
In Vitro Bioactivity of Astaxanthin and Peptides from Hydrolisates of Shrimp (Parapenaeus longirostris) By-Products: From the Extraction Process to Biological Effect Evaluation, as Pilot Actions for the Strategy “From Waste to Profit”

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Proximate Composition and Fatty Acid Profile of P. longirostris By-Product
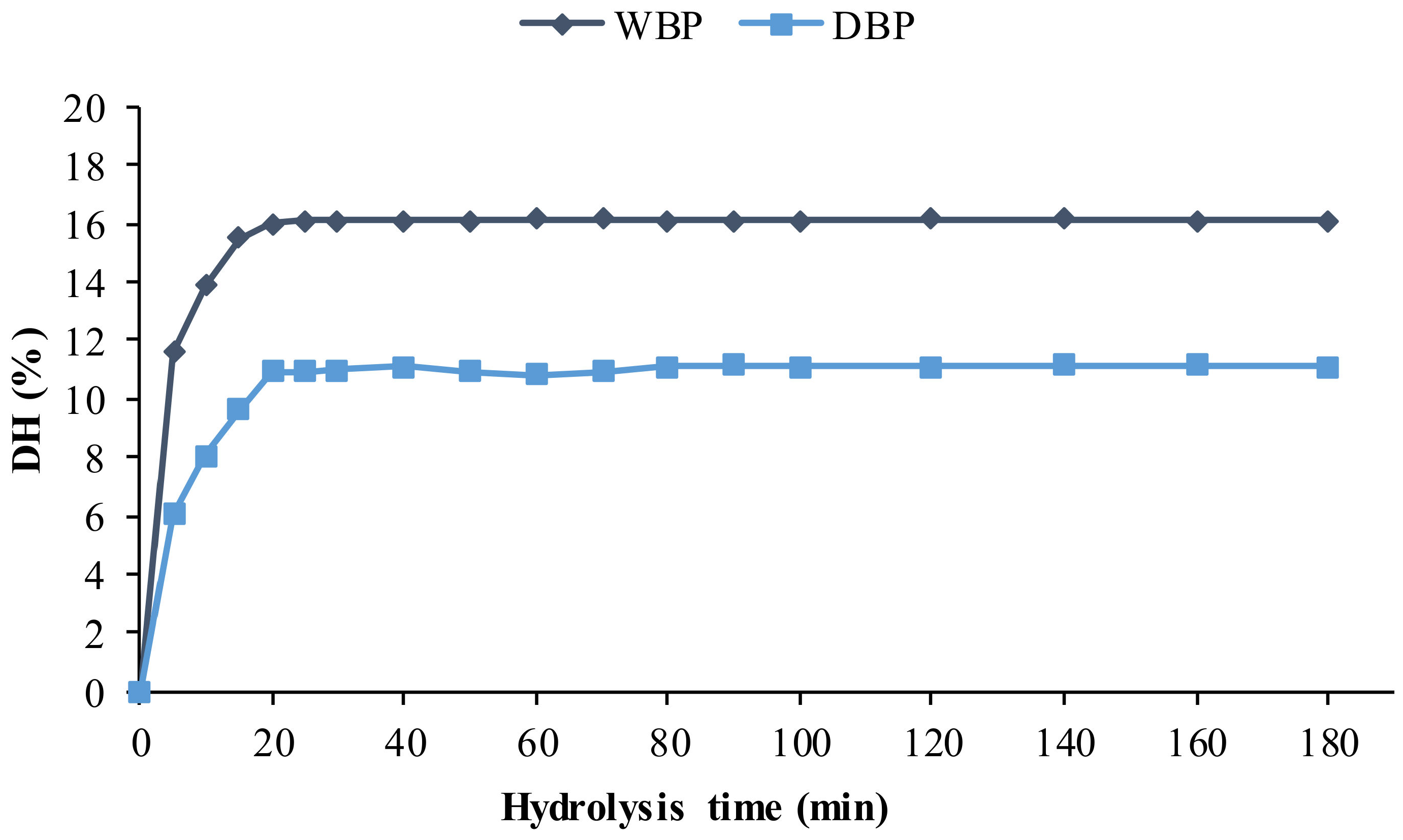
2.2. Enzymatic Hydrolysis
2.3. Hydrolysates Characterization and Bioactive Properties of the Protein Fractions
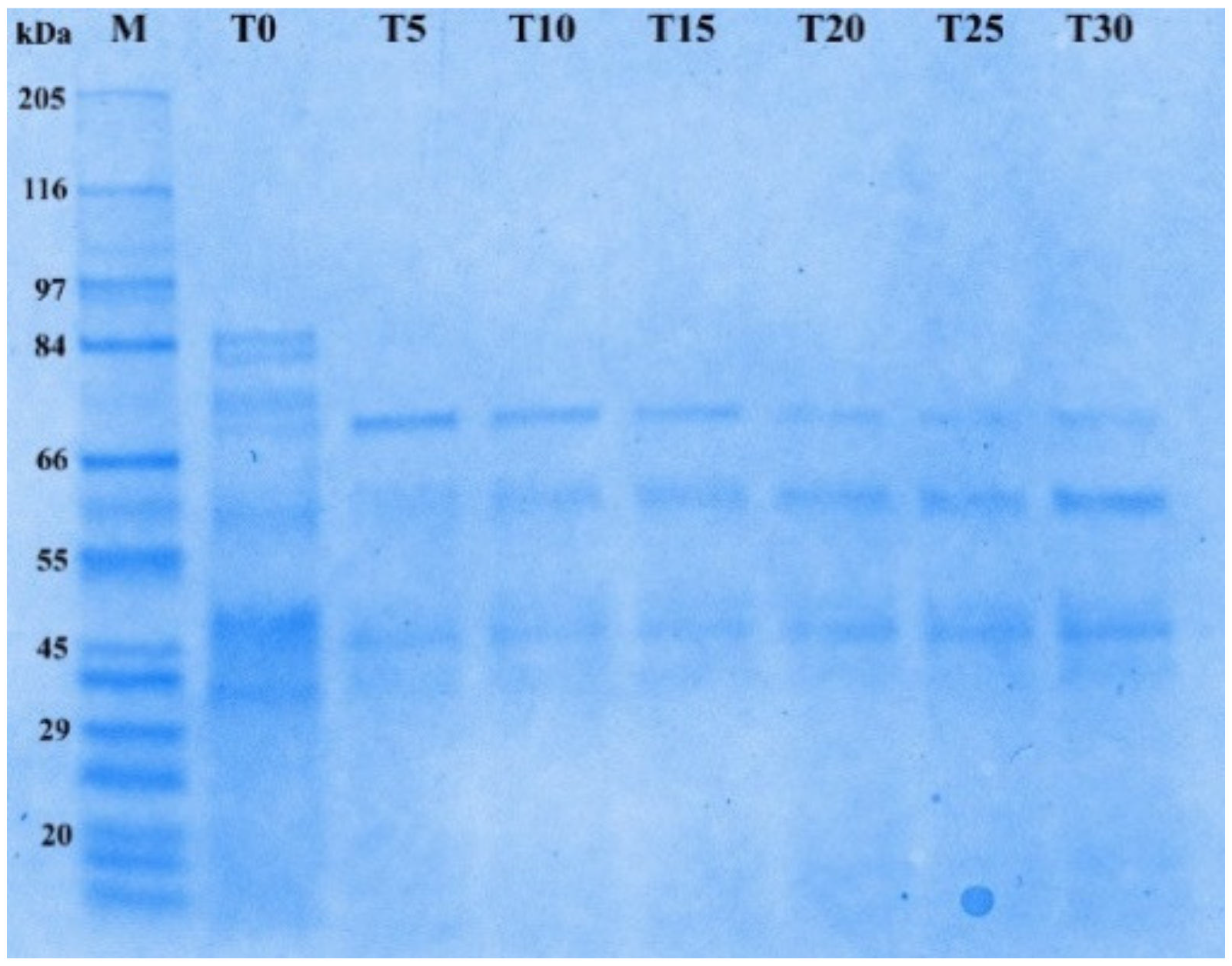
2.3.1. SDS PAGE
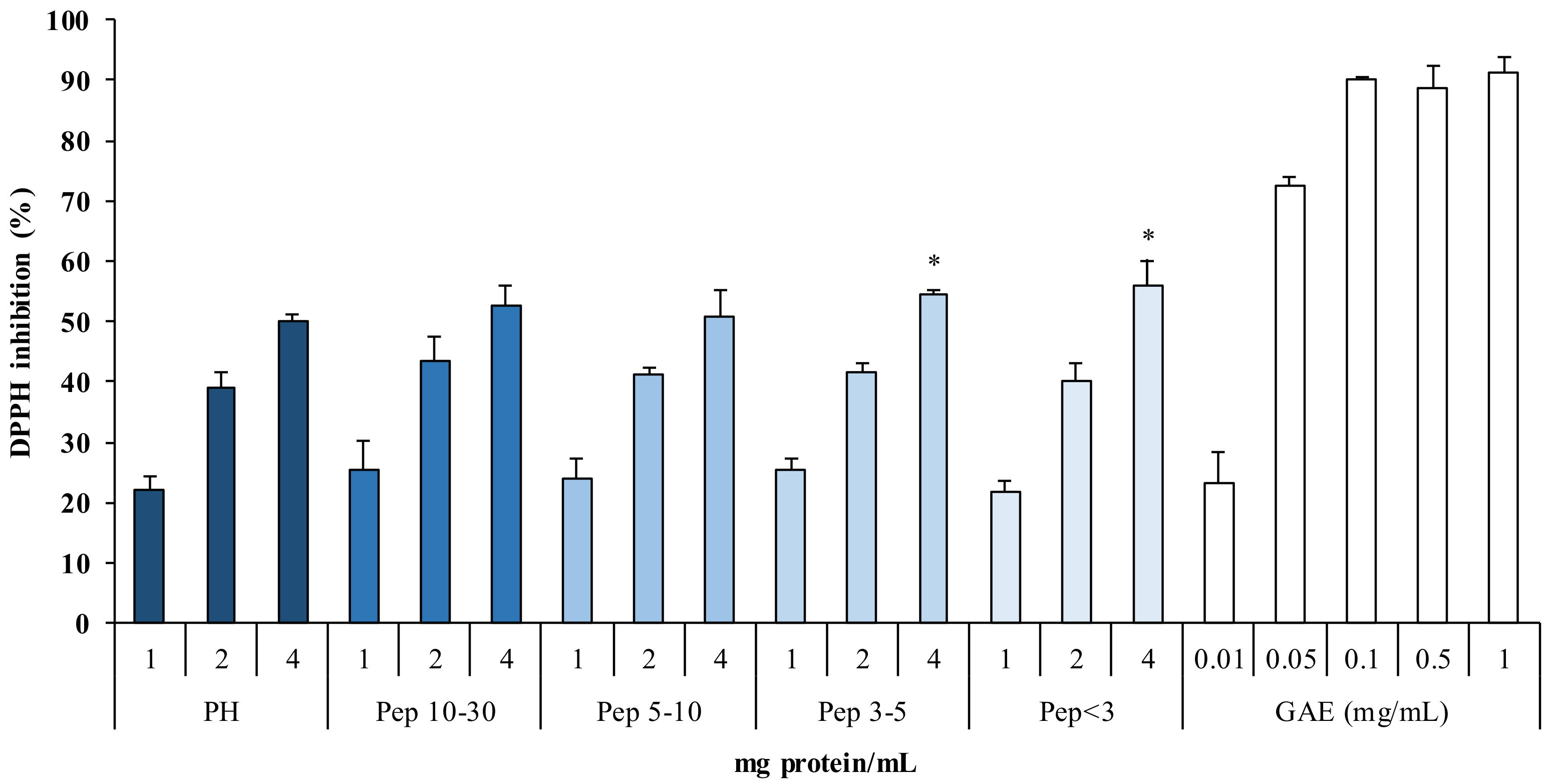
2.3.2. Antioxidant Power of Protein Hydrolysates
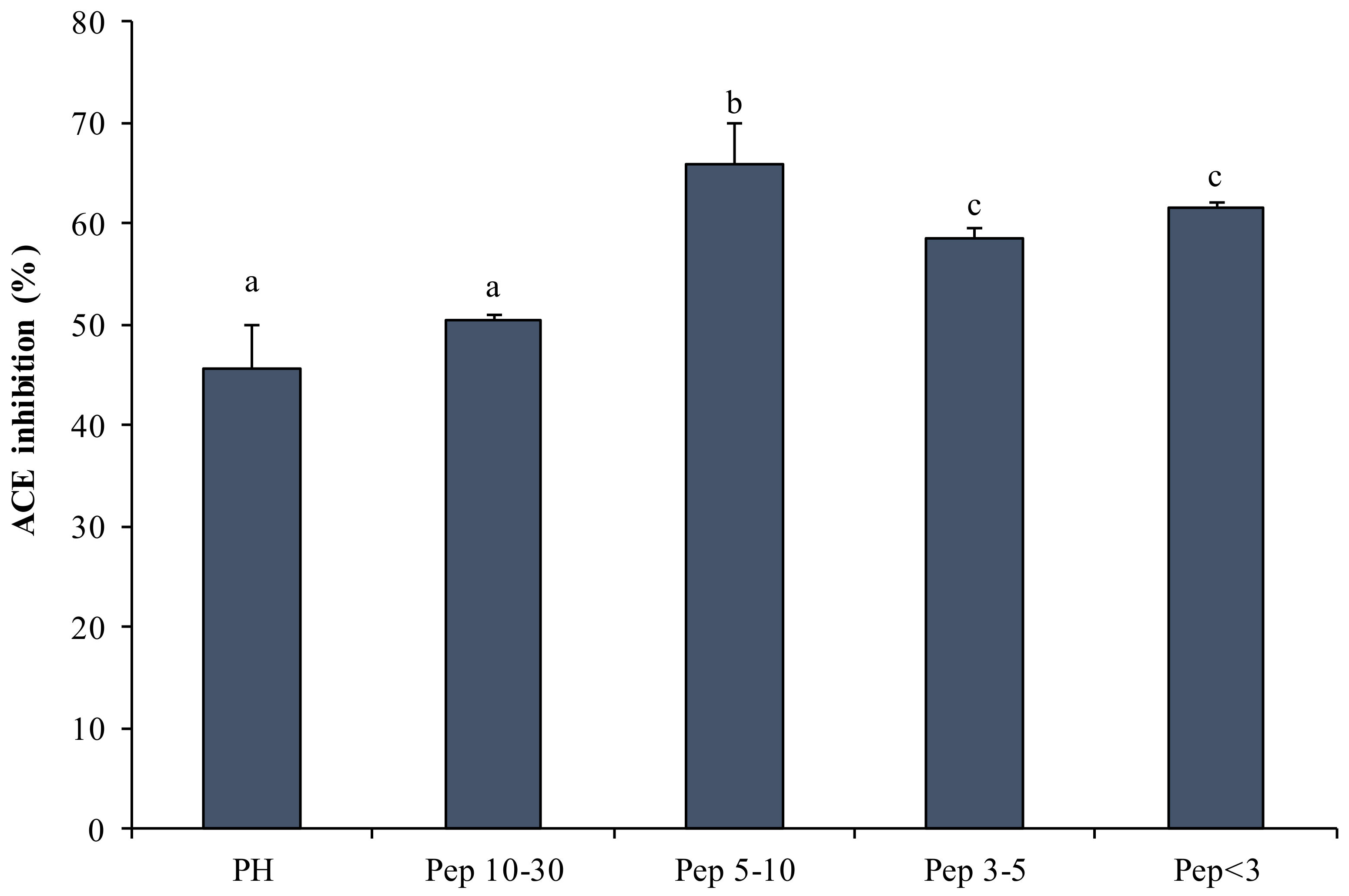
2.3.3. ACE-IA Determined by Protein Hydrolysates
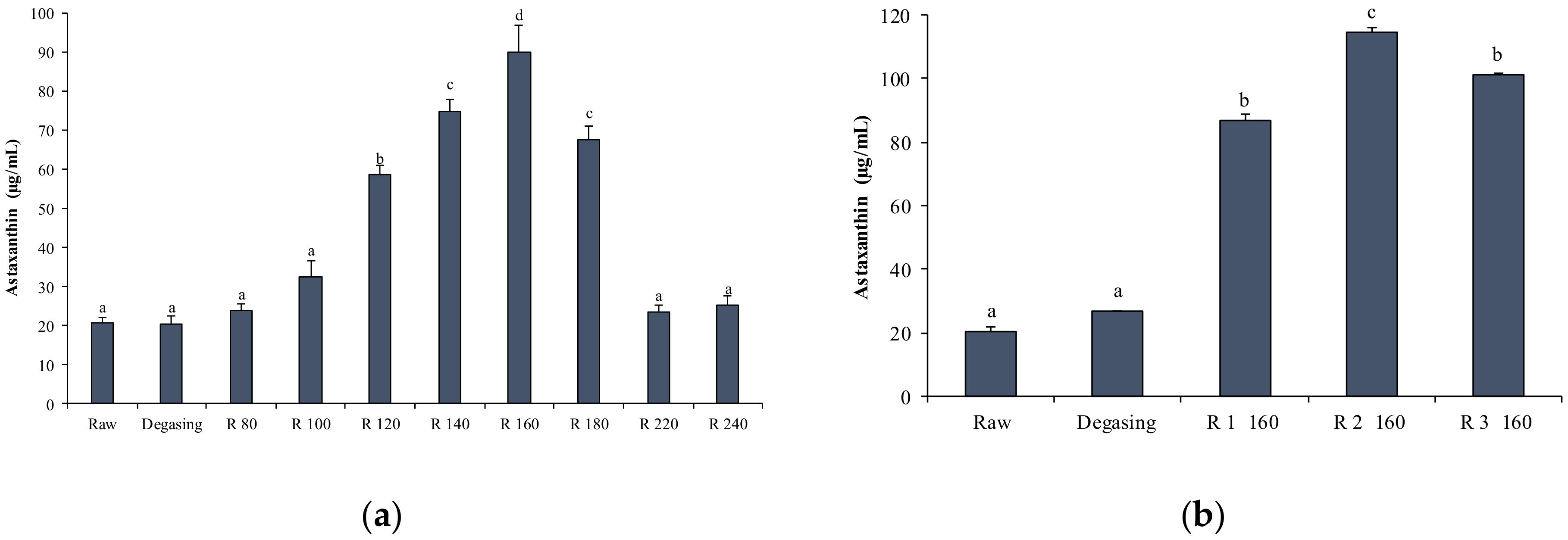
2.4. Extraction, Enrichment and Determination of AST
2.4.1. AST Yields Extracted with Crude Viscera Oil (CVO) and Ethyl Esters
2.4.2. Supercritical CO2 Extraction (SFE)
2.4.3. AST Enrichment by Short Path Distillation (SPD)
2.5. Evaluation of Bioactive Properties of Protein Hydrolysates and AST In Vitro
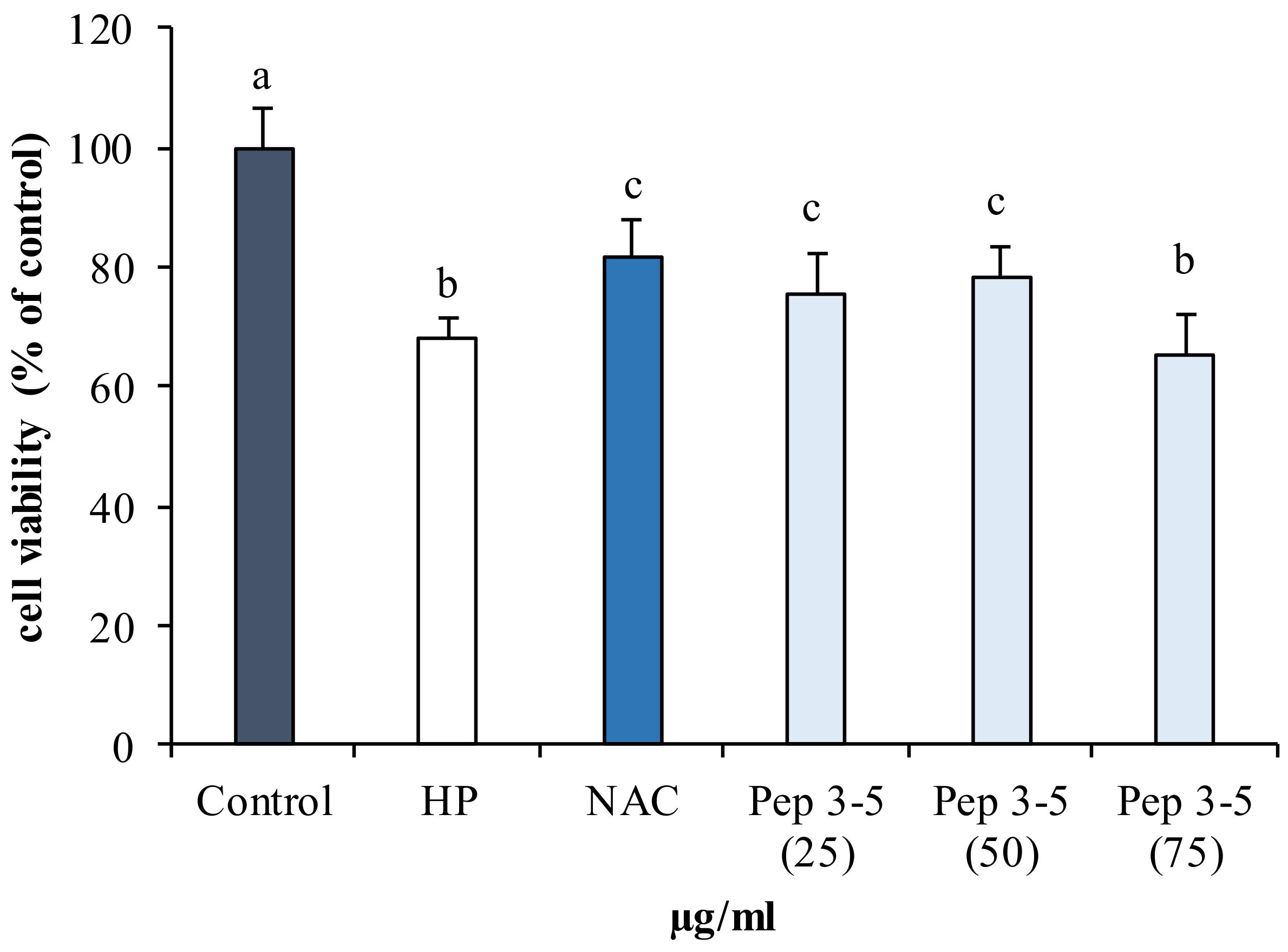
2.5.1. Antioxidant Activity of Hydrolyzed Fractions in Human Fibroblast (142BR)
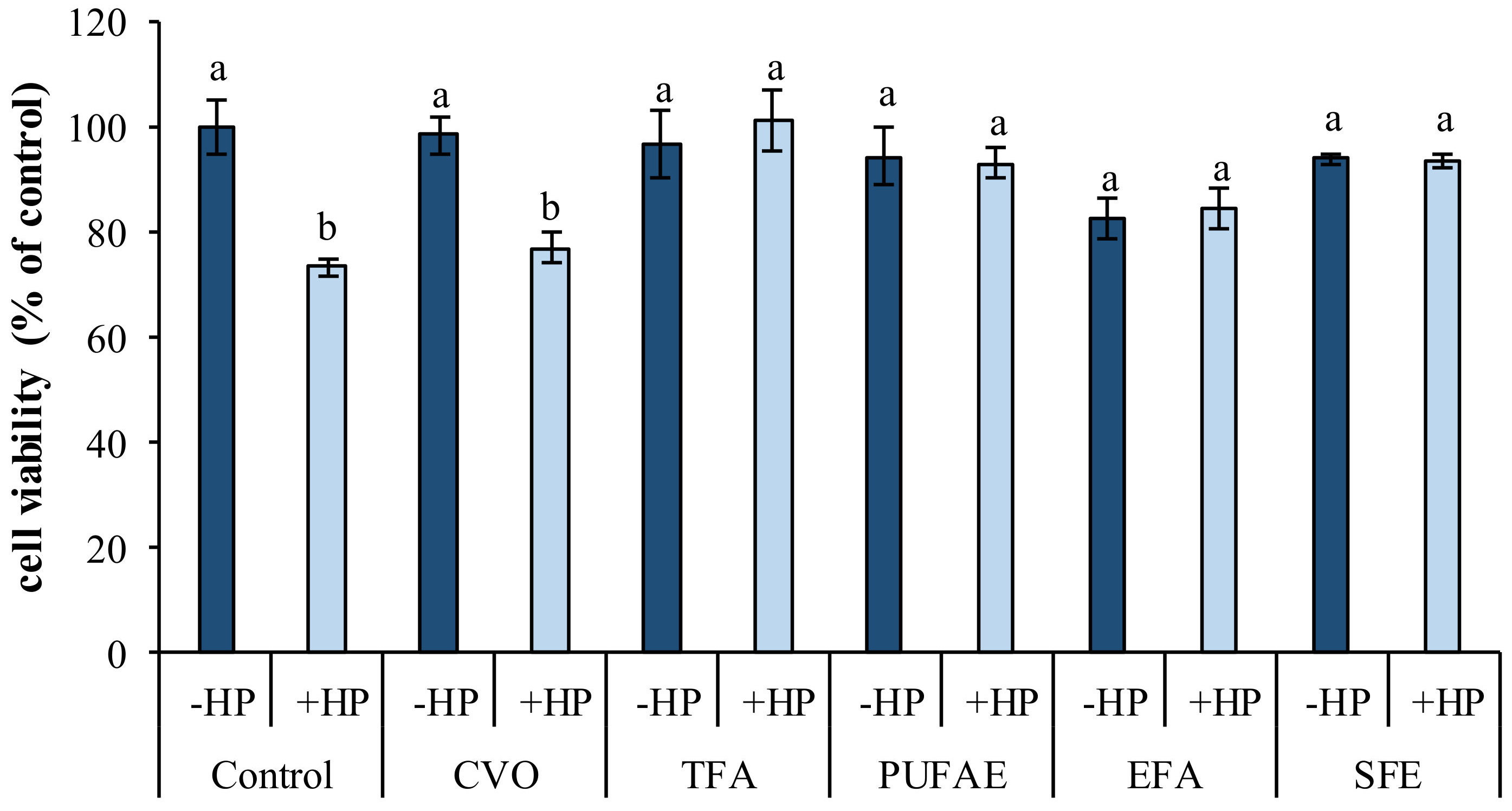
2.5.2. In Vitro Effects of AST Extracted Using Fish Oil, Ethyl Esters, and SFE
3. Materials and Methods
3.1. Sampling and Sample Processing
3.2. Evaluation of Proximate Composition and Fatty Acid Profile of P. longirostris by Products
3.3. Enzymatic Hydrolysis and AST Extraction
3.4. Separation of Protein Hydrolysates and AST
3.5. Protein Hydrolysates Characterization
3.5.1. SDS-PAGE
3.5.2. Fractionation of Proteins
3.5.3. DPPH Radical Scavenging Activity
3.5.4. ACE Inhibition Assay
3.6. AST Supercritical Fluid Extraction (SFE)
3.7. Enrichment of AST by Short Path Distillation (SPD)
3.8. Spectrophotometric Determination of AST
3.9. Cell Culture
3.9.1. Assessment of Antioxidant Activity of Pep in 142BR Cells
3.9.2. Protective Effect AST Extracted by CVO, TFA, PUFAE, EFA, and SFE in 3T3 Cells
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Caruso, G.; Floris, R.; Serangeli, C.; Di Paola, L. Fishery Wastes as a Yet Undiscovered Treasure from the Sea: Biomolecules Sources, Extraction Methods and Valorization. Mar. Drugs 2020, 18, 622. [Google Scholar] [CrossRef] [PubMed]
- Ucak, I.; Afreen, M.; Montesano, D.; Carrillo, C.; Tomasevic, I.; Simal-Gandara, J.; Barba, F.J. Functional and Bioactive Properties of Peptides Derived from Marine Side Streams. Mar. Drugs 2021, 19, 71. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Madrid, R.; Carballo-Uicab, V.M.; Cárdenas-Conejo, Y.; Aguilar-Espinosa, M.; Siva, R. Overview of carotenoids and beneficial effects on human health. In Carotenoids: Properties, Processing and Applications; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1–40. [Google Scholar]
- Giannaccare, G.; Pellegrini, M.; Senni, C.; Bernabei, F.; Scorcia, V.; Cicero, A.F.G. Clinical applications of astaxanthin in the treatment of ocular diseases: Emerging insights. Mar. Drugs 2020, 18, 239. [Google Scholar] [CrossRef] [PubMed]
- Fathalipour, M.; Fathalipour, H.; Safa, O.; Nowrouzi-Sohrabi, P.; Mirkhani, H.; Hassanipour, S. The therapeutic role of carotenoids in diabetic retinopathy: A systematic review. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 2347–2358. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, T.; Patel, K. Carotenoids: Potent to Prevent Diseases Review. Nat. Prod. Bioprospect. 2020, 10, 109–117. [Google Scholar] [CrossRef]
- Hussein, G.; Sankawa, U.; Goto, H.; Matsumoto, K.; Watanabe, H. Astaxanthin, a Carotenoid with Potential in Human Health and Nutrition. J. Nat. Prod. 2006, 69, 443–449. [Google Scholar] [CrossRef]
- Harnedy, P.A.; Fitzgerald, R.J. Bioactive proteins, peptides, and amino acids from macroalgae. J. Phycol. 2011, 47, 218–232. [Google Scholar] [CrossRef]
- Radzali, S.A.; Baharin, B.S.; Othman, R.; Markom, M.; Rahman, R.A. Co-solvent selection for supercritical fluid extraction of astaxanthin and other carotenoids from Penaeus monodon waste. J. Oleo Sci. 2014, 63, 769–777. [Google Scholar] [CrossRef] [Green Version]
- Sila, A.; Ghlissi, Z.; Kamoun, Z.; Makni, M.; Nasri, M.; Bougatef, A.; Sahnoun, Z. Astaxanthin from shrimp by-products ameliorates nephropathy in diabetic rats. Eur. J. Nutr. 2015, 54, 301–307. [Google Scholar] [CrossRef]
- Messina, C.M.; Manuguerra, S.; Renda, G.; Santulli, A. Biotechnological Applications for the Sustainable Use of Marine By-products: In Vitro Antioxidant and Pro-apoptotic Effects of Astaxanthin Extracted with Supercritical CO2 from Parapeneus longirostris. Mar. Biotechnol. 2019, 21, 565–576. [Google Scholar] [CrossRef]
- Guerin, M.; Huntley, M.E.; Olaizola, M. Haematococcus astaxanthin: Applications for human health and nutrition. Trends Biotechnol. 2003, 21, 210–216. [Google Scholar] [CrossRef]
- De Holanda, H.D.; Netto, F.M. Recovery of components from shrimp (Xiphopenaeus kroyeri) processing waste by enzymatic hydrolysis. J. Food Sci. 2006, 71, 1–6. [Google Scholar] [CrossRef]
- Baccouche, B.; Mbarek, S.; Dellaa, A.; Hammoum, I.; Messina, C.M.; Santulli, A.; Ben Chaouacha-Chekir, R. Protective Effect of Astaxanthin on Primary Retinal Cells of the Gerbil Psammomys Obesus Cultured in Diabetic Milieu. J. Food Biochem. 2017, 41, e12274. [Google Scholar] [CrossRef]
- Yeh, P.T.; Huang, H.W.; Yang, C.M.; Yang, W.S.; Yang, C.H. Astaxanthin inhibits expression of retinal oxidative stress and inflammatory mediators in streptozotocin-induced diabetic rats. PLoS ONE 2016, 11, e0146438. [Google Scholar] [CrossRef] [Green Version]
- Parjikolaei, B.R.; Errico, M.; El-Houri, R.B.; Mantell, C.; Fretté, X.C.; Christensen, K.V. Process design and economic evaluation of green extraction methods for recovery of astaxanthin from shrimp waste. Chem. Eng. Res. Des. 2017, 117, 73–82. [Google Scholar] [CrossRef]
- Chemat, F.; Vian, M.A.; Cravotto, G. Green extraction of natural products: Concept and principles. Int. J. Mol. Sci. 2012, 13, 8615–8627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yara-Varón, E.; Li, Y.; Balcells, M.; Canela-Garayoa, R.; Fabiano-Tixier, A.S.; Chemat, F. Vegetable oils as alternative solvents for green oleo-extraction, purification and formulation of food and natural products. Molecules 2017, 22, 1474. [Google Scholar] [CrossRef] [PubMed]
- Sachindra, N.M.; Mahendrakar, N.S. Process optimization for extraction of carotenoids from shrimp waste with vegetable oils. Bioresour. Technol. 2005, 96, 1195–1200. [Google Scholar] [CrossRef] [PubMed]
- Razi Parjikolaei, B.; Bahij El-Houri, R.; Fretté, X.C.; Christensen, K.V. Influence of green solvent extraction on carotenoid yield from shrimp (Pandalus borealis) processing waste. J. Food Eng. 2015, 155, 22–28. [Google Scholar] [CrossRef]
- Razi Parjikolaei, B.; Errico, M.; El-Houri, R.B.; Christensen, K.V.; Fretté, X.C. Green Approaches to Extract Astaxanthin from Shrimp Waste: Process Design and Economic Evaluation. Comput. Aided Chem. Eng. 2016, 38, 649–654. [Google Scholar]
- Shahidi, F.; Synowiecki, J. Isolation and Characterization of Nutrients and Value-Added Products from Snow Crab (Chinoecetes opilio) and Shrimp (Pandalus borealis) Processing Discards. J. Agrie. Food Chem. 1991, 39, 1527–1532. [Google Scholar] [CrossRef]
- Vian, M.; Breil, C.; Vernes, L.; Chaabani, E.; Chemat, F. Green solvents for sample preparation in analytical chemistry. Curr. Opin. Green Sustain. Chem. 2017, 5, 44–48. [Google Scholar] [CrossRef]
- Dátilo, M.N.; Sant’Ana, M.R.; Formigari, G.P.; Rodrigues, P.B.; de Moura, L.P.; da Silva, A.S.R.; Ropelle, E.R.; Pauli, J.R.; Cintra, D.E. Omega-3 from Flaxseed Oil Protects Obese Mice Against Diabetic Retinopathy Through GPR120 Receptor. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Ahmadkelayeh, S.; Hawboldt, K. Extraction of lipids and astaxanthin from crustacean by-products: A review on supercritical CO2 extraction. Trends Food Sci. Technol. 2020, 103, 94–108. [Google Scholar] [CrossRef]
- Al Khawli, F.; Pateiro, M.; Domínguez, R.; Lorenzo, J.M.; Gullón, P.; Kousoulaki, K.; Ferrer, E.; Berrada, H.; Barba, F.J. Innovative green technologies of intensification for valorization of seafood and their by-products. Mar. Drugs 2019, 17, 689. [Google Scholar] [CrossRef] [Green Version]
- Herrero, M.; Cifuentes, A.; Ibañez, E. Sub- and supercritical fluid extraction of functional ingredients from different natural sources: Plants, food-by-products, algae and microalgae—A review. Food Chem. 2006, 98, 136–148. [Google Scholar] [CrossRef] [Green Version]
- Herrero, M.; Mendiola, J.A.; Cifuentes, A.; Ibáñez, E. Supercritical fluid extraction: Recent advances and applications. J. Chromatogr. A 2010, 1217, 2495–2511. [Google Scholar] [CrossRef] [Green Version]
- Sila, A.; Nasri, M.; Bougatef, A. Isolation and characterisation of carotenoproteins from deep-water pink shrimp processing waste. Int. J. Biol. Macromol. 2012, 51, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.B.; Nguyen, D.N.; Nguyen, A.D.; Ngo, V.A. Utilization of Crab Waste for Cost-E ff ective. Mar. Drugs 2020, 18, 523. [Google Scholar] [CrossRef]
- Maschmeyer, T.; Luque, R.; Selva, M. Upgrading of marine (fish and crustaceans) biowaste for high added-value molecules and bio(nano)-materials. Chem. Soc. Rev. 2020, 49, 4527–4563. [Google Scholar] [CrossRef]
- Jafarpour, A.; Gregersen, S.; Marciel Gomes, R.; Marcatili, P.; Hegelund Olsen, T.; Jacobsen, C.; Overgaard, M.T.; Sørensen, A.-D.M. Biofunctionality of Enzymatically Derived Peptides from Codfish (Gadus morhua) Frame: Bulk In Vitro Properties, Quantitative Proteomics, and Bioinformatic Prediction. Mar. Drugs 2020, 18, 599. [Google Scholar] [CrossRef]
- Giannetto, A.; Esposito, E.; Lanza, M.; Oliva, S.; Riolo, K.; Di Pietro, S.; Abbate, J.M.; Briguglio, G.; Cassata, G.; Cicero, L.; et al. Protein Hydrolysates from Anchovy (Engraulis encrasicolus) Waste: In vitro and in vivo biological activities. Mar. Drugs 2020, 18, 86. [Google Scholar] [CrossRef] [PubMed]
- Doan, C.T.; Tran, T.N.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.L. Utilization of Seafood Processing By-Products for Production of Proteases by Paenibacillus sp. TKU052 and Their Application in Biopeptides’ Preparation. Mar. Drugs 2020, 18, 574. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Mendis, E. Bioactive compounds from marine processing byproducts—A review. Food Res. Int. 2006, 39, 383–393. [Google Scholar] [CrossRef]
- Othman, N.; Manan, Z.A.; Alwi, S.R.W.; Sarmidi, M.R. A review of extraction tech for carotenoid and vit E. Appl. Sci. 2010, 10, 1187–1191. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Camargo, A.P.; Meireles, M.Â.A.; Ferreira, A.L.K.; Saito, E.; Cabral, F.A. Extraction of ω-3 fatty acids and astaxanthin from Brazilian redspotted shrimp waste using supercritical CO2 + ethanol mixtures. J. Supercrit. Fluids 2012, 61, 71–77. [Google Scholar] [CrossRef]
- Arbia, W.; Adour, L.; Amrane, A.; Lounici, H. Optimization of medium composition for enhanced chitin extraction from Parapenaeus longirostris by Lactobacillus helveticus using response surface methodology. Food Hydrocoll. 2013, 31, 392–403. [Google Scholar] [CrossRef]
- Díaz-Rojas, E.I.; Argüelles-Monal, W.M.; Higuera-Ciapara, I.; Hernández, J.; Lizardi-Mendoza, J.; Goycoolea, F.M. Determination of Chitin and Protein Contents During the Isolation of Chitin from Shrimp Waste. Macromol. Biosci. 2006, 6, 340–347. [Google Scholar] [CrossRef]
- Routray, W.; Dave, D.; Ramakrishnan, V.V.; Murphy, W. Study of drying kinetics of salmon processing by-products at different temperatures and the quality of extracted fish oil. Dry. Technol. 2017, 35, 1981–1993. [Google Scholar] [CrossRef]
- Yves, H.; Korma, S.A.; Ali, A.H.; Tuyishime, M.A.; Habinshuti, I.; Abed, S.M. Extraction, Refining and Purification of ω-3 PUFA through Different Techniques—A Review. Am. J. Food Sci. Nutr. Res. 2016, 4, 18. [Google Scholar]
- Fox, C.J.; Blow, P.; Brown, J.H.; Watson, I. The effect of various processing methods on the physical and biochemical properties of shrimp head meals and their utilization by juvenile Penaeus monodon Fab. Aquaculture 1994, 122, 209–226. [Google Scholar] [CrossRef]
- Dumay, J.; Donnay-Moreno, C.; Barnathan, G.; Jaouen, P.; Bergé, J.P. Improvement of lipid and phospholipid recoveries from sardine (Sardina pilchardus) viscera using industrial proteases. Process Biochem. 2006, 41, 2327–2332. [Google Scholar] [CrossRef] [Green Version]
- Tkaczewska, J.; Borawska-Dziadkiewicz, J.; Kulawik, P.; Duda, I.; Morawska, M.; Mickowska, B. The effects of hydrolysis condition on the antioxidant activity of protein hydrolysate from Cyprinus carpio skin gelatin. LWT 2020, 117, 108616. [Google Scholar] [CrossRef]
- Sabeena Farvin, K.H.; Andersen, L.L.; Nielsen, H.H.; Jacobsen, C.; Jakobsen, G.; Johansson, I.; Jessen, F. Antioxidant activity of Cod (Gadus morhua) protein hydrolysates: In vitro assays and evaluation in 5% fish oil-in-water emulsion. Food Chem. 2014, 149, 326–334. [Google Scholar] [CrossRef]
- Taheri, A.; Farvin, K.H.S.; Jacobsen, C.; Baron, C.P. Antioxidant activities and functional properties of protein and peptide fractions isolated from salted herring brine. Food Chem. 2014, 142, 318–326. [Google Scholar] [CrossRef]
- Picot, L.; Ravallec, R.; Martine, F.P.; Vandanjon, L.; Jaouen, P.; Chaplain-Derouiniot, M.; Guérard, F.; Chabeaud, A.; Legal, Y.; Alvarez, O.M.; et al. Impact of ultrafiltration and nanofiltration of an industrial fish protein hydrolysate on its bioactive properties. J. Sci. Food Agric. 2010, 90, 1819–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krichen, F.; Sila, A.; Caron, J.; Kobbi, S.; Nedjar, N.; Miled, N.; Blecker, C.; Besbes, S.; Bougatef, A. Identification and molecular docking of novel ACE inhibitory peptides from protein hydrolysates of shrimp waste. Eng. Life Sci. 2018, 18, 682–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.M.; Meyers, S.P. Effect of Antioxidants on Stability of Astaxanthin Pigment in Crawfish Waste and Oil Extract. J. Agric. Food Chem. 1982, 30, 469–473. [Google Scholar] [CrossRef]
- Voda, A.; Homan, N.; Witek, M.; Duijster, A.; van Dalen, G.; van der Sman, R.; Nijsse, J.; van Vliet, L.; Van As, H.; van Duynhoven, J. The impact of freeze-drying on microstructure and rehydration properties of carrot. Food Res. Int. 2012, 49, 687–693. [Google Scholar] [CrossRef]
- Balaban, M.O.; Marshall, M.R.; Cornell, J.A.; Charest, D.J. Astaxanthin Extraction from Crawfish Shells by Supercritical CO2 with Ethanol as Cosolvent. J. Aquat. Food Prod. Technol. 2001, 10, 81–96. [Google Scholar]
- Batistella, C.B.; Moraes, E.B.; Maciel Filho, R.; Wolf Maciel, M.R. Molecular distillation process for recovering biodiesel and carotenoids from palm oil. Appl. Biochem. Biotechnol. Part A Enzym. Eng. Biotechnol. 2002, 98–100, 1149–1159. [Google Scholar] [CrossRef]
- Qian, Z.J.; Jung, W.K.; Byun, H.G.; Kim, S.K. Protective effect of an antioxidative peptide purified from gastrointestinal digests of oyster, Crassostrea gigas against free radical induced DNA damage. Bioresour. Technol. 2008, 99, 3365–3371. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Jimenez, G.-M.; Burgos-Hernandez, A.; Ezquerra-Brauer, J.-M. Bioactive Peptides and Depsipeptides with Anticancer Potential: Sources from Marine Animals. Mar. Drugs 2012, 10, 963–986. [Google Scholar] [CrossRef]
- Castelli, V.; Melani, F.; Ferri, C.; d’Angelo, M.; Catanesi, M.; Grassi, D.; Benedetti, E.; Giordano, A.; Cimini, A.; Desideri, G. Neuroprotective activities of bacopa, lycopene, astaxanthin, and vitamin B12 combination on oxidative stress-dependent neuronal death. J. Cell. Biochem. 2020, 121, 4862–4869. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis: Changes in Official Methods of Analysis Made at the Annual Meeting; AOAC International: Gaithersburg, MD, USA, 1990; Volume 15, ISBN 9780935584752. [Google Scholar]
- AOAC. Association of Official Analytical Chemists Official Method, 981.10 Crude protein in meat block digestion method. J. AOAC Int. 1992, 65, 1339. [Google Scholar]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Lepage, G.; Roy, C.C. Improved recovery of fatty acid through direct transesterification without prior extraction or purification. J. Lipid Res. 1984, 25, 1391–1396. [Google Scholar] [CrossRef]
- Messina, C.M.; Renda, G.; La Barbera, L.; Santulli, A. By-products of farmed European sea bass (Dicentrarchus labrax L.) as a potential source of n-3 PUFA. Biol. 2013, 68, 288–293. [Google Scholar] [CrossRef] [Green Version]
- Adler-Nissen, J. Limited enzymic degradation of proteins: A new approach in the industrial application of hydrolases. J. Chem. Technol. Biotechnol. 1982, 32, 138–156. [Google Scholar] [CrossRef]
- Utne-Palm, A.C.; Bogevik, A.S.; Humborstad, O.B.; Aspevik, T.; Pennington, M.; Løkkeborg, S. Feeding response of Atlantic cod (Gadus morhua) to attractants made from by-products from the fishing industry. Fish. Res. 2020, 227, 105535. [Google Scholar] [CrossRef]
- Messina, C.M.; Arena, R.; Renda, G.; La Barbera, L.; Manuguerra, S.; Santulli, A. Valorization of farmed gilthead seabream (Sparus aurata) by-products for application in nutraceutic and sustainability promotion: Utilization of short-path distillation for ω-3 enriched-oil production and evaluation of its bioactivity in-vitro. Mar. Drugs Drugs 2021, 19, 160. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Messina, C.M.; Gaglio, R.; Morghese, M.; Tolone, M.; Arena, R.; Moschetti, G.; Santulli, A.; Francesca, N.; Settanni, L. Microbiological profile and bioactive properties of insect powders used in food and feed formulations. Foods 2019, 8, 400. [Google Scholar] [CrossRef] [Green Version]
- Messina, C.M.; Troia, A.; Arena, R.; Manuguerra, S.; Ioannou, T.; Curcuraci, E.; Renda, G.; Hellio, C.; Santulli, A. Species-specific antioxidant power and bioactive properties of the extracts obtained from wild mediterranean Calendula Spp. (Asteraceae). Appl. Sci. 2019, 9, 4627. [Google Scholar] [CrossRef] [Green Version]
- Bernatoniene, J.; Masteikova, R.; Davalgiene, J.; Peciura, R.; Gauryliene, R.; Bernatoniene, R.; Majiene, D.; Lazauskas, R.; Civinskiene, G.; Velziene, S.; et al. Topical application of Calendula officinalis (L.): Formulation and evaluation of hydrophilic cream with antioxidant activity. J. Med. Plants Res. 2011, 5, 868–877. [Google Scholar]
- Arena, R.; Manuguerra, S.; Collins, E.; Mahdhi, A.; Renda, G.; Messina, C.M.; Santulli, A. Antioxidant properties of a supercritical fluid extract of the halophyte Mesembryanthemum nodiflorum L. from sicilian coasts: Nutraceutical and cosmeceutical applications. Appl. Sci. 2020, 10, 2374. [Google Scholar] [CrossRef] [Green Version]
- Cushman, D.W.; Cheung, H.S. Spectrophotometric assay and properties of the angiotensin-converting enzyme of rabbit lung. Biochem. Pharmacol. 1971, 20, 1637–1648. [Google Scholar] [CrossRef]
- Qian, Z.J.; Jung, W.K.; Lee, S.H.; Byun, H.G.; Kim, S.K. Antihypertensive effect of an angiotensin I-converting enzyme inhibitory peptide from bullfrog (Rana catesbeiana Shaw) muscle protein in spontaneously hypertensive rats. Process Biochem. 2007, 42, 1443–1448. [Google Scholar] [CrossRef]
- Mazzelli, A.; Luzzi, D.M.; Buonanno, G.; Cicci, A.; Piemonte, V.; Iaquaniello, G. An optimized separation process of microalgal lipidic products by molecular distillation: Techno-economic analysis. Chem. Eng. Sci. 2019, 207, 1187–1195. [Google Scholar] [CrossRef]
- Chen, H.M.; Meyers, S.P. A rapid quantitative method for determination of astaxanthin pigment concentration in oil extracts. J. Am. Oil Chem. Soc. 1984, 61, 1045–1047. [Google Scholar] [CrossRef]
- Abbes, M.; Baati, H.; Guermazi, S.; Messina, C.; Santulli, A.; Gharsallah, N.; Ammar, E. Biological properties of carotenoids extracted from Halobacterium halobium isolated from a Tunisian solar saltern. BMC Complement. Altern. Med. 2013, 13, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhouibi, N.; Manuguerra, S.; Arena, R.; Mahdhi, A.; Messina, C.M.; Santulli, A.; Dhaouadi, H. Screening of Antioxidant Potentials and Bioactive Properties of the Extracts Obtained from Two Centaurea L. Species (C. kroumirensis Coss. and C. sicula L. subsp sicula). Appl. Sci. 2020, 10, 2267. [Google Scholar] [CrossRef] [Green Version]
- Manuguerra, S.; Caccamo, L.; Mancuso, M.; Arena, R.; Rappazzo, A.C.; Genovese, L.; Santulli, A.; Messina, C.M.; Maricchiolo, G. The antioxidant power of horseradish, Armoracia rusticana, underlies antimicrobial and antiradical effects, exerted in vitro. Nat. Prod. Res. 2018, 34, 1567–1570. [Google Scholar] [CrossRef]
- Messina, C.M.; Manuguerra, S.; Catalano, G.; Arena, R.; Cocchi, M.; Morghese, M.; Montenegro, L.; Santulli, A. Green biotechnology for valorisation of residual biomasses in nutraceutic sector: Characterization and extraction of bioactive compounds from grape pomace and evaluation of the protective effects in vitro. Nat. Prod. Res. 2019, 35, 331–336. [Google Scholar] [CrossRef]
- Denizot, F.; Lang, R. Rapid colorimetric assay for cell growth and survival. Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J. Immunol. Methods 1986, 89, 271–277. [Google Scholar] [CrossRef]
- Ortega, Á.L. Oxidative Stress in Diabetic Retinopathy. Antioxidants 2021, 10, 50. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| g/100 g | |
|---|---|
| Moisture | 3.43 ± 0.16 |
| Ash | 36.40 ± 0.61 |
| Lipid | 4.96 ± 0.17 |
| Protein | 38.47 ± 0.46 |
| Chitin | 16.75 ± 1.06 |
| WBP | DBP | |
|---|---|---|
| 14:0 | 1.47 ± 0.06 | 0.83 ± 0.08 |
| 16:0 | 17.67 ± 0.20 | 13.10 ± 0.61 |
| 16:1n-7 | 3.40 ± 0.22 | 2.46 ± 0.13 |
| 16:2n-4 | 0.38 ± 0.05 | 0.29 ± 0.03 |
| 16:3n-4 | 0.70 ± 0.06 | 0.67 ± 0.04 |
| 18:0 | 6.61 ± 0.18 | 6.02 ± 0.13 |
| 18:1n-9 | 18.25 ± 1.91 | 22.84 ± 0.55 |
| 18:1n-7 | 4.03 ± 0.18 | 3.81 ± 0.16 |
| 18:2n-6 | 4.00 ± 1.67 | 15.24 ± 0.51 |
| 18:3n-4 | 0.23 ± 0.05 | 0.19 ± 0.03 |
| 18:3n-3 | 0.52 ± 0.04 | 0.36 ± 0.04 |
| 18:4n-3 | 0.29 ± 0.05 | 0.16 ± 0.02 |
| 20:1n-9 | 1.54 ± 0.07 | 1.34 ± 0.04 |
| 20:4n-6 | 4.68 ± 0.39 | 4.32 ± 0.33 |
| 20:4n-3 | 0.38 ± 0.05 | 0.23 ± 0.06 |
| 20:5n-3 (EPA) | 11.97 ± 1.07 | 9.69 ± 0.43 |
| 22:1n-11 | 0.68 ± 0.06 | 0.53 ± 0.04 |
| 22:1n-9 | 0.31 ± 0.08 | 0.26 ± 0.06 |
| 22:5n-3 | 1.26 ± 0.27 | 1.04 ± 0.14 |
| 22:6n-3 (DHA) | 21.66 ± 1.73 | 16.45 ± 0.90 |
| Saturated | 25.74 ± 0.31 | 19.95 ± 0.74 |
| Monounsaturated | 28.21 ± 1.72 | 31.24 ± 0.45 |
| Tot n-3 | 36.08 ± 2.94 | 27.92 ± 1.47 |
| Tot n-6 | 8.67 ± 1.34 | 19.56 ± 0.51 |
| DHA/EPA | 1.81 ± 0.04 | 1.70 ± 0.05 |
| ER | WBP | DBP | p < 0.05 | |
|---|---|---|---|---|
| CVO | 0.5 | 80.21 ± 2.0 e | 52.56 ± 0.74 e | * |
| TFA | 86.14 ± 1.88 f | 47.81 ± 3.16 d | * | |
| PUFAE | 31.78 ± 4.19 a | 31.89 ± 1.18 b | - | |
| EFA | 27.17 ± 3.54 a | 19.02 ± 3.74 a | * | |
| CVO | 1.0 | 97.99 ± 1.30 g | 64.22 ± 2.05 g | * |
| TFA | 105.23 ± 3.15 h | 58.41 ± 2.88 f | * | |
| PUFAE | 38.83 ± 6.81 b | 38.96 ± 5.54 c | - | |
| EFA | 33.20 ± 2.30 a,b | 23.23 ± 4.08 a | * | |
| CVO | 2.0 | 149.06 ± 0.82 i | 97.68 ± 1.51 h | * |
| TFA | 160.06 ± 8.91 l | 88.85 ± 7.34 h | * | |
| PUFAE | 59.06 ± 4.06 d | 59.26 ± 3.78 f,g | - | |
| EFA | 50.50 ± 0.91 c | 35.34 ± 1.60 b,c | * |
| Degassing | Distillation | |
|---|---|---|
| Flow (htz) | 20 | 5 |
| T (°C) Feed | 40 | 40 |
| T (°C) Condenser | 25 | 60 |
| T (°C) Residue | 60 | 60 |
| T (°C) Evaporator | 80 | 80–240 |
| Vacuum (mbar) | 5 | 2 × 10−3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Messina, C.M.; Manuguerra, S.; Arena, R.; Renda, G.; Ficano, G.; Randazzo, M.; Fricano, S.; Sadok, S.; Santulli, A. In Vitro Bioactivity of Astaxanthin and Peptides from Hydrolisates of Shrimp (Parapenaeus longirostris) By-Products: From the Extraction Process to Biological Effect Evaluation, as Pilot Actions for the Strategy “From Waste to Profit”. Mar. Drugs 2021, 19, 216. https://doi.org/10.3390/md19040216
Messina CM, Manuguerra S, Arena R, Renda G, Ficano G, Randazzo M, Fricano S, Sadok S, Santulli A. In Vitro Bioactivity of Astaxanthin and Peptides from Hydrolisates of Shrimp (Parapenaeus longirostris) By-Products: From the Extraction Process to Biological Effect Evaluation, as Pilot Actions for the Strategy “From Waste to Profit”. Marine Drugs. 2021; 19(4):216. https://doi.org/10.3390/md19040216
Chicago/Turabian StyleMessina, Concetta Maria, Simona Manuguerra, Rosaria Arena, Giuseppe Renda, Giovanna Ficano, Mariano Randazzo, Stefano Fricano, Saloua Sadok, and Andrea Santulli. 2021. "In Vitro Bioactivity of Astaxanthin and Peptides from Hydrolisates of Shrimp (Parapenaeus longirostris) By-Products: From the Extraction Process to Biological Effect Evaluation, as Pilot Actions for the Strategy “From Waste to Profit”" Marine Drugs 19, no. 4: 216. https://doi.org/10.3390/md19040216