
Characterization of ACE Inhibitory Peptides Prepared from Pyropia pseudolinearis Protein

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
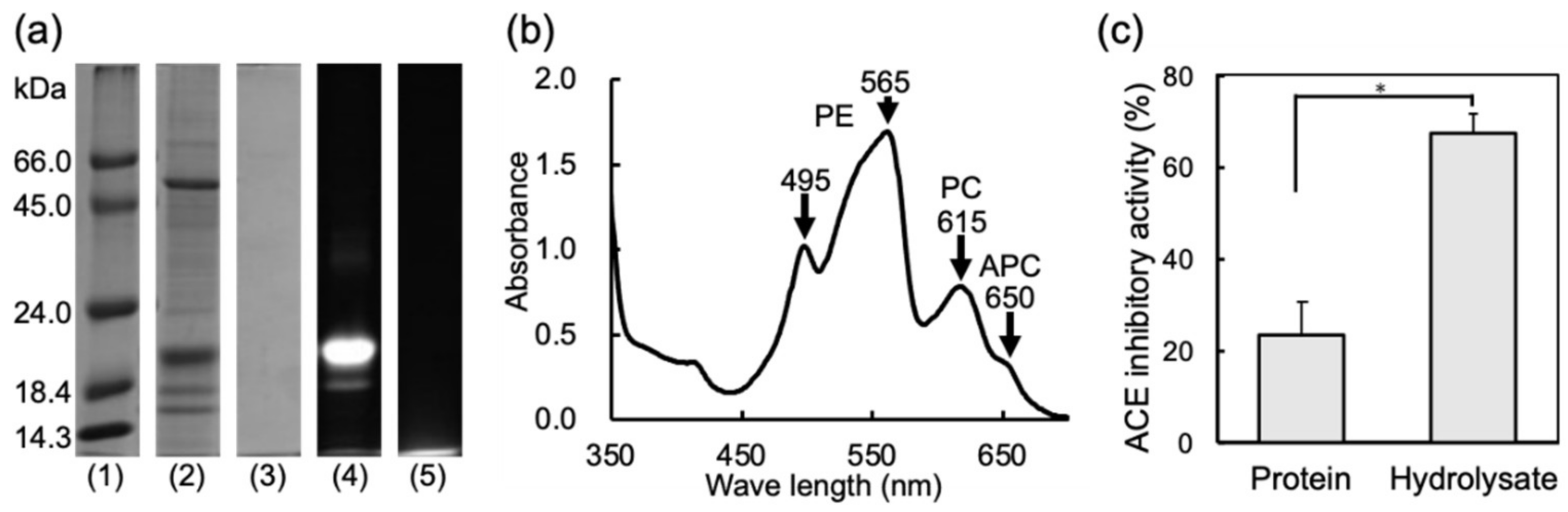
2.1. Properties and ACE Inhibitory Activities of Water Soluble Protein (WSP) and the Thermolysin Hydrolysate of the WSP
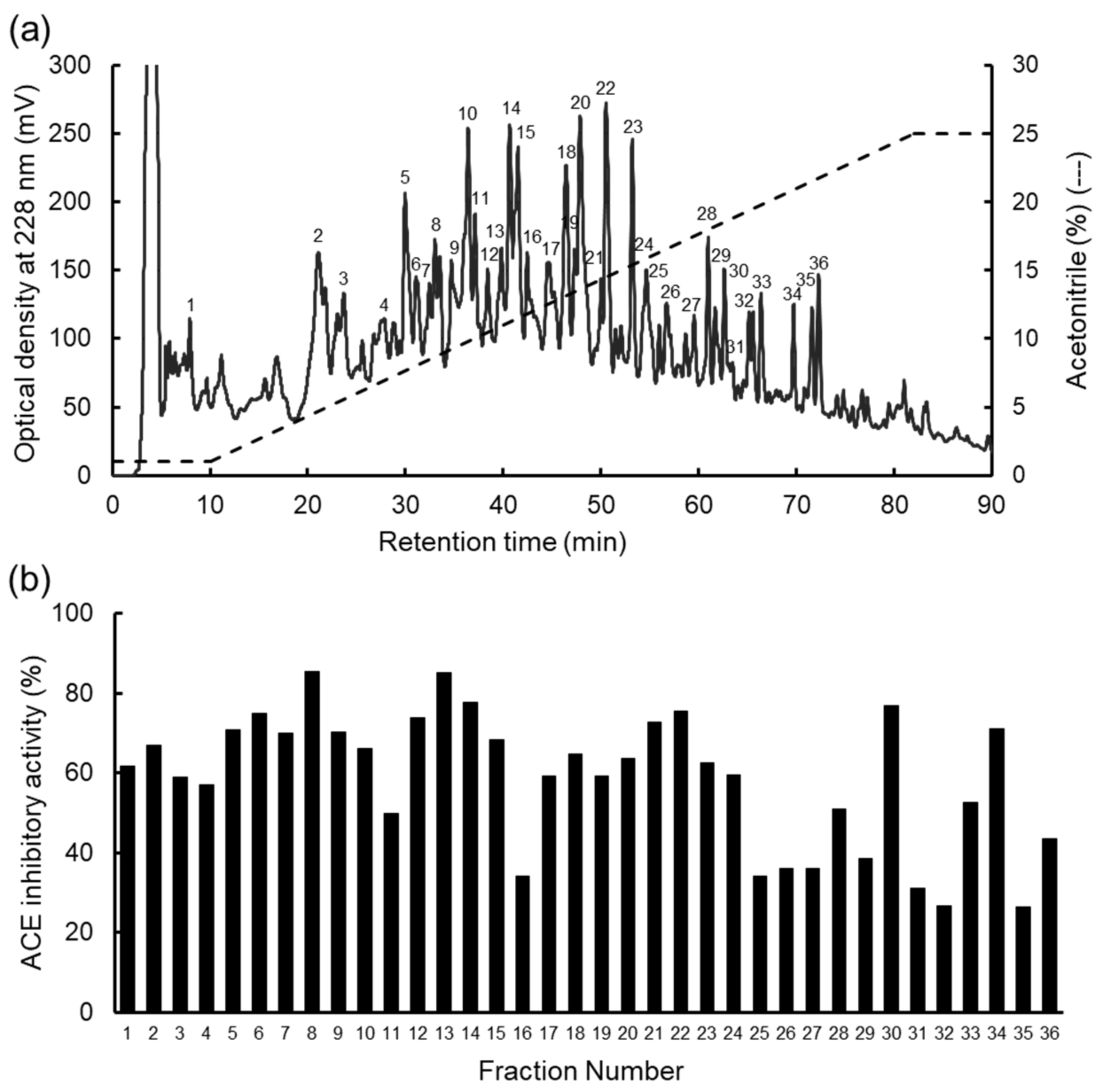
2.2. Fractionation of the Hydrolysate of the WSP
2.3. Identification of Peptides from the WSP
2.4. Sequencing and Comparison of Phycobiliproteins
2.5. Identification of Novel ACE Inhibitory Peptides from Red Algae
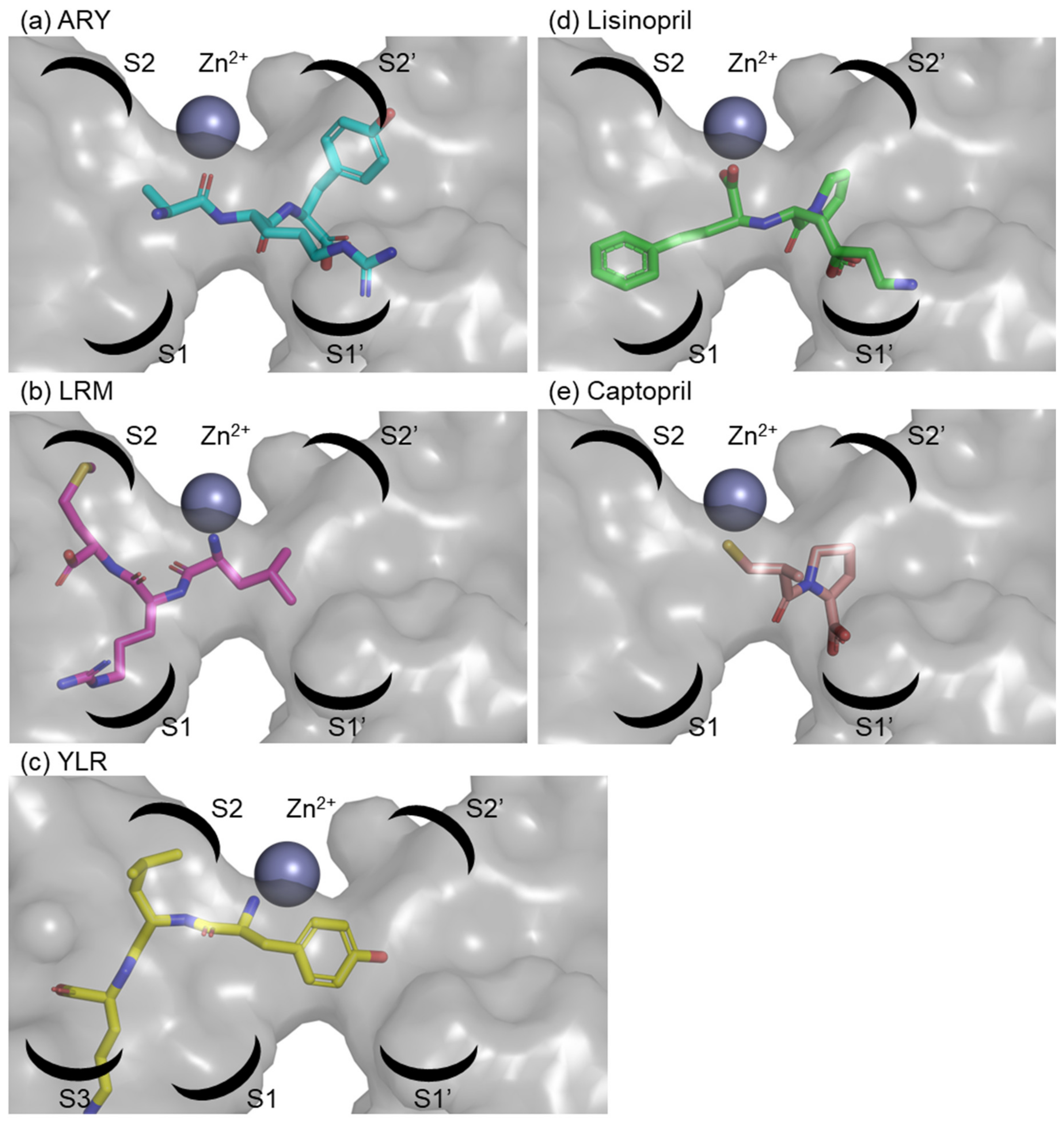
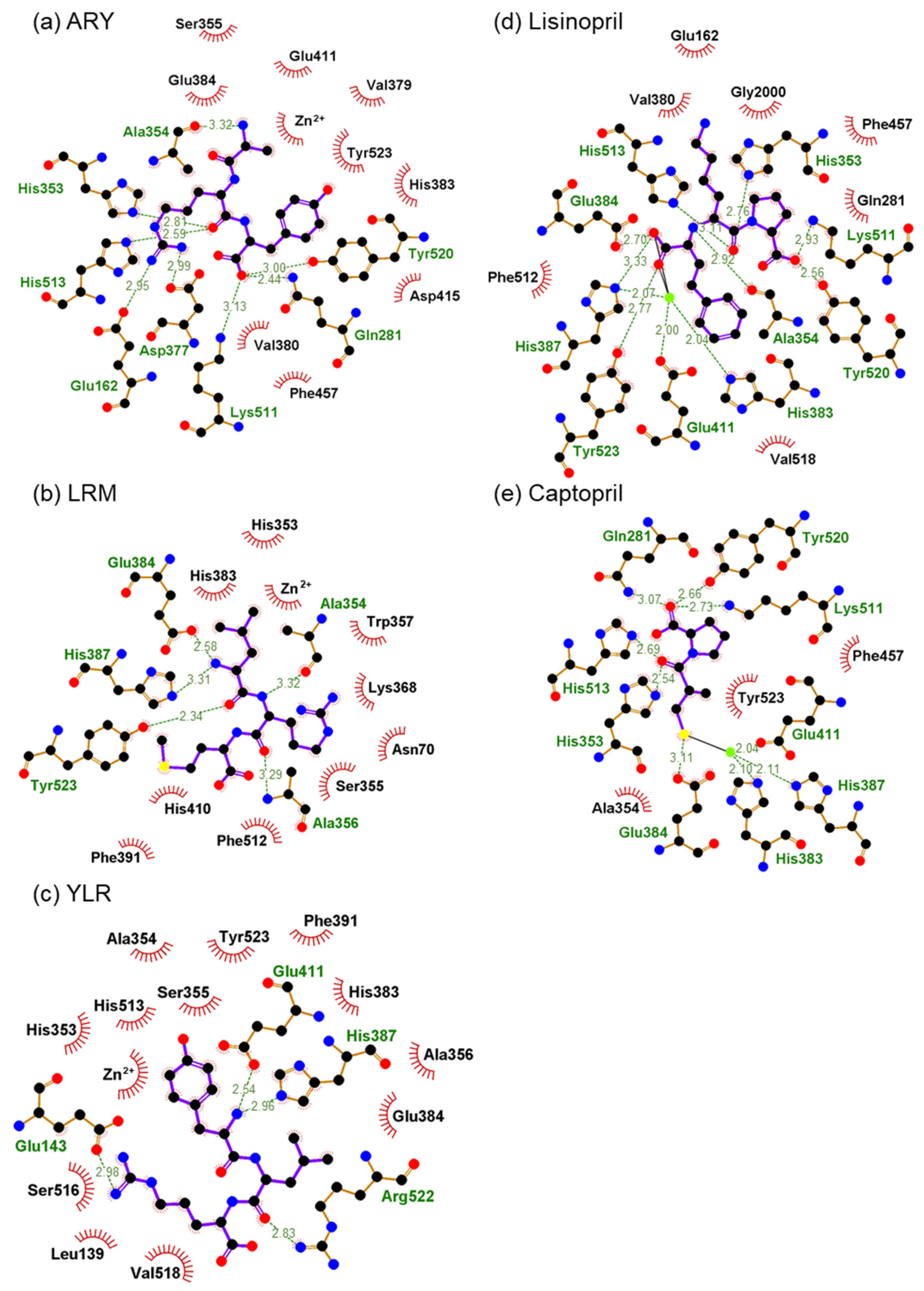
2.6. In Silico Analysis of Binding of Peptides and ACE
3. Materials and Methods
3.1. Materials
3.2. Preparation of the WSP and Its Hydrolysate
3.3. Separation of WSP Hydrolysate and Determination of Peptide Structures
3.4. Assay of ACE Inhibitory Activity
3.5. Extraction and Sequencing of Chloroplast DNA
3.6. Bioinformatics Analysis of ACE Inhibitory Peptides
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tu, M.; Cheng, S.; Lu, W.; Du, M. Advancement and prospects of bioinformatics analysis for studying bioactive peptides from food-derived protein: Sequence, structure, and functions. Trac. Trend. Anal. Chem. 2018, 105, 7–17. [Google Scholar] [CrossRef]
- Matsufuji, H.; Matsui, T.; Seki, E.; Osajima, K.; Nakashima, M.; Osajima, Y. Angiotensin I-converting enzyme inhibitory peptides in an alkaline protease hydrolyzate derived from sardine muscle. Biosci. Biotechnol. Biochem. 1994, 58, 2244–2245. [Google Scholar] [CrossRef] [PubMed]
- Nakano, D.; Ogura, K.; Miyakoshi, M.; Ishii, F.; Kawanishi, H.; Kurumazuka, D.; Kwak, C.-J.; Ikemura, K.; Takaoka, M.; Moriguchi, S.; et al. Antihypertensive effect of angiotensin I-converting enzyme inhibitory peptides from a sesame protein hydrolysate in spontaneously hypertensive rats. Biosci. Biotechnol. Biochem. 2006, 70, 1118–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daskaya-Dikmen, C.; Yucetepe, A.; Karbancioglu-Guler, F.; Daskaya, H.; Ozcelik, B. Angiotensin-I-converting enzyme (ACE)-inhibitory peptides from plants. Nutrients 2017, 9, 316. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, F.D.; Whelton, P.K. High Blood Pressure and Cardiovascular Disease. Hypertension 2020, 75, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Schmieder, R.E.; Hilgers, K.F.; Schlaich, M.P.; Schmidt, B.M.W. Reninangiotensin system and cardiovascular risk. Lancet 2007, 369, 1208–1219. [Google Scholar] [CrossRef]
- Umemura, S.; Arima, H.; Arima, S.; Asayama, K.; Dohi, Y.; Hirooka, Y.; Horio, T.; Hoshide, S.; Ikeda, S.; Ishimitsu, T.; et al. The Japanese Society of Hypertension Guidelines for the Management of Hypertension (JSH 2019). Hypertens. Res. 2019, 42, 1235–1481. [Google Scholar]
- Wijesekara, I.; Pangestuti, R.; Kim, S.-K. Biological activities and potential health benefits of sulfated polysaccharides derived from marine algae. Carbohydr. Polym. 2011, 84, 14–21. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Kishimura, H.; Kinoshita, Y.; Saburi, W.; Kumagai, Y.; Yasui, H.; Ojima, T. Enzymatic production of xylooligosaccharides from red alga dulse (Palmaria sp.) wasted in Japan. Process. Biochem. 2019, 82, 117–122. [Google Scholar] [CrossRef]
- Chen, H.; Yan, X.; Zhu, P.; Lin, J. Antioxidant activity and hepatoprotective potential of agaro-oligosaccharides in vitro and in vivo. Nutr. J. 2006, 5, 31. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Kumagai, Y.; Yamamoto, Y.; Yasui, H.; Kishimura, H. Identification of a key enzyme for the hydrolysis of β-(1→3)-xylosyl linkage in red alga dulse xylooligosaccharide from Bifidobacterium adolescentis. Mar. Drugs 2020, 18, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Ruiter, G.A.; Rudolph, B. Carrageenan biotechnology. Trends Food Sci. Technol. 1997, 8, 389–395. [Google Scholar] [CrossRef]
- Cherry, P.; Yadav, S.; Strain, C.R.; Allsopp, P.J.; McSorley, E.M.; Ross, R.P.; Stanton, C. Prebiotics from seaweeds: An ocean of opportunity? Mar. Drugs 2019, 17, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurpilhares, D.D.B.; Cinelli, L.P.; Simas, N.K.; Pessoa, A., Jr.; Sette, L.D. Marine prebiotics: Polysaccharides and oligosaccharides obtained by using microbial enzymes. Food Chem. 2019, 280, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Roleda, M.Y.; Marfaing, H.; Desnica, N.; Jónsdóttir, R.; Skjermo, J.; Rebours, C.; Nitschke, U. Variations in polyphenol and heavy metal contents of wild-harvested and cultivated seaweed bulk biomass: Health risk assessment and implication for food applications. Food Control 2019, 95, 121–134. [Google Scholar] [CrossRef]
- Rosa, G.P.; Tavares, W.R.; Sousa, P.M.C.; Pagès, A.K.; Seca, A.M.L.; Pinto, D.C.G.A. Seaweed secondary metabolites with beneficial health effects: An overview of successes in in vivo studies and clinical trials. Mar. Drugs 2020, 18, 8. [Google Scholar] [CrossRef] [Green Version]
- Kendel, M.; Couzinet-Mossion, A.; Viau, M.; Fleurence, J.; Barnathan, G.; Wielgosz-Collin, G. Seasonal composition of lipids, fatty acids, and sterols in the edible red alga Grateloupia turuturu. J. Appl. Phycol. 2013, 25, 425–432. [Google Scholar] [CrossRef]
- Schubert, N.; García-Mendoza, E.; Pacheco-Ruiz, I. Carotenoid composition of marine red algae 1. J. Phycol. 2006, 42, 1208–1216. [Google Scholar] [CrossRef]
- Aryee, A.N.A.; Agyei, D.; Akanbi, T.O. Recovery and utilization of seaweed pigments in food processing. Curr. Opin. Food Sci. 2018, 19, 113–119. [Google Scholar] [CrossRef]
- Harnedy, P.A.; FitzGerald, R.J. Bioactive proteins, peptides, and amino acid from macroalgae. J. Appl. Phycol. 2020, 47, 218–232. [Google Scholar] [CrossRef] [PubMed]
- Mai, K.; Mercer, J.P.; Donlon, J. Comparative studies on the nutrition of two species of abalone, Haliotis tuberculata L. and Haliotis discus hannai Ino: II. Amino acid composition of abalone and six species of macroalgae with an assessment of their nutritional value. Aquaculture 1994, 128, 115–130. [Google Scholar] [CrossRef]
- Paiano, M.O.; Cortona, A.D.; Costa, J.F.; Liu, S.-L.; Verbruggen, H.; Clerck, O.D.; Necchi, O., Jr. Organization of plastid genomes in the freshwater red algal order Batrachospermales (Rhodphyta). J. Phycol. 2018, 54, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Yang, E.C.; Boo, S.M.; Bhattacharya, D.; Saunders, G.W.; Knoll, A.H.; Fredericq, S.; Graf, L.; Yoon, H.S. Divergence time estimates and the evolution of major lineages in the florideophyte red algae. Sci. Rep. 2016, 6, 21361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glazer, A.N. Light harvesting by phycobilisomes. Ann. Rev. Biophys. Chem. 1985, 14, 47–77. [Google Scholar] [CrossRef] [PubMed]
- Miyabe, Y.; Furuta, T.; Takeda, T.; Kanno, G.; Shimizu, T.; Tanaka, Y.; Gai, Z.; Yasui, H.; Kishimura, H. Structural properties of phycoerythrin from dulse Palmaria palmata. J. Food Biochem. 2017, 41, e12301. [Google Scholar] [CrossRef] [Green Version]
- Kitade, Y.; Miyabe, Y.; Yamamoto, Y.; Takeda, H.; Shimizu, T.; Yasui, H.; Kishimura, H. Structural characteristics of phycobiliproteins from red alga Mazzaella japonica. J. Food Biochem. 2018, 42, e12436. [Google Scholar] [CrossRef] [Green Version]
- Sfriso, A.A.; Gallo, M.; Baldi, F. Phycoerythrin productivity and diversity from five red macroalgae. J. Appl. Phycol. 2018, 30, 2523–2531. [Google Scholar] [CrossRef]
- Wijesekara, I.; Kim, S.-K. Angiotensin-I-converting enzyme (ACE) inhibitors from marine resources: Prospects in the pharmaceutical industry. Mar. Drugs 2010, 8, 1080–1093. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Xu, X.; Sun, X.; Zhang, X.; Chen, X.; Xu, N. Preparation and identification of ACE inhibitory peptides from the marine macroalga Ulva intestinalis. Mar. Drugs 2019, 17, 179. [Google Scholar] [CrossRef] [Green Version]
- Noda, H. Health benefits and nutritional properties of nori. J. Appl. Phycol. 1993, 5, 255–258. [Google Scholar] [CrossRef]
- He, H.-L.; Chen, X.-L.; Wu, H.; Sun, C.-Y.; Zhang, Y.-Z.; Zhou, B.-C. High throughput and rapid screening of marine protein hydrolysates enriched in peptides with angiotensin-I-converting enzyme inhibitory activity by capillary electrophoresis. Bioresour. Technol. 2007, 98, 3499–3505. [Google Scholar] [CrossRef]
- Cian, R.E.; Alaiz, M.; Vioque, J.; Drago, S.R. Enzyme proteolysis enhanced extraction of ACE inhibitory and antioxidant compounds (peptides and polyphenols) from Porphyra columbina residual cake. J. Appl. Phycol. 2013, 25, 1197–1206. [Google Scholar] [CrossRef]
- Suetsuna, K. Purification and identification of angiotensin I converting enzyme inhibitors from the red alga Porphyra yezoensis. J. Mar. Biotechnol. 1998, 6, 163–167. [Google Scholar]
- Furuta, T.; Miyabe, Y.; Yasui, H.; Kinoshita, Y.; Kishimura, H. Angiotensin I converting enzyme inhibitory peptides derived from phycobiliproteins of dulse Palmaria palmata. Mar. Drugs 2016, 14, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumagai, Y.; Kitade, Y.; Kobayashi, M.; Watanabe, K.; Kurita, H.; Takeda, H.; Yasui, H.; Kishimura, H. Identification of ACE inhibitory peptides from red alga Mazzaella japonica. Eur. Food Res. Technol. 2020, 246, 2225–2231. [Google Scholar] [CrossRef]
- Fitzgerald, C.; Aluko, R.E.; Hossain, M.; Rai, D.K.; Hayes, M. Potential of a renin inhibitory peptide from the red seaweed Palmaria palmata as a functional food ingredient following confirmation and characterization of a hypotensive effect in spontaneously hypertensive rats. J. Agric. Food Chem. 2014, 62, 8352–8356. [Google Scholar] [CrossRef]
- Harnedy, P.A.; O’Keeffe, M.B.; FitzGerald, R.J. Purification and identification of dipeptidyl peptidase (DPP) IV inhibitory peptides from the macroalga Palmaria palmata. Food Chem. 2015, 172, 400–406. [Google Scholar] [CrossRef]
- Fitzgerald, C.; Gallagher, E.; Tasdemir, D.; Hayes, M. Heart health peptides from macroalgae and their potential use in functional foods. J. Agric. Food Chem. 2011, 59, 6829–6836. [Google Scholar] [CrossRef]
- Sonani, R.R.; Singh, N.K.; Kumar, J.; Thakar, D.; Madamwar, D. Concurrent purification and antioxidant activity of phycobiliproteins from Lyngbya sp. A09DM: An antioxidant and anti-aging potential of phycoerythrin in Caenorhabditis elegans. Process. Biochem. 2014, 49, 1757–1766. [Google Scholar] [CrossRef]
- Sato, N.; Furuta, T.; Takeda, T.; Miyabe, Y.; Ura, K.; Takagi, Y.; Yasui, H.; Kumagai, Y.; Kishimura, H. Antioxidant activity of proteins extracted from red alga dulse harvested in Japan. J. Food Biochem. 2019, 43, e12709. [Google Scholar] [CrossRef]
- Sakai, S.; Komura, Y.; Nishimura, Y.; Sugawara, T.; Hirata, T. Inhibition of mast cell degranulation by phycoerythrin and its pigment moiety phycoerythrobilin, prepared from Porphyra yezoensis. Food Sci. Technol. Res. 2011, 17, 171–177. [Google Scholar] [CrossRef] [Green Version]
- Cian, R.E.; Garzon, A.G.; Ancona, D.B.; Guerrero, L.C.; Drago, S.R. Hydrolyzates from Pyropia columbina seaweed have antiplatelet aggregation, antioxidant and ACE I inhibitory peptides which maintain bioactivity after simulated gastrointestinal digestion. LWT Food Sci. Technol. 2015, 64, 881–888. [Google Scholar] [CrossRef]
- Sato, M.; Hosokawa, T.; Yamaguchi, T.; Nakano, T.; Muramoto, K.; Kahara, T.; Funayama, K.; Kobayashi, A.; Nakano, T. Angiotensin I-converting enzyme inhibitory peptides derived from Wakame (Undaria pinnatifida) and their antihypertensive effect in spontaneously hypertensive rats. J. Agric. Food Chem. 2002, 50, 6245–6252. [Google Scholar] [CrossRef]
- Suetsuna, K.; Maekawa, K.; Chen, J.-R. Antihypertensive effects of Undaria pinnatifida (wakame) peptide on blood pressure in spontaneously hypertensive rats. J. Nutr. Biochem. 2004, 15, 267–272. [Google Scholar] [CrossRef]
- Kumagai, Y.; Miyabe, Y.; Takeda, T.; Adachi, K.; Yasui, H.; Kishimura, H. In silico analysis of relationship between proteins from plastid genome of red alga Palmaria sp. (Japan) and angiotensin I converting enzyme inhibitory peptides. Mar. Drugs 2019, 17, 190. [Google Scholar] [CrossRef] [Green Version]
- Sumikawa, K.; Takei, K.; Kumagai, Y.; Shimizu, T.; Yasui, H.; Kishimura, H. In silico analysis of ACE inhibitory peptides from chloroplast proteins of red alga Grateloupia asiatica. Mar. Biotechnol. 2020, 22, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Ng, P.K.; Lin, S.M.; Lim, P.E.; Liu, L.C.; Chen, C.M.; Pai, T.W. Complete chloroplast genome of Gracilaria firma (Gracilariaceae, Rhodophyta), with discussion on the use of chloroplast phylogenomics in the subclass Rhodymeniophycidae. BMC Genom. 2017, 18, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Kim, K.M.; Yang, E.C.; Miller, K.A.; Boo, S.M.; Bhattacharya, D.; Yoon, H.S. Reconstructing the complex evolutionary history of mobile plasmids in red algal genomes. Sci. Rep. 2016, 6, 23744. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.-E.; Deng, Y.-Y.; Xu, G.-P.; Russell, S.; Lu, Q.-Q.; Brodie, J. Redefining Pyropia (Bangiales, Rhodophyta): Four new genera, resurrection of Porphyrella and description of Calidia pseudolobata sp. nov. from China. J. Phycol. 2020, 56, 862–879. [Google Scholar] [CrossRef] [PubMed]
- Saunders, G.W.; Hommersand, M.H. Assessing red algal supraordinal diversity and taxonomy in the context of contemporary systematic data. Am. J. Bot. 2004, 91, 1494–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Aluko, R.E.; Nakai, S. Structural requirements of angiotensin I-converting enzyme inhibitory peptides: Quantitative structure−activity relationship study of di- and tripeptides. J. Agric. Food Chem. 2006, 54, 732–738. [Google Scholar] [CrossRef]
- Baldwin, J.E.; Adlington, R.M.; Russell, A.T.; Smith, M.L. Carbon based nucleophilic ring opening of activated monocyclic β-lactams; Synthesis and stereochemical assignment of the ACE inhibitor WF-10129. Tetrahedron 1995, 51, 4733–4762. [Google Scholar] [CrossRef]
- Hooper, N.M. Angiotensin converting enzyme: Implications from molecular biology for its physiological functions. Int. J. Biochem. 1991, 23, 641–647. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Cota-Sánchez, J.H.; Remarchuk, K.; Ubayasena, K. Ready-to-use DNA extracted with a CTAB method adapted for herbarium specimens and mucilaginous plant tissue. Plant Mol. Biol. Rep. 2006, 24, 161. [Google Scholar] [CrossRef]
- Kumagai, Y.; Tsubouchi, R.; Miyabe, Y.; Takeda, T.; Adachi, K.; Yasui, H.; Kishimura, H. Complete sequence of mitochondrial DNA of red alga dulse Palmaria palmata (Linnaeus) Weber & Mohr in Japan. Mitochondrial DNA Part B 2019, 4, 3177–3178. [Google Scholar] [PubMed]
- Watanabe, K.; Kishimoto, T.; Kumagai, Y.; Shimizu, T.; Uji, T.; Yasui, H.; Kishimura, H. Complete sequence of mitochondrial DNA of Gloiopeltis furcata (Postels and Ruprecht) J. Agardh. Mitochondrial DNA Part B 2019, 4, 2543–2544. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Jin, B.; Li, H.; Huang, S.-Y. HPEPDOCK: A web server for blind peptide–protein docking based on a hierarchical algorithm. Nucleic Acids Res. 2018, 46, W443–W450. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FN | Peptide | Number of Peptides in Chloroplast Proteins | In Silico Thermolysin Digestion | |
|---|---|---|---|---|
| ◯ a | × b | |||
| 3 | AYR | 6 | rbcL | apcE, atpA, cpcG, psaL, rps7 |
| 4 | MTFF | 0 | - c | - |
| 5 | VRFK d | 1 | gltB | - |
| 6 | KFR | 4 | - | accA, cemA, chlI, rpl16 |
| 6 | WHKPA | 0 | - | - |
| 6 | FFKWEF | 0 | - | - |
| 7 | FGGR | 1 | petJ | - |
| 7 | LVER | 0 | - | - |
| 8 | YRD | 7 | - | cpeA, moeB, odpA, orf148, rps14, ycf3, ycf21 |
| 8 | FFR d | 4 | psaB, ycf22 | psbT, ycf63 |
| 9 | ARY | 6 | atpI, chlB, gltB | apcA, orf263, ycf46 |
| 9 | RFR | 3 | - | odpA, rpoC1 (2) e |
| 10 | FAR d | 7 | clpC, orf174, ycf26 | atpI, chlB, cpeA, ilvH |
| 10 | YLR | 12 | - | accA, apcA, apcB, apcD, apcE, apcF, carA, cpcA, orf114, rpl3, rpl19, rps9 |
| 10 | VYRT | 1 | - | cpeA |
| 10 | FVCG | 0 | - | - |
| 11 | FFLREF | 0 | - | - |
| 12 | SRGL | 1 | - | rpl23 |
| 13 | ACWR | 0 | - | - |
| 13 | RFAPR | 0 | - | - |
| 14 | ACPSGT | 0 | - | - |
| 16 | WER | 1 | - | psbA |
| 17 | LDY | 17 | chlN, cpcA, cpeA, infB, rbcL, ycf16 | accA, apcA, apcB, atpA, infC, orf111, orf565, rpoA, rps1, syh, ycf24 |
| 17 | LLEER | 0 | - | - |
| 18 | PGCRR | 0 | - | - |
| 18 | FLWWLR | 0 | - | - |
| 20 | AAGRFP | 1 | - | cpeA |
| 23 | LVFFGH | 0 | - | - |
| 24 | PVAFN | 0 | - | - |
| 25 | LRM d | 2 | cpcA | psbB |
| 26 | LRY | 8 | apcB, apcE, apcF, cpcB, cpeB | apcD, apcE, rbcR |
| 27 | CPSNN | 0 | - | - |
| 27 | AWRRP | 0 | - | - |
| 27 | LWT | 4 | trpG | psaB, psbD, psbZ |
| 28 | YRF | 3 | - | cpcG, odpA, syh |
| 28 | FRV d | 8 | petB, rbcL, rps5, rpoB (2), ycf26 | pgmA, rpoB |
| 29 | VNLF | 1 | - | ycf38 |
| 29 | PGDTY | 0 | - | - |
| 30 | EWYPH | 0 | - | - |
| 32 | KTFPY | 0 | - | - |
| 34 | FGRPF d | 1 | rbcL | - |
| 36 | VESR | 2 | - | rbcL, rpoA |
| Total | 101 peptides in 66 kinds of proteins (31 peptides were produced by in silico digestion) | |||
| Species | Accession No. | Identity (%) | |||||
|---|---|---|---|---|---|---|---|
| cpeA a | cpeB a | cpcA b | cpcB b | apcA c | apcB c | ||
| Pyropia pulchra | NC_029861.1 | 100 | 100 | 100 | 98.8 | 99.4 | 100 |
| Porphyra purpurea | NC_000925 | 100 | 99.4 | 99.4 | 99.4 | 100 | 100 |
| Neopyropia yezoensis | KC51072 | 100 | 100 | 100 | 98.3 | 100 | 100 |
| Palmaria palmata in Japan | AB807662 | 89.0 | 88.1 | 88.9 | 90.1 | 95.0 | 98.1 |
| Grateloupia asiatica | AP018129 | 90.2 | 91.5 | 92.6 | 90.1 | 96.3 | 93.8 |
| Peptide | IC50 (μmol) | Reference |
|---|---|---|
| ARY | 1.3 | This study |
| YLR | 5.8 | |
| LRM | 0.15 | |
| VYRT | 0.14 | [35] |
| LDY | 6.1 | |
| FEQWAS | >2.8 | |
| LRY | 0.044 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumagai, Y.; Toji, K.; Katsukura, S.; Morikawa, R.; Uji, T.; Yasui, H.; Shimizu, T.; Kishimura, H. Characterization of ACE Inhibitory Peptides Prepared from Pyropia pseudolinearis Protein. Mar. Drugs 2021, 19, 200. https://doi.org/10.3390/md19040200
Kumagai Y, Toji K, Katsukura S, Morikawa R, Uji T, Yasui H, Shimizu T, Kishimura H. Characterization of ACE Inhibitory Peptides Prepared from Pyropia pseudolinearis Protein. Marine Drugs. 2021; 19(4):200. https://doi.org/10.3390/md19040200
Chicago/Turabian StyleKumagai, Yuya, Keigo Toji, Satoshi Katsukura, Rie Morikawa, Toshiki Uji, Hajime Yasui, Takeshi Shimizu, and Hideki Kishimura. 2021. "Characterization of ACE Inhibitory Peptides Prepared from Pyropia pseudolinearis Protein" Marine Drugs 19, no. 4: 200. https://doi.org/10.3390/md19040200