, R.; Bonnes, S. Gut Health = Mental Health? The Impact of Diet and Dietary Supplements on Mood Disorders. Curr. Nutr. Rep. 2020, 9, 361–368. [Google Scholar] [CrossRef]
- Dronkers, T.M.G.; Ouwehand, A.C.; Rijkers, G.T. Global Analysis of Clinical Trials with Probiotics. Heliyon 2020, 6, e04467. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Chen, C.; Wen, T.; Zhao, Q. Probiotics in the Prevention of Antibiotic-Associated Diarrhea in Adults: A Meta-Analysis of Randomized Placebo-Controlled Trials. J. Clin. Gastroenterol. 2021, 55, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Fakhri, S.; Yarmohammadi, A.; Yarmohammadi, M.; Farzaei, M.H.; Echeverria, J. Marine Natural Products: Promising Candidates in the Modulation of Gut-Brain Axis towards Neuroprotection. Mar. Drugs 2021, 19, 165. [Google Scholar] [CrossRef] [PubMed]
- Toumazi, D.; Constantinou, C. A Fragile Balance: The Important Role of the Intestinal Microbiota in the Prevention and Management of Colorectal Cancer. Oncology 2020, 98, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Clavel, T.; Lagkouvardos, I.; Stecher, B. From Complex Gut Communities to Minimal Microbiomes via Cultivation. Curr. Opin. Microbiol. 2017, 38, 148–155. [Google Scholar] [CrossRef]
- Arnold, J.W.; Roach, J.; Azcarate-Peril, M.A. Emerging Technologies for Gut Microbiome Research. Trends Microbiol. 2016, 24, 887–901. [Google Scholar] [CrossRef] [Green Version]
- Wiciński, M.; Gębalski, J.; Gołębiewski, J.; Malinowski, B. Probiotics for the Treatment of Overweight and Obesity in Humans—A Review of Clinical Trials. Microorganisms 2020, 8, 1148. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Dong, X.; Yan, C.; Ai, C.; Zhou, D.; Yang, J.; Zhang, H.; Liu, X.; Song, S.; Xiao, H.; et al. Structural Features and Digestive Behavior of Fucosylated Chondroitin Sulfate from Sea Cucumbers Stichopus Japonicus. J. Agric. Food Chem. 2019, 67, 10534–10542. [Google Scholar] [CrossRef]
- Hehemann, J.H.; Correc, G.; Barbeyron, T.; Helbert, W.; Czjzek, M.; Michel, G. Transfer of Carbohydrate-Active Enzymes from Marine Bacteria to Japanese Gut Microbiota. Nature 2010, 464, 908–912. [Google Scholar] [CrossRef]
- Strain, C.R.; Collins, K.C.; Naughton, V.; McSorley, E.M.; Stanton, C.; Smyth, T.J.; Soler-Vila, A.; Rea, M.C.; Ross, P.R.; Cherry, P.; et al. Effects of a Polysaccharide-Rich Extract Derived from Irish-Sourced Laminaria Digitata on the Composition and Metabolic Activity of the Human Gut Microbiota Using an in Vitro Colonic Model. Eur. J. Nutr. 2020, 59, 309–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imbs, T.I.; Zvyagintseva, T.N.; Ermakova, S.P. Is the Transformation of Fucoidans in Human Body Possible? Int. J. Biol. Macromol. 2020, 142, 778–781. [Google Scholar] [CrossRef] [PubMed]
- Kadena, K.; Tomori, M.; Iha, M.; Nagamine, T. Absorption Study of Mozuku Fucoidan in Japanese Volunteers. Mar. Drugs 2018, 16, 254. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fonseca, R.J.C.; Mourão, P.A.S. Pharmacological Activities of Sulfated Fucose-Rich Polysaccharides after Oral Administration: Perspectives for the Development of New Carbohydrate-Based Drugs. Mar. Drugs 2021, 19, 425. https://doi.org/10.3390/md19080425
Fonseca RJC, Mourão PAS. Pharmacological Activities of Sulfated Fucose-Rich Polysaccharides after Oral Administration: Perspectives for the Development of New Carbohydrate-Based Drugs. Marine Drugs. 2021; 19(8):425. https://doi.org/10.3390/md19080425
Chicago/Turabian StyleFonseca, Roberto J. C., and Paulo A. S. Mourão. 2021. "Pharmacological Activities of Sulfated Fucose-Rich Polysaccharides after Oral Administration: Perspectives for the Development of New Carbohydrate-Based Drugs" Marine Drugs 19, no. 8: 425. https://doi.org/10.3390/md19080425