
Farmed Gilthead Sea Bream (Sparus aurata) by-Products Valorization: Viscera Oil ω-3 Enrichment by Short-Path Distillation and In Vitro Bioactivity Evaluation

 , , , , and
, , , , and
Abstract
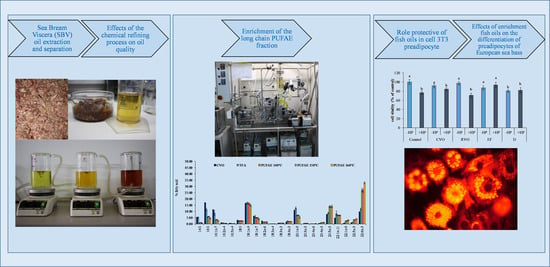
:
1. Introduction
2. Results and Discussion
2.1. Proximate Composition of Sea Bream Viscera (SBV) By-Product
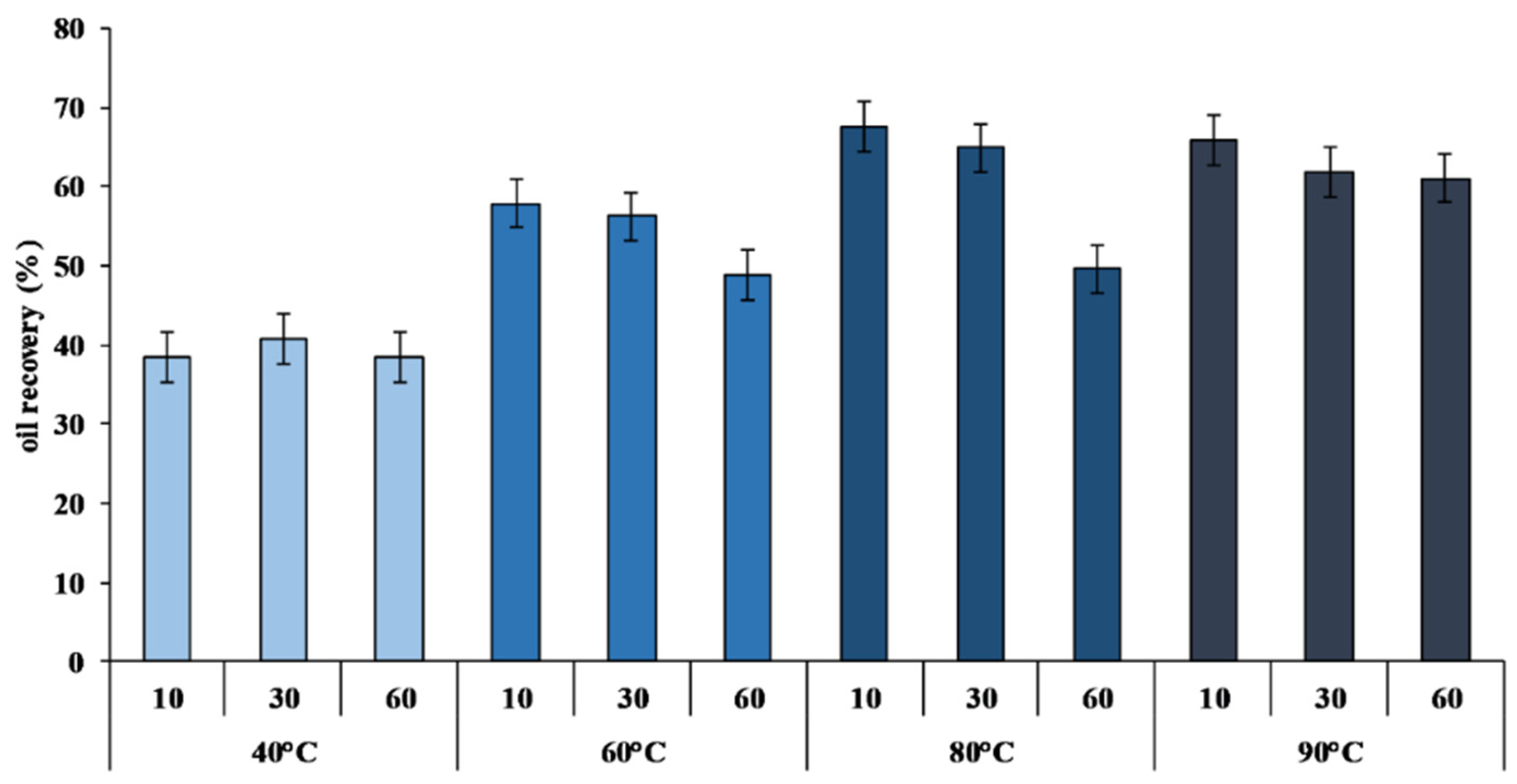
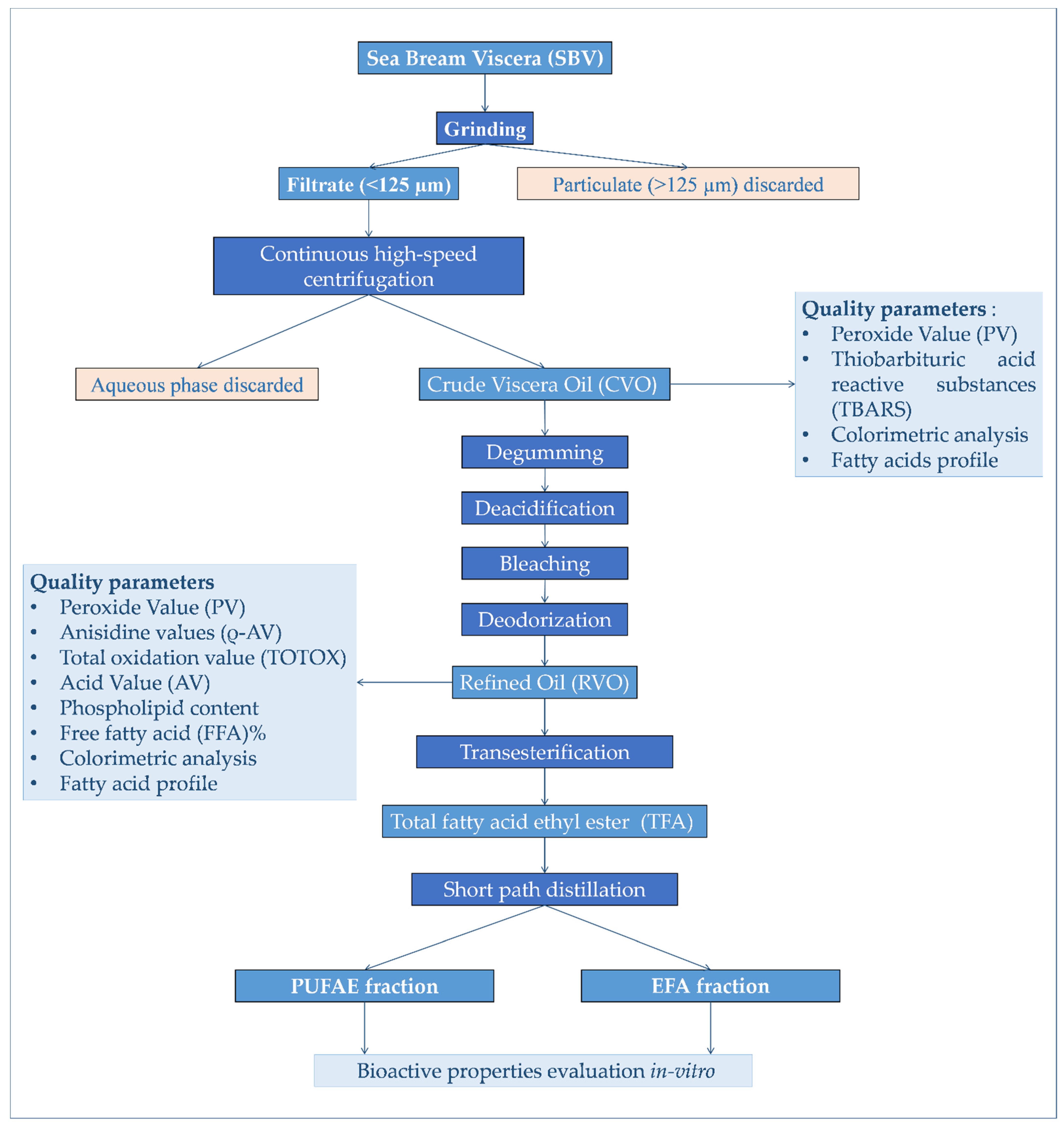
2.2. Yield and Quality of Crude Viscera Oil (CVO)
2.3. Crude Oil Refining: Effects of the Chemical Processes on Oil Quality
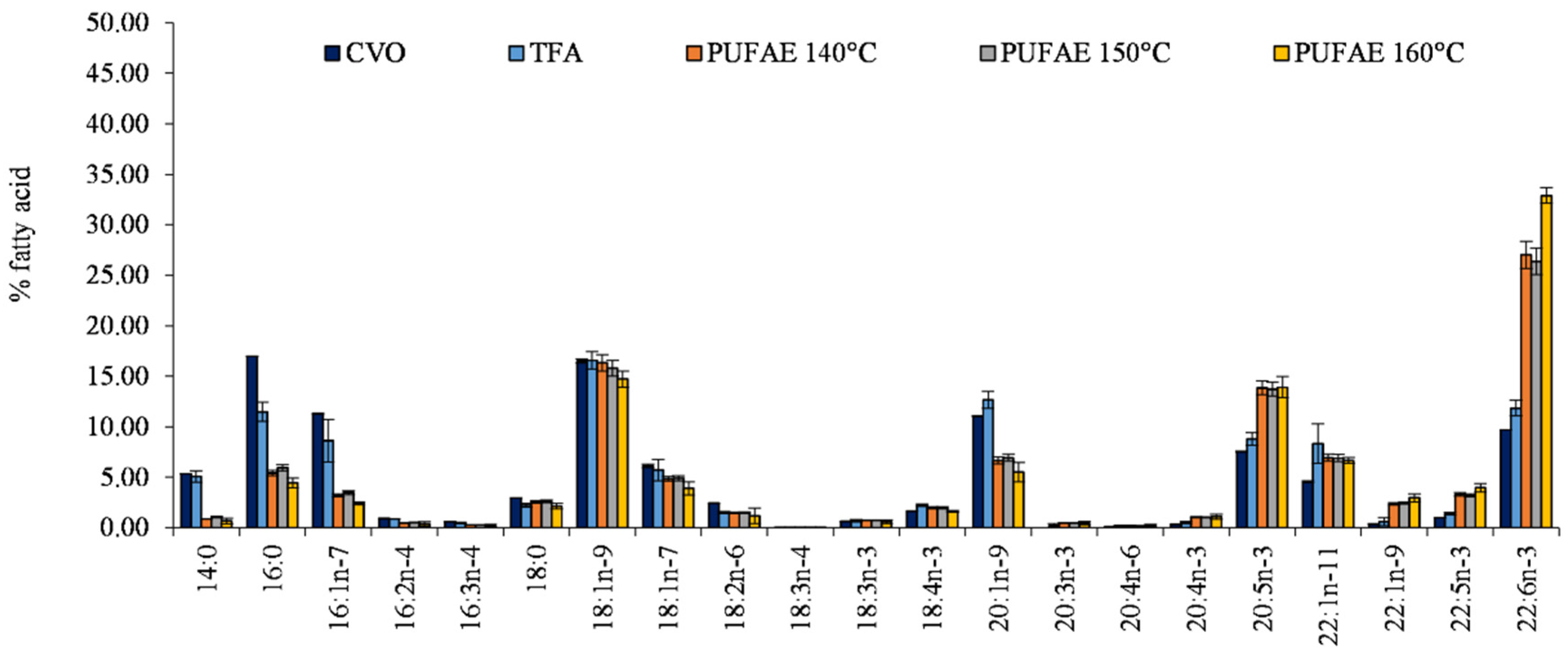
2.4. PUFA Enrichment
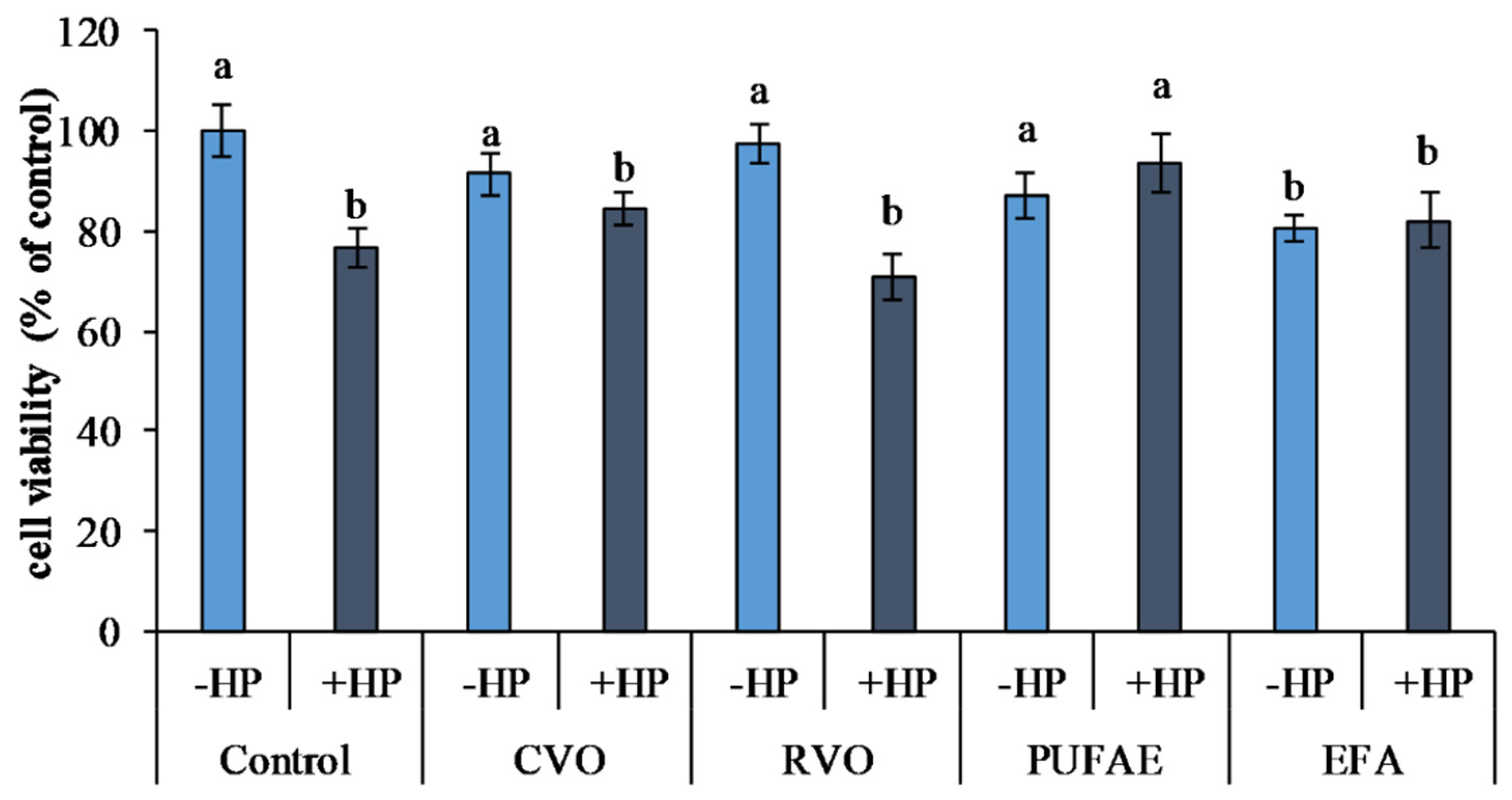
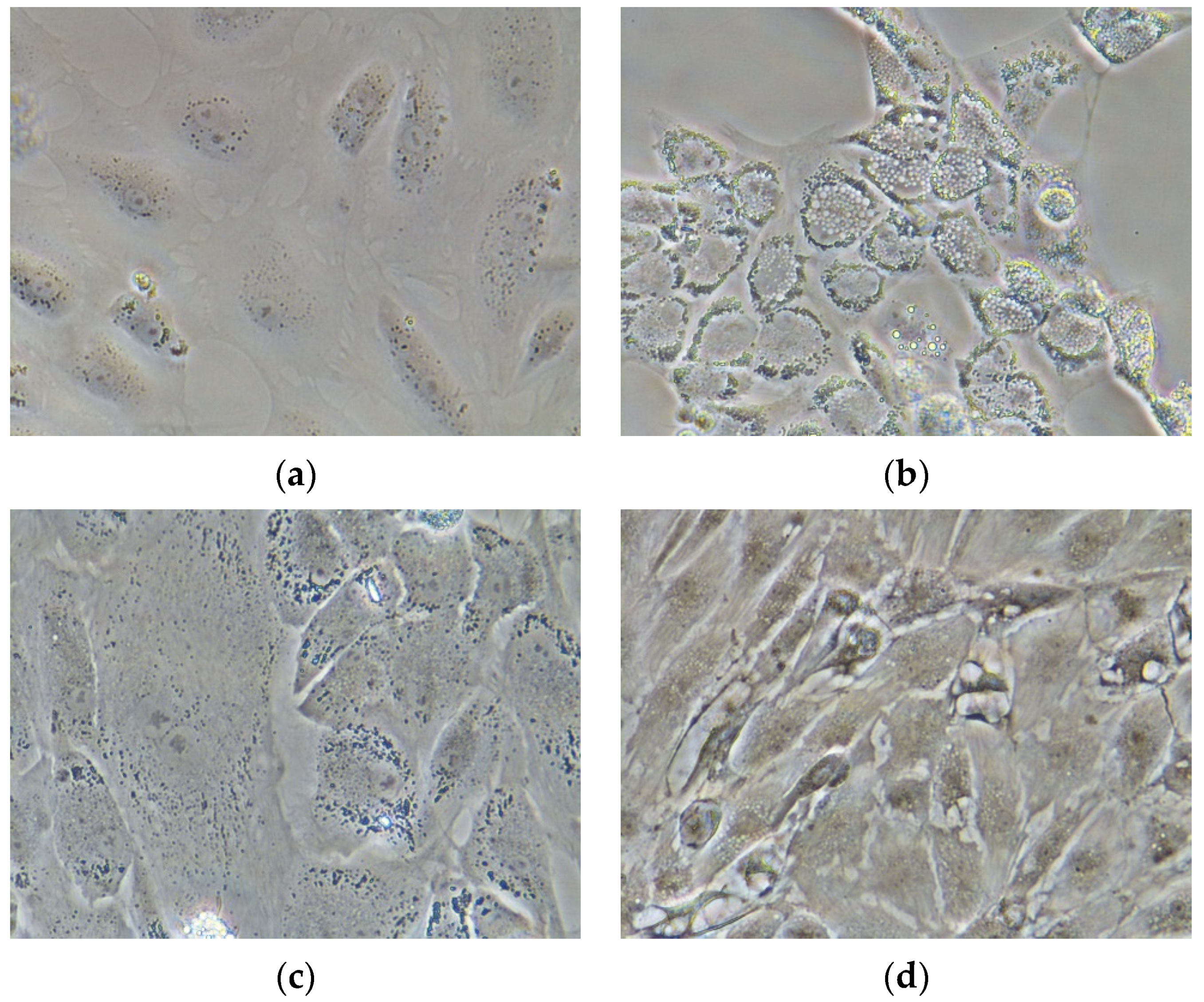
2.5. In Vitro Bioactive Properties of the Refined Oil
3. Materials and Methods
3.1. Sampling
3.2. Proximate Composition and Fatty Acid Profile
3.3. Extraction of Crude Oil
3.4. Chemical Refining Process of CVO
3.5. Assessment of Oil Quality
3.5.1. Peroxide Value (PV)
3.5.2. Thiobarbituric Acid Reactive Substances Analysis (TBARS)
3.5.3. Content of ρ-Anisidine (ρ-AV)
3.5.4. Total Oxidation Value (TOTOX)
3.5.5. Phospholipid Content
3.5.6. Acid Value
3.5.7. Colorimetric Analysis
3.6. PUFA Enrichment
- -
- EPA and DHA%.
- -
- Fatty acid ratio ® (4) [72]:
- -
- Enrichment factor for EPA, DHA, and PUFA.
- -
- Ratio of total PUFA to total saturated fatty acids (PUFA/saturated).
3.7. In Vitro Bioactive Properties
Image Acquisition
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Coppola, D.; Lauritano, C.; Esposito, F.P.; Riccio, G.; Rizzo, C.; de Pascale, D. Fish Waste: From Problem to Valuable Resource. Mar. Drugs 2021, 19, 116. [Google Scholar] [CrossRef] [PubMed]
- García-De-Vinuesa, A.; Demestre, M.; Carreño, A.; Lloret, J. The Bioactive Potential of Trawl Discard: Case Study from A Crinoid Bed off Blanes (North-Western Mediterranean). Mar. Drugs 2021, 19, 83. [Google Scholar] [CrossRef]
- Alfio, V.G.; Manzo, C.; Micillo, R. From Fish Waste to Value: An Overview of the Sustainable Recovery of Omega-3 for Food Supplements. Molecules 2021, 26, 1002. [Google Scholar] [CrossRef]
- Lund, E.K. Health benefits of seafood; Is it just the fatty acids? In Proceedings of the Food Chemistry; Elsevier: Amsterdam, The Netherlands, 2013; Volume 140, pp. 413–420. [Google Scholar]
- Rimm, E.B.; Appel, L.J.; Chiuve, S.E.; Djoussé, L.; Engler, M.B.; Kris-Etherton, P.M.; Mozaffarian, D.; Siscovick, D.S.; Lichtenstein, A.H. A Seafood Long-Chain n-3 Polyunsaturated Fatty Acids and Cardiovascular Disease: A Science Advisory From the American Heart Association. Circulation 2018, 138, e35–e47. [Google Scholar] [CrossRef] [PubMed]
- Zárate, R.; El Jaber-Vazdekis, N.; Tejera, N.; Pérez, J.A.; Rodríguez, C. Significance of long chain polyunsaturated fatty acids in human health. Clin. Transl. Med. 2017, 6, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Caruso, G.; Floris, R.; Serangeli, C.; Di Paola, L. Fishery Wastes as a Yet Undiscovered Treasure from the Sea: Biomolecules Sources, Extraction Methods and Valorization. Mar. Drugs 2020, 18, 622. [Google Scholar] [CrossRef]
- Pateiro, M.; Munekata, P.E.S.; Domínguez, R.; Wang, M.; Barba, F.J.; Bermúdez, R.; Lorenzo, J.M. Nutritional Profiling and the Value of Processing By-Products from Gilthead Sea Bream (Sparus aurata). Mar. Drugs 2020, 18, 101. [Google Scholar] [CrossRef] [Green Version]
- Marsol-Vall, A.; Aitta, E.; Guo, Z.; Yang, B. Green technologies for production of oils rich in n-3 polyunsaturated fatty acids from aquatic sources. Crit. Rev. Food Sci. Nutr. 2021, 1–21. [Google Scholar] [CrossRef]
- Mentoor, I.; Engelbrecht, A.-M.; Nell, T. Fatty acids: Adiposity and breast cancer chemotherapy, a bad synergy? Prostaglandins Leukot. Essent. Fat. Acids 2019, 140, 18–33. [Google Scholar] [CrossRef] [PubMed]
- Tarasiuk, A.; Mosińska, P.; Fichna, J. The mechanisms linking obesity to colon cancer: An overview. Obes. Res. Clin. Pr. 2018, 12, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Field, C.J.; Schley, P.D.; Brindley, D.N. Omega-3 polyunsaturated fatty acids alter raft lipid composition and decrease epidermal growth factor receptor levels in lipid rafts of human breast cancer cells. FASEB J. 2007, 21, A165. [Google Scholar] [CrossRef]
- Siddiqui, R.A.; Harvey, K.; Stillwell, W. Anticancer properties of oxidation products of docosahexaenoic acid. Chem. Phys. Lipids 2008, 153, 47–56. [Google Scholar] [CrossRef]
- Omabe, M.; Ezeani, M.; Omabe, K.N. Lipid metabolism and cancer progression: The missing target in metastatic cancer treatment. J. Appl. Biomed. 2015, 13, 47–59. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Wu, J.H.Y. Omega-3 fatty acids and cardiovascular disease: Effects on risk factors, molecular pathways, and clinical events. J. Am. Coll. Cardiol. 2011, 58, 2047–2067. [Google Scholar] [CrossRef] [Green Version]
- Zamora-Sillero, J.; Gharsallaoui, A.; Prentice, C.; Vázquez, J.A.; Meduíña, A.; Durán, A.I.; Nogueira, M.; Fernández-Compás, A.; Pérez-Martín, R.I.; Rodríguez-Amado, I.; et al. Impact of omega-3 polyunsaturated fatty acids on vascular function and blood pressure: Relevance for cardiovascular outcomes. Circulation 2019, 138, 191–200. [Google Scholar]
- Guillen, J.; Natale, F.; Carvalho, N.; Casey, J.; Hofherr, J.; Druon, J.-N.; Fiore, G.; Gibin, M.; Zanzi, A.; Martinsohn, J.T. Global seafood consumption footprint. Ambio 2019, 48, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Turchini, G.M.; Ng, W.K.; Tocher, D.R. Fish Oil Replacement and Alternative Lipid Sources in Aquaculture Feeds; CRC Press: Boca Raton, FL, USA, 2010; ISBN 9781439808634. [Google Scholar]
- Shepherd, C.J.; Jackson, A.J. Global fishmeal and fish-oil supply: Inputs, outputs and marketsa. J. Fish Biol. 2013, 83, 1046–1066. [Google Scholar] [CrossRef]
- Fountoulaki, E.; Vasilaki, A.; Hurtado, R.; Grigorakis, K.; Karacostas, I.; Nengas, I.; Rigos, G.; Kotzamanis, Y.; Venou, B.; Alexis, M. Fish oil substitution by vegetable oils in commercial diets for gilthead sea bream (Sparus aurata L.); effects on growth performance, flesh quality and fillet fatty acid profile. Aquaculture 2009, 289, 317–326. [Google Scholar] [CrossRef]
- Messina, C.M.; Renda, G.; La Barbera, L.; Santulli, A. By-products of farmed European sea bass (Dicentrarchus labrax L.) as a potential source of n-3 PUFA. Biologia 2013, 68, 288–293. [Google Scholar] [CrossRef] [Green Version]
- Messina, M.; Piccolo, G.; Tulli, F.; Cardinaletti, G.; Tibaldi, E. Lipid composition and metabolism of European sea bass (Dicentrarchus labrax L.) fed diets containing wheat gluten and legume meals as substitutes for fish meal. Aquaculture 2013, 376–379, 6–14. [Google Scholar] [CrossRef]
- Monge-Ortiz, R.; Tomás-Vidal, A.; Rodriguez-Barreto, D.; Martínez-Llorens, S.; Pérez, J.; Jover-Cerdá, M.; Lorenzo, A. Replacement of fish oil with vegetable oil blends in feeds for greater amberjack (Seriola dumerili) juveniles: Effect on growth performance, feed efficiency, tissue fatty acid composition and flesh nutritional value. Aquac. Nutr. 2018, 24, 605–615. [Google Scholar] [CrossRef]
- Torrecillas, S.; Mompel, D.; Caballero, M.; Montero, D.; Merrifield, D.; Rodiles, A.; Robaina, L.; Zamorano, M.; Karalazos, V.; Kaushik, S.; et al. Effect of fishmeal and fish oil replacement by vegetable meals and oils on gut health of European sea bass (Dicentrarchus labrax). Aquaculture 2017, 468, 386–398. [Google Scholar] [CrossRef]
- Šimat, V.; Vlahović, J.; Soldo, B.; Skroza, D.; Ljubenkov, I.; Mekinić, I.G. Production and Refinement of Omega-3 Rich Oils from Processing By-Products of Farmed Fish Species. Foods 2019, 8, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rustad, T.; Storrø, I.; Slizyte, R. Possibilities for the utilisation of marine by-products. Int. J. Food Sci. Technol. 2011, 46, 2001–2014. [Google Scholar] [CrossRef]
- Antelo, L.T.; Lopes, C.; Franco-Uría, A.; Alonso, A.A. Fish discards management: Pollution levels and best available removal techniques. Mar. Pollut. Bull. 2012, 64, 1277–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galanakis, C.M. Recovery of high added-value components from food wastes: Conventional, emerging technologies and commercialized applications. Trends Food Sci. Technol. 2012, 26, 68–87. [Google Scholar] [CrossRef]
- Stevens, J.R.; Newton, R.W.; Tlusty, M.; Little, D.C. The rise of aquaculture by-products: Increasing food production, value, and sustainability through strategic utilisation. Mar. Policy 2018, 90, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Maschmeyer, T.; Luque, R.; Selva, M. Upgrading of marine (fish and crustaceans) biowaste for high added-value molecules and bio(nano)-materials. Chem. Soc. Rev. 2020, 49, 4527–4563. [Google Scholar] [CrossRef]
- Villamil, O.; Váquiro, H.; Solanilla, J.F. Fish viscera protein hydrolysates: Production, potential applications and functional and bioactive properties. Food Chem. 2017, 224, 160–171. [Google Scholar] [CrossRef]
- Ucak, I.; Afreen, M.; Montesano, D.; Carrillo, C.; Tomasevic, I.; Simal-Gandara, J.; Barba, F. Functional and Bioactive Properties of Peptides Derived from Marine Side Streams. Mar. Drugs 2021, 19, 71. [Google Scholar] [CrossRef]
- Cervera, M.; Ángel, R.; Venegas, E.V.; Bueno, R.P.R.; Medina, M.D.S.; Guerrero, J.L.G. Docosahexaenoic acid purification from fish processing industry by-products. Eur. J. Lipid Sci. Technol. 2015, 117, 724–729. [Google Scholar] [CrossRef]
- Šimat, V.; Vlahović, J.; Soldo, B.; Mekinić, I.G.; Čagalj, M.; Hamed, I.; Skroza, D. Production and characterization of crude oils from seafood processing by-products. Food Biosci. 2020, 33, 100484. [Google Scholar] [CrossRef]
- Kusunoki, C.; Yang, L.; Yoshizaki, T.; Nakagawa, F.; Ishikado, A.; Kondo, M.; Morino, K.; Sekine, O.; Ugi, S.; Nishio, Y.; et al. Omega-3 polyunsaturated fatty acid has an anti-oxidant effect via the Nrf-2/HO-1 pathway in 3T3-L1 adipocytes. Biochem. Biophys. Res. Commun. 2013, 430, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Sakai, C.; Ishida, M.; Ohba, H.; Yamashita, H.; Uchida, H.; Yoshizumi, M.; Ishida, T. Fish oil omega-3 polyunsaturated fatty acids attenuate oxidative stress-induced DNA damage in vascular endothelial cells. PLoS ONE 2017, 12, e0187934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todorčević, M.; Vegusdal, A.; Gjøen, T.; Sundvold, H.; Torstensen, B.E.; Kjær, M.A.; Ruyter, B. Changes in fatty acids metabolism during differentiation of Atlantic salmon preadipocytes; Effects of n-3 and n-9 fatty acids. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2008, 1781, 326–335. [Google Scholar] [CrossRef]
- Riera-Heredia, N.; Lutfi, E.; Sánchez-Moya, A.; Gutiérrez, J.; Capilla, E.; Navarro, I. Short-Term Responses to Fatty Acids on Lipid Metabolism and Adipogenesis in Rainbow Trout (Oncorhynchus mykiss). Int. J. Mol. Sci. 2020, 21, 1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hathwar, S.C.; Bijinu, B.; Rai, A.K.; Narayan, B. Simultaneous Recovery of Lipids and Proteins by Enzymatic Hydrolysis of Fish Industry Waste Using Different Commercial Proteases. Appl. Biochem. Biotechnol. 2011, 164, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, J.A.; Meduíña, A.; Durán, A.I.; Nogueira, M.; Fernández-Compás, A.; Pérez-Martín, R.I.; Rodríguez-Amado, I. Production of Valuable Compounds and Bioactive Metabolites from By-Products of Fish Discards Using Chemical Processing, Enzymatic Hydrolysis, and Bacterial Fermentation. Mar. Drugs 2019, 17, 139. [Google Scholar] [CrossRef] [Green Version]
- Sinanoglou, V.; Houhoula, D.; Kyrana, V.; Lougovois, V. Visceral oil from farmed Sparus aurata, Dicentrarchus labrax and Diplodus puntazzo as a source of ω-3 PUFA. Czech J. Food Sci. 2017, 35, 414–423. [Google Scholar] [CrossRef] [Green Version]
- Kandyliari, A.; Mallouchos, A.; Papandroulakis, N.; Golla, J.P.; Lam, T.T.; Sakellari, A.; Karavoltsos, S.; Vasiliou, V.; Kapsokefalou, M. Nutrient Composition and Fatty Acid and Protein Profiles of Selected Fish By-Products. Foods 2020, 9, 190. [Google Scholar] [CrossRef] [Green Version]
- Lenas, D.S.; Triantafillou, D.J.; Chatziantoniou, S.; Nathanailides, C. Fatty acid profile of wild and farmed gilthead sea bream (Sparus aurata). J. Consum. Prot. Food Saf. 2011, 6, 435–440. [Google Scholar] [CrossRef]
- Gelibolu, S.; Yanar, Y.; Genc, M.A.; Genç, E. Effect of Mannan-oligosaccharide Supplementation on Body Growth, Fatty Acid Profile and Organ Morphology of Gilthead Seabream, Sparus aurata. Pak. J. Zool. 2018, 50, 229–240. [Google Scholar] [CrossRef]
- Grigorakis, K. Compositional and organoleptic quality of farmed and wild gilthead sea bream (Sparus aurata) and sea bass (Dicentrarchus labrax) and factors affecting it: A review. Aquaculture 2007, 272, 55–75. [Google Scholar] [CrossRef]
- Yıldız, M.; Eroldoğan, T.O.; Ofori-Mensah, S.; Engin, K.; Baltacı, M.A. The effects of fish oil replacement by vegetable oils on growth performance and fatty acid profile of rainbow trout: Re-feeding with fish oil finishing diet improved the fatty acid composition. Aquaculture 2018, 488, 123–133. [Google Scholar] [CrossRef]
- Benedito-Palos, L.; Bermejo-Nogales, A.; Karampatos, A.I.; Ballester-Lozano, G.F.; Navarro, J.C.; Diez, A.; Bautista, J.M.; Bell, J.G.; Tocher, D.R.; Obach, A.; et al. Modelling the predictable effects of dietary lipid sources on the fillet fatty acid composition of one-year-old gilthead sea bream (Sparus aurata L.). Food Chem. 2011, 124, 538–544. [Google Scholar] [CrossRef] [Green Version]
- Honold, P.J.; Nouard, M.-L.; Jacobsen, C. Fish oil extracted from fish-fillet by-products is weakly linked to the extraction temperatures but strongly linked to the omega-3 content of the raw material. Eur. J. Lipid Sci. Technol. 2015, 118, 874–884. [Google Scholar] [CrossRef]
- Sathivel, S.; Prinyawiwatkul, W.; King, J.M.; Grimm, C.C.; Lloyd, S. Oil production from catfish viscera. J. Am. Oil Chem. Soc. 2003, 80, 377–382. [Google Scholar] [CrossRef]
- Chantachum, S. Separation and quality of fish oil from precooked and non-precooked tuna heads. Food Chem. 2000, 69, 289–294. [Google Scholar] [CrossRef]
- Genç, ÍY.; Esteves, E.; Aníbal, J.; Diler, A. Effects of different thawing methods on the quality of meagre fillets. Ankara Üniv. Vet. Fak. Derg. 2015, 62, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization of the United Nations. Codex Alimentarius Commision Standard for Fish Oil. In Codex Stan; Food and Agriculture Organization of the United Nations: Rome, Italy, 2017; p. 329. [Google Scholar]
- Yin, H.; Solval, K.M.; Huang, J.; Bechtel, P.J.; Sathivel, S. Effects of Oil Extraction Methods on Physical and Chemical Properties of Red Salmon Oils (Oncorhynchus nerka). J. Am. Oil Chem. Soc. 2011, 88, 1641–1648. [Google Scholar] [CrossRef]
- Eke-Ejiofor, J.A. Effect of Extraction Methods on the Quality Characteristics of Catfish (Clarias gariepinus) Oil. Am. J. Food Sci. Technol. 2018, 6, 199–203. [Google Scholar] [CrossRef]
- CIE. Commission Internationale de l’Eclairage Colorimetry, 3rd ed.; CIE 0152004 Color; CIE: Tiruchirappalli, India, 2004; Volume 15, pp. 1–82. [Google Scholar]
- Shabanpour, B.; Asghari, M.; Heydari, S.; Baee, H.; Ghorbani, A.; Jafar, A. Comparing of qualitative changes among the carps culturing in a pond, an under controlled place and marine carp during refrigeration. J. Anim. Res. Iran. J. Biol. 2016, 28, 466–480. [Google Scholar]
- Sabzipour, F.; Naseri, M.; Babaei, S.; Imani, A. Effect of various postmortem processing times and blanching methods on quality of rainbow trout (Oncorhynchus mykiss) waste oil. Food Sci. Nutr. 2019, 7, 3093–3102. [Google Scholar] [CrossRef] [Green Version]
- Franklin, E.C.; Haq, M.; Roy, V.C.; Park, J.; Chun, B. Supercritical CO2 extraction and quality comparison of lipids from Yellowtail fish (Seriola quinqueradiata) waste in different conditions. J. Food Process. Preserv. 2020, 44, 1–12. [Google Scholar] [CrossRef]
- Yves, H.; Korma, S.A.; Ali, A.H.; Tuyishime, M.A.; Habinshuti, I.; Abed, S.M. Extraction, Refining and Purification of ω-3 PUFA through Different Techniques—A Review. Am. J. Food Sci. Nutr. Res. 2016, 4, 18–26. [Google Scholar] [CrossRef]
- European Food Safety Authority. Scientific Opinion on Fish Oil for Human Consumption. Food Hygiene, including Rancidity. EFSA J. 2010, 8, 1–48. [Google Scholar] [CrossRef]
- Chakraborty, K.; Joseph, D. Production and Characterization of Refined Oils Obtained from Indian Oil Sardine (Sardinella longiceps). J. Agric. Food Chem. 2015, 63, 998–1009. [Google Scholar] [CrossRef]
- Kuo, C.-H.; Liao, H.-Z.; Wang, Y.-H.; Wang, H.-M.D.; Shieh, C.-J.; Tseng, C.-Y. Highly efficient extraction of EPA/DHA-enriched oil from cobia liver using homogenization plus sonication. Eur. J. Lipid Sci. Technol. 2017, 119, 1–8. [Google Scholar] [CrossRef]
- FAO. A Joint FAO/WHO Food Standards Programme Codex Committee on Fats and Oils. Codex Alimentarius Commission, Procedural Manual, 20th ed.; FAO: Rome, Italy, 2011. [Google Scholar]
- Soldo, B.; Šimat, V.; Vlahović, J.; Skroza, D.; Ljubenkov, I.; Mekinić, I.G. High Quality Oil Extracted from Sardine By-Products as an Alternative to Whole Sardines: Production and Refining. Eur. J. Lipid Sci. Technol. 2019, 121, 1–10. [Google Scholar] [CrossRef]
- Mei, L.; Wang, L.; Li, Q.; Yu, J.; Xu, X. Comparison of acid degumming and enzymatic degumming process forSilybum marianumseed oil. J. Sci. Food Agric. 2013, 93, 2822–2828. [Google Scholar] [CrossRef]
- Vaisali, C.; Charanyaa, S.; Belur, P.D.; Regupathi, I. Refining of edible oils: A critical appraisal of current and potential technologies. Int. J. Food Sci. Technol. 2014, 50, 13–23. [Google Scholar] [CrossRef]
- Chaiyasit, W.; Elias, R.J.; McClements, D.J.; Decker, E.A. Role of Physical Structures in Bulk Oils on Lipid Oxidation. Crit. Rev. Food Sci. Nutr. 2007, 47, 299–317. [Google Scholar] [CrossRef] [PubMed]
- García-Moreno, P.J.; Guadix, A.; Gómez-Robledo, L.; Melgosa, M.; Guadix, E.M. Optimization of bleaching conditions for sardine oil. J. Food Eng. 2013, 116, 606–612. [Google Scholar] [CrossRef]
- Afaq, F.; Malik, A.; Syed, D.; Maes, D.; Matsui, M.S.; Mukhtar, H. Pomegranate Fruit Extract Modulates UV-B–mediated Phosphorylation of Mitogen-activated Protein Kinases and Activation of Nuclear Factor Kappa B in Normal Human Epidermal Keratinocytes. Photochem. Photobiol. 2005, 81, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.C.M.; Miller, M.R. Purification of Alaskan Walleye Pollock (Gadus chalcogrammus) and New Zealand Hoki (Macruronus novaezelandiae) Liver Oil Using Short Path Distillation. Nutrients 2014, 6, 2059–2076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solaesa, Á.G.; Sanz, M.T.; Falkeborg, M.; Beltrán, S.; Guo, Z. Production and concentration of monoacylglycerols rich in omega-3 polyunsaturated fatty acids by enzymatic glycerolysis and molecular distillation. Food Chem. 2016, 190, 960–967. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.-H.; Hwang, L.S. Fractionation of squid visceral oil ethyl esters by short-path distillation. J. Am. Oil Chem. Soc. 2000, 77, 773–777. [Google Scholar] [CrossRef]
- Breivik, H.; Haraldsson, G.G.; Kristinsson, B. Preparation of highly purified concentrates of eicosapentaenoic acid and docosahexaenoic acid. J. Am. Oil Chem. Soc. 1997, 74, 1425–1429. [Google Scholar] [CrossRef]
- Valverde, L.M.; Moreno, P.A.G.; Callejón, M.J.J.; Cerdán, L.E.; Medina, A.R. Concentration of eicosapentaenoic acid (EPA) by selective alcoholysis catalyzed by lipases. Eur. J. Lipid Sci. Technol. 2013, 115, 990–1004. [Google Scholar] [CrossRef]
- Wang, W.; Li, T.; Ning, Z.; Wang, Y.; Yang, B.; Ma, Y.; Yang, X. A process for the synthesis of PUFA-enriched triglycerides from high-acid crude fish oil. J. Food Eng. 2012, 109, 366–371. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Xie, D.; Zou, S.; Jin, Q.; Wang, X. Synthesis and concentration of 2-monoacylglycerols rich in polyunsaturated fatty acids. Food Chem. 2018, 250, 60–66. [Google Scholar] [CrossRef]
- Fard, S.G.; Wang, F.; Sinclair, A.J.; Elliott, G.; Turchini, G.M. How does high DHA fish oil affect health? A systematic review of evidence. Crit. Rev. Food Sci. Nutr. 2019, 59, 1684–1727. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.; Huang, X.; Zhen, J.; Van Halm-Lutterodt, N.; Wang, J.; Zhou, C.; Yuan, L. Dietary Vitamin E Status Dictates Oxidative Stress Outcomes by Modulating Effects of Fish Oil Supplementation in Alzheimer Disease Model APPswe/PS1dE9 Mice. Mol. Neurobiol. 2018, 55, 9204–9219. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.-C.; Chen, H.-Y. Effects of different fatty acids on cell differentiation and lipid accumulation in preadipocytes of warm water fish cobia (Rachycentron canadum Linnaeus, 1766). Aquac. Res. 2013, 46, 590–601. [Google Scholar] [CrossRef]
- Vegusdal, A.; Sundvold, H.; Gjøen, T.; Ruyter, B. An in vitro method for studying the proliferation and differentiation of Atlantic salmon preadipocytes. Lipids 2003, 38, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Santulli, A.; Messina, C.M.; D’Amelio, V. Variations of lipid and apolipoprotein content in lipoproteins during fasting in European sea bass (Dicentrarchus labrax L.). Comp. Biochem. Physiol. Part A Physiol. 1997, 118, 1233–1239. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis of AOAC International, 18th ed.; Horwitz, W., Ed.; AOAC International: Gaithersburg, MD, USA, 2005; ISBN 978-0-935584-75-2. [Google Scholar]
- AOAC. AOAC Association of Official Analytical Chemists Official Method, 981.10 Crude protein in meat block digestion method. J. AOAC Int. 1992, 65, 1339. [Google Scholar]
- Folch, J.; Lees, M.; Stanley, G.S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- AOCS. AOCS Peroxide value. In Official Methods and Recommended Practices of the American Oil Chemists’ Society Official Method Cd 8-53; AOCS Press: Champaign, IL, USA, 1998. [Google Scholar]
- Botsoglou, N.A.; Fletouris, D.J.; Papageorgiou, G.E.; Vassilopoulos, V.N.; Mantis, A.J.; Trakatellis, A.G. Rapid, Sensitive, and Specific Thiobarbituric Acid Method for Measuring Lipid Peroxidation in Animal Tissue, Food, and Feedstuff Samples. J. Agric. Food Chem. 1994, 42, 1931–1937. [Google Scholar] [CrossRef]
- AOCS. AOCS Sampling and Analysis of Commercial Fats and Oil: ρ-Anisidine Value, Official Method Cd 18-90; AOAC International: Gaithersburg, MD, USA, 1993. [Google Scholar]
- Holm, U. Abstracts, International Society for Fat Research Congress; ISF: Urbana, IL, USA, 1972. [Google Scholar]
- AOCS. AOCS Method Cd 3d-63. Acid Value. In Official Methods and Recommended Practices of the AOCS, 5th ed.; AOCS Press: Champaign, IL, USA, 1999. [Google Scholar]
- Aryee, A.N.A.; Simpson, B.K.; Phillip, L.E.; Cue, R.I. Effect of Temperature and Time on the Stability of Salmon Skin Oil During Storage. J. Am. Oil Chem. Soc. 2011, 89, 287–292. [Google Scholar] [CrossRef]
- Vazquez, L.; Akoh, C.C. Fractionation of Short and Medium Chain Fatty Acid Ethyl Esters from a Blend of Oils via Ethanolysis and Short-Path Distillation. J. Am. Oil Chem. Soc. 2010, 87, 917–928. [Google Scholar] [CrossRef]
- Messina, C.M.; Manuguerra, S.; Renda, G.; Santulli, A. Biotechnological Applications for the Sustainable Use of Marine By-products: In Vitro Antioxidant and Pro-apoptotic Effects of Astaxanthin Extracted with Supercritical CO2 from Parapeneus longirostris. Mar. Biotechnol. 2019, 21, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Abbes, M.; Baati, H.; Guermazi, S.; Messina, C.; Santulli, A.; Gharsallah, N.; Ammar, E. Biological properties of carotenoids extracted from Halobacterium halobium isolated from a Tunisian solar saltern. BMC Complement. Altern. Med. 2013, 13, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manuguerra, S.; Caccamo, L.; Mancuso, M.; Arena, R.; Rappazzo, A.C.; Genovese, L.; Santulli, A.; Messina, C.M.; Maricchiolo, G. The antioxidant power of horseradish, Armoracia rusticana, underlies antimicrobial and antiradical effects, exerted in vitro. Nat. Prod. Res. 2018, 34, 1567–1570. [Google Scholar] [CrossRef] [PubMed]
- Messina, C.M.; Troia, A.; Arena, R.; Manuguerra, S.; Ioannou, T.; Curcuraci, E.; Renda, G.; Hellio, C.; Santulli, A. Species-Specific Antioxidant Power and Bioactive Properties of the Extracts Obtained from Wild Mediterranean Calendula Spp. (Asteraceae). Appl. Sci. 2019, 9, 4627. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Ljubojević, D.B.; Pelic, M.; Novakov, N.; Lazić, S.; Lupulović, D.; Ćirković, M.; Todorčević, M. The establishment of the cell culture of preadipocytes of common carp (Cyprinus carpio L.). Food Feed. Res. 2014, 41, 103–108. [Google Scholar] [CrossRef]
- Underwood, A.J. Experiments in Ecology. Their Logical Design and Interpretation Using Analysis of Variance; Cambridge University Press: Cambridge, UK, 1997; ISBN 0521556961. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | g/100 g SBV |
|---|---|
| Lipid | 51.79 ± 12.92 |
| Moisture | 40.81 ± 4.86 |
| Protein | 5.67 ± 0.02 |
| Ash | 1.43 ± 0.55 |
| Fatty Acids | % |
|---|---|
| 14:0 | 4.99 ± 0.17 |
| 16:0 | 15.56 ± 0.18 |
| 16:1n−7 | 10.72 ± 0.18 |
| 16:2n−4 | 0.89 ± 0.03 |
| 16:3n−4 | 0.59 ± 0.02 |
| 18:0 | 2.96 ± 0.06 |
| 18:1n−9 | 16.11 ± 0.40 |
| 18:1n−7 | 6.05 ± 0.15 |
| 18:2n−6 | 2.42 ± 0.16 |
| 18:3n−4 | 0.01 ± 0.00 |
| 18:3n−3 | 0.62 ± 0.04 |
| 18:4n−3 | 1.68 ± 0.06 |
| 20:1n−9 | 11.14 ± 0.27 |
| 20:4n−6 | 0.13 ± 0.03 |
| 20:4n−3 | 0.38 ± 0.08 |
| EPA | 8.19 ± 0.12 |
| 22:1n−11 | 4.71 ± 0.17 |
| 22:1n−9 | 0.38 ± 0.02 |
| 22:5n−3 | 1.19 ± 0.08 |
| DHA | 11.27 ± 0.30 |
| SFA | 23.51 ± 0.28 |
| MUFA | 49.12 ± 0.76 |
| PUFA | 27.38 ± 0.48 |
| Sample | Parameters | ||
|---|---|---|---|
| Temperatures | Times | PV | TBARS |
| 40 | 10 | 6.17 ± 0.47 b | 16.01 ± 0.02 b |
| 30 | 6.51 ± 0.22 b | 15.59 ± 0.03 b | |
| 60 | 10.29 ± 2.37 c | 21.18 ± 5.85 bc | |
| 60 | 10 | 5.42 ± 0.13 b | 14.15 ± 0.07 b |
| 30 | 6.38 ± 0.03 b | 16.58 ± 0.01 b | |
| 60 | 11.33 ± 0.75 cd | 14.31 ± 0.22 b | |
| 80 | 10 | 11.01 ± 1.77 cd | 17.50 ± 2.41 b |
| 30 | 13.27 ± 2.13 d | 25.38 ± 8.59 c | |
| 60 | 19.69 ± 0.90 e | 19.89 ± 2.29 bc | |
| 90 | 10 | 10.99 ± 0.53 cd | 31.41 ± 6.33 d |
| 30 | 19.67 ± 2.10 e | 33.09 ± 1.31 d | |
| 60 | 22.74 ± 2.27 f | 33.43 ± 5.63 d | |
| CO | 2.10 ± 0.53 a | 5.51 ± 0.81 a | |
| Samples | SFA | MUFA | PUFA | EPA | DHA | |
|---|---|---|---|---|---|---|
| SBV | 23.51 ± 0.28 a | 49.12 ± 0.76 ab | 27.38 ± 0.48 e | 8.19 ± 0.12 eg | 11.27± 0.30 e | |
| CVO | ||||||
| Temperature | Time | |||||
| 40 | 10 | 24.98 ± 1.03 ab | 50.09 ± 0.66 b | 24.93 ± 0.37 bcd | 7.12 ± 0.52 bcd | 10.15 ± 0.06 cd |
| 30 | 24.26± 0.58 a | 50.28 ± 0.15 b | 25.46 ± 0.58 cde | 7,78 ± 0.18 def | 10.03 ± 0.36 cd | |
| 60 | 26.61 ± 1.35 bc | 49.81 ± 0.28 b | 23.58 ±1.64 bc | 6.91 ± 0.51 bc | 9.25 ± 1.00 bc | |
| 60 | 10 | 25.28 ± 0.01 ab | 49.84 ± 0.11 b | 24.88 ± 0.10 bcd | 7.54 ± 0.05 cdef | 9.69 ± 0.02 bcd |
| 30 | 25.69 ± 1.41 abc | 49.43 ± 0.06 b | 24.88 ± 1.36 bcd | 7.71 ± 0.56 def | 9.61 ± 0.49 bcd | |
| 60 | 25.17 ± 0.79 ab | 48.86 ± 0.49 b | 25.97 ± 1.28 de | 7.92 ± 0.69 ef | 9.85 ± 0.67 cd | |
| 80 | 10 | 26.62 ± 1.66 bc | 47.41± 0.69 a | 25.97 ± 2.35 de | 7.56 ± 0.35 cdef | 10.49 ± 1.37 de |
| 30 | 27.23 ± 0.80 c | 49.83 ± 0.42 b | 22.94 ± 0.37 b | 6.73 ± 0.08 b | 8.71 ± 0.37 ab | |
| 60 | 24.78 ± 0.77 a | 49.42 ± 0.38 b | 25.79 ± 1.15 cde | 7.38 ± 0.56 bcde | 10.61 ± 0.54 de | |
| 90 | 10 | 24.48 ± 0.62 a | 48.54 ± 0.33 ab | 26.98 ± 0.97 de | 8.64 ± 0.42 g | 10.40 ± 0.52 cde |
| 30 | 25.25 ± 0.76 ab | 48.72 ± 0.39 b | 25.76 ± 0.84 cde | 7.44 ± 0.30 cdef | 9.68 ± 0.45 bcd | |
| 60 | 26.69 ± 0.21 bc | 52.30 ± 2.75 c | 21.02 ± 1.90 a | 6.09 ± 0.45 a | 7.97 ± 0.43 a | |
| Parameters | |||||||
|---|---|---|---|---|---|---|---|
| Samples | CVO Extraction Temperature | PV (meqO2/kg) | ρ-AV | TBARS (MDA µg/g) | TOTOX | Phospholipids (mg kg−1) | Acid Value (%FFA) |
| CVO | 60 °C | 5.41 ± 0.14 b | 28.98 ± 0.90 d | 14.15 ± 0.07 d | 39.80 ± 1.18 c | 24.75 ± 1.61 d | 8.23 ± 0.82 d |
| RVO | 3.90 ± 1.15 b | 13.49 ± 0.20 b | 6.62 ± 0.39 b | 21.30 ± 2.50 b | 8.00 ± 1.10 a | 1.74 ± 0.33 b | |
| CVO | 80 °C | 11.17 ± 1.31 d | 34.56 ± 2.36 e | 17.50 ± 2.41 e | 56.90 ± 0.26 e | 47.47 ± 14.05 e | 7.41 ± 0.49 c |
| RVO | 8.63 ± 0.31 c | 24.06 ± 1.13 c | 8.96 ± 0.86 c | 41.32 ± 0.51 d | 14.42 ± 3.90 c | 6.62 ± 0.74 c | |
| CO | 2.10 ± 0.53 a | 4.96 ± 0.89 a | 5.51 ± 0.81 a | 10.83 ± 0.24 a | 10.74 ± 2.45 b | 0.50 ± 0.04 a | |
| Fatty Acids | TFA | PUFAE | EFA |
|---|---|---|---|
| 14:0 | 5.07 ± 0.57 b | 0.66 ± 0.28 a | 4.35 ± 0.05 b |
| 16:0 | 11.47 ± 0.96 b | 4.45 ± 0.46 a | 11.26 ± 0.03 b |
| 16:1n−7 | 8.63 ± 2.10 b | 2.43 ± 0.16 a | 7.48 ± 0.02 b |
| 16:2n−4 | 0.86 ± 0.02 b | 0.38 ± 0.21 a | 1.01 ± 0.06 b |
| 16:3n−4 | 0.48 ± 0.06 b | 0.22 ± 0.12 a | 0.65 ± 0.08 b |
| 18:0 | 2.24 ± 0.19 | 2.15 ± 0.28 | 2.36 ± 0.23 |
| 18:1n−9 | 16.58 ± 0.87 b | 14.72 ± 0.77 a | 18.32 ± 0.27 b |
| 18:1n−7 | 5.71 ± 1.07 | 3.90 ± 0.66 | 5.27 ± 0.57 |
| 18:2n−6 | 1.48 ± 0.13 | 1.17 ± 0.75 | 1.78 ± 0.06 |
| 18:3n−4 | 0.01 ± 0.01 a | 0.02 ± 0.01 a | 0.17 ± 0.00 b |
| 18:3n−3 | 0.69 ± 0.11 a | 0.60 ± 0.19 a | 0.95 ± 0.02 b |
| 18:4n−3 | 2.23 ± 0.11 b | 1.61 ± 0.10 a | 2.48 ± 0.14 c |
| 20:1n−9 | 12.67 ± 0.83 b | 5.51 ± 0.95 a | 11.44 ± 0.05 b |
| 20:3n−3 | 0.22 ± 0.19 | 0.48 ± 0.18 | 0.43 ± 0.00 |
| 20:4n−6 | 0.16 ± 0.08 | 0.20 ± 0.14 | 0.26 ± 0.00 |
| 20:4n−3 | 0.53 ± 0.11 a | 1.09 ± 0.25 b | 0.97 ± 0.01 b |
| EPA-20:5n−3 | 8.80 ± 0.63 a | 13.92 ± 1.05 b | 9.28 ± 0.03 a |
| 22:1n−11 | 8.33 ± 1.95 b | 6.67 ± 0.28 a | 8.31 ± 0.19 b |
| 22:1n−9 | 0.59 ± 0.39 a | 2.96 ± 0.38 b | 1.03 ± 0.05 a |
| 22:5n−3 | 1.39 ± 0.11 a | 3.97 ± 0.42 b | 1.40 ± 0.05 a |
| DHA-22:6n−3 | 11.84 ± 0.78 a | 32.90 ± 0.76 b | 10.78 ± 0.19 a |
| SFA | 18.78 ± 1.29 a | 7.27 ± 0.17 b | 17.97 ± 0.19 a |
| MUFA | 52.51 ± 0.95 b | 36.19 ± 0.88 a | 51.84 ± 0.24 b |
| PUFA | 28.71 ± 2.11 a | 56.55 ± 0.93 b | 30.19 ± 0.14 a |
| R | 0.74 ± 0.05 a | 2.45 ± 0.20 b | 0.68 ± 0.01 a |
| EPA Enrichment Factor | 1.00 ± 0.07 a | 1.58 ± 0.12 b | 1.05 ± 0.00 a |
| DHA Enrichment Factor | 1.00 ± 0.07 a | 2.78 ± 0.06 b | 0.91 ± 0.02 a |
| PUFA enrichment facto | 1.00 ± 0.07 a | 1.97 ± 0.03 b | 1.05 ± 0.00 a |
| PUFA/SFA | 1.54 ± 0.21 a | 7.79 ± 0.26 b | 1.68 ± 0.02 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Messina, C.M.; Arena, R.; Manuguerra, S.; Renda, G.; Laudicella, V.A.; Ficano, G.; Fazio, G.; La Barbera, L.; Santulli, A. Farmed Gilthead Sea Bream (Sparus aurata) by-Products Valorization: Viscera Oil ω-3 Enrichment by Short-Path Distillation and In Vitro Bioactivity Evaluation. Mar. Drugs 2021, 19, 160. https://doi.org/10.3390/md19030160
Messina CM, Arena R, Manuguerra S, Renda G, Laudicella VA, Ficano G, Fazio G, La Barbera L, Santulli A. Farmed Gilthead Sea Bream (Sparus aurata) by-Products Valorization: Viscera Oil ω-3 Enrichment by Short-Path Distillation and In Vitro Bioactivity Evaluation. Marine Drugs. 2021; 19(3):160. https://doi.org/10.3390/md19030160
Chicago/Turabian StyleMessina, Concetta Maria, Rosaria Arena, Simona Manuguerra, Giuseppe Renda, Vincenzo Alessandro Laudicella, Giovanna Ficano, Gioacchino Fazio, Laura La Barbera, and Andrea Santulli. 2021. "Farmed Gilthead Sea Bream (Sparus aurata) by-Products Valorization: Viscera Oil ω-3 Enrichment by Short-Path Distillation and In Vitro Bioactivity Evaluation" Marine Drugs 19, no. 3: 160. https://doi.org/10.3390/md19030160