
Marine Natural Products from Tunicates and Their Associated Microbes




Abstract
:
1. Introduction
2. Ecological Importance of Tunicates
3. Database Search on Tunicate MNPs
4. Profile of MNPs from Tunicates and Associated Microbes
5. Antimicrobial Applications
6. Anticancer and Antitumor Applications
7. Antifouling and Anti-Deterrent Activities
8. Miscellaneous Applications
9. Issues in Extraction & Identification of Tunicate MNPs
10. Metabolic Origin of Some Tunicates and Their Predators
11. Utilization of Invasive Tunicates Resources
12. Research Gaps and Future Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shenkar, N.; Swalla, B.J. Global diversity of Ascidiacea. PLoS ONE 2011, 6, e20657. [Google Scholar] [CrossRef]
- Holland, L.Z. Tunicates. Curr. Biol. 2016, 26, R141–R156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasparini, F.; Ballarin, L. Reproduction in Tunicates. In Encyclopedia of Reproduction, 2nd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2018; Volume 6, pp. 546–553. [Google Scholar]
- Bone, Q.; Carré, C.; Chang, P. Tunicate feeding filters. J. Mar. Biol. Assoc. UK 2003, 83, 907–919. [Google Scholar] [CrossRef]
- Delsuc, F.; Philippe, H.; Tsagkogeorga, G.; Simion, P.; Tilak, M.K.; Turon, X.; López-Legentil, S.; Piette, J.; Lemaire, P.; Douzery, E.J.P. A phylogenomic framework and timescale for comparative studies of tunicates. BMC Biol. 2018, 16, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berna, L.; Alvarez-Valin, F. Evolutionary genomics of fast evolving tunicates. Genome Biol. Evol. 2014, 6, 1724–1738. [Google Scholar] [CrossRef] [Green Version]
- Swalla, B.J. Building divergent body plans with similar genetic pathways. Heredity 2006, 97, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, W.R. Tunicates: Models for Chordate Evolution and Development at Low Genomic Complexity. In Comparative Genomics; Clark, M.S., Ed.; Springer Science + Business Media: New York, NY, USA, 2000; pp. 43–69. [Google Scholar]
- Lemaire, P. Evolutionary crossroads in developmental biology: The tunicates. Development 2011, 138, 2143–2152. [Google Scholar] [CrossRef] [Green Version]
- Zhan, A.; Briski, E.; Bock, D.G.; Ghabooli, S.; MacIsaac, H.J. Ascidians as models for studying invasion success. Mar. Biol. 2015, 162, 2449–2470. [Google Scholar] [CrossRef]
- Watters, D.J. Ascidian toxins with potential for drug development. Mar. Drugs 2018, 16, 162. [Google Scholar] [CrossRef] [Green Version]
- Gouiffes, D.; Juge, M.; Grimaud, N.; Welin, L.; Sauviat, M.P.; Barbin, Y.; Laurent, D.; Roussakis, C.; Henichart, J.P.; Verbist, J.F. Bistramide A, a new toxin from the urochordata Lissoclinum bistratum Sluiter: Isolation and preliminary characterization. Toxicon 1988, 26, 1129–1136. [Google Scholar] [CrossRef]
- Oh, K.-S.; Kim, J.-S.; Heu, M.-S. Food Constituents of Edible Ascidians Halocynthia roretzi and Pyura michaelseni. Korean J. Food Sci. Technol. 1997, 29, 955–962. [Google Scholar]
- Ali, A.J.H.; Tamilselvi, M. Ascidians in Coastal Water: A Comprehensive Inventory of Ascidian Fauna from the Indian Coast; Springer Nature: Cham, Switzerland, 2016; ISBN 9783319291185. [Google Scholar]
- DeFilippo, J.; Beck, G. Tunicate Immunology. In Reference Module in Life Sciences; Elsevier: Amsterdam, The Netherlands, 2018; pp. 1–10. [Google Scholar]
- Ayuningrum, D.; Liu, Y.; Riyanti; Sibero, M.T.; Kristiana, R.; Asagabaldan, M.A.; Wuisan, Z.G.; Trianto, A.; Radjasa, O.K.; Sabdono, A.; et al. Tunicate-associated bacteria show a great potential for the discovery of antimicrobial compounds. PLoS ONE 2019, 14, e0213797. [Google Scholar] [CrossRef]
- Franchi, N.; Ballarin, L. Immunity in protochordates: The tunicate perspective. Front. Immunol. 2017, 8, 674. [Google Scholar] [CrossRef] [PubMed]
- Walters, T.L.; Gibson, D.M.; Frischer, M.E. Cultivation of the marine pelagic tunicate Dlioletta gegenbauri (Uljanin 1884) for experimental studies. J. Vis. Exp. 2019, 150, e59832. [Google Scholar] [CrossRef]
- Fusetani, N. Drugs from the Sea; Krager: Basel, Switzerland, 2000. [Google Scholar]
- Michibara, H.; Uyama, T.; Ueki, T.; Kanamori, K. The mechanism of accumulation and reduction of vanadium by ascidians. In The Biology of Ascidians; Sawada, H., Yokosawa, H., Lambert, C.C., Eds.; Springer: Tokyo, Japan, 2001; pp. 363–373. [Google Scholar]
- Shen, G.Q.; Baker, B.J. Biosynthetic studies of the eudistomins in the tunicate Eudistoma olivaceum. Tetrahedron Lett. 1994, 35, 1141–1144. [Google Scholar] [CrossRef]
- Le, V.H.; Inai, M.; Williams, R.M.; Kan, T. Ecteinascidins. A review of the chemistry, biology and clinical utility of potent tetrahydroisoquinoline antitumor antibiotics. Nat. Prod. Rep. 2015, 32, 328–347. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Kersten, R.D.; Nam, S.J.; Lu, L.; Al-Suwailem, A.M.; Zheng, H.; Fenical, W.; Dorrestein, P.C.; Moore, B.S.; Qian, P.Y. Bacterial biosynthesis and maturation of the didemnin anti-cancer agents. J. Am. Chem. Soc. 2012, 134, 8625–8632. [Google Scholar] [CrossRef] [Green Version]
- Dewapriya, P.; Khalil, Z.G.; Prasad, P.; Salim, A.A.; Cruz-Morales, P.; Marcellin, E.; Capon, R.J. Talaropeptides A-D: Structure and biosynthesis of extensively N-methylated linear peptides from an Australian marine tunicate-derived Talaromyces sp. Front. Chem. 2018, 6, 394. [Google Scholar] [CrossRef]
- Dou, X.; Dong, B. Origins and bioactivities of natural compounds derived from marine ascidians and their symbionts. Mar. Drugs 2019, 17, 670. [Google Scholar] [CrossRef] [Green Version]
- Casertano, M.; Menna, M.; Imperatore, C. The ascidian-derived metabolites with antimicrobial properties. Antibiotics 2020, 9, 510. [Google Scholar] [CrossRef]
- Palanisamy, S.K.; Rajendran, N.M.; Marino, A. Natural products diversity of marine ascidians (Tunicates; Ascidiacea) and successful drugs in clinical development. Nat. Prod. Bioprospect. 2017, 7, 1–111. [Google Scholar] [CrossRef] [Green Version]
- Petersen, J.K.; Riisgard, H.U. Filtration capacity of the ascidian Ciona intestinalis and its grazing impact in a shallow fjord. Mar. Ecol. Prog. Ser. 1992, 88, 9–17. [Google Scholar] [CrossRef]
- Morris, R.J.; Bone, Q.; Head, R.; Braconnot, J.C.; Nival, P. Role of salps in the flux of organic matter to the bottom of the Ligurian Sea. Mar. Biol. 1988, 97, 237–241. [Google Scholar] [CrossRef]
- Pomeroy, L.R.; Deibel, D. Aggregation of organic matter by pelagic tunicates. Limnol. Oceanogr. 1980, 25, 643–652. [Google Scholar] [CrossRef]
- Gorsky, G.; Da Silva, N.L.; Dallot, S.; Laval, P.; Braconnot, J.C.; Prieur, L. Midwater tunicates: Are they related to the permanent front of the Ligurian Sea (NW Mediterranean)? Mar. Ecol. Prog. Ser. 1991, 74, 195–204. [Google Scholar] [CrossRef]
- Sutherland, K.R.; Madin, L.P.; Stocker, R. Filtration of submicrometer particles by pelagic tunicates. Proc. Natl. Acad. Sci. USA 2010, 107, 15129–15134. [Google Scholar] [CrossRef] [Green Version]
- Hirose, E.; Nozawa, Y. Latitudinal difference in the species richness of photosymbiotic ascidians along the east coast of Taiwan. Zool. Stud. 2020, 59, e19. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.R. Alkaloids and ascidian chemical defense: Evidence for the ecological role of natural products from Eudistoma olivaceum. Mar. Biol. 1991, 111, 375–379. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, J. Excellent chemical and material cellulose from tunicates: Diversity in cellulose production yield and chemical and morphological structures from different tunicate species. Cellulose 2014, 21, 3427–3441. [Google Scholar] [CrossRef]
- Menna, M.; Aiello, A. The Chemistry of Marine Tunicates. In Handbook of Marine Natural Products; Fattorusso, E., Gerwick, W.H., Taglialatela-Scafati, O., Eds.; Springer Science + Business Media B.V.: Berlin/Heidelberg, Germany, 2012; pp. 295–385. [Google Scholar]
- Franchi, N.; Ballarin, L. Cytotoxic cells of compound Ascidians. In Lessons in Immunity: From Single-Cell Organisms to Mammals; Ballarin, L., Cammarata, M., Eds.; Elsevier Inc.: London, UK, 2016; pp. 193–199. [Google Scholar]
- Parrinello, N. Cytotoxic activity of tunicate hemocytes. In Invertebrate Immunology; Rinkevich, B., Müller, W.E.G., Eds.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 190–217. [Google Scholar]
- Liu, L.; Zheng, Y.-Y.; Shao, C.-L.; Wang, C.-Y. Metabolites from marine invertebrates and their symbiotic microorganisms: Molecular diversity discovery, mining, and application. Mar. Life Sci. Technol. 2019, 1, 60–94. [Google Scholar] [CrossRef] [Green Version]
- Leal, M.C.; Sheridan, C.; Osinga, R.; Dionísio, G.; Rocha, R.; Silva, B.; Rosa, R.; Calado, C. Marine Microorganism-Invertebrate Assemblages: Perspectives to Solve the “Supply Problem” in the Initial Steps of Drug Discovery. Mar. Drugs 2014, 12, 3929–3952. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Hu, J.S.; Xu, J.L.; Shao, C.L.; Wang, G.Y. Biological and chemical diversity of ascidian-associated microorganisms. Mar. Drugs 2018, 16, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oltz, E.M.; Smith, M.J.; Nakanishi, K.; Bruening, R.C.; Kustin, K. The tunichromes. A class of reducing blood pigments from sea squirts: Isolation, structures, and vanadium chemistry. J. Am. Chem. Soc. 1988, 110, 6162–6172. [Google Scholar] [CrossRef] [PubMed]
- Kustin, K.; Robinson, W.E.; Smith, M.J. Tunichromes, vanadium, and vacuolated blood cells in tunicates. Invertebr. Reprod. Dev. 1990, 17, 129–139. [Google Scholar] [CrossRef]
- Lee, I.H.; Zhao, C.; Cho, Y.; Harwig, S.S.L.; Cooper, E.L.; Lehrer, R.I. Clavanins, α-helical antimicrobial peptides from tunicate hemocytes. FEBS Lett. 1997, 400, 158–162. [Google Scholar] [CrossRef] [Green Version]
- Cima, F.; Franchi, N.; Ballarin, L. Origin and functions of tunicate hemocytes. In The Evolution of the Immune System: Conservation and Diversification; Malagoli, D., Ed.; Academic Press: London, UK, 2016; pp. 29–49. ISBN 9780128020135. [Google Scholar]
- Galinier, R.; Roger, E.; Sautiere, P.E.; Aumelas, A.; Banaigs, B.; Mitta, G. Halocyntin and papillosin, two new antimicrobial peptides isolated from hemocytes of the solitary tunicate, Halocynthia papillosa. J. Pept. Sci. 2009, 15, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Jang, W.S.; Kim, K.N.; Lee, Y.S.; Nam, M.H.; Lee, I.H. Halocidin: A new antimicrobial peptide from hemocytes of the solitary tunicate, Halocynthia aurantium. FEBS Lett. 2002, 521, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Gorbman, A.; Whiteley, A.; Kavanaugh, S. Pheromonal stimulation of spawning release of gametes by gonadotropin releasing hormone in the chiton, Mopalia sp. Gen. Comp. Endocrinol. 2003, 131, 62–65. [Google Scholar] [CrossRef]
- Evans, J.S.; Erwin, P.M.; Shenkar, N.; López-Legentil, S. Introduced ascidians harbor highly diverse and host-specific symbiotic microbial assemblages. Sci. Rep. 2017, 7, 11033. [Google Scholar] [CrossRef] [Green Version]
- Tianero, M.D.B.; Kwan, J.C.; Wyche, T.P.; Presson, A.P.; Koch, M.; Barrows, L.R.; Bugni, T.S.; Schmidt, E.W. Species specificity of symbiosis and secondary metabolism in ascidians. ISME J. 2015, 9, 615–628. [Google Scholar] [CrossRef] [Green Version]
- Flood, P. Yellow-stained oikopleurid appendicularians are caused by bacterial parasitism. Mar. Ecol. Prog. Ser. 1991, 71, 291–295. [Google Scholar] [CrossRef]
- Cuadrado, A.; García-Fernández, L.F.; González, L.; Suárez, Y.; Losada, A.; Alcaide, V.; Martínez, T.; Máa Fernández-Sousa, J.; Sánchez-Puelles, J.M.; Muñoz, A. AplidinTM induces apoptosis in human cancer cells via glutathione depletion and sustained activation of the epidermal growth factor receptor, Src, JNK, and p38 MAPK. J. Biol. Chem. 2003, 278, 241–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitsiades, C.S.; Ocio, E.M.; Pandiella, A.; Maiso, P.; Gajate, C.; Garayoa, M.; Vilanova, D.; Montero, J.C.; Mitsiades, N.; McMullan, C.J.; et al. Aplidin, a marine organism-derived compound with potent antimyeloma activity in vitro and in vivo. Cancer Res. 2008. [Google Scholar] [CrossRef] [Green Version]
- Urdiales, J.L.; Morata, P.; De Castro, I.N.; Sánchez-Jiménez, F. Antiproliferative effect of dehydrodidemnin B (DDB), a depsipeptide isolated from Mediterranean tunicates. Cancer Lett. 1996, 102, 31–37. [Google Scholar] [CrossRef]
- Fedorov, S.N.; Radchenko, O.S.; Shubina, L.K.; Balaneva, N.N.; Bode, A.M.; Stonik, V.A.; Dong, Z. Evaluation of cancer-preventive activity and structure– activity relationships of 3-demethylubiquinone Q2, isolated from the ascidian Aplidium glabrum, and it synthetic analogs. Pharm. Res. 2006, 23, 70–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido, L.; Zubía, E.; Ortega, M.J.; Salvá, J. Haouamines A and B: A new class of alkaloids from the ascidian Aplidium haouarianum. J. Org. Chem. 2003, 68, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Park, N.S.; Park, Y.K.; Ramalingam, M.; Yadav, A.K.; Cho, H.R.; Hong, V.S.; More, K.N.; Bae, J.H.; Bishop-Bailey, D.; Kano, J.; et al. Meridianin C inhibits the growth of YD-10B human tongue cancer cells through macropinocytosis and the down-regulation of Dickkopf-related protein-3. J. Cell. Mol. Med. 2018, 22, 5833–5846. [Google Scholar] [CrossRef]
- Núñez-Pons, L.; Carbone, M.; Vázquez, J.; Rodríguez, J.; Nieto, R.M.; Varela, M.M.; Gavagnin, M.; Avila, C. Natural products from antarctic colonial ascidians of the genera Aplidium and Synoicum: Variability and defensive role. Mar. Drugs 2012, 10, 1741–1764. [Google Scholar] [CrossRef] [Green Version]
- Paul, V.J.; Lindquist, N.; Fenical, W. Chemical defenses of the tropical ascidian Atapozoa sp. and its nudibranch predators Nembrotha spp. Mar. Ecol. Prog. Ser. 1990, 59, 109–118. [Google Scholar] [CrossRef]
- Choi, H.; Hwang, H.; Chin, J.; Kim, E.; Lee, J.; Nam, S.J.; Lee, B.C.; Rho, B.J.; Kang, H. Tuberatolides, potent FXR antagonists from the korean marine tunicate Botryllus tuberatus. J. Nat. Prod. 2011, 74, 90–94. [Google Scholar] [CrossRef]
- Kubanek, J.; Williams, D.E.; de Silva, E.D.; Allen, T.; Andersen, R.J. Cytotoxic alkaloids from the flatworm Prostheceraeus villatus and its tunicate prey Clavelina lepadiformis. Tetrahedron Lett. 1995, 36, 6189–6192. [Google Scholar] [CrossRef]
- Raub, M.F.; Cardellina, J.H.; Choudhary, M.I.; Ni, C.Z.; Clardy, J.; Alley, M.C. Clavepictines A and B: Cytotoxic Quinolizidines from the Tunicate Clavelina picta. J. Am. Chem. Soc. 1991, 113, 3178–3180. [Google Scholar] [CrossRef]
- Abourriche, A.; Abboud, Y.; Maoufoud, S.; Mohou, H.; Seffaj, T.; Charrouf, M.; Chaib, N.; Bennamara, A.; Bontemps, N.; Francisco, C. Cynthichlorine: A bioactive alkaloid from the tunicate Cynthia savignyi. Farmaco 2003, 58, 1351–1354. [Google Scholar] [CrossRef]
- Kobayashi, J.; Cheng, J.F.; Nakamura, H.; Ohizumi, Y.; Walchli, M.R.; Hirata, Y.; Sasaki, T. Cystodytins A, B, and C, novel tetracyclic aromatic alkaloids with potent antineoplastic activity from the Okinawan tunicate Cystodytes dellechiajei. J. Org. Chem. 1988, 53, 1800–1804. [Google Scholar] [CrossRef]
- Kobayashi, J.; Tsuda, M.; Tanabe, A.; Ishibashi, M.; Cheng, J.F.; Yamamura, S.; Sasaki, T. Cystodytins D-I, new cytotoxic tetracyclic aromatic alkaloids from the okinawan marine tunicate Cystodytes dellechiajei. J. Nat. Prod. 1991, 54, 1634–1638. [Google Scholar] [CrossRef] [PubMed]
- Dassonneville, L.; Wattez, N.; Baldeyrou, B.; Mahieu, C.; Lansiaux, A.; Banaigs, B.; Bonnard, I.; Bailly, C. Inhibition of topoisomerase II by the marine alkaloid ascididemin and induction of apoptosis in leukemia cells. Biochem. Pharmacol. 2000, 60, 527–537. [Google Scholar] [CrossRef]
- López-Legentil, S.; Turon, X.; Schupp, P. Chemical and physical defenses against predators in Cystodytes (Ascidiacea). J. Exp. Mar. Biol. Ecol. 2006, 332, 27–36. [Google Scholar] [CrossRef]
- Uchimasu, H.; Matsumura, K.; Tsuda, M.; Kumagai, K.; Akakabe, M.; Fujita, M.J.; Sakai, R. Mellpaladines and dopargimine, novel neuroactive guanidine alkaloids from a Palauan Didemnidae tunicate. Tetrahedron 2016, 72, 7185–7193. [Google Scholar] [CrossRef]
- Torii, M.; Hitora, Y.; Kato, H.; Koyanagi, Y.; Kawahara, T.; Losung, F.; Mangindaan, R.E.P.; Tsukamoto, S. Siladenoserinols M-P, sulfonated serinol derivatives from a tunicate. Tetrahedron 2018, 74, 7516–7521. [Google Scholar] [CrossRef]
- Machida, K.; Arai, D.; Katsumata, R.; Otsuka, S.; Yamashita, J.K.; Ye, T.; Tang, S.; Fusetani, N.; Nakao, Y. Sameuramide A, a new cyclic depsipeptide isolated from an ascidian of the family Didemnidae. Bioorg. Med. Chem. 2018, 26, 3852–3857. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.D.; Goclik, E.; König, G.M.; Kaminsky, R. Lepadins D-F: Antiplasmodial and antitrypanosomal decahydroquinoline derivatives from the tropical marine tunicate Didemnum sp. J. Med. Chem. 2002, 45, 3067–3072. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.S.; Rhodes, D.; Bushman, F.D.; Faulkner, D.J. Cyclodidemniserinol trisulfate, a sulfated serinolipid from the Palauan ascidian Didemnum guttatum that inhibits HIV-1 integrase. Org. Lett. 2000, 2, 1605–1607. [Google Scholar] [CrossRef]
- Seleghim, M.H.R.; De Lira, S.P.; Campana, P.T.; Berlinck, R.G.S.; Custódio, M.R. Localization of granulatimide alkaloids in the tissues of the ascidian Didemnum granulatum. Mar. Biol. 2007, 150, 967–975. [Google Scholar] [CrossRef]
- Smith, T.E.; Pond, C.D.; Pierce, E.; Harmer, Z.P.; Kwan, J.; Zachariah, M.M.; Harper, M.K.; Wyche, T.P.; Matainaho, T.K.; Bugni, T.S.; et al. Accessing chemical diversity from the uncultivated symbionts of small marine animals. Nat. Chem. Biol. 2018, 14, 179–185. [Google Scholar] [CrossRef]
- Donia, M.S.; Wang, B.; Dunbar, D.C.; Desai, P.V.; Patny, A.; Avery, M.; Hamann, M.T. Mollamides B and C, cyclic hexapeptides from the indonesian tunicate Didemnum molle. J. Nat. Prod. 2008, 71, 941–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oku, N.; Matsunaga, S.; Fusetani, N. Shishijimicins A-C, novel enediyne antitumor antibiotics from the ascidian Didemnum proliferum. J. Am. Chem. Soc. 2003, 125, 2044–2045. [Google Scholar] [CrossRef]
- Takeara, R.; Jimenez, P.C.; Wilke, D.V.; Moraes, M.O.d.; Pessoa, C.; Lopes, N.P.; Lopes, J.L.C.; Lotufo, T.M.d.C.; Costa-Lotufo, L.V. Antileukemic effects of Didemnum psammatodes (Tunicata: Ascidiacea) constituents. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2008, 151, 363–369. [Google Scholar] [CrossRef]
- Bracegirdle, J.; Robertson, L.P.; Hume, P.A.; Page, M.J.; Sharrock, A.V.; Ackerley, D.F.; Carroll, A.R.; Keyzers, R.A. Lamellarin Sulfates from the Pacific Tunicate Didemnum ternerratum. J. Nat. Prod. 2019, 82, 2000–2008. [Google Scholar] [CrossRef]
- Charyulu, G.A.; McKee, T.C.; Ireland, C.M. Diplamine, a cytotoxic polyaromatic alkaloid from the tunicate Diplosoma sp. Tetrahedron Lett. 1989, 30, 4201–4202. [Google Scholar] [CrossRef]
- Ogi, T.; Taira, J.; Margiastuti, P.; Ueda, K. Cytotoxic metabolites from the Okinawan ascidian Diplosoma virens. Molecules 2008, 13, 595–602. [Google Scholar] [CrossRef]
- Cvetkovic, R.S.; Figgitt, D.P.; Plosker, G.L. ET-743. Drugs 2002, 62, 1185–1192. [Google Scholar] [CrossRef]
- Rashid, M.A.; Gustafson, K.R.; Boyd, M.R. New cytotoxic n-methylated β-carboline alkaloids from the marine ascidian Eudistoma gilboverde. J. Nat. Prod. 2001, 64, 1454–1456. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, J.; Harbour, G.C.; Gilmore, J.; Rinehart, K.L. Eudistomins A, D, G, H, I, J, M, N, O, P, and Q, Bromo-, Hydroxy-, Pyrrolyl-, and 1-Pyrrolinyl-β-carbolines from the antiviral Caribbean tunicate Eudistoma olivaceum. J. Am. Chem. Soc. 1984, 106, 1526–1528. [Google Scholar] [CrossRef]
- Rinehart, K.L.; Kobayashi, J.; Harbour, G.C.; Hughes, R.G.; Mizsak, S.A.; Scahill, T.A. Eudistomins C, E, K, and L, potent antiviral compounds containing a novel oxathiazepine ring from the Caribbean tunicate Eudistoma olivaceum. J. Am. Chem. Soc. 1984, 106, 1524–1526. [Google Scholar] [CrossRef]
- Jimenez, P.C.; Wilke, D.V.; Ferreira, E.G.; Takeara, R.; De Moraes, M.O.; Silveira, E.R.; Lotufo, T.M.D.C.; Lopes, N.P.; Costa-Lotufo, L.V. Structure elucidation and anticancer activity of 7-oxostaurosporine derivatives from the Brazilian endemic tunicate Eudistoma vannamei. Mar. Drugs 2012, 10, 1092–1102. [Google Scholar] [CrossRef] [Green Version]
- Rajesh, R.P.; Annappan, M. Anticancer effects of brominated indole alkaloid eudistomin H from marine ascidian Eudistoma viride against cervical cancer cells (HeLa). Anticancer Res. 2015, 35, 283–294. [Google Scholar]
- Tapiolas, D.M.; Bowden, B.F.; Abou-Mansour, E.; Willis, R.H.; Doyle, J.R.; Muirhead, A.N.; Liptrot, C.; Llewellyn, L.E.; Wolff, C.W.W.; Wright, A.D.; et al. Eusynstyelamides A, B, and C, nNOS inhibitors, from the ascidian Eusynstyela latericius. J. Nat. Prod. 2009, 72, 1115–1120. [Google Scholar] [CrossRef]
- Devi, S.; Rajasekharan, K.; Padmakumar, K.; Tanaka, J.; Higa, T. Biological activity and chemistry of the compound ascidian Eusynstyela tineta. In The Biology of Ascidians; Sawada, H., Yokosawa, H., Lambert, C.C., Eds.; Springer: Tokyo, Japan, 2001; pp. 341–354. [Google Scholar]
- Tsukamotol, S.; Kato, H.; Hirota, H.; Fusetane, N. Lumichrome Is a putative intrinsic substance inducing larval metamorphosis in the ascidian Halocynthia roretzi. In The Biology of Ascidians; Sawada, H., Yokosawa, H., Lambert, C.C., Eds.; Springer: Tokyo, Japan, 2001; pp. 335–340. [Google Scholar]
- Azumi, K.; Yokosawa, H.; Ishii, S.I. Halocyamines: Novel antimicrobial tetrapeptide-like substances isolated from the hemocytes of the solitary ascidian Halocynthia roretzi. Biochemistry 1990, 29, 159–165. [Google Scholar] [CrossRef]
- Oda, T.; Fujiwara, T.; Liu, H.; Ukai, K.; Mangindaan, R.E.P.; Mochizuki, M.; Namikoshi, M. Effects of lissoclibadins and lissoclinotoxins, isolated from a tropical ascidian Lissoclinum cf. badium, on IL-8 production in a PMA-stimulated promyelocytic leukemia cell line. Mar. Drugs 2006, 4, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Kumaran, N.S.; Bragadeeswaran, S.; Meenakshi, V.K.; Balasubramanian, T. Bioactivity potential of extracts from ascidian Lissoclinum fragile. Afr. J. Pharm. Pharmacol. 2012, 6, 1854–1859. [Google Scholar] [CrossRef] [Green Version]
- Corley, D.G.; Moore, R.E.; Paul, V.J. Patellazole B: A novel cytotoxic thiazole-containing macrolide from the marine tunicate Lissoclinum patella. J. Am. Chem. Soc. 1988, 110, 7920–7922. [Google Scholar] [CrossRef]
- Zabriskie, T.M.; Mayne, C.L.; Ireland, C.M. Patellazole C: A novel cytotoxic macrolide from Lissoclinum patella. J. Am. Chem. Soc. 1988, 110, 7919–7920. [Google Scholar] [CrossRef]
- Odate, S.; Pawlik, J.R. The role of vanadium in the chemical defense of the solitary tunicate, Phallusia nigra. J. Chem. Ecol. 2007, 33, 643–654. [Google Scholar] [CrossRef]
- Casertano, M.; Imperatore, C.; Luciano, P.; Aiello, A.; Putra, M.Y.; Gimmelli, R.; Ruberti, G.; Menna, M. Chemical investigation of the indonesian tunicate Polycarpa aurata and evaluation of the effects against Schistosoma mansoni of the novel alkaloids polyaurines A and B. Mar. Drugs 2019, 17, 278. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.; Fenical, W. Polycarpine dihydrochloride: A cytotoxic dimeric disulfide alkaloid from the Indian ocean ascidian Polycarpa clavata. Tetrahedron Lett. 1996, 37, 2369–2372. [Google Scholar] [CrossRef]
- Guo, P.; Wang, Z.; Li, G.; Liu, Y.; Xie, Y.; Wang, Q. First discovery of polycarpine, polycarpaurines A and C, and their derivatives as novel antiviral and antiphytopathogenic fungus agents. J. Agric. Food Chem. 2016, 64, 4264–4272. [Google Scholar] [CrossRef]
- Fujita, M.; Nakao, Y.; Matsunaga, S.; Nishikawa, T.; Fusetani, N. Sodium 1-(12-hydroxy)octadecanyl sulfate, an MMP2 inhibitor, isolated from a tunicate of the family polyclinidae. J. Nat. Prod. 2002, 65, 1936–1938. [Google Scholar] [CrossRef]
- Nicolaou, K.C.; Li, R.; Lu, Z.; Pitsinos, E.N.; Alemany, L.B. Total synthesis and full structural assignment of namenamicin. J. Am. Chem. Soc. 2018, 140, 8091–8095. [Google Scholar] [CrossRef]
- Cheng, M.T.; Rinehart, K.L. Polyandrocarpidines: Antimicrobial and Cytotoxic Agents from a Marine Tunicate (Polyandrocarpa sp.) from the Gulf of California. J. Am. Chem. Soc. 1978, 100, 7409–7411. [Google Scholar] [CrossRef]
- Lindquist, N.; Hay, M.E.; Fenical, W. Defense of ascidians and their conspicuous larvae: Adult vs. larval chemical defenses. Ecol. Monogr. 1992, 62, 547–568. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, N.; Katsuyama, Y.; Kawamura, K.; Fujiwara, S. Regeneration of the gut requires retinoic acid in the budding ascidian Polyandrocarpa misakiensis. Dev. Growth Differ. 2010, 52, 457–468. [Google Scholar] [CrossRef]
- Wang, W.; Kim, H.; Nam, S.J.; Rho, B.J.; Kang, H. Antibacterial butenolides from the korean tunicate Pseudodistoma antinboja. J. Nat. Prod. 2012, 27, 574–577. [Google Scholar] [CrossRef] [PubMed]
- Appleton, D.R.; Page, M.J.; Lambert, G.; Berridge, M.V.; Copp, B.R. Kottamides A–D: Novel bioactive imidazolone-containing alkaloids from the New Zealand ascidian Pycnoclavella kottae. J. Org. Chem. 2002, 67, 5402–5404. [Google Scholar] [CrossRef]
- Aiello, A.; Carbonelli, S.; Fattorusso, E.; Iuvone, T.; Menna, M. New bioactive sulfated metabolites from the Mediterranean tunicate Sidnyum turbinatum. J. Nat. Prod. 2001, 64, 219–221. [Google Scholar] [CrossRef]
- Davies-Coleman, M.T.; Cantrell, C.L.; Gustafson, K.R.; Beutler, J.A.; Pannell, L.K.; Boyd, M.R. Stolonic acids A and B, new cytotoxic cyclic peroxides from an Indian Ocean ascidian Stolonica species. J. Nat. Prod. 2000, 63, 1411–1413. [Google Scholar] [CrossRef]
- Menzel, L.P.; Lee, I.H.; Sjostrand, B.; Lehrer, R.I. Immunolocalization of clavanins in Styela clava hemocytes. Dev. Comp. Immunol. 2002, 26, 505–515. [Google Scholar] [CrossRef]
- Raftos, D.A.; Hutchinson, A. Cytotoxicity reactions in the solitary tunicate Styela plicata. Dev. Comp. Immunol. 1995, 19, 463–471. [Google Scholar] [CrossRef]
- Miyata, Y. Ecdysteroids from the Antarctic tunicate Synoicum adareanum. J. Nat. Prod. 2007, 70, 1859–1864. [Google Scholar] [CrossRef]
- Rinehart, K.L.; Gloer, J.B.; Hughes, R.G.; Renis, H.E.; Patrick McGovren, J.; Swynenberg, E.B.; Stringfellow, D.A.; Kuentzel, S.L.; Li, L.H. Didemnins: Antiviral and antitumor depsipeptides from a Caribbean tunicate. Science 1981, 212, 933–935. [Google Scholar] [CrossRef]
- Rodríguez-Martínez, R.E.; Jordán-Garza, A.G.; Baker, D.M.; Jordán-Dahlgren, E. Competitive interactions between corals and Trididemnum solidum on Mexican Caribbean reefs. Coral Reefs 2012, 31, 571–577. [Google Scholar] [CrossRef]
- Schofield, M.M.; Jain, S.; Porat, D.; Dick, G.J.; Sherman, D.H. Identification and analysis of the bacterial endosymbiont specialized for production of the chemotherapeutic natural product ET-743. Environ. Microbiol. 2015, 17, 3964–3975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayanetti, D.R.; Braun, D.R.; Barns, K.J.; Rajski, S.R.; Bugni, T.S. Bulbiferates A and B: Antibacterial acetamidohydroxybenzoates from a marine proteobacterium, Microbulbifer sp. J. Nat. Prod. 2019, 82, 1930–1934. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, H.; Nakayama, W.; Takahashi, O.; Kirikoshi, R.; Izumikawa, Y.; Iwasaki, K.; Toraiwa, K.; Ukai, K.; Rotinsulu, H.; Wewengkang, D.S.; et al. Verruculides A and B, two new protein tyrosine phosphatase 1B inhibitors from an Indonesian ascidian-derived Penicillium verruculosum. Bioorganic Med. Chem. Lett. 2015, 25, 3087–3090. [Google Scholar] [CrossRef]
- Pinkerton, D.M.; Banwell, M.G.; Garson, M.J.; Kumar, N.; De Moraes, M.O.; Cavalcanti, B.C.; Barros, F.W.A.; Pessoa, C. Antimicrobial and cytotoxic activities of synthetically derived tambjamines C and E-J, BE-18591, and a related alkaloid from the marine bacterium Pseudoalteromonas tunicata. Chem. Biodivers. 2010, 7, 1311–1324. [Google Scholar] [CrossRef]
- Burke, C.; Thomas, T.; Egan, S.; Kjelleberg, S. The use of functional genomics for the identification of a gene cluster encoding for the biosynthesis of an antifungal tambjamine in the marine bacterium Pseudoalteromonas tunicata: Brief report. Environ. Microbiol. 2007, 9, 814–818. [Google Scholar] [CrossRef]
- Wang, K.L.; Xu, Y.; Lu, L.; Li, Y.; Han, Z.; Zhang, J.; Shao, C.L.; Wang, C.Y.; Qian, P.Y. Low-toxicity diindol-3-ylmethanes as potent antifouling compounds. Mar. Biotechnol. 2015, 17, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Takagi, M.; Motohashi, K.; Izumikawa, M.; Khan, S.T.; Hwang, J.-H.; Shin-Ya, K. JBIR-66, a new metabolite isolated from tunicate-derived Saccharopolyspora sp. SS081219JE-28. Biosci. Biotechnol. Biochem. 2010, 74, 2355–2357. [Google Scholar] [CrossRef] [Green Version]
- Wasserman, H.H.; Friedland, D.J.; Morrison, D.A. A novel dipyrrolyldipyrromethene prodigiosin analog from Serratia marcescens. Tetrahedron Lett. 1968, 6, 641–644. [Google Scholar] [CrossRef]
- Sung, A.A.; Gromek, S.M.; Balunas, M.J. Upregulation and identification of antibiotic activity of a marine-derived Streptomyces sp. via co-cultures with human pathogens. Mar. Drugs 2017, 15, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukimoto, M.; Nagaoka, M.; Shishido, Y.; Fujimoto, J.; Nishisaka, F.; Matsumoto, S.; Harunari, E.; Imada, C.; Matsuzaki, T. Bacterial production of the tunicate-derived antitumor cyclic depsipeptide didemnin B. J. Nat. Prod. 2011, 74, 2329–2331. [Google Scholar] [CrossRef]
- Karthikeyan, M.M.; Ananthan, G.; Balasubramanian, T. Antimicrobial activity of crude extracts of some ascidians (Urochordata: Ascidiacea), from Palk Strait, (Southeast Coast of India). World J. Fish. Mar. Sci. 2009, 1, 262–267. [Google Scholar]
- Ayuningrum, D.; Kristiana, R.; Nisa, A.A.; Radjasa, S.K.; Muchlissin, S.I.; Radjasa, O.K.; Sabdono, A.; Trianto, A. Bacteria associated with tunicate, Polycarpa aurata, from Lease sea, Maluku, Indonesia exhibiting anti-multidrug resistant bacteria. Biodiversitas 2019, 20, 956–964. [Google Scholar] [CrossRef] [Green Version]
- Litaay, M.; Christine, G.; Gobel, R.B.; Dwyana, Z. Bioactivity of endo-symbiont bacteria of tunicate Polycarpa aurata as antimicrobial. In Proceedings of the 23 National Seminar of Indonesia Biology Society, Jayapura, Indonesia, 18 September 2015. [Google Scholar]
- Menna, M.; Fattorusso, E.; Imperatore, C. Alkaloids from marine ascidians. Molecules 2011, 16, 8694–8732. [Google Scholar] [CrossRef]
- Franks, A.; Haywood, P.; Holmström, C.; Egan, S.; Kjelleberg, S.; Kumar, N. Isolation and structure elucidation of a novel yellow pigment from the marine bacterium Pseudoalteromonas tunicata. Molecules 2005, 10, 1286–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franks, A.; Egan, S.; Holmström, C.; James, S.; Lappin-Scott, H.; Kjelleberg, S. Inhibition of fungal colonization by Pseudoalteromonas tunicata provides a competitive advantage during surface colonization. Appl. Environ. Microbiol. 2006, 72, 6079–6087. [Google Scholar] [CrossRef] [Green Version]
- Egan, S.; James, S.; Holmström, C.; Kjelleberg, S. Correlation between pigmentation and antifouling compounds produced by Pseudoalteromonas tunicata. Environ. Microbiol. 2002, 4, 433–442. [Google Scholar] [CrossRef]
- Matz, C.; Webb, J.S.; Schupp, P.J.; Phang, S.Y.; Penesyan, A.; Egan, S.; Steinberg, P.; Kjelleberg, S. Marine biofilm bacteria evade eukaryotic predation by targeted chemical defense. PLoS ONE 2008, 3, e2744. [Google Scholar] [CrossRef] [Green Version]
- Holmström, C.; James, S.; Neilan, B.A.; White, D.C.; Kjelleberg, S. Pseudoalteromonas tunicata sp. nov., a bacterium that produces antifouling agents. Int. J. Syst. Bacteriol. 1998, 48, 1205–1212. [Google Scholar] [CrossRef] [Green Version]
- James, S.G.; Holmström, C.; Kjelleberg, S. Purification and characterization of a novel antibacterial protein from the marine bacterium D2. Appl. Environ. Microbiol. 1996, 62, 2783–2788. [Google Scholar] [CrossRef] [Green Version]
- Fedders, H.; Michalek, M.; Grötzinger, J.; Leippe, M. An exceptional salt-tolerant antimicrobial peptide derived from a novel gene family of haemocytes of the marine invertebrate Ciona intestinalis. Biochem. J. 2008, 416, 65–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nurfadillah, A.; Litaay, M.; Gobel, R.B.; Haedar, N. Potency of tunicate Polycarpa aurata as inoculum source of sebagai sumber endosimbyotic fungi that produce antimicrobe. J. Alam Lingkung. 2015, 6, 10–16. [Google Scholar]
- Tahir, E.; Litaay, M.; Gobel, R.B.; Haedar, N.; Al, E. Potency of tunicate Rhopalaea crassa as inoculum source of endosymbiont fungi that produce antimicrobe. Spermonde 2016, 2, 33–37. [Google Scholar]
- Shaala, L.A.; Youssef, D.T.A. Identification and bioactivity of compounds from the fungus Penicillium sp. CYE-87 isolated from a marine tunicate. Mar. Drugs 2015, 13, 1698–1709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canonico, P.G.; Pannier, W.L.; Huggins, J.W.; Rienehart, K.L. Inhibition of RNA viruses in vitro and in Rift Valley fever-infected mice by didemnins A and B. Antimicrob. Agents Chemother. 1982, 22, 696–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, A.M.S.; Glaser, K.B.; Cuevas, C.; Jacobs, R.S.; Kem, W.; Little, R.D.; McIntosh, J.M.; Newman, D.J.; Potts, B.C.; Shuster, D.E. The odyssey of marine pharmaceuticals: A current pipeline perspective. Trends Pharmacol. Sci. 2010, 31, 255–265. [Google Scholar] [CrossRef]
- Zelek, L.; Yovine, A.; Brain, E.; Turpin, F.; Taamma, A.; Riofrio, M.; Spielmann, M.; Jimeno, J.; Misset, J.L. A phase II study of Yondelis® (trabectedin, ET-743) as a 24-h continuous intravenous infusion in pretreated advanced breast cancer. Br. J. Cancer 2006, 94, 1610–1614. [Google Scholar] [CrossRef] [Green Version]
- Atmaca, H.; Bozkurt, E.; Uzunoglu, S.; Uslu, R.; Karaca, B. A diverse induction of apoptosis by trabectedin in MCF-7 (HER2−/ER+) and MDA-MB-453 (HER2+/ER−) breast cancer cells. Toxicol. Lett. 2013, 221, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Grosso, F.; Jones, R.L.; Demetri, G.D.; Judson, I.R.; Blay, J.-Y.; Cesne, A.L.; Lippo, R.S.; Casieri, P.; Collini, P.; Dileo, P.; et al. Effi cacy of trabectedin (ecteinascidin-743) in advanced pretreated myxoid liposarcomas: A retrospective study. Lancet Oncol. 2007, 8, 595–602. [Google Scholar] [CrossRef]
- Sessa, C.; De Braud, F.; Perotti, A.; Bauer, J.; Curigliano, G.; Noberasco, C.; Zanaboni, F.; Gianni, L.; Marsoni, S.; Jimeno, J.; et al. Trabectedin for women with ovarian carcinoma after treatment with platinum and taxanes fails. J. Clin. Oncol. 2005, 23, 1867–1874. [Google Scholar] [CrossRef]
- Krasner, C.N.; McMeekin, D.S.; Chan, S.; Braly, P.S.; Renshaw, F.G.; Kaye, S.; Provencher, D.M.; Campos, S.; Gore, M.E. A Phase II study of trabectedin single agent in patients with recurrent ovarian cancer previously treated with platinum-based regimens. Br. J. Cancer 2007, 97, 1618–1624. [Google Scholar] [CrossRef] [Green Version]
- Monk, B. A randomized phase III study of trabectedin with pegylated liposomal doxorubicin (PLD) versus PLD in relapsed, recurrent ovarian cancer (OC). Eur. J. Cancer Suppl. 2008, 19, viii1–viii4. [Google Scholar]
- Rath, C.M.; Janto, B.; Earl, J.; Ahmed, A.; Hu, F.Z.; Hiller, L.; Dahlgren, M.; Kreft, R.; Yu, F.; Wolff, J.J.; et al. Meta-omic characterization of the marine invertebrate microbial consortium that produces the chemotherapeutic natural product ET-743. ACS Chem. Biol. 2011, 6, 1244–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tohme, R.; Darwiche, N.; Gali-Muhtasib, H. A journey under the sea: The quest for marine anti-cancer alkaloids. Molecules 2011, 16, 9665–9696. [Google Scholar] [CrossRef]
- González-Santiago, L.; Suárez, Y.; Zarich, N.; Muñoz-Alonso, M.J.; Cuadrado, A.; Martínez, T.; Goya, L.; Iradi, A.; Sáez-Tormo, G.; Maier, J.V.; et al. Aplidin® induces JNK-dependent apoptosis in human breast cancer cells via alteration of glutathione homeostasis, Rac1 GTPase activation, and MKP-1 phosphatase downregulation. Cell Death Differ. 2006, 13, 1968–1981. [Google Scholar] [CrossRef] [PubMed]
- Rinehart, K.L. Antitumor compounds from tunicates. Med. Res. Rev. 2000, 20, 1–27. [Google Scholar] [CrossRef]
- Kobayashi, J.; Cheng, J.F.; Nakamura, H.; Ohta, T.; Nozoe, S.; Hirata, Y.; Sasaki, T. Lejimalides A and B, novel 24-membered macrolides with potent antileukemic activity from the Okinawan tunicate Eudistoma cf. rigida. J. Org. Chem. 1988, 53, 6147–6150. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Ishibashi, M.; Sasaki, T.; Kobayashi, J. Lejimalides C and D, new antineoplastic 24-membered macrolide sulfates from the okinawan marine tunicate Eudistoma cf. rigida. Tetrahedron Lett. 1991, 32, 789–797. [Google Scholar] [CrossRef]
- Nguyen, M.H.; Imanishi, M.; Kurogi, T.; Wan, X.; Ishmael, J.E.; McPhail, K.L.; Smith, A.B. Synthetic access to the mandelalide family of macrolides: Development of an anion relay chemistry strategy. J. Org. Chem. 2018, 83, 4287–4306. [Google Scholar] [CrossRef]
- Rinehart, K.L.; Holt, T.G.; Fregeau, N.L.; Stroh, J.G.; Keifer, P.A.; Sun, F.; Li, L.H.; Martin, D.G. Ecteinascidins 729, 743, 745, 759A, 759B, and 770: Potent antitumor agents from the Caribbean tunicate Ecteinascidia turbinata. J. Org. Chem. 1990, 55, 4512–4515. [Google Scholar] [CrossRef]
- Izbicka, E.; Lawrence, R.; Raymond, E.; Eckhardt, G.; Faircloth, G.; Jimeno, J.; Clark, G.; Von Hoff, D.D. In vitro antitumor activity of the novel marine agent, Ecteinascidin-743 (ET-743, NSC-648766) against human tumors explanted from patients. Ann. Oncol. 1998, 9, 981–987. [Google Scholar] [CrossRef]
- Berlmck, R.G.S.; Britton, R.; Piers, E.; Lim, L.; Roberge, M.; Moreira Da Rocha, R.; Andersen, R.J. Granulatimide and isogranulatimide, aromatic alkaloids with G2 checkpoint inhibition activity isolated from the Brazilian ascidian Didemnum granulatum: Structure elucidation and synthesis. J. Org. Chem. 1998, 63, 9850–9856. [Google Scholar] [CrossRef]
- Holmstrom, C.; James, S.; Egan, S.; Kjelleberg, S. Inhibition of common fouling organisms by marine bacterial isolates with special reference to the role of pigmented bacteria. Biofouling 1996, 10, 251–259. [Google Scholar] [CrossRef]
- Holmstrom, C.; Rittschof, D.; Kjelleberg, S. Inhibition of settlement by larvae of Balanus amphitrite and Ciona intestinalis by a surface-colonizing marine bacterium. Appl. Environ. Microbiol. 1992, 58, 2111–2115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoecker, D. Resistance of a tunicate to fouling. Biol. Bull. 1978, 155, 615–626. [Google Scholar] [CrossRef]
- Vervoort, H.C.; Pawlik, J.R.; Fenical, W. Chemical defense of the Caribbean ascidian Didemnum conchyliatum. Mar. Ecol. Prog. Ser. 1998, 164, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Lindquist, N.; Hay, M.E. Can small rare prey be chemically defended? The case for marine larvae. Ecology 1995, 76, 1347–1358. [Google Scholar] [CrossRef]
- Lindquist, N.; Fenical, W. New tambjamine class alkaloids from the marine ascidian Atapozoa sp. and its nudibranch predators. Origin of the tambjamines in Atapozoa. Experientia 1991, 47, 504–506. [Google Scholar] [CrossRef]
- McClintock, J.B.; Amsler, M.O.; Koplovitz, G.; Amsler, C.D.; Baker, B.J. Observations on an association between the dexaminid amphipod Polycheria antarctica f. acanthopoda and its ascidian host Distaplia cylindrica. J. Crustac. Biol. 2009, 29, 605–608. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, T.; Shibata, H.; Ebine, M.; Tsuchikawa, H.; Matsumori, N.; Murata, M.; Yoshida, M.; Morisawa, M.; Lin, S.; Yamauchi, K.; et al. Synthesis and complete structure determination of a sperm-activating and -attracting factor isolated from the ascidian ascidia sydneiensis. J. Nat. Prod. 2018, 81, 985–997. [Google Scholar] [CrossRef]
- Mikami, N.; Hosokawa, M.; Miyashita, K. Effects of sea squirt (Halocynthia roretzi) lipids on white adipose tissue weight and blood glucose in diabetic/obese KK-Ay mice. Mol. Med. Rep. 2010, 3, 449–453. [Google Scholar] [PubMed]
- Thakur, N.L. Studies on Some Bioactive Aspects of Selected Marine Organisms; Goa University: Goa, India, 2001. [Google Scholar]
- Kazlauskas, R.; Marwood, J.F.; Murphy, P.T.; Wells, R.J. A blue pigment from a compound ascidian. Aust. J. Chem. 1982, 35, 215–217. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.E.; Forleo, D.A.; Gunawardana, G.P.; Gunasekera, S.P.; Koehn, F.E.; McConnell, O.J. Antitumor tetrahydroisoquinoline alkaloids from the colonial ascidian Ecteinascidia turbinata. J. Org. Chem. 1990, 55, 4508–4515. [Google Scholar] [CrossRef]
- Kott, P. Didemnid-algal symbiosis: Host species in the Western Pacific with notes on the symbiosis. Micronesica 1982, 18, 95–127. [Google Scholar]
- Sings, H.L.; Rinehart, K.L. Compounds produced from potential tunicate-blue-green algal symbiosis: A review. J. Ind. Microbiol. Biotechnol. 1996, 17, 385–396. [Google Scholar] [CrossRef]
- Carté, B.; Faulkner, D.J. Defensive metabolites from three nembrothid nudibranchs. J. Org. Chem. 1983, 48, 2314–2318. [Google Scholar] [CrossRef]
- Garson, M.J. Marine natural products as antifeedants. In Comprehensive Natural Products II: Chemistry and Biology; Elsevier Ltd.: Amsterdam, The Netherlands, 2010; pp. 503–537. ISBN 9780080453828. [Google Scholar]
- Lins, D.M.; de Marco, P.; Andrade, A.F.A.; Rocha, R.M. Predicting global ascidian invasions. Divers. Distrib. 2018, 24, 692–704. [Google Scholar] [CrossRef] [Green Version]
- Gittenberger, A.; Moons, J.J.S. Settlement and possible competition for space between the invasive violet tunicate Botrylloides violaceus and the native star tunicate Botryllus schlosseri in The Netherlands. Aquat. Invasions 2011, 6, 435–440. [Google Scholar] [CrossRef]
- Sephton, D.; Vercaemer, B.; Nicolas, J.M.; Keays, J. Monitoring for invasive tunicates in Nova Scotia, Canada (2006–2009). Aquat. Invasions 2011, 6, 391–403. [Google Scholar] [CrossRef]
- Seo, K.S.; Lee, Y. A first assessment of invasive marine species on Chinese and Korean coasts. In Biological Invasions in Marine Ecosystems, Ecological Studies 204; Rilov, G., Crooks, J.A., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 577–585. [Google Scholar]
- Costello, K.E.; Lynch, S.A.; McAllen, R.; O’Riordan, R.M.; Culloty, S.C. The role of invasive tunicates as reservoirs of molluscan pathogens. Biol. Invasions 2021, 23, 641–655. [Google Scholar] [CrossRef]
- Daigle, R.M.; Herbinger, C.M. Ecological interactions between the vase tunicate (Ciona intestinalis) and the farmed blue mussel (Mytilus edulis) in Nova Scotia, Canada. Aquat. Invasions 2009, 4, 177–187. [Google Scholar] [CrossRef]
- Ramesh, C.H.; Koushik, S.; Shunmugaraj, T.; Murthy, M.V.R. Infestation of colonial ascidians on reef biota of Gulf of Mannar Marine Biosphere Reserve, India. J. New Biol. Rep. 2019, 8, 187–189. [Google Scholar]
- Vargas-Ángel, B.; Godwin, L.S.; Asher, J.; Brainard, R.E. Invasive didemnid tunicate spreading across coral reefs at remote Swains Island, American Sāmoa. Coral Reefs 2009, 28, 53. [Google Scholar] [CrossRef] [Green Version]
- Tatián, M.; Schwindt, E.; Lagger, C.; Varela, M.M. Colonization of Patagonian harbours (SW Atlantic) by an invasive sea squirt. Spixiana 2010, 33, 111–117. [Google Scholar]
- Pleus, A.; LeClair, L.; Schultz, J.; Lambert, G. 2007–09 Tunicate Management Plan; In Coordination with the Tunicate Response Advisory Committee; Washington State Department of Fish and Wildlife, Aquatic Invasive Species Unit: Olympia, WA, USA, 2008; pp. 1–64.
- Cordell, J.R.; Levy, C.; Toft, J.D. Ecological implications of invasive tunicates associated with artificial structures in Puget Sound, Washington, USA. Biol. Invasions 2013, 15, 1303–1318. [Google Scholar] [CrossRef]
- Abdul Jaffar, H.; Soban Akram, A.; Kaleem Arshan, M.L.; Sivakumar, V.; Tamilselvi, M. Distribution and invasiveness of a colonial ascidian, Didemnum psammathodes, along the southern Indian coastal water. Oceanologia 2016, 58, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Carman, M.R.; Grunden, D.W. First occurrence of the invasive tunicate Didemnum vexillum in eelgrass habitat. Aquat. Invasions 2010, 5, 23–29. [Google Scholar] [CrossRef]
- Griffith, K.; Mowat, S.; Holt, R.H.F.; Ramsay, K.; Bishop, J.D.D.; Lambert, G.; Jenkins, S.R. First records in Great Britain of the invasive colonial ascidian Didemnum vexillum Kott, 2002. Aquat. Invasions 2009, 4, 581–590. [Google Scholar] [CrossRef]
- Sorte, C.J.B.; Williams, S.L.; Zerebecki, R.A. Ocean warming increases threat of invasive species in a marine fouling community. Ecology 2010, 91, 2198–2204. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Database | No. of Known Compounds | No. of Unknown Compounds | Known Chemical Compound | Biological Properties |
|---|---|---|---|---|
| BIAdb | 1 | - | Polycarpine | Cytotoxic, antiviral, and antifungal |
| BindingDB | 2 | - | Tuberatolides, Sodium 1-(12-hydroxy) octadecanyl sulfate | Farnesoid X receptor antagonists, matrix metalloproteinase 2 inhibitor |
| ChemDB | 2 | - | Patellazole B, Patellazole C | Antimicrobial, cytotoxic |
| ChEMBL | 2 | Ascididemin, Trabectedin | Anticancer | |
| ChemSpider | 1 | - | Trabectedin | Anticancer |
| DrugBank | - | - | ||
| HIT | - | - | ||
| HMDB | 1 | - | Trabectedin | Anticancer |
| KEGG | 1 | - | Trabectedin | Anticancer |
| NCI | - | - | ||
| NPACT | - | - | ||
| PDB-Bind | - | - | ||
| PDBeChem | 16 | >30 | Cystodytin D, cystodytin F, cystodytin E, cystodytin G, cystodytin H, cystodytin I, Diplosoma ylidene 1, Diplosoma ylidene 2, Lejimalide A, lejimalide B, lissoclibadin 1, lissoclibadin 2, lissoclibadin 3, lamellarin alpha 20-sulfate, plitidepsin, trabectedin | Cytotoxic, anticancer |
| PharmaGKB | 1 | - | Trabectedin | Anticancer |
| PubChem | 4 | 2 | Patellazole B, Patellazole C, GnRH-II, GnRH-I | Antimicrobial and cytotoxic, induces spawning |
| SMPDB | - | - | ||
| SuperDrug | 1 | - | Trabectedin | Anticancer |
| TTD | - | - | ||
| UniProt | 1 | 1 | Retinoic acid | Regeneration of gut |
| ZINC | 1 | - | Trabectedin | Anticancer |
| Body Component | Chemical Compound | Function | Application | Reference |
|---|---|---|---|---|
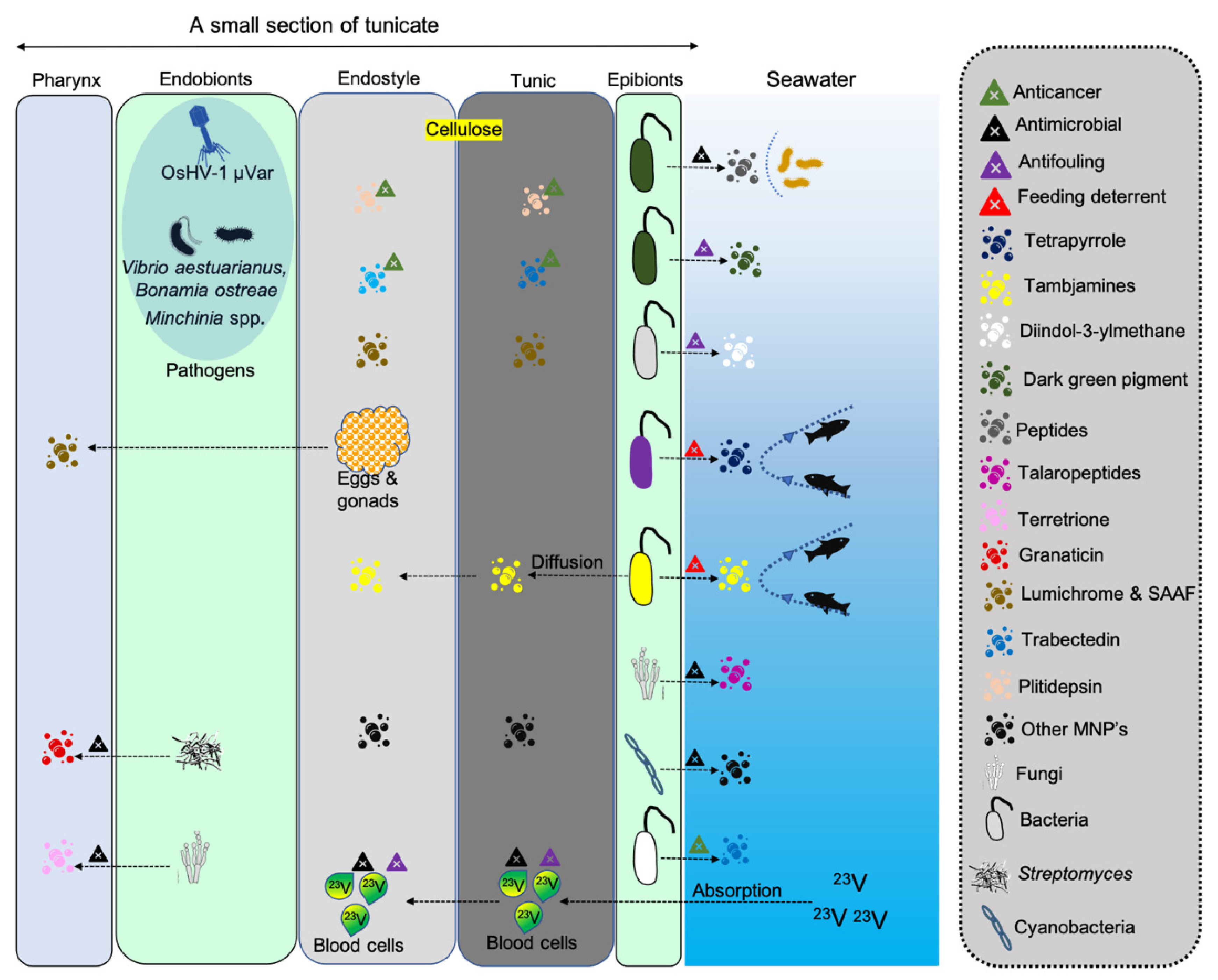
| Tunic (Ascidia sp., Ciona intestinalis, Halocynthia roretzi, and Styela plicata) | Tunicin (cellulose) | Protection | Material cellulose | [35] |
| Blood (Ascidia nigra, Molgula manhattensis) | Vanadium | Oxygen transport | [42] | |
| Blood (Ascidia nigra) | Tunichromes | Vanadium binding and reduces blood pigments | [42,43] | |
| Hemocytes (Styela clava) | Clavanins | Multiple functions | Antimicrobial | [44,45] |
| Hemocytes (Halocynthia papillosa) | Halocyntin and papillosin | Antimicrobial | [46] | |
| Hemocytes (Halocynthia aurantium) | Halocidin | Antimicrobial | [47] | |
| Gonad (Unknown sp.) | GnRH-2 peptide | Pheromone-like function | Induce spawning | [48] |
| MNPs from Tunicates | Chemical Compound | Function | Application | Reference |
|---|---|---|---|---|
| Aplidium albicans | Aplidin | Anticancer | [52,53] | |
| Aplidium albicans | Dehydrodidemnin B | Antitumor | [54] | |
| Aplidium glabrum | Quinones | Anticancer, cytotoxic | [55] | |
| Aplidium haouarianum | Haouamine A | Cytotoxic activity | [56] | |
| Aplidium meridianum | Meridianins | Anticancer, antibiofilm | [57] | |
| Aplidium & Synoicum | Meridianins | Feeding deterrents | Antibacterial | [58] |
| Atapozoa sp. | Tambjamine | Feeding deterrents | [59] | |
| Botryllus tuberatus | Tuberatolides | Farnesoid X receptor antagonists | [60] | |
| Clavelina lepadiformis | Lepadins and villatamines | Antiparasitic, anticancer | [61] | |
| Clavelina picta | Clavepictine A and B | Antimicrobial, cytotoxicity | [62] | |
| Cynthia savignyi | Cynthichlorine | Antifungal, cytotoxicity | [63] | |
| Cystodytes dellechiajei | Cystodytins A-I | Antitumor, cytotoxic | [64,65] | |
| Cystodytes dellechiajei | Ascididemin | Antitumor | [66] | |
| Cystodytes sp. | Ascididemin | Feeding deterrents | Antifeedant | [67] |
| Didemnidae | Mellpaladine and dopargimine | Neuroactive | [68] | |
| Didemnidae | Siladenoserinols A and B | Antitumor | [69] | |
| Didemnidae | Sameuramide A | Colony formation | [70] | |
| Didemnum sp. | Lepadins D-F | Antiplasmodial and antitrypanosomal | [71] | |
| Didemnum guttatum | Cyclodidemniserinol trisulfate | Anti-retroviral | [72] | |
| Didemnum granulatum | Granulatamides | Deterrent activity | [73] | |
| Didemnum molle | Lanthipeptide divamide A | anti-HIV drug | [74] | |
| Didemnum molle | Mollamide B | Anticancer | [75] | |
| Didemnum proliferum | Shishijimicins | Antitumor | [76] | |
| Didemnum psammatodes | Methyl esters | Antiproliferative | [77] | |
| Didemnum ternerratum | Lamellarin Sulfates | Anticancer | [78] | |
| Diplosoma sp. | Diplamine | Antibacterial and cytotoxic | [79] | |
| Diplosoma virens | Diplosoma ylidene 1, Diplosoma ylidene 2 | Anticancer | [80] | |
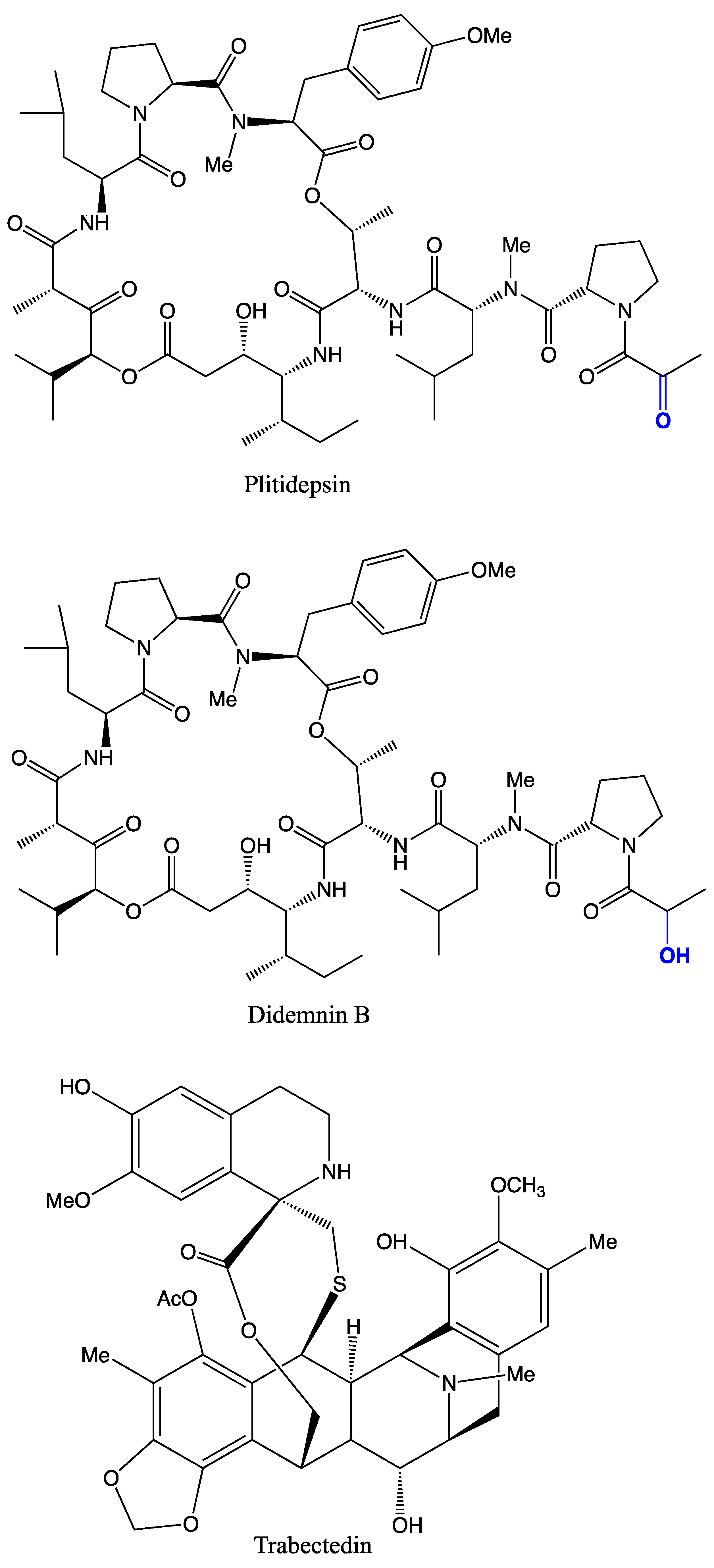
| Ecteinascidia turbinata | Ecteinascidin 743 (Trabectedin) | Anticancer | [81] | |
| Eudistoma gilboverde | Methyleudistomins | Antitumor | [82] | |
| Eudistoma olivaceum | Eudistomins G and H | Chemical defense | Antifouling | [34] |
| Eudistoma olivaceum | Eudistomins A, D, G, H, I, J, M, N, O, P, and Q | Antiviral | [83] | |
| Eudistoma olivaceum | Eudistomins C, E, K, and L | Antiviral | [84] | |
| Eudistoma vannamei | 7-Oxostaurosporine | Anticancer | [85] | |
| Eudistoma viride | Eudistomins H | Anticancer | [86] | |
| Eusynstyela latericius | Eusynstyelamides A, B | Antibacterial | [87] | |
| Eusynstyela tincta | Kuanoniamine A | Chemical defense | Antimicrobial, antitumor, antifouling | [88] |
| Halocynthia aurantium | Halocidin | Antimicrobial | [47] | |
| Halocynthia papillosa | Halocyntin and papillosin | Antimicrobial | [46] | |
| Halocynthia roretzi | Lumichrome | Larval metamorphosis | [89] | |
| Halocynthia roretzi | Halocyamine A and B | Antimicrobial, anticancer | [90] | |
| Lissoclinum cf. badium | Lissoclibadins | Anticancer | [91] | |
| Lissoclinum fragile | Antimicrobial, hemolytic, and cytotoxic | [92] | ||
| Lissoclinum patella | Patellazole B and C | Antimicrobial, cytotoxic | [93,94] | |
| Phallusia nigra | Vanadium chloride, vanadyl sulfate | Antimicrobial | [95] | |
| Polycarpa aurata | Polyaurines A and B | Antiparasitic | [96] | |
| Polycarpa clavata | Polycarpine dihydrochloride | Cytotoxic | [97] | |
| Polycarpa clavata | Polycarpaurines A and C | Antiviral, antifungal | [98] | |
| Polyclinidae | Sodium 1-(12-hydroxy) octadecanyl sulfate | Matrix metalloproteinase 2 inhibitor | [99] | |
| Polysyncraton lithostrotum | Namenamicin | Cytotoxic, antitumor | [100] | |
| Polyandrocarpa sp. | Polyandrocarpidines | Antimicrobial, cytotoxic, and deterrent activities | [101,102] | |
| Polyandrocarpa misakiensis | Retinoic acid | Regeneration of gut | [103] | |
| Pseudodistoma antinboja | Cadiolides J-M | Antibacterial | [104] | |
| Pycnoclavella kottae | Kottamide D | Cytotoxic, anti-inflammatory, and antimetabolic activities | [105] | |
| Sidnyum turbinatum | Alkyl sulfates | Antiproliferative | [106] | |
| Stolonica sp. | Stolonic acid A and B | Antiproliferative | [107] | |
| Styela clava | Clavanins | Antimicrobial | [108] | |
| Styela plicata | Hemocytes | Cytotoxic | [109] | |
| Synoicum adareanum | Hyousterones and Abeohyousterone | Cytotoxic and anticancer | [110] | |
| Trididemnum solidum | Didemnins A, B, and C | Antiviral, cytotoxic | [111,112] | |
| MNPs from associated microbes | ||||
| Candidatus Endoecteinascidia frumentensis | Tetrahydroisoquinoline | [113] | ||
| Microbulbifer sp. | Bulbiferates A and B | Antibacterial | [114] | |
| Penicillium verruculosum | Verruculides A, chrodrimanins A and H | Protein tyrosine phosphatase 1B inhibition | [115] | |
| Pseudoalteromonas rubra | Isatin | Microbial defense | Antibacterial | [16] |
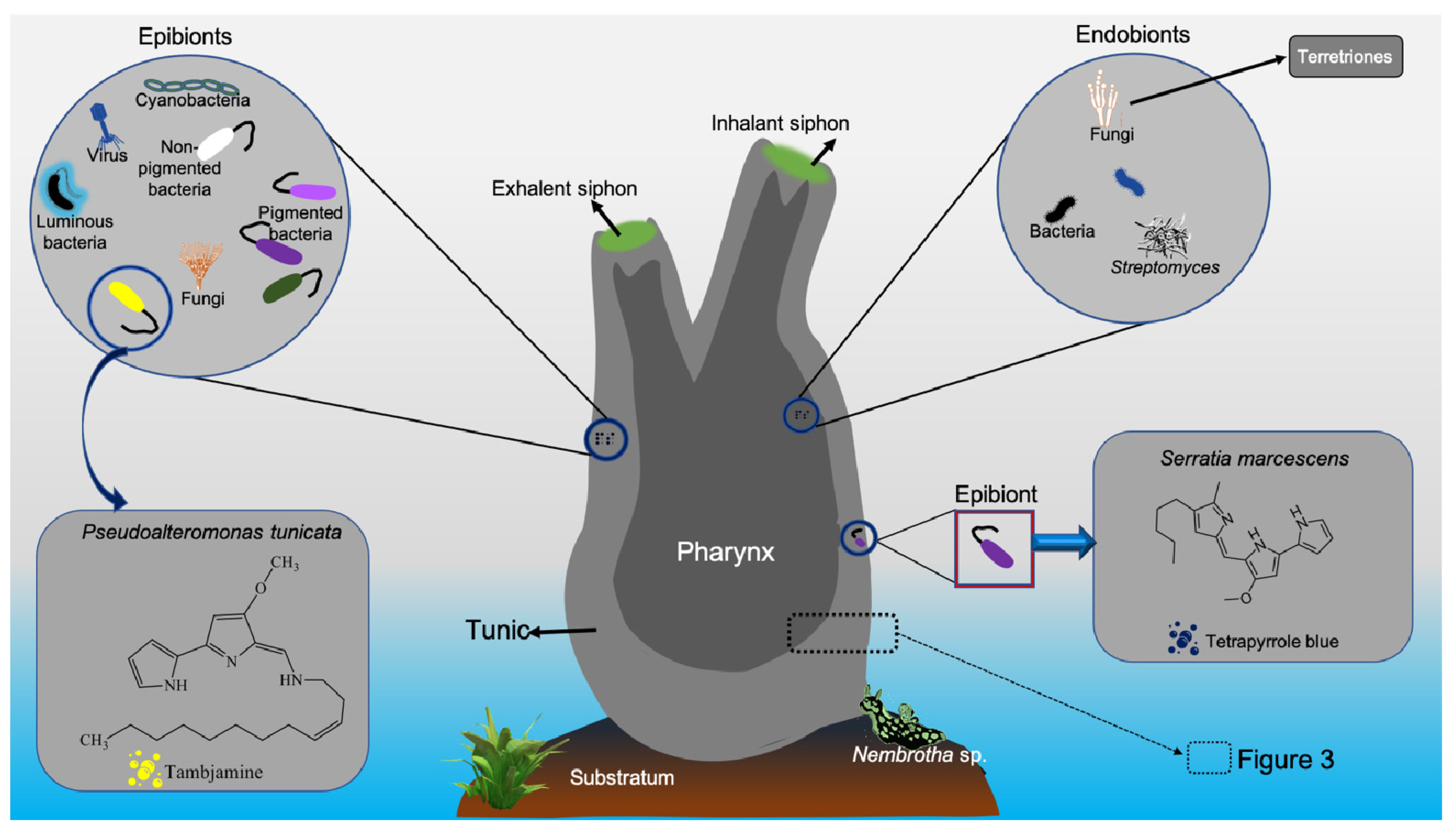
| Pseudoalteromonas tunicata | Tambjamine | Feeding deterrents | [116] | |
| Pseudoalteromonas tunicata | Tambjamine | Antifungal | [117] | |
| Pseudovibrio denitrificans | Diindol-3-ylmethanes | Antifouling | [118] | |
| Saccharopolyspora sp. | JBIR-66 | Cytotoxic | [119] | |
| Serratia marcescens | Tetrapyrrole pigment | Feeding deterrents | [120] | |
| Streptomyces sp. | Granaticin, granatomycin D, and dihydrogranaticin B | Antibacterial | [121] | |
| Talaromyces sp. | Talaropeptides A-D | Plasma stability, Antibacterial, antifungal, cytotoxic | [24] | |
| Tistrella mobilis and Tistrella bauzanensis | Didemnin | Anticancer | [23,122] |
| Application | Compound | Activity against | Dose/ Concentration | Growth Inhibition (Diameter/ Percentage) | Assay Method | Reference |
|---|---|---|---|---|---|---|
| Antimicrobial | ||||||
| Clavanins | E. coli, L. monocytogenes, C. albicans | 1.6–3.5 μg/mL | - | Radial diffusion assay | [44] | |
| Diplamine | E. coli, S. aureus | - | [79] | |||
| Halocidin | Methicillin-resistant Staphylococcus aureus and multidrug-resistant Pseudomonas aeruginosa | 100–200 μg/mL | 5–11 mm | Radial diffusion assay | [47] | |
| Isatin | Bacillus cereus, Bacillus megaterium, Escherichia coli, Micrococcus luteus, | MIC 200 μg/mL | 7–>21 mm | Disk diffusion assay | [16] | |
| Kuanoniamine A | B. Subtilis, E. coli, S. aureus, V. cholerae, V. parahaemolyticus and fungus A. jumigatus and C. albicans | 25 μg/mL | 7–13 mm | Disk diffusion assay | [88] | |
| Cynthichlorine | A. radiobacter, E. coli, P. aeruginosa, Botrytis cinerea, Verticillium albo atrum | 6–10 mm | Disc diffusion assay | [63] | ||
| Talaropeptides A and B | Bacillus subtilis | IC50 1.5–3.7 µM | 50% | Microtiter plate assay | [24] | |
| Terretrione C and D | Candida albicans | MIC 32 µg/mL | 17–19 mm | Disc diffusion assay | [136] | |
| Anticancer & antitumor | ||||||
| Aplidin | Multiple myeloma cell lines, MDA-MB-231 breast cancer cells, A-498 and ACHN cell lines | IC50 1 to 15 nmol/L | Nuclear Staining Assay; MTT assay | [52,53] | ||
| Clavepictines A and B | Murine leukemia and human solid tumor cell lines | IC50 12 μg/mL | Microculture tetrazolium assay | [62] | ||
| Dehydrodidemnin B | Ehrlich carcinoma cells | 2.5 μg/mouse | 70–90% | MTT assay | [54] | |
| Didemnins A and B | Leukaemia P388 cells | IC50 1.5–25 μg/mL | - | [111] | ||
| Diplamine | Leukemia L1210 cells | IC50 2×10-2 μg/mL | - | [79] | ||
| Ecteinascidin 743 (Trabectedin) | Leukemia L1210 cells | IC50 0.5 μg/mL | - | [152] | ||
| Eudistomins H | HeLa cell lines | IC50 0.49 μg/mL | 60% | MTT assay | [86] | |
| Halocyamine A and B | Rat neuronal cells, mouse neuroblastoma N-18 cells, and human Hep-G2 cells | - | [90] | |||
| Kuanoniamine A | Dalton’s lymphoma and Ehrlich ascites tumour cell lines | 25 μg/mL | 100% | Trypan blue exclusion test | [88] | |
| Lamellarin Sulfates | HCT-116 human colon tumor cells | IC50 9.7 μM | MTS cell proliferation assay | [78] | ||
| Namenamicin | P388 leukemia cells, 3Y1, and HeLa | IC50 3.5 nM; IC50 3.3–13 pM | Biochemical prophage induction assay | [100] | ||
| Polycarpine dihydrochloride | HCT-116 human colon tumor cells | ED50 1.9 μg/mL | - | [97] | ||
| 7-oxostaurosporine | HL-60, Molt-4, Jurkat, K562, HCT-8, MDA MB-435, and SF-295 cell lines | IC50 10–58 nM | 95% | MTT assay | [85] | |
| Terretrione C and D | Human breast cancer cells | IC50 16.5 and 17.6 μM | Sulforhodamine B assay | [136] | ||
| Antifouling | ||||||
| Diindol-3-ylmethanes | Barnacle, Balanus amphitrite and bryozoan, Bugula neritina | EC50 18.57 | Microtiter plate assay | [118] | ||
| Eudistomins G and H | Fish and other larvae | Antifeedant assay | [34] |
| Invasive Tunicate | Country | Origin Type | Negative Impacts | Reference |
|---|---|---|---|---|
| Ascidiella aspersa | Argentina | Exotic | Space competition | [179] |
| Botrylloides violaceus | Netherlands | Exotic | Space competition | [172] |
| Botryllus schlosseri | Netherlands | Indigenous | Space competition | [172] |
| Botryllus schlosseri, Botrylloides violaceus, Ciona intestinalis, Ciona savignyi, Didemnum vexillum, Molgula manhattensis, Styela clava | USA | Exotic | Competitors for food and space | [180,181] |
| Ciona intestinalis | Canada | Exotic | Mussel mortality | [176] |
| Ciona intestinalis | Korea | Exotic | Space competition and damage to aquaculture | [174] |
| Didemnum psammathodes | India | Indigenous | Space competition | [182] |
| Didemnum vexillum | USA | Exotic | Threat to eelgrass | [183] |
| Didemnum vexillum | Wales | Exotic | Space competition | [184] |
| Diplosoma similis | American Sāmoa | Indigenous | Kill corals | [178] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramesh, C.; Tulasi, B.R.; Raju, M.; Thakur, N.; Dufossé, L. Marine Natural Products from Tunicates and Their Associated Microbes. Mar. Drugs 2021, 19, 308. https://doi.org/10.3390/md19060308
Ramesh C, Tulasi BR, Raju M, Thakur N, Dufossé L. Marine Natural Products from Tunicates and Their Associated Microbes. Marine Drugs. 2021; 19(6):308. https://doi.org/10.3390/md19060308
Chicago/Turabian StyleRamesh, Chatragadda, Bhushan Rao Tulasi, Mohanraju Raju, Narsinh Thakur, and Laurent Dufossé. 2021. "Marine Natural Products from Tunicates and Their Associated Microbes" Marine Drugs 19, no. 6: 308. https://doi.org/10.3390/md19060308