
EPS364, a Novel Deep-Sea Bacterial Exopolysaccharide, Inhibits Liver Cancer Cell Growth and Adhesion


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
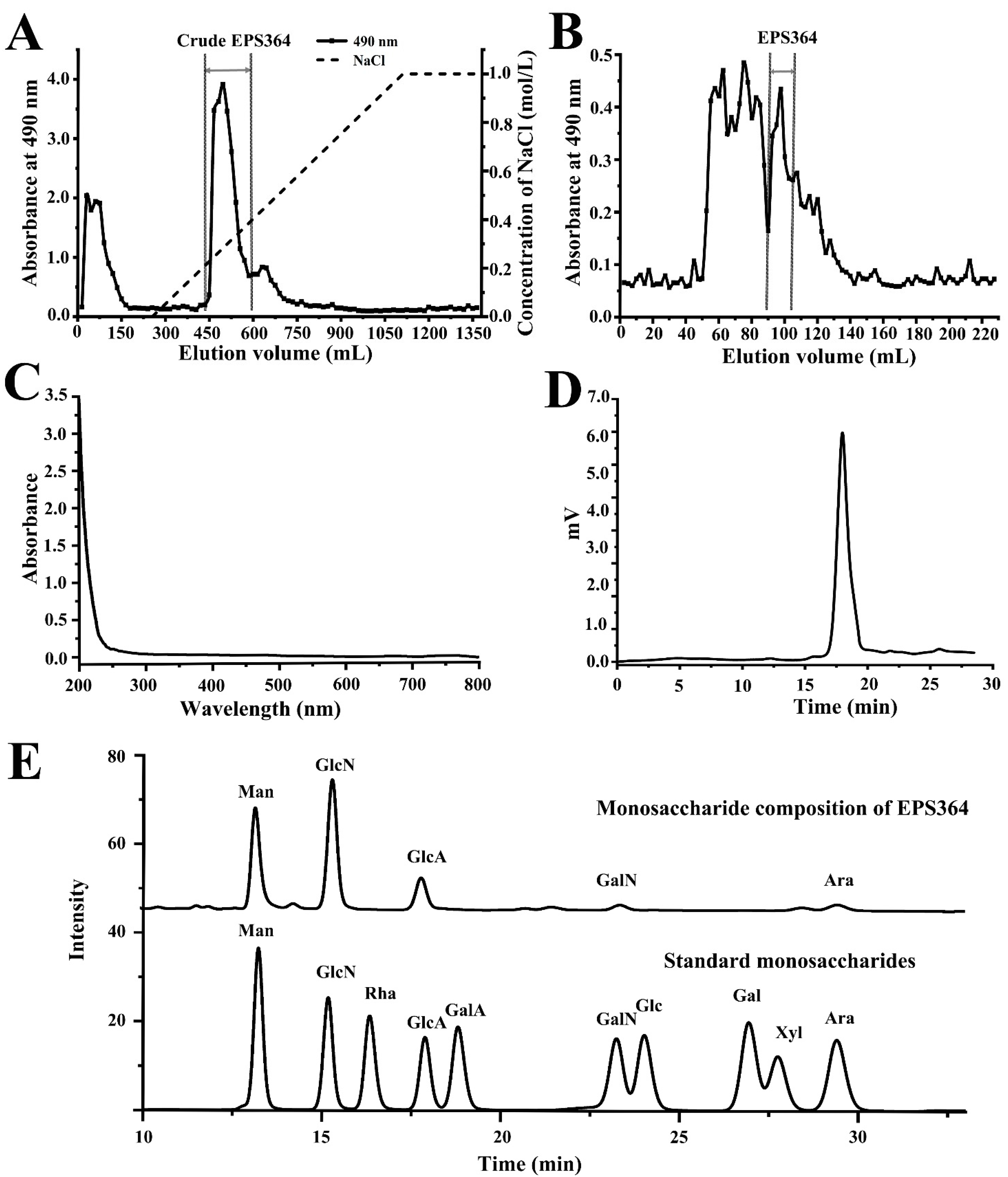
2.1. Isolation, Extraction and Purification of Exopolysaccharide EPS364
2.2. Characterization of EPS364
2.2.1. Molecular Weight (Mw) and Monosaccharide Composition of EPS364
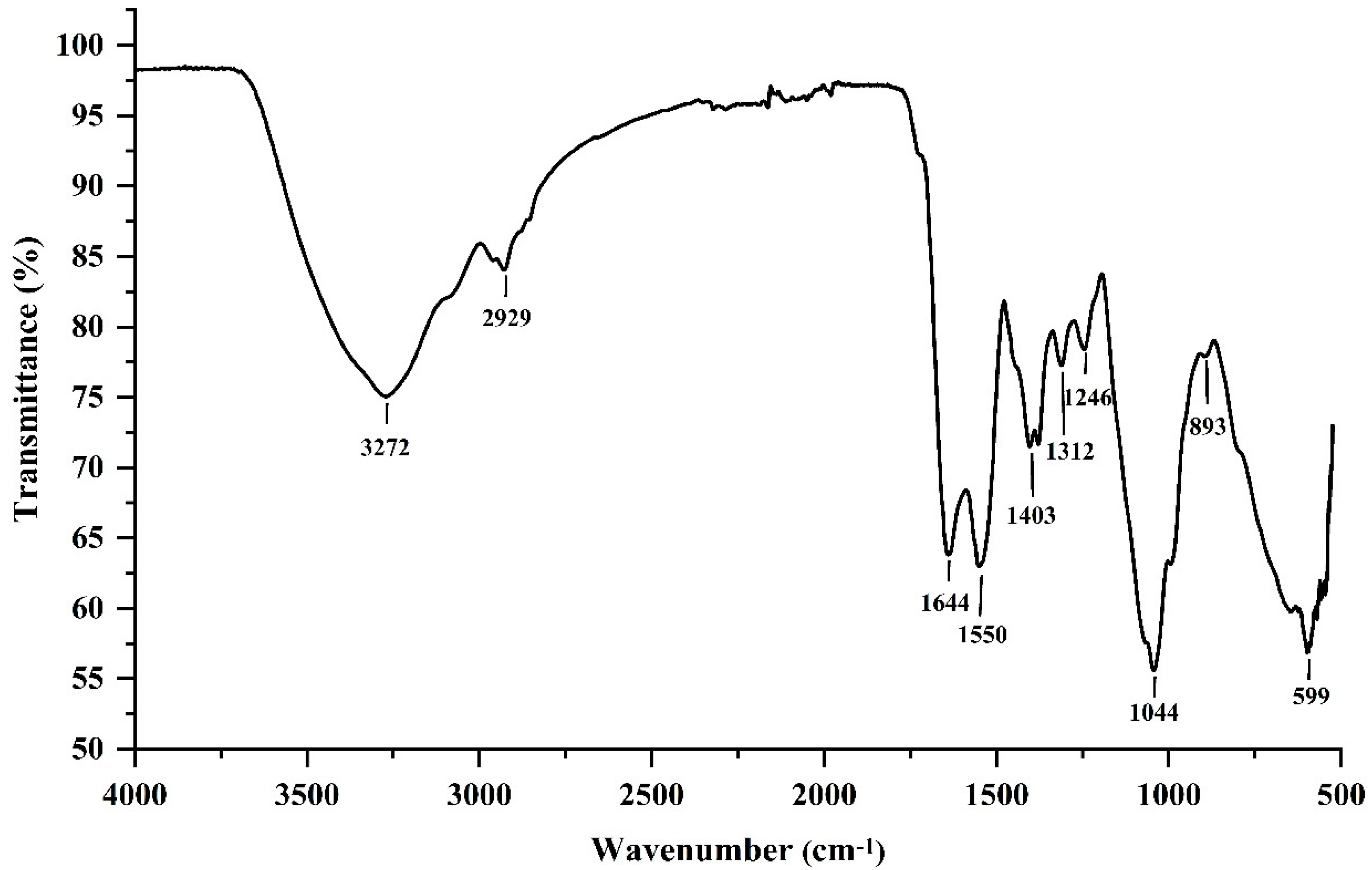
2.2.2. Fourier-Transform Infrared (FTIR) Spectrum Analysis
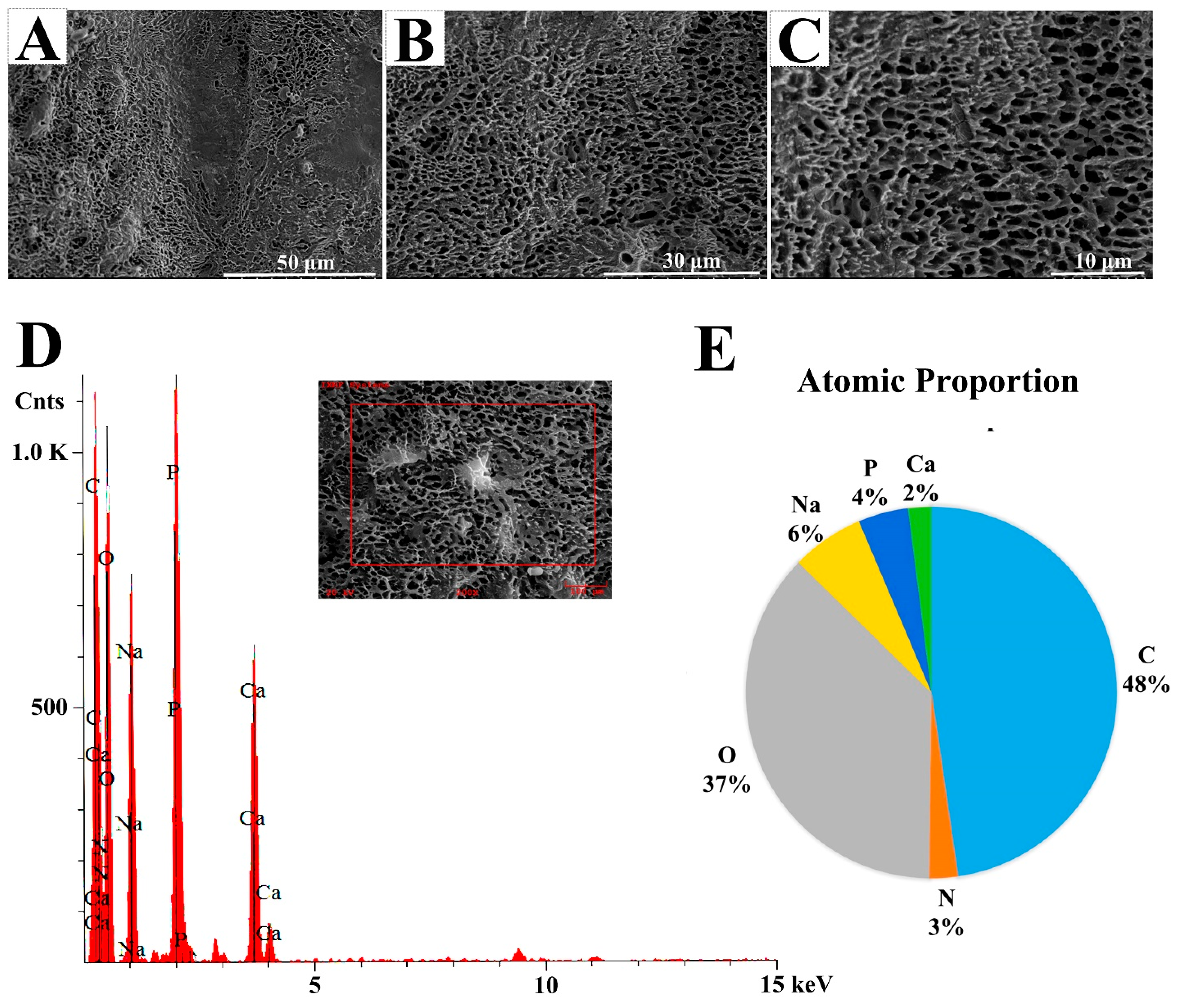
2.2.3. Scanning Electron Microscopy (SEM) and Energy-Dispersive Spectrum (EDS) Observations of EPS364
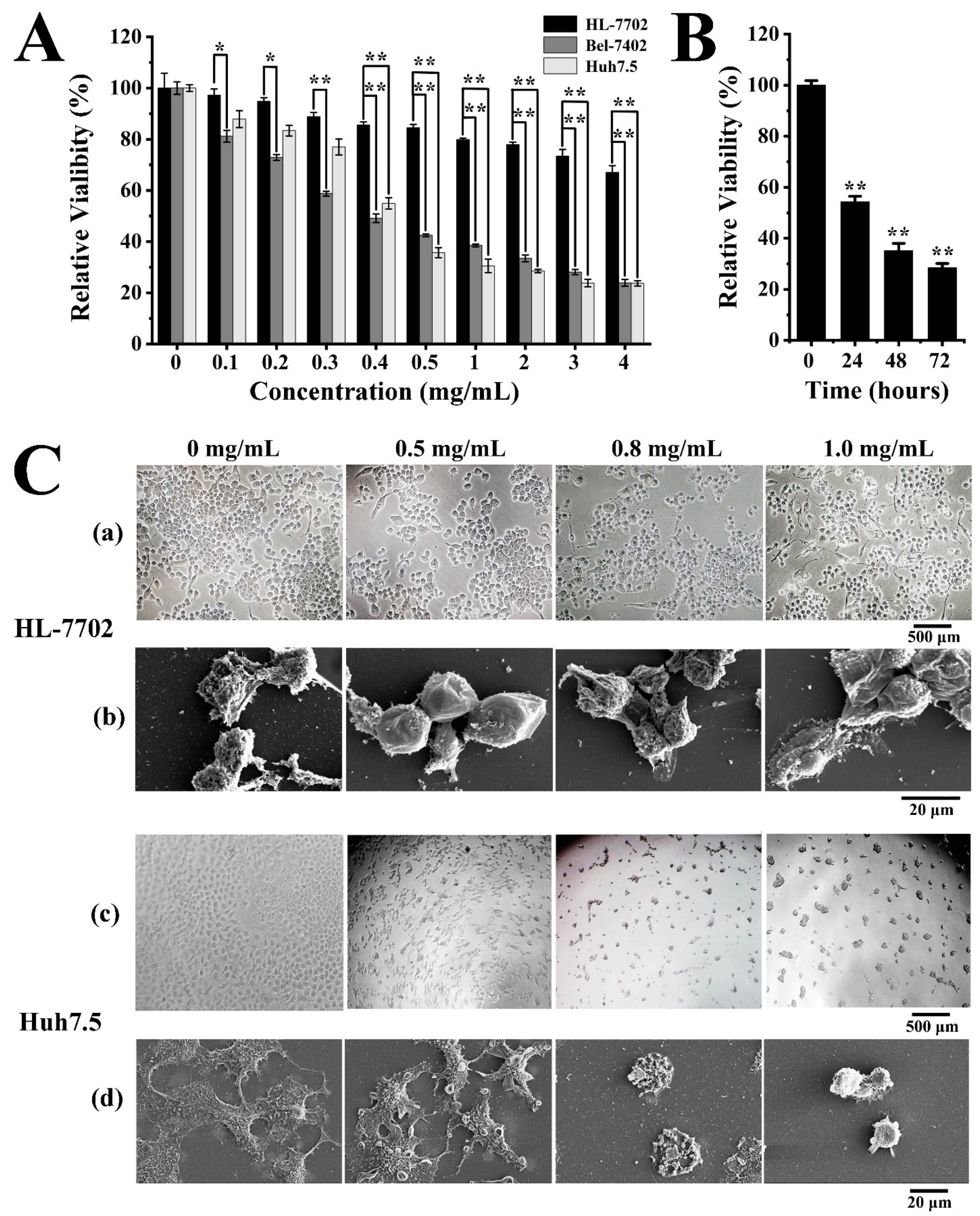
2.3. Antitumor Activities of Exopolysaccharide EPS364
2.3.1. EPS364 Inhibited Cell Proliferation and Adhesion
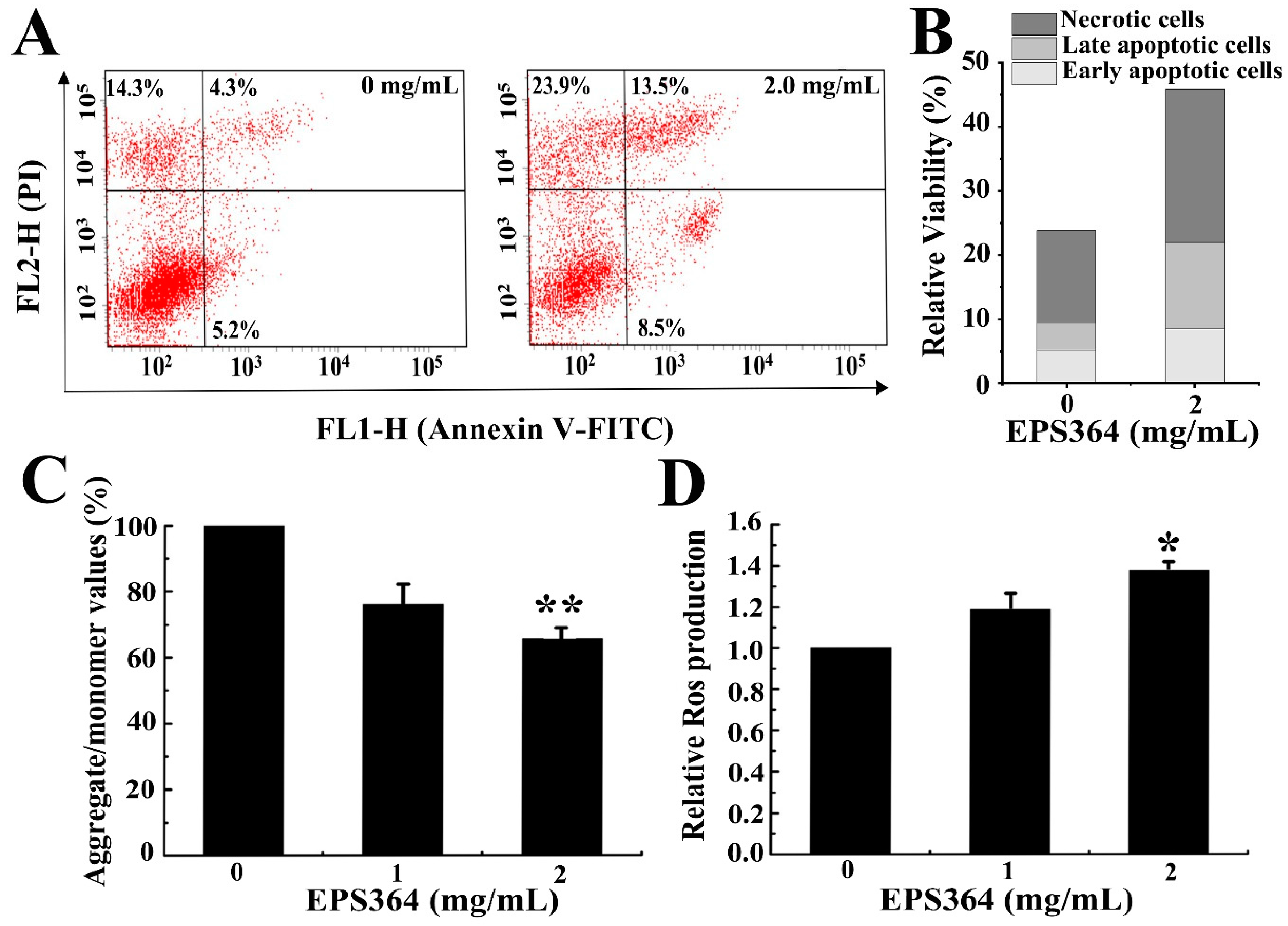
2.3.2. EPS364 Induced Apoptosis, Loss of Mitochondrial Membrane Potential (MMP) and Reactive Oxygen Species (ROS)
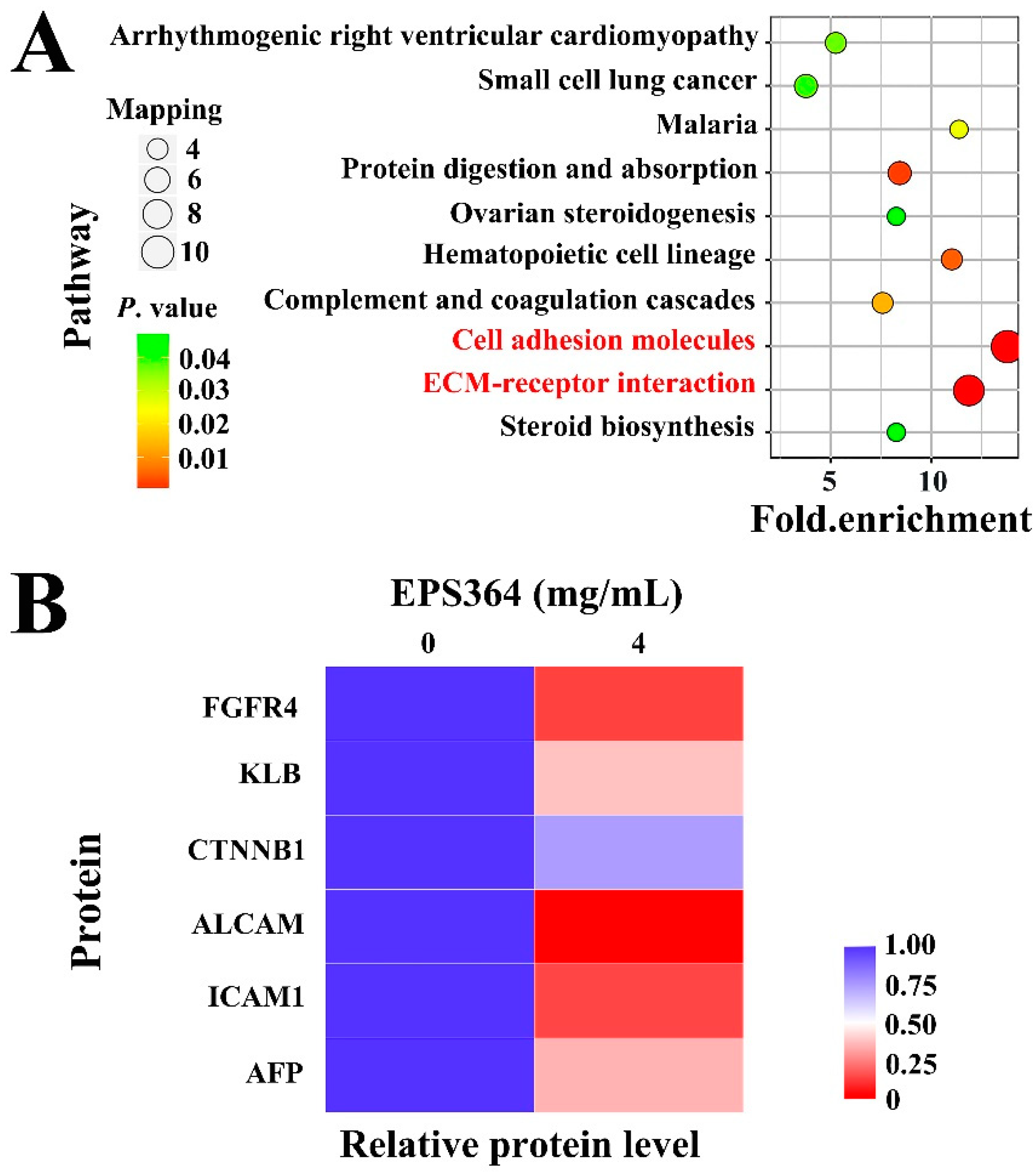
2.3.3. Proteomic Analysis Growth Inhibition of Huh7.5 Cells after EPS364 Treatment
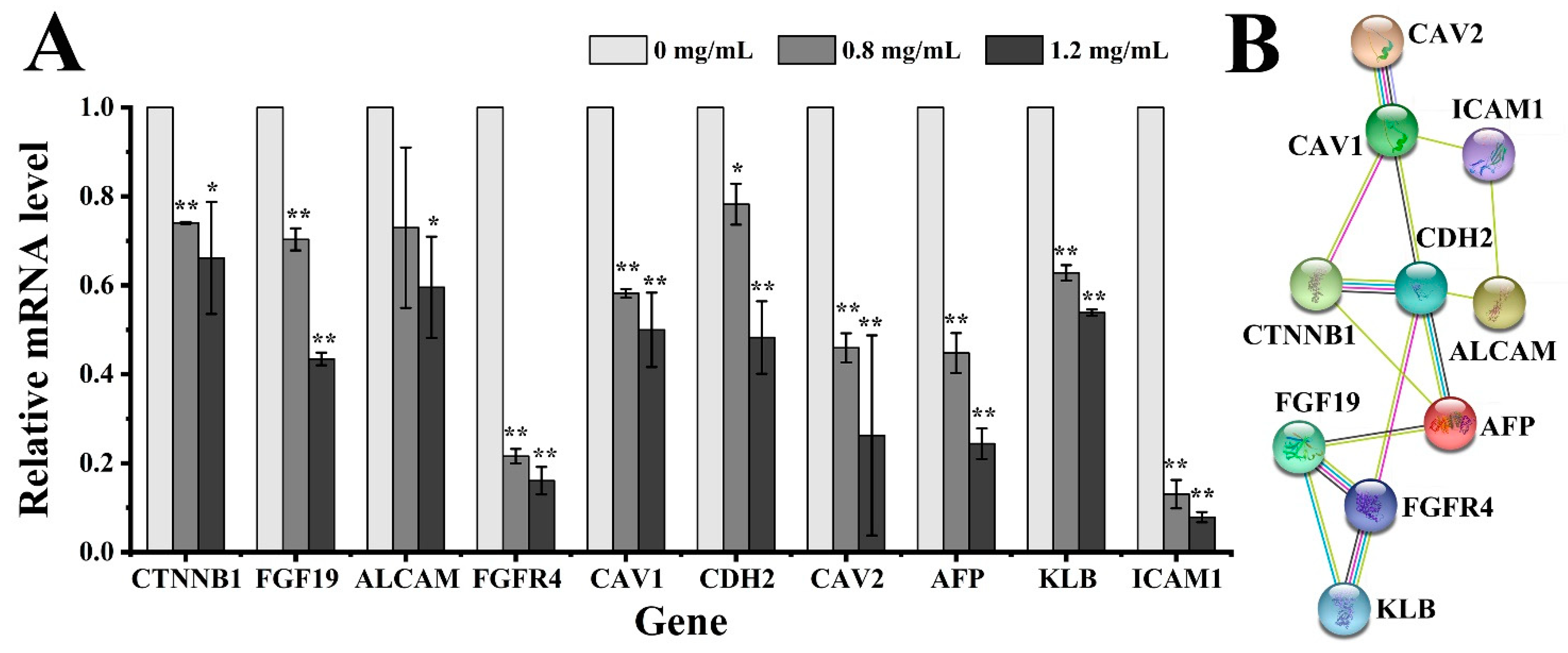
2.3.4. Quantitative Real-Time PCR (qRT-PCR) and Protein Interaction Analyses
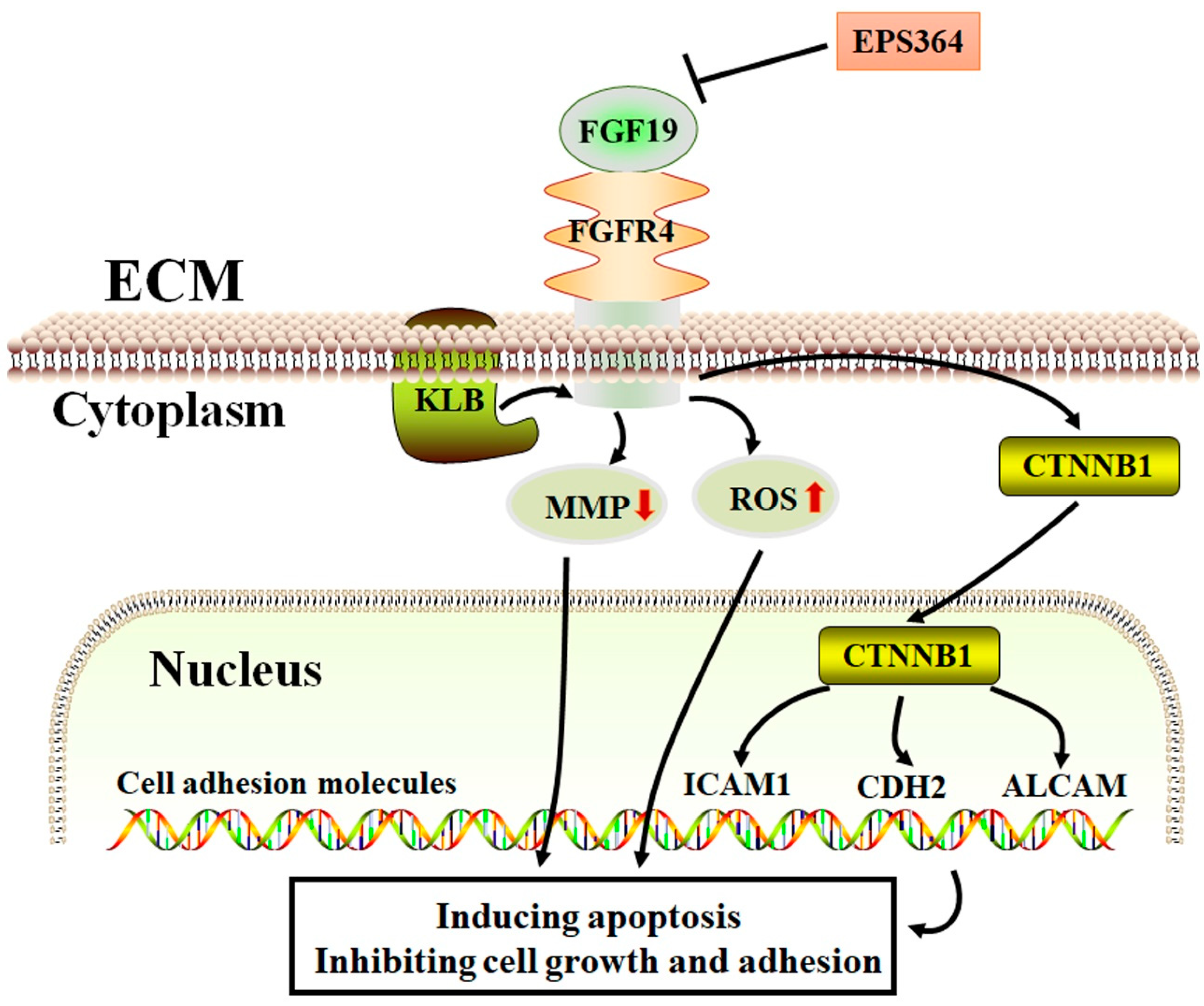
3. Discussion
4. Materials and Methods
4.1. Isolation, Extraction and Purification of Exopolysaccharide EPS364
4.2. Characterization of Polysaccharide EPS364
4.2.1. Molecular Weight Analysis
4.2.2. Monosaccharide Composition Analysis
4.2.3. Fourier-Transformation Infrared (FTIR) Spectroscopy Analysis
4.2.4. Scanning Electron Microscopy (SEM) and Energy-Dispersive Spectrum (EDS) Analyses of Polysaccharide EPS364
4.3. Measurement of Anticancer Activities
4.3.1. Cell Culture
4.3.2. Cell Proliferation Assay
4.3.3. Cell Morphology Observation by Inverted Phase Contrast Microscope and Scanning Electron Microscopy (SEM)
4.3.4. Cell Apoptosis, Mitochondrial Membrane Potential (MMP) and Reactive Oxygen Species (ROS) Detection
4.3.5. Proteomic Analysis
4.3.6. Quantitative Real-Time PCR (qRT-PCR) and Protein Interaction Analyses
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forner, A.; Reig, M.; Bruix, J. Hepatocellular carcinoma. Lancet 2018, 391, 1301–1314. [Google Scholar] [CrossRef]
- Lurje, I.; Czigany, Z.; Bednarsch, J.; Roderburg, C.; Isfort, P.; Neumann, U.P.; Lurje, G. Treatment strategies for hepatocellular carcinoma—a multidisciplinary approach. Int. J. Mol. Sci. 2019, 20, 1465. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Luo, D.M.; Luesch, H. Advances in exploring the therapeutic potential of marine natural products. Pharm. Res. 2019, 147, 104373. [Google Scholar] [CrossRef]
- Newman, D.; Cragg, G. Drugs and drug candidates from marine sources: An assessment of the current “state of play”. Planta Med. 2016, 82, 775–789. [Google Scholar] [CrossRef] [Green Version]
- Niu, G.Q.; Li, W.L. Next-generation drug discovery to combat antimicrobial resistance. Trends Biochem. Sci. 2019, 44, 961–972. [Google Scholar] [CrossRef]
- Wiese, J.; Imhoff, J.F. Marine bacteria and fungi as promising source for new antibiotics. Drug Dev. Res. 2019, 80, 24–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.P.; Ding, W.; Li, Y.X.; Tam, C.; Bougouffa, S.; Wang, R.J.; Pei, B.T.; Chiang, H.Y.; Leung, P.; Lu, Y.H.; et al. Marine biofilms constitute a bank of hidden microbial diversity and functional potential. Nat. Commun. 2019, 10, 517. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, X.C.; Pu, H.M.; Liu, S.; Kan, J.; Jin, C.H. Recent advances in endophytic exopolysaccharides: Production, structural characterization, physiological role and biological activity. Carbohydr. Polym. 2017, 157, 1113–1124. [Google Scholar] [CrossRef]
- Moscovici, M. Present and future medical applications of microbial exopolysaccharides. Front. Microbiol. 2015, 6, 1012. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Liu, G.; Ma, W.P.; Lu, Z.X.; Sun, C.M. Marine bacterial polysaccharide EPS11 inhibits cancer cell growth and metastasis via blocking cell adhesion and attenuating filiform structure formation. Mar. Drugs 2019, 17, 50. [Google Scholar] [CrossRef] [Green Version]
- Cao, R.; Jin, W.; Shan, Y.; Wang, J.; Liu, G.; Kuang, S.; Sun, C. Marine bacterial polysaccharide EPS11 inhibits cancer cell growth via blocking cell adhesion and stimulating anoikis. Mar. Drugs 2018, 16, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, X.; Li, Z.; Ma, K.; Zhang, C.L.; Chen, X.H.; Wang, G.X.; Yang, L.; Dong, M.S.; Rui, X.; Zhang, Q.Q.; et al. Structural characterization and immunomodulatory activity of an exopolysaccharide produced by Lactobacillus helveticus LZ-R-5. Carbohydr. Polym. 2020, 235, 115977. [Google Scholar] [CrossRef]
- Sardari, R.R.R.; Kulcinskaja, E.; Ron, E.Y.C.; Bjornsdottir, S.; Fridjonsson, O.H.; Hreggvidsson, G.O.; Karlsson, E.N. Evaluation of the production of exopolysaccharides by two strains of the thermophilic bacterium Rhodothermus marinus. Carbohydr. Polym. 2017, 156, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, K.; Zeng, Y.T.; Yang, M.L.; Chen, S.J.; He, L.; Ao, X.L.; Zou, L.K.; Liu, S.L. Production, purification and structural study of an exopolysaccharide from Lactobacillus plantarum BC-25. Carbohydr. Polym. 2016, 144, 205–214. [Google Scholar] [CrossRef]
- Addoun, N.; Boual, Z.; Delattre, C.; Ursu, A.V.; Desbrieres, J.; Le Cerf, D.; Gardarin, C.; Hentati, F.; El-Hadj, M.D.O.; Michaud, P.; et al. Structural features and rheological behavior of a water-soluble polysaccharide extracted from the seeds of Plantago ciliata Desf. Int. J. Biol. Macromol. 2020, 155, 1333–1341. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Deng, H.; Jin, S.; Yang, W.; Wang, H.; Meng, C.; Wang, H.; Yang, S. A polysaccharide from dried aerial parts of Agrimonia pilosa: Structural characterization and its potential therapeutic activity for steroid-induced necrosis of the femoral head (SANFH). Carbohydr. Polym. 2019, 214, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Huang, G.; Huang, H. Preparation, analysis, antioxidant activities in vivo of phosphorylated polysaccharide from Momordica charantia. Carbohydr. Polym. 2021, 252, 117179. [Google Scholar] [CrossRef]
- Yang, Y.F.; Feng, F.; Zhou, Q.Q.; Zhao, F.K.; Du, R.P.; Zhou, Z.J.; Han, Y. Isolation, purification and characterization of exopolysaccharide produced by Leuconostoc pseudomesenteroides YF32 from soybean paste. Int. J. Biol. Macromol. 2018, 114, 529–535. [Google Scholar] [CrossRef]
- Rani, R.P.; Anandharaj, M.; Ravindran, A.D. Characterization of a novel exopolysaccharide produced by Lactobacillus gasseri FR4 and demonstration of its in vitro biological properties. Int. J. Biol. Macromol. 2018, 109, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Jacquemet, G.; Hamidi, H.; Ivaska, J. Filopodia in cell adhesion, 3D migration and cancer cell invasion. Curr. Opin. Cell Biol. 2015, 36, 23–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakamuru, S.; Attene-Ramos, M.S.; Xia, M. Mitochondrial membrane potential assay. Methods Mol. Biol. 2016, 1473, 17–22. [Google Scholar]
- Ma, Z.J.; Lu, L.; Yang, J.J.; Wang, X.X.; Su, G.; Wang, Z.L.; Chen, G.H.; Sun, H.M.; Wang, M.Y.; Yang, Y. Lariciresinol induces apoptosis in HepG2 cells via mitochondrial-mediated apoptosis pathway. Eur. J. Pharm. 2018, 821, 1–10. [Google Scholar] [CrossRef]
- Gao, L.X.; Loveless, J.; Shay, C.; Teng, Y. Targeting ROS-mediated crosstalk between autophagy and apoptosis in cancer. Adv. Exp. Med. Biol. 2020, 1260, 1–12. [Google Scholar] [PubMed]
- Moloney, J.N.; Cotter, T.G. ROS signalling in the biology of cancer. Semin. Cell Dev. Biol. 2018, 80, 50–64. [Google Scholar] [CrossRef]
- Figenschau, S.L.; Knutsen, E.; Urbarova, I.; Fenton, C.; Elston, B.; Perander, M.; Mortensen, E.S.; Fenton, K.A. ICAM1 expression is induced by proinflammatory cytokines and associated with TLS formation in aggressive breast cancer subtypes. Sci. Rep. 2018, 8, 11720. [Google Scholar] [CrossRef] [Green Version]
- Inaguma, S.; Lasota, J.; Wang, Z.; Czapiewski, P.; Langfort, R.; Rys, J.; Szpor, J.; Waloszczyk, P.; Okon, K.; Biernat, W.; et al. Expression of ALCAM (CD166) and PD-L1 (CD274) independently predicts shorter survival in malignant pleural mesothelioma. Hum. Pahuol. 2018, 71, 1–7. [Google Scholar] [CrossRef]
- Jia, G.D.; Wang, X.; Yan, M.; Chen, W.T.; Zhang, P. CD166-mediated epidermal growth factor receptor phosphorylation promotes the growth of oral squamous cell carcinoma. Oral Oncol. 2016, 59, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Saga, K.; Park, J.; Nimura, K.; Kawamura, N.; Ishibashi, A.; Nonomura, N.; Kaneda, Y. NANOG helps cancer cells escape NK cell attack by downregulating ICAM1 during tumorigenesis. J. Exp. Clin. Canc. Res. 2019, 38, 416. [Google Scholar] [CrossRef] [PubMed]
- Rozario, T.; DeSimone, D.W. The extracellular matrix in development and morphogenesis: A dynamic view. Dev. Biol. 2010, 341, 126–140. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.J.; Tao, J.; Sheng, L.; Hu, X.; Rong, R.M.; Xu, M.; Zhu, T.Y. Twist2 promotes kidney cancer cell proliferation and invasion by regulating ITGA6 and CD44 expression in the ECM-receptor interaction pathway. Onco Targets Ther. 2016, 9, 1801–1812. [Google Scholar] [CrossRef]
- Zou, Y.; Zheng, S.; Deng, X.; Yang, A.; Xie, X.; Tang, H.; Xie, X. The role of circular RNA CDR1as/ciRS-7 in regulating tumor microenvironment: A pan-cancer analysis. Biomolecules 2019, 9, 429. [Google Scholar] [CrossRef] [Green Version]
- Li, M.S.; Li, P.F.; He, S.P.; Du, G.G.; Li, G. The promoting molecular mechanism of alpha-fetoprotein on the growth of human hepatoma Bel7402 cell line. World J. Gastroenterol. 2002, 8, 469–475. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, Y.; Zhang, L.; Zhang, L.; Mao, J. Silencing alpha-fetoprotein expression induces growth arrest and apoptosis in human hepatocellular cancer cell. Cancer Lett. 2008, 271, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Su, N.; Yang, J.; Tan, Q.; Huang, S.; Jin, M.; Ni, Z.; Zhang, B.; Zhang, D.; Luo, F.; et al. FGF/FGFR signaling in health and disease. Signal Transduct Target Ther. 2020, 5, 181. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Mege, R.M. N-Cadherin and fibroblast growth factor receptors crosstalk in the control of developmental and cancer cell migrations. Eur. J. Cell Biol. 2016, 95, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.Y.; Tang, J.; Chen, L.; Wang, B.; Weng, S.; Xie, Z.; Wu, Z.Y.; Shen, Z.; Bai, B.; Yang, L. Imperatorin promotes osteogenesis and suppresses osteoclast by activating AKT/GSK3 β/β-catenin pathways. J. Cell Mol. Med. 2019, 24, 2330–2341. [Google Scholar] [CrossRef]
- Zhao, H.; Lv, F.; Liang, G.; Huang, X.; Wu, G.; Zhang, W.; Yu, L.; Shi, L.; Teng, Y. FGF19 promotes epithelial-mesenchymal transition in hepatocellular carcinoma cells by modulating the GSK3β/β- catenin signaling cascade via FGFR4 activation. Oncotarget 2015, 7, 13575–13586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.M.; Ji, S.; Li, Y.; Fu, L.Y.; Jiang, T.; Meng, F.D. beta-Catenin promotes cell proliferation, migration, and invasion but induces apoptosis in renal cell carcinoma. Onco Targets Ther. 2017, 10, 711–724. [Google Scholar] [CrossRef] [Green Version]
- Silva, R.d.S.; Lombardi, A.P.G.; de Souza, D.S.; Vicente, C.M.; Porto, C.S. Activation of estrogen receptor beta (ERβ) regulates the expression of N-cadherin, E-cadherin and β-catenin in androgen-independent prostate cancer cells. Int. J. Biochem. Cell. Biol. 2018, 96, 40–50. [Google Scholar] [CrossRef]
- Rutkowski, R.; Chrzanowski, R.; Trwoga, M.; Kochanowicz, J.; Turek, G.; Mariak, Z.; Reszeć, J. Expression of N-cadherin and β-catenin in human meningioma in correlation with peritumoral edema. Int. J. Neurosci. 2018, 128, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Rogers, C.D.; Sorrells, L.K.; Bronner, M.E. A catenin-dependent balance between N-cadherin and E-cadherin controls neuroectodermal cell fate choices. Mech. Dev. 2018, 152, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Qiu, F.; Shin, Y.; Chen, D.; Cheng, R.; Chen, Q.; Zhou, K.; Larrick, J.W.; Mendelson, A.R.; Ma, J.X. Anti-angiogenic effect of a humanized antibody blocking the Wnt/beta-catenin signaling pathway. Microvasc. Res. 2018, 119, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Oh, B.Y.; Kim, S.-Y.; Lee, Y.S.; Hong, H.K.; Kim, T.W.; Kim, S.H.; Lee, W.Y.; Cho, Y.B. Twist1-induced epithelial-mesenchymal transition according to microsatellite instability status in colon cancer cells. Oncotarget 2016, 7, 57066–57076. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Ma, N.; Xiong, Y.L.; Wang, L.; Li, W.M.; Lai, Y.Y.; Zhang, C.X.; Zhang, Z.P.; Li, X.F.; Zhao, J.B. Aberrantly high expression of NOK/STYK1 is tightly associated with the activation of the AKT/GSK3β/N-Cadherin pathway in non-small cell lung cancer. Onco Targets Ther. 2019, 12, 10299–10309. [Google Scholar] [CrossRef] [Green Version]
- Fu, C.; Liu, P.; Li, P.; Liu, W.; Huang, X.; Liang, Y. FSP1 promotes the biofunctions of adventitial fibroblast through the crosstalk among RAGE, JAK2/STAT3 and Wnt3a/beta-catenin signalling pathways. J. Cell Mol. Med. 2019, 23, 7246–7260. [Google Scholar] [CrossRef] [Green Version]
- Devis, L.; Martinez-Garcia, E.; Moiola, C.P.; Quiles, M.T.; Arbos, M.A.; Stirbat, T.V.; Brochard-Wyart, F.; Garcia, A.; Alonso-Alconada, L.; Abal, M.; et al. ALCAM shedding at the invasive front of the tumor is a marker of myometrial infiltration and promotes invasion in endometrioid endometrial cancer. Oncotarget 2018, 9, 16648–16664. [Google Scholar] [CrossRef] [Green Version]
- Ali, R.M.; Al-Shorbagy, M.Y.; Helmy, M.W.; El-Abhar, H.S. Role of Wnt4/β-catenin, Ang II/TGFβ, ACE2, NF-κB, and IL-18 in attenuating renal ischemia/reperfusion-induced injury in rats treated with Vit D and pioglitazone. Eur. J. Pharmacol. 2018, 831, 68–76. [Google Scholar] [CrossRef]
- Lang, L.; Shull, A.Y.; Teng, Y. Interrupting the FGF19-FGFR4 axis to therapeutically disrupt cancer progression. Curr. Cancer Drug Targets 2018, 19, 17–25. [Google Scholar] [CrossRef]
- Lin, B.C.; Wang, M.; Blackmore, C.; Desnoyers, L.R. Liver-specific activities of FGF19 require Klotho beta. J. Biol. Chem. 2007, 282, 27277–27284. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.X.; Wang, X.L.; Tang, Y.L.; Huang, S.; Hu, C.A.A.; Teng, Y. FGF19/FGFR4 signaling contributes to the resistance of hepatocellular carcinoma to sorafenib. J. Exp. Clin. Canc. Res. 2017, 36, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raja, A.; Park, I.; Haq, F.; Ahn, S.M. FGF19-FGFR4 signaling in hepatocellular carcinoma. Cells 2019, 8, 536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, S.Y.; Hao, Y.L.; Yuan, Y.; Liu, R.; Chen, Q.M. Role of fibroblast growth factor receptor 4 in cancer. Cancer Sci. 2018, 109, 3024–3031. [Google Scholar] [CrossRef] [PubMed]
- Kamibeppu, T.; Yamasaki, K.; Nakahara, K.; Nagai, T.; Terada, N.; Tsukino, H.; Mukai, S.; Kamoto, T. Caveolin-1 and -2 regulate cell motility in castration-resistant prostate cancer. Res. Rep. Urol. 2018, 10, 135–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ketteler, J.; Klein, D. Caveolin-1, cancer and therapy resistance. Int. J. Cancer 2018, 143, 2092–2104. [Google Scholar] [CrossRef]
- Park, S.Y.; Park, J.W.; Lee, G.W.; Li, L.; Chun, Y.S. Inhibition of neddylation facilitates cell migration through enhanced phosphorylation of caveolin-1 in PC3 and U373MG cells. BMC Cancer 2018, 18, 30. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Qi, X.; Liu, H.; Xue, K.; Xu, S.; Tian, Z. The anti-cancer effects of fucoidan: A review of both in vivo and in vitro investigations. Cancer Cell Int. 2020, 20, 154. [Google Scholar] [CrossRef]
- Pai, R.; Dunlap, D.; Qing, J.; Mohtashemi, I.; Hotzel, K.; French, D.M. Inhibition of fibroblast growth factor 19 reduces tumor growth by modulating beta-catenin signaling. Cancer Res. 2008, 68, 5086–5095. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.; Hou, X.; Zhang, D.; Zhang, Q.; Yan, C. Two novel polysaccharides from rhizomes of Cibotium barometz promote bone formation via activating the BMP2/SMAD1 signaling pathway in MC3T3-E1 cells. Carbohydr. Polym. 2020, 231, 115732. [Google Scholar] [CrossRef]
- Dai, J.; Wu, Y.; Chen, S.W.; Zhu, S.; Yin, H.P.; Wang, M.; Tang, J.A. Sugar compositional determination of polysaccharides from Dunaliella salina by modified RP-HPLC method of precolumn derivatization with 1-phenyl-3-methyl-5-pyrazolone. Carbohydr. Polym. 2010, 82, 629–635. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, R.; Xi, S.C.; Cai, R.N.; Zhang, X.; Sun, C.M. A novel bacterial thiosulfate oxidation pathway provides a new clue about the formation of zero-valent sulfur in deep sea. ISME J. 2020, 14, 2261–2274. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Kuang, S.; Cao, R.B.; Wang, J.; Peng, Q.C.; Sun, C.M. Sorafenib kills liver cancer cells by disrupting SCD1-mediated synthesis of monounsaturated fatty acids via the ATP-AMPK-mTOR-SREBP1 signaling pathway. Faseb J. 2019, 33, 10089–10103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Kuang, S.; Wu, S.M.; Jin, W.H.; Sun, C.M. A novel polysaccharide from Sargassum integerrimum induces apoptosis in A549 cells and prevents angiogensis in vitro and in vivo. Sci. Rep. 2016, 6, 26722. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Liu, G.; Liu, R.; Wei, M.; Zhang, J.; Sun, C. EPS364, a Novel Deep-Sea Bacterial Exopolysaccharide, Inhibits Liver Cancer Cell Growth and Adhesion. Mar. Drugs 2021, 19, 171. https://doi.org/10.3390/md19030171
Wang Y, Liu G, Liu R, Wei M, Zhang J, Sun C. EPS364, a Novel Deep-Sea Bacterial Exopolysaccharide, Inhibits Liver Cancer Cell Growth and Adhesion. Marine Drugs. 2021; 19(3):171. https://doi.org/10.3390/md19030171
Chicago/Turabian StyleWang, Yun, Ge Liu, Rui Liu, Maosheng Wei, Jinxiang Zhang, and Chaomin Sun. 2021. "EPS364, a Novel Deep-Sea Bacterial Exopolysaccharide, Inhibits Liver Cancer Cell Growth and Adhesion" Marine Drugs 19, no. 3: 171. https://doi.org/10.3390/md19030171