
Antioxidant Compounds from Microalgae: A Review


Abstract
:1. Introduction
2. Antioxidant and Reactive Oxygen Species (ROS)
3. The Antioxidants Molecules of Microalgae
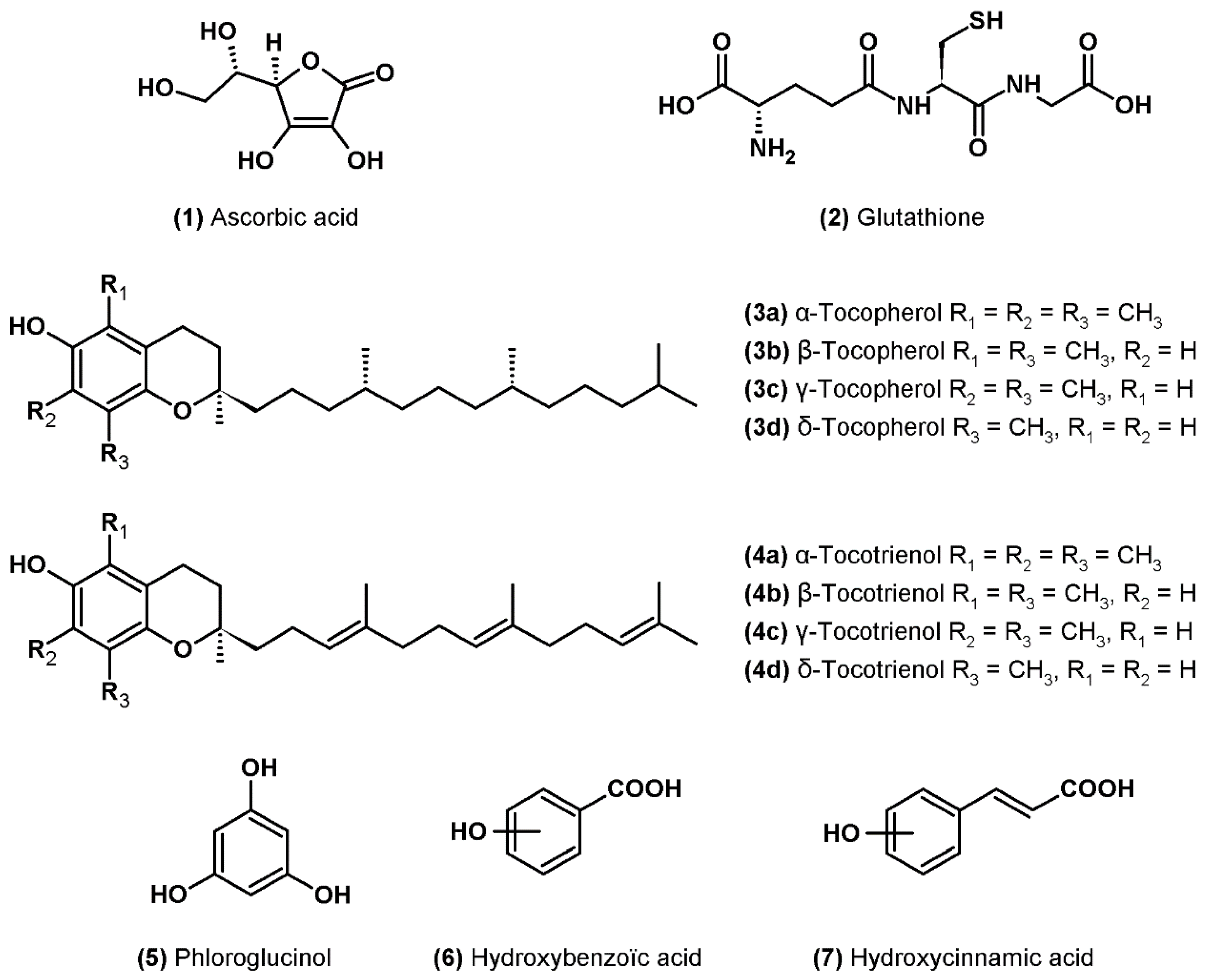
3.1. Ascorbic Acid
3.2. Glutathione
3.3. Tocopherols
3.4. Phenolic Compounds
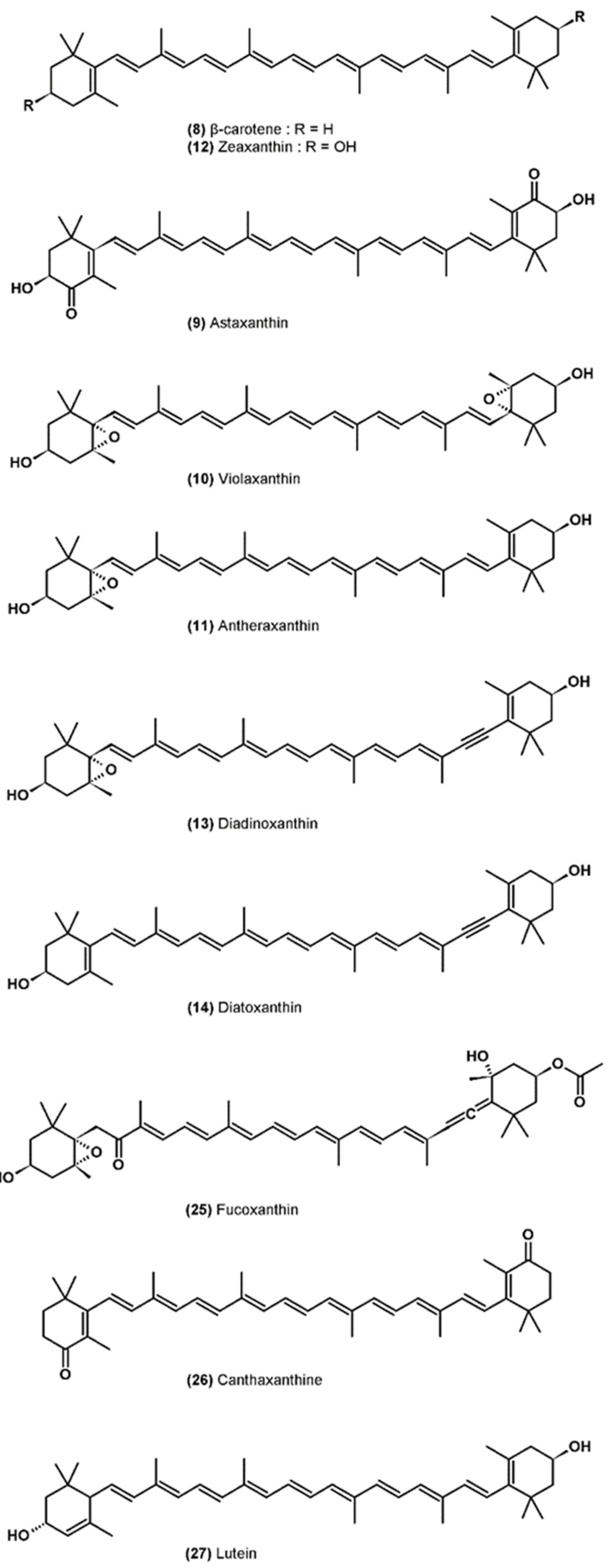
3.5. Carotenoids
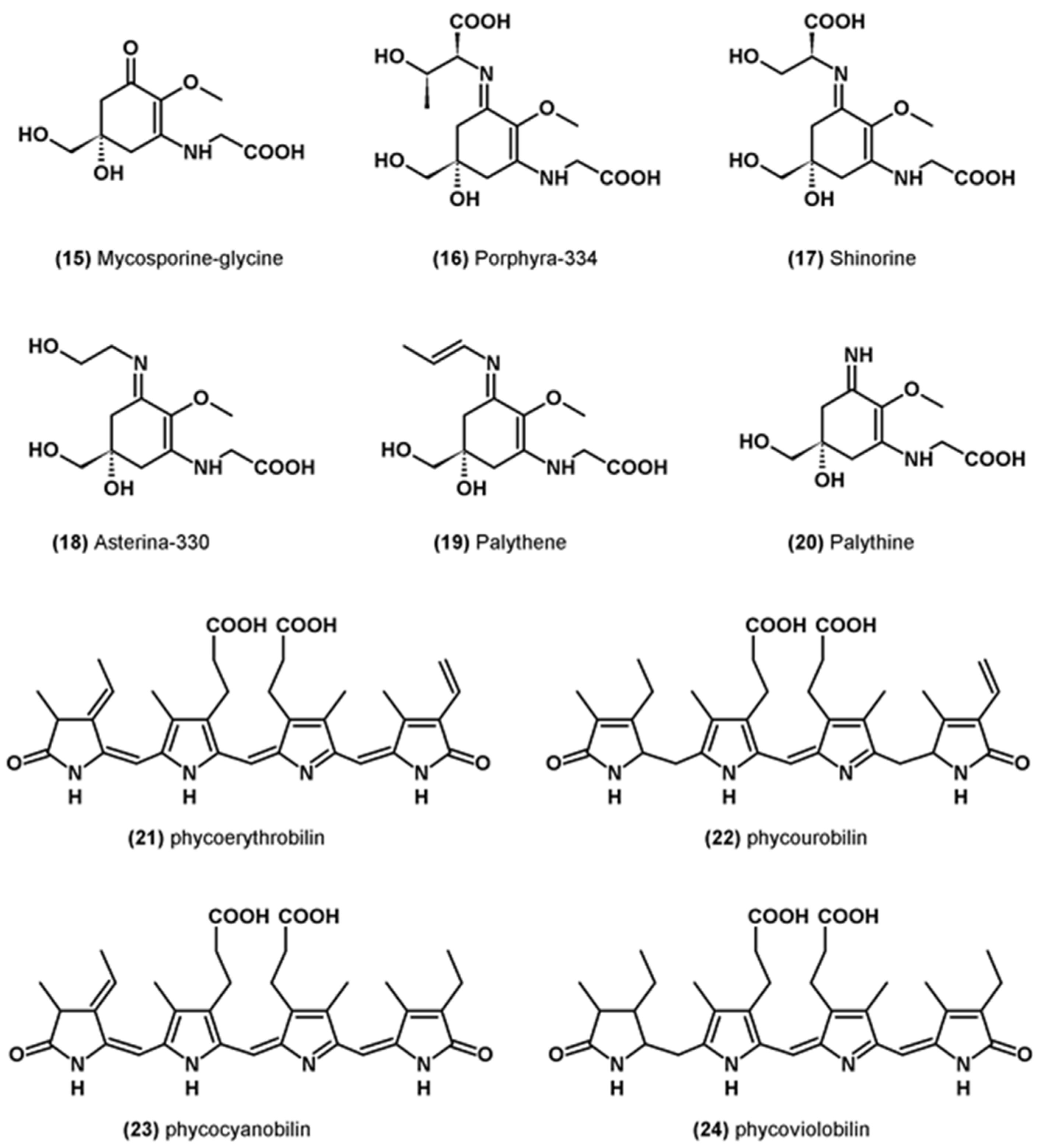
3.6. Miscellaneous Antioxidants
4. Common and Recent Assays Used to Evaluate Antioxidant Activity of Microalgae
5. Antioxidant Activity of Microalgae
6. Applications in the Food Industry
7. Conclusion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Mimouni, V.; Ulmann, L.; Pasquet, V.; Mathieu, M.; Picot, L.; Bougaran, G.; Cadoret, J.-P.; Morant-Manceau, A.; Schoefs, B. The Potential of Microalgae for the Production of Bioactive Molecules of Pharmaceutical Interest. Curr. Pharm. Biotechnol. 2012, 13, 2733–2750. [Google Scholar] [CrossRef]
- Wijffels, R.H.; Barbosa, M.J.; Eppink, M.H.M. Microalgae for the Production of Bulk Chemicals and Biofuels. Biofuels. Bioprod. Biorefining 2010, 4, 287–295. [Google Scholar] [CrossRef] [Green Version]
- Aklakur, M. Natural Antioxidants from Sea: A Potential Industrial Perspective in Aquafeed Formulation. Rev. Aquacult. 2016, 10, 385–399. [Google Scholar] [CrossRef]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as Sources of High Added-Value Compounds—a Brief Review of Recent Work. Biotechnol Prog. 2011, 27, 597–613. [Google Scholar] [CrossRef]
- Abad, M.J.; Bedoya, L.M.; Bermejo, P. Natural Marine Anti-Inflammatory Products. Mini-Rev. Med. Chem. 2008, 8, 740–754. [Google Scholar] [CrossRef]
- Assunção, M.F.G.; Amaral, R.; Martins, C.B.; Ferreira, J.D.; Ressurreição, S.; Santos, S.D.; Varejão, J.M.T.B.; Santos, L.M.A. Screening Microalgae as Potential Sources of Antioxidants. J. Appl. Phycol. 2016, 29, 865–877. [Google Scholar] [CrossRef]
- Coulombier, N.; Nicolau, E.; Le Déan, L.; Antheaume, C.; Jauffrais, T.; Lebouvier, N. Impact of Light Intensity on Antioxidant Activity of Tropical Microalgae. Mar. Drugs 2020, 18, 122. [Google Scholar] [CrossRef] [Green Version]
- Safafar, H.; van Wagenen, J.; Møller, P.; Jacobsen, C. Carotenoids, Phenolic Compounds and Tocopherols Contribute to the Antioxidative Properties of Some Microalgae Species Grown on Industrial Wastewater. Mar. Drugs 2015, 13, 7339–7356. [Google Scholar] [CrossRef] [Green Version]
- Sathasivam, R.; Ki, J.-S. A Review of the Biological Activities of Microalgal Carotenoids and Their Potential Use in Healthcare and Cosmetic Industries. Mar. Drugs 2018, 16, 26. [Google Scholar] [CrossRef] [Green Version]
- Goiris, K.; Muylaert, K.; Fraeye, I.; Foubert, I.; Brabanter, J.D.; Cooman, L.D. Antioxidant Potential of Microalgae in Relation to Their Phenolic and Carotenoid Content. J. Appl. Phycol. 2012, 24, 1477–1486. [Google Scholar] [CrossRef]
- Cezare-Gomes, E.A.; del Carmen Mejia-da-Silva, L.; Pérez-Mora, L.S.; Matsudo, M.C.; Ferreira-Camargo, L.S.; Singh, A.K.; de Carvalho, J.C.M. Potential of Microalgae Carotenoids for Industrial Application. Appl. Biochem. Biotechnol. 2019, 188, 602–634. [Google Scholar] [CrossRef]
- Ito, N.; Hirose, M.; Fukushima, S.; Tsuda, H.; Shirai, T.; Tatematsu, M. Studies on Antioxidants: Their Carcinogenic and Modifying Effects on Chemical Carcinogenesis. Food Chem. Toxicol. 1986, 24, 1071–1082. [Google Scholar] [CrossRef]
- Li, H.-B.; Cheng, K.-W.; Wong, C.-C.; Fan, K.-W.; Chen, F.; Jiang, Y. Evaluation of Antioxidant Capacity and Total Phenolic Content of Different Fractions of Selected Microalgae. Food Chem. 2007, 102, 771–776. [Google Scholar] [CrossRef]
- Safer, A.M.; Al-Nughamish, A.J. Hepatotoxicity Induced by the Anti-Oxidant Food Additive, Butylated Hydroxytoluene (BHT), in Rats: An Electron Microscopical Study. Histol. Histopathol. 1999, 14, 391–406. [Google Scholar]
- Batista, A.P.; Niccolai, A.; Fradinho, P.; Fragoso, S.; Bursic, I.; Rodolfi, L.; Biondi, N.; Tredici, M.R.; Sousa, I.; Raymundo, A. Microalgae Biomass as an Alternative Ingredient in Cookies: Sensory, Physical and Chemical Properties, Antioxidant Activity and in Vitro Digestibility. Algal Res. 2017, 26, 161–171. [Google Scholar] [CrossRef]
- Batista, A.P.; Niccolai, A.; Bursic, I.; Sousa, I.; Raymundo, A.; Rodolfi, L.; Biondi, N.; Tredici, M.R. Microalgae as Functional Ingredients in Savory Food Products: Application to Wheat Crackers. Foods 2019, 8, 611. [Google Scholar] [CrossRef] [Green Version]
- Goiris, K.; Muylaert, K.; De Cooman, L. Microalgae as a Novel Source of Antioxidants for Nutritional Applications. In Handbook of Marine Microalgae; Kim, S.-K., Ed.; Academic Press: Boston, MA, USA, 2015; pp. 269–280. ISBN 978-0-12-800776-1. [Google Scholar]
- Sansone, C.; Brunet, C. Promises and Challenges of Microalgal Antioxidant Production. Antioxidants 2019, 8, 199. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B. How to Characterize an Antioxidant: An Update. Biochem. Soc. Symp. 1995, 61, 73–101. [Google Scholar] [CrossRef]
- Chen, B.; Wan, C.; Mehmood, M.A.; Chang, J.-S.; Bai, F.; Zhao, X. Manipulating Environmental Stresses and Stress Tolerance of Microalgae for Enhanced Production of Lipids and Value-Added Products–A Review. Bioresour. Technol. 2017, 244, 1198–1206. [Google Scholar] [CrossRef]
- Coulombier, N.; Blanchier, P.; Le Dean, L.; Barthelemy, V.; Lebouvier, N.; Jauffrais, T. The Effects of CO2-Induced Acidification on Tetraselmis Biomass Production, Photophysiology and Antioxidant Activity: A Comparison Using Batch and Continuous Culture. J. Biotechnol. 2021, 325, 312–324. [Google Scholar] [CrossRef]
- Coulombier, N.; Nicolau, E.; Le Déan, L.; Barthelemy, V.; Schreiber, N.; Brun, P.; Lebouvier, N.; Jauffrais, T. Effects of Nitrogen Availability on the Antioxidant Activity and Carotenoid Content of the Microalgae Nephroselmis sp. Mar. Drugs 2020, 18, 453. [Google Scholar] [CrossRef]
- Gauthier, M.R.; Senhorinho, G.N.A.; Scott, J.A. Microalgae under Environmental Stress as a Source of Antioxidants. Algal Res. 2020, 52, 102104. [Google Scholar] [CrossRef]
- Goiris, K.; Van Colen, W.; Wilches, I.; León-Tamariz, F.; De Cooman, L.; Muylaert, K. Impact of Nutrient Stress on Antioxidant Production in Three Species of Microalgae. Algal Res. 2015, 7, 51–57. [Google Scholar] [CrossRef]
- Guedes, A.C.; Amaro, H.M.; Pereira, R.D.; Malcata, F.X. Effects of Temperature and PH on Growth and Antioxidant Content of the Microalga Scenedesmus obliquus. Biotechnol. Prog. 2011, 27, 1218–1224. [Google Scholar] [CrossRef]
- Paliwal, C.; Mitra, M.; Bhayani, K.; Bharadwaj, V.S.V.; Ghosh, T.; Dubey, S.; Mishra, S. Abiotic Stresses as Tools for Metabolites in Microalgae. Bioresour. Technol. 2017, 244, 1216–1226. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Diplock, A.T. Current Status of Antioxidant Therapy. Free Radic. Biol. Med. 1993, 15, 77–96. [Google Scholar] [CrossRef]
- Saed-Moucheshi, A.; Shekoofa, A.; Pessarakli, M. Reactive Oxygen Species (ROS) Generation and Detoxifying in Plants. J. Plant. Nutr. 2014, 37, 1573–1585. [Google Scholar] [CrossRef]
- Cirulis, J.T.; Scott, J.A.; Ross, G.M. Management of Oxidative Stress by Microalgae. Can. J. Physiol. Pharmacol. 2013, 91, 15–21. [Google Scholar] [CrossRef]
- Demidchik, V. Mechanisms of Oxidative Stress in Plants: From Classical Chemistry to Cell Biology. Environ. Exp. Bot. 2015, 109, 212–228. [Google Scholar] [CrossRef]
- Triantaphylidès, C.; Havaux, M. Singlet Oxygen in Plants: Production, Detoxification and Signaling. Trends Plant. Sci. 2009, 14, 219–228. [Google Scholar] [CrossRef]
- Telfer, A.; Pascal, A.; Gall, A. Carotenoids in Photosynthesis. In Carotenoids: Volume 4: Natural Functions; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser: Basel, Switzerland, 2008; pp. 265–308. ISBN 978-3-7643-7499-0. [Google Scholar]
- Gill, S.S.; Tuteja, N. Reactive Oxygen Species and Antioxidant Machinery in Abiotic Stress Tolerance in Crop Plants. Plant. Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Ledford, H.K.; Niyogi, K.K. Singlet Oxygen and Photo-Oxidative Stress Management in Plants and Algae. Plant. Cell Environ. 2005, 28, 1037–1045. [Google Scholar] [CrossRef]
- Pospíšil, P.; Yamamoto, Y. Damage to Photosystem II by Lipid Peroxidation Products. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 457–466. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 5th ed.; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 21703. [Google Scholar] [CrossRef] [Green Version]
- Richards, S.L.; Wilkins, K.A.; Swarbreck, S.M.; Anderson, A.A.; Habib, N.; Smith, A.G.; McAinsh, M.; Davies, J.M. The Hydroxyl Radical in Plants: From Seed to Seed. J. Exp. Bot. 2015, 66, 37–46. [Google Scholar] [CrossRef]
- Gulcin, İ. Antioxidants and Antioxidant Methods: An Updated Overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef] [Green Version]
- Duarte, T.L.; Lunec, J. When Is an Antioxidant Not an Antioxidant? A Review of Novel Actions and Reactions of Vitamin C. Free Radic. Res. 2005, 39, 671–686. [Google Scholar] [CrossRef]
- Herrera, E.; Barbas, C. Vitamin E: Action, Metabolism and Perspectives. J. Physiol. Biochem. 2001, 57, 43–56. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Alegre, L. The Function of Tocopherols and Tocotrienols in Plants. Crit. Rev. Plant. Sci. 2002, 21, 31–57. [Google Scholar] [CrossRef]
- Dai, J.; Mumper, R.J. Plant Phenolics: Extraction, Analysis and Their Antioxidant and Anticancer Properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Aremu, A.O.; Neményi, M.; Stirk, W.A.; Ördög, V.; van Staden, S. Manipulation of Nitrogen Levels and Mode of Cultivation Are Viable Methods to Improve the Lipid, Fatty Acids, Phytochemical Content, and Bioactivities in Chlorella minutissima. J. Phycol. 2015, 51, 659–669. [Google Scholar] [CrossRef]
- Custódio, L.; Justo, T.; Silvestre, L.; Barradas, A.; Duarte, C.V.; Pereira, H.; Barreira, L.; Rauter, A.P.; Alberício, F.; Varela, J. Microalgae of Different Phyla Display Antioxidant, Metal Chelating and Acetylcholinesterase Inhibitory Activities. Food Chem. 2012, 131, 134–140. [Google Scholar] [CrossRef]
- Hajimahmoodi, M.; Faramarzi, M.A.; Mohammadi, N.; Soltani, N.; Oveisi, M.R.; Nafissi-Varcheh, N. Evaluation of Antioxidant Properties and Total Phenolic Contents of Some Strains of Microalgae. J. Appl. Phycol. 2010, 22, 43–50. [Google Scholar] [CrossRef]
- Jayshree, A.; Jayashree, S.; Thangaraju, N. Chlorella vulgaris and Chlamydomonas reinhardtii: Effective Antioxidant, Antibacterial and Anticancer Mediators. Indian J. Pharm. Sci. 2016, 78, 575–581. [Google Scholar] [CrossRef] [Green Version]
- Aremu, A.O.; Masondo, N.A.; Molnár, Z.; Stirk, W.A.; Ördög, V.; Van Staden, S. Changes in Phytochemical Content and Pharmacological Activities of Three Chlorella Strains Grown in Different Nitrogen Conditions. J. Appl. Phycol. 2016, 28, 149–159. [Google Scholar] [CrossRef]
- Chaudhuri, D.; Ghate, N.B.; Deb, S.; Panja, S.; Sarkar, R.; Rout, J.; Mandal, N. Assessment of the Phytochemical Constituents and Antioxidant Activity of a Bloom Forming Microalgae Euglena tuba. Biol. Res. 2014, 47, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goiris, K.; Muylaert, K.; Voorspoels, S.; Noten, B.; Paepe, D.D.; Baart, G.J.E.; Cooman, L.D. Detection of Flavonoids in Microalgae from Different Evolutionary Lineages. J. Phycol. 2014, 50, 483–492. [Google Scholar] [CrossRef]
- Haoujar, I.; Cacciola, F.; Abrini, J.; Mangraviti, D.; Giuffrida, D.; Oulad El Majdoub, Y.; Kounnoun, A.; Miceli, N.; Fernanda Taviano, M.; Mondello, L.; et al. The Contribution of Carotenoids, Phenolic Compounds, and Flavonoids to the Antioxidative Properties of Marine Microalgae Isolated from Mediterranean Morocco. Molecules 2019, 24, 4037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López, A.; Rico, M.; Santana-Casiano, J.M.; González, A.G.; González-Dávila, M. Phenolic Profile of Dunaliella Tertiolecta Growing under High Levels of Copper and Iron. Environ. Sci. Pollut. Res. 2015, 22, 14820–14828. [Google Scholar] [CrossRef]
- Smerilli, A.; Balzano, S.; Maselli, M.; Blasio, M.; Orefice, I.; Galasso, C.; Sansone, C.; Brunet, C. Antioxidant and Photoprotection Networking in the Coastal Diatom Skeletonema marinoi. Antioxidants 2019, 8, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Wang, Y.; Sun, J.; Bian, F.; Chen, G.; Zhang, Y.; Bi, Y.; Wu, Y. Antioxidant Activity of Alcohol Aqueous Extracts of Crypthecodinium cohnii and Schizochytrium sp. J. Zhejiang Univ.-Sci. B 2017, 18, 797–806. [Google Scholar] [CrossRef] [Green Version]
- Shahidi, F.; Ambigaipalan, P. Phenolics and Polyphenolics in Foods, Beverages and Spices: Antioxidant Activity and Health Effects—A Review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Leopoldini, M.; Russo, N.; Toscano, M. The Molecular Basis of Working Mechanism of Natural Polyphenolic Antioxidants. Food Chem. 2011, 125, 288–306. [Google Scholar] [CrossRef]
- Pouvreau, J.-B.; Morançais, M.; Taran, F.; Rosa, P.; Dufossé, L.; Guérard, F.; Pin, S.; Fleurence, J.; Pondaven, P. Antioxidant and Free Radical Scavenging Properties of Marennine, a Blue-Green Polyphenols Pigment from the Diatom Haslea ostrearia (Gaillon/Bory) Simonsen Responsible for the Natural Greening of Cultured Oysters. J. Agric. Food Chem. 2008, 56, 6278–6286. [Google Scholar] [CrossRef]
- Takaichi, S. Carotenoids in Algae: Distributions, Biosyntheses and Functions. Mar. Drugs 2011, 9, 1101–1118. [Google Scholar] [CrossRef] [PubMed]
- Stahl, W.; Sies, H. Antioxidant Activity of Carotenoids. Mol. Asp. Med. 2003, 24, 345–351. [Google Scholar] [CrossRef]
- Solovchenko, A.E. Physiology and Adaptive Significance of Secondary Carotenogenesis in Green Microalgae. Russ. J. Plant. Physiol. 2013, 60, 167–176. [Google Scholar] [CrossRef]
- Huang, J.J.; Lin, S.; Xu, W.; Cheung, P.C.K. Occurrence and Biosynthesis of Carotenoids in Phytoplankton. Biotechnol. Adv. 2017, 35, 597–618. [Google Scholar] [CrossRef] [PubMed]
- Goss, R.; Jakob, T. Regulation and Function of Xanthophyll Cycle-Dependent Photoprotection in Algae. Photosynth. Res. 2010, 106, 103–122. [Google Scholar] [CrossRef] [PubMed]
- Jahns, P.; Holzwarth, A.R. The Role of the Xanthophyll Cycle and of Lutein in Photoprotection of Photosystem II. Biochim. Biophys. Acta 2012, 1817, 182–193. [Google Scholar] [CrossRef] [Green Version]
- Edge, R.; McGarvey, D.J.; Truscott, T.G. The Carotenoids as Antioxidants—A Review. J. Photochem. Photobiol. B Biol. 1997, 41, 189–200. [Google Scholar] [CrossRef]
- El-Agamey, A.; McGarvey, D.J. Carotenoid Radicals and Radical Ions. In Carotenoids; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser: Basel, Switzerland, 2008; Volume 4, pp. 119–154. ISBN 978-3-7643-7498-3. [Google Scholar]
- Ribeiro, D.; Freitas, M.; Silva, A.M.S.; Carvalho, F.; Fernandes, E. Antioxidant and Pro-Oxidant Activities of Carotenoids and Their Oxidation Products. Food Chem. Toxicol. 2018, 120, 681–699. [Google Scholar] [CrossRef]
- Wada, N.; Sakamoto, T.; Matsugo, S. Mycosporine-Like Amino Acids and Their Derivatives as Natural Antioxidants. Antioxidants 2015, 4, 603–646. [Google Scholar] [CrossRef]
- Llewellyn, C.A.; Airs, R.L. Distribution and Abundance of MAAs in 33 Species of Microalgae across 13 Classes. Mar. Drugs 2010, 8, 1273–1291. [Google Scholar] [CrossRef] [Green Version]
- Sinha, R.P.; Singh, S.P.; Häder, D.-P. Database on Mycosporines and Mycosporine-like Amino Acids (MAAs) in Fungi, Cyanobacteria, Macroalgae, Phytoplankton and Animals. J. Photochem. Photobiol. B Biol. 2007, 89, 29–35. [Google Scholar] [CrossRef]
- Raposo, M.F.D.J.; De Morais, R.M.S.C.; de Morais, B.A.M.M. Bioactivity and Applications of Sulphated Polysaccharides from Marine Microalgae. Mar. Drugs 2013, 11, 233–252. [Google Scholar] [CrossRef] [Green Version]
- Balavigneswaran, C.K.; Sujin Jeba Kumar, T.; Moses Packiaraj, R.; Veeraraj, A.; Prakash, S. Anti-Oxidant Activity of Polysaccharides Extracted from Isochrysis galbana Using RSM Optimized Conditions. Int. J. Biol. Macromol. 2013, 60, 100–108. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, H.; Guo, G.; Pu, Y.; Yan, B. The Isolation and Antioxidant Activity of Polysaccharides from the Marine Microalgae Isochrysis galbana. Carbohydr. Polym. 2014, 113, 22–31. [Google Scholar] [CrossRef]
- Tannin-Spitz, T.; Bergman, M.; Van-Moppes, D.; Grossman, S.; Arad, S. Antioxidant Activity of the Polysaccharide of the Red Microalga Porphyridium sp. J. Appl. Phycol. 2005, 17, 215–222. [Google Scholar] [CrossRef]
- Yu, M.; Chen, M.; Gui, J.; Huang, S.; Liu, Y.; Shentu, H.; He, J.; Fang, Z.; Wang, W.; Zhang, Y. Preparation of Chlorella vulgaris Polysaccharides and Their Antioxidant Activity in Vitro and in Vivo. Int. J. Biol. Macromol. 2019, 137, 139–150. [Google Scholar] [CrossRef]
- Zhong, Q.; Wei, B.; Wang, S.; Ke, S.; Chen, J.; Zhang, H.; Wang, H. The Antioxidant Activity of Polysaccharides Derived from Marine Organisms: An Overview. Mar. Drugs 2019, 17, 674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glazer, A.N. Phycobiliproteins—A Family of Valuable, Widely Used Fluorophores. J. Appl. Phycol. 1994, 6, 105–112. [Google Scholar] [CrossRef]
- Sonani, R.R.; Rastogi, R.P.; Madamwar, D. Antioxidant Potential of Phycobiliproteins: Role in Anti-Aging Research. Biochem. Anal. Biochem. 2015, 4, 1009–2161. [Google Scholar] [CrossRef] [Green Version]
- Carocho, M.; Ferreira, I.C.F.R. A Review on Antioxidants, Prooxidants and Related Controversy: Natural and Synthetic Compounds, Screening and Analysis Methodologies and Future Perspectives. Food Chem. Toxicol. 2013, 51, 15–25. [Google Scholar] [CrossRef]
- Frankel, E.N.; Meyer, A.S. The Problems of Using One-Dimensional Methods to Evaluate Multifunctional Food and Biological Antioxidants. J. Sci. Food Agric. 2000, 80, 1925–1941. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized Methods for the Determination of Antioxidant Capacity and Phenolics in Foods and Dietary Supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef]
- Plaza, M.; Santoyo, S.; Jaime, L.; García-Blairsy Reina, G.; Herrero, M.; Señoráns, F.J.; Ibáñez, E. Screening for Bioactive Compounds from Algae. J. Pharm. Biomed. Anal. 2010, 51, 450–455. [Google Scholar] [CrossRef]
- Aremu, A.O.; Masondo, N.A.; Stirk, W.A.; Ördög, V.; Staden, J.V. Influence of Culture Age on the Phytochemical Content and Pharmacological Activities of Five Scenedesmus Strains. J. Appl. Phycol. 2014, 26, 407–415. [Google Scholar] [CrossRef]
- Herrero, M.; Jaime, L.; Martín-Alvarez, P.J.; Cifuentes, A.; Ibáñez, E. Optimization of the Extraction of Antioxidants from Dunaliella salina Microalga by Pressurized Liquids. J. Agric. Food Chem. 2006, 54, 5597–5603. [Google Scholar] [CrossRef]
- Natrah, F.M.I.; Yusoff, F.M.; Shariff, M.; Abas, F.; Mariana, N.S. Screening of Malaysian Indigenous Microalgae for Antioxidant Properties and Nutritional Value. J. Appl. Phycol. 2007, 19, 711–718. [Google Scholar] [CrossRef]
- Simic, S.; Kosanic, M.; Rankovic, B. Evaluation of In Vitro Antioxidant and Antimicrobial Activities of Green Microalgae Trentepohlia umbrina. Not. Bot. Horti Agrobot. 2012, 40, 86–91. [Google Scholar] [CrossRef] [Green Version]
- Goh, S.-H.; Yusoff, F.M.; Loh, S.P. A Comparison of the Antioxidant Properties and Total Phenolic Content in a Diatom, Chaetoceros sp. and a Green Microalga, Nannochloropsis sp. J. Agric. Sci. 2010, 2, 123. [Google Scholar] [CrossRef] [Green Version]
- Guzmán, S.; Gato, A.; Calleja, J.M. Antiinflammatory, Analgesic and Free Radical Scavenging Activities of the Marine Microalgae Chlorella stigmatophora and Phaeodactylum tricornutum. Phytother. Res. 2001, 15, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Azizan, A.; Ahamad Bustamam, M.S.; Maulidiani, M.; Shaari, K.; Ismail, I.S.; Nagao, N.; Abas, F. Metabolite Profiling of the Microalgal Diatom Chaetoceros calcitrans and Correlation with Antioxidant and Nitric Oxide Inhibitory Activities via 1H NMR-Based Metabolomics. Mar. Drugs 2018, 16, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.-H.; Lee, J.-B.; Lee, K.-W.; Jeon, Y.-J. Antioxidant Properties of Tidal Pool Microalgae, Halochlorococcum porphyrae and Oltamannsiellopsis unicellularis from Jeju Island, Korea. Algae 2010, 25, 45–56. [Google Scholar] [CrossRef]
- Buono, S.; Langellotti, A.L.; Martello, A.; Bimonte, M.; Tito, A.; Carola, A.; Apone, F.; Colucci, G.; Fogliano, V. Biological Activities of Dermatological Interest by the Water Extract of the Microalga Botryococcus braunii. Arch. Derm. Res. 2012, 304, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.Ø.; Romano, G.; Ianora, A. Bioactivity Screening of Microalgae for Antioxidant, Anti-Inflammatory, Anticancer, Anti-Diabetes, and Antibacterial Activities. Front. Mar. Sci. 2016, 3, 68. [Google Scholar] [CrossRef] [Green Version]
- Karawita, R.; Senevirathne, M.; Athukorala, Y.; Affan, A.; Lee, Y.-J.; Kim, S.-K.; Lee, J.-B.; Jeon, Y.-J. Protective Effect of Enzymatic Extracts from Microalgae against DNA Damage Induced by H2O2. Mar. Biotechnol. 2007, 9, 479–490. [Google Scholar] [CrossRef]
- Affan, A.; Karawita, R.; Jeon, Y.-J.; Kim, B.-Y.; Lee, J.-B. Growth Characteristics, Bio-Chemical Composition and Antioxidant Activities of Benthic Diatom Grammatophora marina from Jeju Coast, Korea. Algae 2006, 21, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Agregán, R.; Munekata, P.E.S.; Franco, D.; Carballo, J.; Barba, F.J.; Lorenzo, J.M. Antioxidant Potential of Extracts Obtained from Macro- (Ascophyllum nodosum, Fucus vesiculosus and Bifurcaria bifurcata) and Micro-Algae (Chlorella vulgaris and Spirulina platensis) Assisted by Ultrasound. Medicines 2018, 5, 33. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, F.; Fanning, K.; Netzel, M.; Turner, W.; Li, Y.; Schenk, P.M. Profiling of Carotenoids and Antioxidant Capacity of Microalgae from Subtropical Coastal and Brackish Waters. Food Chem. 2014, 165, 300–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaro, H.M.; Fernandes, F.; Valentão, P.; Andrade, P.B.; Sousa-Pinto, I.; Malcata, F.X.; Guedes, A.C. Effect of Solvent System on Extractability of Lipidic Components of Scenedesmus obliquus (M2-1) and Gloeothece sp. on Antioxidant Scavenging Capacity Thereof. Mar. Drugs 2015, 13, 6453–6471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banskota, A.H.; Sperker, S.; Stefanova, R.; McGinn, P.J.; O’Leary, S.J.B. Antioxidant Properties and Lipid Composition of Selected Microalgae. J. Appl. Phycol. 2018, 31, 309–318. [Google Scholar] [CrossRef]
- Bauer, L.M.; Costa, J.A.V.; da Rosa, A.P.C.; Santos, L.O. Growth Stimulation and Synthesis of Lipids, Pigments and Antioxidants with Magnetic Fields in Chlorella Kessleri Cultivations. Bioresour. Technol. 2017, 244, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Bulut, O.; Akın, D.; Sönmez, Ç.; Öktem, A.; Yücel, M.; Öktem, H.A. Phenolic Compounds, Carotenoids, and Antioxidant Capacities of a Thermo-Tolerant Scenedesmus sp. (Chlorophyta) Extracted with Different Solvents. J. Appl. Phycol. 2019, 31, 1675–1683. [Google Scholar] [CrossRef]
- Choochote, W.; Suklampoo, L.; Ochaikul, D. Evaluation of Antioxidant Capacities of Green Microalgae. J. Appl. Phycol. 2014, 26, 43–48. [Google Scholar] [CrossRef]
- Custódio, L.; Soares, F.; Pereira, H.; Barreira, L.; Vizetto-Duarte, C.; Rodrigues, M.J.; Rauter, A.P.; Alberício, F.; Varela, J. Fatty Acid Composition and Biological Activities of Isochrysis galbana T-ISO, Tetraselmis sp. and Scenedesmus sp.: Possible Application in the Pharmaceutical and Functional Food Industries. J. Appl. Phycol. 2014, 26, 151–161. [Google Scholar] [CrossRef]
- Elshobary, M.E.; El-Shenody, R.A.; Ashour, M.; Zabed, H.M.; Qi, X. Antimicrobial and Antioxidant Characterization of Bioactive Components from Chlorococcum minutum. Food Biosci. 2020, 35, 100567. [Google Scholar] [CrossRef]
- Foo, S.C.; Yusoff, F.M.; Ismail, M.; Basri, M.; Khong, N.M.H.; Chan, K.W.; Yau, S.K. Efficient Solvent Extraction of Antioxidant-Rich Extract from a Tropical Diatom, Chaetoceros calcitrans (Paulsen) Takano 1968. Asian Pac. J. Trop. Biomed. 2015, 5, 834–840. [Google Scholar] [CrossRef]
- Foo, S.C.; Yusoff, F.M.; Ismail, M.; Basri, M.; Yau, S.K.; Khong, N.M.H.; Chan, K.W.; Ebrahimi, M. Antioxidant Capacities of Fucoxanthin-Producing Algae as Influenced by Their Carotenoid and Phenolic Contents. J. Biotechnol. 2017, 241, 175–183. [Google Scholar] [CrossRef]
- Guedes, A.C.; Gião, M.S.; Seabra, R.; Ferreira, A.C.S.; Tamagnini, P.; Moradas-Ferreira, P.; Malcata, F.X. Evaluation of the Antioxidant Activity of Cell Extracts from Microalgae. Mar. Drugs 2013, 11, 1256–1270. [Google Scholar] [CrossRef] [Green Version]
- Gürlek, C.; Yarkent, Ç.; Köse, A.; Tuğcu, B.; Gebeloğlu, I.K.; Öncel, S.Ş.; Elibol, M. Screening of Antioxidant and Cytotoxic Activities of Several Microalgal Extracts with Pharmaceutical Potential. Health Technol. 2020, 10, 111–117. [Google Scholar] [CrossRef]
- Jaime, L.; Mendiola, J.A.; Ibáñez, E.; Martin-Álvarez, P.J.; Cifuentes, A.; Reglero, G.; Señoráns, F.J. β-Carotene Isomer Composition of Sub- and Supercritical Carbon Dioxide Extracts. Antioxidant Activity Measurement. J. Agric. Food Chem. 2007, 55, 10585–10590. [Google Scholar] [CrossRef]
- Jerez-Martel, I.; García-Poza, S.; Rodríguez-Martel, G.; Rico, M.; Afonso-Olivares, C.; Gómez-Pinchetti, J.L. Phenolic Profile and Antioxidant Activity of Crude Extracts from Microalgae and Cyanobacteria Strains. J. Food Qual. 2017, 2017, 2924508. [Google Scholar] [CrossRef] [Green Version]
- Maadane, A.; Merghoub, N.; Ainane, T.; El Arroussi, H.; Benhima, R.; Amzazi, S.; Bakri, Y.; Wahby, I. Antioxidant Activity of Some Moroccan Marine Microalgae: Pufa Profiles, Carotenoids and Phenolic Content. J. Biotechnol. 2015, 215, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Matos, J.; Cardoso, C.; Gomes, A.; Campos, A.M.; Falé, P.; Afonso, C.; Bandarra, N.M. Bioprospection of Isochrysis galbana and Its Potential as a Nutraceutical. Food Funct. 2019, 10, 7333–7342. [Google Scholar] [CrossRef] [PubMed]
- Millao, S.; Uquiche, E. Antioxidant Activity of Supercritical Extracts from Nannochloropsis Gaditana: Correlation with Its Content of Carotenoids and Tocopherols. J. Supercrit. Fluids 2016, 111, 143–150. [Google Scholar] [CrossRef]
- Morowvat, M.H.; Ghasemi, Y. Evaluation of Antioxidant Properties of Some Naturally Isolated Microalgae: Identification and Characterization of the Most Efficient Strain. Biocatal. Agric. Biotechnol. 2016, 8, 263–269. [Google Scholar] [CrossRef]
- Pereira, H.; Custódio, L.; Rodrigues, M.J.; De, S.; Oliveira, M.; Barreira, L.; Neng, N.D.R.; Nogueira, J.M.F.; Alrokayan, S.A.; Mouffouk, F.; et al. Biological Activities and Chemical Composition of Methanolic Extracts of Selected Autochthonous Microalgae Strains from the Red Sea. Mar. Drugs 2015, 13, 3531–3549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Meizoso, I.; Jaime, L.; Santoyo, S.; Señoráns, F.J.; Cifuentes, A.; Ibáñez, E. Subcritical Water Extraction and Characterization of Bioactive Compounds from Haematococcus pluvialis Microalga. J. Pharm. Biomed. Anal. 2010, 51, 456–463. [Google Scholar] [CrossRef] [Green Version]
- Sansone, C.; Galasso, C.; Orefice, I.; Nuzzo, G.; Luongo, E.; Cutignano, A.; Romano, G.; Brunet, C.; Fontana, A.; Esposito, F.; et al. The Green Microalga Tetraselmis suecica Reduces Oxidative Stress and Induces Repairing Mechanisms in Human Cells. Sci. Rep. 2017, 7, 41215. [Google Scholar] [CrossRef]
- Sharma, A.K.; General, T. Variation of Both Chemical Composition and Antioxidant Properties of Newly Isolated Parachlorella kessleri GB1, by Growing in Different Culture Conditions. LWT 2019, 112, 108205. [Google Scholar] [CrossRef]
- Singh, P.; Baranwal, M.; Reddy, S.M. Antioxidant and Cytotoxic Activity of Carotenes Produced by Dunaliella Salina under Stress. Pharm. Biol. 2016, 54, 2269–2275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, S.-S.; Yang, E.J.; Lee, S.G.; Youn, U.J.; Han, S.J.; Kim, I.-C.; Kim, S. Bioactivities of Ethanol Extract from the Antarctic Freshwater Microalga, Chloromonas sp. Int. J. Med. Sci. 2017, 14, 560–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, S.-S.; Kim, S.-M.; Kim, J.E.; Hong, J.-M.; Lee, S.G.; Youn, U.J.; Han, S.J.; Kim, I.-C.; Kim, S. Anticancer Activities of Ethanol Extract from the Antarctic Freshwater Microalga, Botryidiopsidaceae sp. BMC Complemen. Altern. Med. 2017, 17, 509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, K.M.; Habte-Tsion, H.-M.; Thompson, K.R.; Filer, K.; Tidwell, J.H.; Kumar, V. Freshwater Microalgae (Schizochytrium sp.) as a Substitute to Fish Oil for Shrimp Feed. Sci. Rep. 2019, 9, 6178. [Google Scholar] [CrossRef] [Green Version]
- El-Bahr, S.; Shousha, S.; Shehab, A.; Khattab, W.; Ahmed-Farid, O.; Sabike, I.; El-Garhy, O.; Albokhadaim, I.; Albosadah, K. Effect of Dietary Microalgae on Growth Performance, Profiles of Amino and Fatty Acids, Antioxidant Status, and Meat Quality of Broiler Chickens. Animals 2020, 10, 761. [Google Scholar] [CrossRef]
- Long, S.F.; Kang, S.; Wang, Q.Q.; Xu, Y.T.; Pan, L.; Hu, J.X.; Li, M.; Piao, X.S. Dietary Supplementation with DHA-Rich Microalgae Improves Performance, Serum Composition, Carcass Trait, Antioxidant Status, and Fatty Acid Profile of Broilers. Poult. Sci. 2018, 97, 1881–1890. [Google Scholar] [CrossRef]
- Marques, A.E.M.L.; Balen, R.E.; da Silva Pereira Fernandes, L.; Motta, C.M.; de Assis, H.C.S.; Taher, D.M.; Meurer, F.; Vargas, J.V.C.; Mariano, A.B.; Cestari, M.M. Diets Containing Residual Microalgae Biomass Protect Fishes against Oxidative Stress and DNA Damage. J. Appl. Phycol. 2019, 31, 2933–2940. [Google Scholar] [CrossRef]
- Nacer, W.; Baba Ahmed, F.Z.; Merzouk, H.; Benyagoub, O.; Bouanane, S. Evaluation of the Anti-Inflammatory and Antioxidant Effects of the Microalgae Nannochloropsis gaditana in Streptozotocin-Induced Diabetic Rats. J. Diabetes Metab. Disord. 2020, 19, 1483–1490. [Google Scholar] [CrossRef]
- Qiao, H.; Hu, D.; Ma, J.; Wang, X.; Wu, H.; Wang, J. Feeding Effects of the Microalga Nannochloropsis sp. on Juvenile Turbot (Scophthalmus maximus L.). Algal Res. 2019, 41, 101540. [Google Scholar] [CrossRef]
- Rahman, N.A.; Khatoon, H.; Yusuf, N.; Banerjee, S.; Haris, N.A.; Lananan, F.; Tomoyo, K. Tetraselmis Chuii Biomass as a Potential Feed Additive to Improve Survival and Oxidative Stress Status of Pacific White-Leg Shrimp Litopenaeus Vannamei Postlarvae. Int. Aquat. Res. 2017, 9, 235–247. [Google Scholar] [CrossRef] [Green Version]
- Ranga Rao, A.; Raghunath Reddy, R.L.; Baskaran, V.; Sarada, R.; Ravishankar, G.A. Characterization of Microalgal Carotenoids by Mass Spectrometry and Their Bioavailability and Antioxidant Properties Elucidated in Rat Model. J. Agric. Food Chem. 2010, 58, 8553–8559. [Google Scholar] [CrossRef] [PubMed]
- Ranga Rao, A.; Baskaran, V.; Sarada, R.; Ravishankar, G.A. In Vivo Bioavailability and Antioxidant Activity of Carotenoids from Microalgal Biomass—A Repeated Dose Study. Food Res. Int. 2013, 54, 711–717. [Google Scholar] [CrossRef]
- Sheikhzadeh, N.; Tayefi-Nasrabadi, H.; Khani Oushani, A.; Najafi Enferadi, M.H. Effects of Haematococcus pluvialis Supplementation on Antioxidant System and Metabolism in Rainbow Trout (Oncorhynchus mykiss). Fish. Physiol. Biochem. 2012, 38, 413–419. [Google Scholar] [CrossRef]
- Carocho, M.; Morales, P.; Ferreira, I.C.F.R. Antioxidants: Reviewing the Chemistry, Food Applications, Legislation and Role as Preservatives. Trends Food Sci. Technol. 2018, 71, 107–120. [Google Scholar] [CrossRef] [Green Version]
- Lourenço, S.C.; Moldão-Martins, M.; Alves, V.D. Antioxidants of Natural Plant Origins: From Sources to Food Industry Applications. Molecules 2019, 24, 4132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Commission Règlement (UE). 2015/2283 du Parlement Européen et du Conseil du 25 Novembre 2015 Relatif aux Nouveaux Aliments, Modifiant le Règlement (UE) No. 1169/2011 du Parlement Européen et du Conseil et Abrogeant le Règlement (CE) n° 258/97 du Parlement Européen et du Conseil et le Règlement (CE) No. 1852/2001 de la Commission (Texte Présentant de L’intérêt pour l’EEE). Available online: http://data.europa.eu/eli/reg/2015/2283/oj/fra (accessed on 15 May 2020).
- Turck, D.; Bresson, J.-L.; Burlingame, B.; Dean, T.; Fairweather-Tait, S.; Heinonen, M.; Hirsch-Ernst, K.I.; Mangelsdorf, I.; McArdle, H.; Naska, A.; et al. Guidance on the Preparation and Presentation of an Application for Authorisation of a Novel Food in the Context of Regulation (EU) 2015/2283. EFSA J. 2016, 14, e04594. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, K.A.; Robertson, A. Domoic Acid and Human Exposure Risks: A Review. Toxicon 2010, 56, 218–230. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, C.H.; Bates, S.S.; Martin, J.L.; Haigh, N.; Howland, K.L.; Lewis, N.I.; Locke, A.; Peña, A.; Poulin, M.; Rochon, A.; et al. Three Decades of Canadian Marine Harmful Algal Events: Phytoplankton and Phycotoxins of Concern to Human and Ecosystem Health. Harmful Algae 2021, 102, 101852. [Google Scholar] [CrossRef]
- Kilcoyne, J.; Nulty, C.; Jauffrais, T.; McCarron, P.; Herve, F.; Foley, B.; Rise, F.; Crain, S.; Wilkins, A.L.; Twiner, M.J.; et al. Isolation, Structure Elucidation, Relative LC-MS Response, and in Vitro Toxicity of Azaspiracids from the Dinoflagellate Azadinium Spinosum. J. Nat. Prod. 2014, 77, 2465–2474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CEVA. Macroalgues et Microalgues Alimentaires—Statut Règlementaire en France et en Europe; CEVA: Marseille, France, 2019; p. 15. [Google Scholar]
- European Commission Règlement (UE). No. 68/2013 de la Commission du 16 Janvier 2013 Relatif au Catalogue des Matières Premières pour Aliments des Animaux Texte Présentant de L’intérêt pour l’EEE. Available online: https://eur-lex.europa.eu/legal-content/FR/ALL/?uri=CELEX%3A32013R0068 (accessed on 15 May 2020).




{kind=link}
{kind=link}
{kind=link}
| Name of the method | Principle | Mode of Detection | Ref. | |
|---|---|---|---|---|
| In vitro | ORAC (oxygen radical absorbance capacity) assay | measure the chain breaking capacity against peroxyl radical generated by the thermal decomposition of AAPH (2,2′-azobis (2-amidino-propane) dihydrochloride). The peroxyl radical reacts with fluorescein (fluorescent probe), causing a fluorescence loss over time | fluorimetry | [81] |
| β-carotene bleaching assay | measure the inhibition capacity of β-carotene oxidation induced by radical products resulting from the peroxidation of linoleic acid. The discoloration of β-carotene is measured at 434 nm | photocolorimetry | [82] | |
| TEAC (trolox equivalent antioxidant capacity) assay | measure the scavenging capacity of the blue chromophore ABTS (2,2′-azino-bis (3-éthylbenzothiazoline-6-sulphonique)) radical cation, which is reduced to a colorless compound in the presence of a radical scavenger. The discoloration is followed by absorbance measure at 734 nm | photocolorimetry | [83] | |
| DPPH (2,2-diphenyl-1-picrylhydrazyl) radical scavenging capacity assay | measure the scavenging capacity of the purple DPPH radical which is reduced to a pale-yellow compound in the presence of a radical scavenger. The absorbance decrease is measured at 515 nm | photocolorimetry | [84] | |
| Reducing power assay | measure the reduction capacity of potassium ferricyanide to potassium ferrocyanide which produces a ferric ferrocyanide blue complex by reaction with ferric chloride. The absorbance of the complex is measured at 700 nm | photocolorimetry | [85] | |
| FRAP (ferric-reducing antioxidant power) assay | measure the reduction capacity of ferric-TPTZ (tripyridyltriazine) to ferrous-TPTZ, the latter forming a blue complex at acidic pH which is measured at an absorbance of 593 nm | photocolorimetry | [46] | |
| TAC (total antioxidant capacity) assay or phosphomolybdenum assay | measure the reduction capacity of molybdenum Mo(vi) to Mo(v), the latter forming a green phosphate-Mo(v) complex at low pH which is followed by absorbance measure at 695 nm | photocolorimetry | [8] | |
| FCA (ferrous-chelating activity) assay | measure the ferrous-chelating activity by following the formation of a magenta-colored Fe2+-ferrozine complex at an absorbance of 562 nm. Coexisting chelator acts as competing agents results in decrease in the absorbance | photocolorimetry | [86] | |
| CCA (copper-chelating activity) assay | measure the copper-chelating activity by following the dissociation of the blue complex of pyrocatechol violet (PV) with CuSO4. The color turned to yellow when PV dissociated a Cu ion in the presence of chelating agents. The change in color is measured at 632 nm. | photocolorimetry | [45] | |
| TBARS (thiobarbituric acid reactive substances) assay | measure of the end-product of lipid peroxidation which formed a pink complex with thiobarbituric acid at 100 °C in acidic condition. The formation of the complex is measured at an absorbance of 534 nm | photocolorimetry | [84] | |
| Superoxide radical scavenging activity assay | measure the scavenging capacity of superoxide radical generated by the reaction of NADH with phenazine methosulfate or by the oxidation of hypoxanthine by the xanthine oxidase. The inhibition of the reduction of nitroblue tetrazolium in blue-colored formazan by superoxide radical is followed at an absorbance of 560 nm. | photocolorimetry | [49,87] | |
| Hydrogen peroxide scavenging activity by FOX (ferrous ion oxidation–xylenol orange) assay | measure the scavenging capacity of hydrogen peroxide. Hydrogen peroxide oxidizes ferrous ion to ferric ion, which then forms a blue-purple complex with xylenol orange. The decrease in absorbance in presence of scavenger is read at 560 nm | photocolorimetry | [49] | |
| Hydroxyl radical scavenging activity assay | measure the scavenging capacity of hydroxyl radical which is generated by the Fenton reaction. 2-deoxyribose is oxidized by hydroxyl radical and degraded to malondialdehyde. It forms a pink complex with thiobarbituric acid at 100 °C in acidic condition which is measured at an absorbance of 532 nm. | photocolorimetry | [87] | |
| In vitro or on cell | Nitric oxide scavenging activity assay | measure the scavenging capacity of nitric oxide (NO), generated from sodium nitroprusside. NO reacts with oxygen to produce nitrite which can be estimated by use of Griess reagent (mix of sulphanilamide, phosphoric acid and naphthylethylenediamine dihydrochloride). Scavengers of NO compete with oxygen leading to reduced production of nitrite. The absorbance of the chromophore formed by the reaction of Griess reagent and nitrite was read at 546 nm. Nitrite oxide scavenging capacity could also be evaluated with a cellular-based assay. NO release by cells is determined by measurement of nitrite concentration in culture supernatant using the Griess reagent. | photocolorimetry | [88,89] |
| On cell | ROS (reactive oxygen species) assay | measure the decrease in ROS produced by cells after stress induction in presence of antioxidant. The cells are incubated with the fluorescent dye CM-DCFDA (5-(e-6)-clorometil-2,7-dichloro dihydrofluorescein diacetate), and the fluorescence of the sample is measured at 535 nm (excitation 490 nm) to follow ROS production. | fluorimetry | [90] |
| CLPAA (cellular lipid peroxidation antioxidant activity) assay | measure inhibition of lipid peroxidation in cellular membranes by monitoring red (590/632 nm) and green (485/520 nm) fluorescent products generated by the lipophilic probe C-11-BODIPY after addition of cumene hydroperoxide. | fluorimetry | [91] | |
| CAA (cellular antioxidant activity) assay | measure the inhibition of oxidation of a fluorescent probe. The nonfluorescent DCFH (2′,7′-dichlorofluorescein) is entrapped in cell and oxidized by peroxyl radical derived from ABAP (2,2′-azobis(2-amidopropane)) or AAPH decomposition producing fluorescent DCF (dichlorofluorescein). Antioxidant prevent oxidation of the probe and attenuate cellular fluorescence (excitation and emission at 485 and 520 nm) | fluorimetry | [91] | |
| Comet assay (single-cell gel electrophoresis) | measure the nuclear DNA protection by an antioxidant after applying hydrogen peroxide oxidative stress on cells. Treated cells are embedded in agarose and are lysed to form nucleoids containing supercoiled loops of DNA linked to the nuclear matrix. After electrophoresis, the DNA is stained with a fluorescent dye and results in structures resembling comets observed by fluorescence microscopy; the intensity of the comet tail relative to the head reflects the number of DNA breaks. | fluorescence microscopy | [92] |
| Microalgae Species | Antioxidant Assay | Composition Analyses | Antioxidant Activity | Positive Control | Molecules Involved in Antioxidant Activity | Method of Extraction | Ref. |
|---|---|---|---|---|---|---|---|
| Grammatophora marina | (i) DPPH (ii) FCA (iii) hydrogen peroxide scavenging activity (iv) superoxide radical scavenging activity (v) hydroxyl radical scavenging activity (vi) nitric oxide scavenging activity | - | extracts at 2000 µg mL−1 (i) 41–86% inhib. (ii) 21–81% inhib. (iii) 14–25% inhib. (iv) 24–45% inhib. (v) 10–35% inhib. (vi) 12–33% inhib. | α-toco. and BHT at 2000 µg mL−1 (i) 70 and 72% inhib (ii) 10 and 11% inhib. (iii) 74 and 67% inhib. (iv) 33 and 64% inhib (v) 79 and 77% inhib. (vi) 43 and 56% inhib. | - | maceration 80% MeOH or enzymatic lysis (5 carbohydrases and 5 proteases tested) | [93] |
| Chlorella vulgaris | (i) DPPH (ii) TEAC (iii) ORAC (iv) FRAP | TPC | (i) 0.8 µmol TE g−1 DW (ii) 15 µmol TE g−1 DW (iii) 31 µmol TE g−1 DW (iv) 0.6 µmol TE g−1 DW | - | phenolic compounds | US (30 min, room temp.) EtOH 50% | [94] |
| Dunaliella salina, Dunaliella tertiolecta, Phaeodactylum tricornutum, Chaetoceros muelleri, Pavlova salina, Pavlova lutheri, Tetraselmis suecica, Tetraselmis sp., Tetraselmis chui, Nannochloropsis sp., Isochrysis galbana | ORAC | TPC, total carotenoids | 45–577 µmol TE g−1 DW | - | - | maceration + EtOAC, Hex or H2O | [95] |
| Scenedesmus obliquus | (i) DPPH (ii) TEAC (iii) superoxide radical scavenging activity (iv) nitric oxide scavenging activity | carotenoids, PUFA | (i) IC50: 412–878 µg mL−1 (ii) IC50: 41–648 µg mL−1 (iii) IC50: 520–1236 µg mL−1 (iv) IC50 = 60 µg mL−1 | - | - | maceration (20 min 40 °C) + EtOH, Ac, ethyl lactate or Hex/i-PrOH (3/2) | [96] |
| Scenedesmus sp. + 4 Scenedesmus quadricauda strains | (i) DPPH (ii) β-carotene bleaching | TPC, tannins, iridoids | (i) 6–70% inhib. (extracts at 200 µg mL−1) (ii) 24–92% inhib. (extracts at 400 µg mL−1) | (i) AA: 98% inhib. at200 µg mL−1 (ii) BHT: 70% inhib. at 400 µg mL−1 | phenolic compounds | maceration + US (30 min, in ice) + MeOH 50%, PE or DCM | [82] |
| Chlorella minutissima | (i) DPPH (ii) β-carotene bleaching | TPC, tannins, iridoids, pigments | (i) 10–70% inhib. (extracts at 200 µg mL−1) (ii) IC50: 75–600 µg mL−1 | (i) AA: 97% inhib. at 200 µg mL−1 (ii) IC50 BHT = 60.7 µg mL−1 | carotenoids, phenolic compounds | maceration (1 night) + US (30 min, in ice) + MeOH, PE or DCM | [44] |
| Chlorella minutissima + 2 Chlorella sp. strains. | (i) DPPH (ii) β-carotene bleaching | TPC, tannins, flavonoids, iridoids | (i) 25–100% inhib. (extracts at 200 µg mL−1) (ii) IC50: 25–450 µg mL−1 | (i) AA: 97% inhib. at 200 µg mL−1 (ii) IC50 BHT = 61 µg mL−1 | - | maceration (1 night) + US (30 min, in ice) MeOH, PE or DCM | [48] |
| Ammatoidea normanii, Ruttnera lamellose, Pavlova granifera, Apistonema sp., 2 Cryptomonas pyrenoidifera strains, Porphyridium aerugineum, Porphyridium sordidum, Audorinella sp., Phragmonema sordidum, 3 Characiopsis aquilonaris strains, Characiopsis ovalis, 2 Characiopsis sp. strains, Characiopsis minima, Pseudostaurastrum enorme, Goniochloris sculpta, Eustigmatos sp., Vischeria helvetica, Chlorobotrys gloeothece, Chlorobotrys sp., Dioxys sp., Coronastrum aestivale, Chlorella vulgaris, Mychonastes homosphaera, Gloeococcus minor, Pectodyction cubicum, Jaagiella apicola, Schizomeris leibleinii, Interfilum paradoxum, Micrasterias radiosa var. elegantior, Haematococcus pluvialis, Lobomonas sp., Stephanosphaera pluvialis, Bumilleria sicula, Euglena cantabrica | (i) DPPH (ii) TEAC | - | (i) IC50: 44–1421 mg FW mL−1 (ii) 5–195 mg AAE 100 g−1 FW and 17–258 mg TE 100 g−1 FW | - | - | US (30 min, dark) + maceration (1 night, −4 °C) + EtOH | [6] |
| Botryococcus braunii, Chlorella sorokiniana, Nannochloropsis granulata, Neochloropsis oleabundans, Phaeodactylum tricornutum, Porphyridium aerugineum, Scenedesmus obliquus, Scenedesmus sp., Tetraselmis chuii | (i) DPPH (ii) ORAC | TPC, carotenoids, lipids, FA | (i) <50% inhib. (extracts at 200 µg mL−1) (ii) 7–53 µmol TE g−1 DW | - | phenolic compounds and lipids | maceration MeOH (DPPH) or PLE Hex/DCM (50/50)(70 °C) and then Ac/H2O/AcOH (70/29.5/0.5) (80 °C) (ORAC) | [97] |
| Chlorella kessleri | (i) DPPH (ii) TEAC (iii) reducing power | total carotenoids, chlorophylls a and b | (i) 1–4% inhib. (extracts at 2500 µg mL−1) (ii) 196–346 µmol TE g−1 extract (iii) ABS700: 0,266–0,473 (extracts at 2500 µg mL−1) | - | - | maceration MeOH | [98] |
| Scenedesmus sp. | (i) DPPH (ii) FRAP | TPC, flavonoids, carotenoids | (i) 0.6–3.7 µmol TE g−1 DW (ii) 2.8–47.0 µmol TE g−1 DW | - | - | US (20 min) + maceration (1 h) EtOH/H2O (3:1), Hex, EtOAc, or H2O | [99] |
| Botryococcus braunii | ORAC | - | 43 µmol TE g−1 extract | - | - | grinding + PBS | [90] |
| Euglena tuba | (i) DPPH (ii) TBARS (iii) superoxide radical scavenging activity (iv) hydrogen peroxide scavenging activity (v) peroxynitrite scavenging activity (vi) singlet oxygen scavenging activity (vii) hypochlorous acid scavenging activity | TPC, flavonoids, tannins, alkaloids, AA | (i) IC50 = 146 µg mL−1 (ii) IC50 = 42 µg mL−1 (iii) IC50 = 5.8 µg mL−1 (iv) IC50 = 47340 µg mL−1 (v) IC50 = 278 µg mL−1 (vi) IC50 = 2821 µg mL−1 (vii) IC50 = 879 µg mL−1 (viii) IC50 = 223 µg mL−1 | (i) IC50 AA = 5.3 µg mL−1 (ii) IC50 mannitol = 571.4 µg mL−1 (ii) IC50 quercetin = 42.1 µg mL−1 (iv) IC50 sodium pyruvate = 3.2 mg mL−1 (v) IC50 curcumin = 90.8 µg mL−1 (vi) IC50 gallic acid = 0.88 mg mL−1 (vii) IC50 lipoic acid = 0.05 mg mL−1 (viii) IC50 AA = 236.0 µg mL−1 | - | maceration (15 h) + MeOH 70% | [49] |
| 3 Chlorella sp. strain | (i) DPPH (ii) FCA (iii)TBARS | TPC | (i) IC50: 810–1400 µg mL−1 (ii) IC50: 1220–1500 µg mL−1 (iii) 5.9–88% inhib. (extracts at 4000 µg mL−1) | (i) IC50 BHT = 50 µg mL−1 (ii) IC50 EDTA = 28 µg mL−1 (iii) BHT 94% inhib. (conc. not specified) | - | grinding (20 min) + H2O 80 °C 20 min or maceration (24h) + EtOH 95% | [100] |
| Nephroselmis sp., Tetraselmis sp., Dunaliella sp., Picochlorum sp., Schizochlamydella sp., 2 Nitzschia sp. strain, Thalassiosira weissflogi, Entomoneis punctulata, Cylindrotheca closterium, Chaetoceros sp., Bacillaria sp. | (i) DPPH (ii) TEAC (iii) ORAC (iv) TBARS | carotenoids composition | (i) IC50 from 484 to >1000 µg mL−1 (ii) IC50 from 193 to >1000 µg mL−1 (iii) 0–190 µg TE mg−1 extract (iv) IC50: 15.4–473.6 µg mL−1 extract | (i) IC50 trolox = 4.7 µg mL−1, α-toco. = 6.2 µg mL−1, AA = 8.7 µg mL−1, β-carotene = 257.3 µg mL−1, astaxanthin = 228.6 µg mL−1 (ii) IC50 trolox = 6.4 µg mL−1, α-toco. = 10.8 µg mL−1, AA = 6.1 µg mL−1, β-carotene = 37.0 µg mL−1, astaxanthin = 98.5 µg mL−1 (iv) IC50 trolox = 0.2 µg mL−1, α-toco. = 1.3 µg mL−1 | carotenoids | US (60 min) + MeOH/DCM (50/50) | [7] |
| Nephroselmis sp. | ORAC | carotenoids composition | 63.6–154.9 µmol TE g−1 DW | - | carotenoids | grinding + maceration (30 min, room temp., dark) + EtOH | [22] |
| Tetraselmis sp. | TBARS | - | IC50: 3.4–11.3 µg mL−1 extract | IC50 trolox = 0.2 µg mL−1, IC50 α-toco. = 1.3 µg mL−1 | - | grinding + US (10 min., ice bath, dark) + MeOH/DCM (50/50) | [21] |
| Tetraselmis chuii, Nannochloropsis oculata, Chlorella minutissima, Rhodomonas salina | (i) DPPH (ii) FCA (iii) CCA | TPC | extracts at 1000 µg mL−1 (i) 0–21% inhib. (ii) 12–98% inhib. (iii) 12–22% inhib. | conc. at 1000 µg mL−1 (i) BHT: 88% inhib. (ii) EDTA: 95% inhib. (iii) EDTA: 74% inhib. | - | grinding + maceration (1 nuit) + Hex or MeOH | [45] |
| Isochrysis galbana T-iso, Tetraselmis sp., Scenedesmus sp. | (i) DPPH (ii) FCA (iii) CCA | TPC, FA | (i) IC50 > 1000 µg mL−1 (ii) IC50: 730–4110 µg mL−1 (iii) IC50: 900 µg mL−1 to >10000 µg mL−1 | (i) IC50 BHT = 70 µg mL−1 (ii) IC50 EDTA = 100 µg mL−1 (iii) IC50 EDTA = 280 µg mL−1 | - | grinding + Hex, and, Ac and H2O in sequential order | [101] |
| Chlorococcum minutum | (i) TAC (ii) reducing power | TPC | (i) 2.5–10 mg AAE g−1 extract (ii) 1–4 mg AAE g−1 extract | - | phenolic compounds | maceration (72 h) EtOH, MeOH, or Ac | [102] |
| Chaetoceros calcitrans | (i) DPPH (ii) TEAC (iii) FCA | TPC, major phenolic compounds, total carotenoids totaux, fucoxanthin | (i) 0.1–1.4 mg TE g−1 DW (ii) 1.2–10.6 mg TE g−1 DW (iii) 0.3–18.5 mg Na-EDTA Eq g−1 DW | - | carotenoids and phenolic compounds | grinding + US (30 min, room temp.) + MeOH, EtOH, Ac, Ac 90%, Ac/CHCl3 (90/10) or Ac/CHCl3/MeOH (80/10/10) | [103] |
| Chaetoceros calcitrans, Isochrysis galbana, Skeletonema costatum, Odontella sinensis, Phaedactylum tricornatum | (i) TEAC (ii) FRAP (iii) FCA (iv) β-carotene bleaching | TPC, major phenolic compounds, total carotenoids totaux, fucoxanthin | (i) 2.0–21.5 mg TE g−1 DW (ii) 0.2–2.0 mg TE g−1 DW (iii) 1.5–13.4 mg EDTA eq g−1 DW (iv) 0.1–1.4 mg TE g−1 DW | - | carotenoids and phenolic compounds | grinding + MeOH | [104] |
| Chaetoceros sp., Nannochloropsis sp. | (i) DPPH (ii) FRAP (iii) FCA (iv) superoxide radical scavenging activity | TPC | (i) 14.0–106.7 µmol TE g−1 extract (ii) 171.5–609.8 µmol TE g−1 extract (iii) 3.2–82.4 µmol EDTA Eq g−1 extract (iv) 227.9–3224.5 µmol TE g−1 extract | - | - | maceration (24 h) + Hex, DCM, CHCl3 or MeOH | [86] |
| Nannochloropsis oculata, Nannochloropsis sp., Isochrysis sp., Isochrysis ISO-T, Tetraselmis sp., Tetraselmis suecica, Botryococcus braunii, Porphyridium cruentum, Neochloris oleabundans, Chaetoceros calcitrans, Chlorella vulgaris, Haematococcus pluvialis (red and green phase), Parachlorella kessleri, Phaeodactylum tricornutum, Schizochytrium sp. | (i) TEAC (ii) FRAP (iii) AIOLA | TPC, total carotenoids | (i) 0–69 µmol TE g−1 DW (ii) 3.3–90 µmol TE g−1 DW (iii) 1.8–89.7 µmol TE g−1 DW | - | carotenoids and phenolic compounds | grinding + maceration (30 min) + EtOH/H2O (3/1) or Hex, EtOAc and H2O (80 °C) in sequential order | [10] |
| Phaeodactylum tricurnutum, 2 Chlorella vulgaris strains, Haematococcus pluvialis, Scenedesmus maximus, Scenedesmus obliquus, Scenedesmus quadricauda, Desmodesmus pleimorphus, Nannochloropsis sp., Pavlova lutheri, Porphyridium aerugineum | TEAC | carotenoids | 0.8–149 mg L−1 AAE µg−1 chlorophyll a | - | - | grinding + EtOH 50% | [105] |
| Galdieria sulphuraria, Ettlia carotinosa, Neochloris texensis, Chlorella minutissima, Stichococcus bacillaris, Schizochytrium limacinum, Crypthecodinium cohnii, Chlorella vulgaris | DPPH | TPC | 89–95% inhib. (extracts at 250 µg mL−1) | BHT: 98% inhib. at 250 µg mL−1 | TPC | US (20 min) MeOH or maceration H2O (100 °C, 30 min) | [106] |
| Chlorella stigmatophora, Phaeodactylum tricornutum | (i) Superoxide radical scavenging activity (ii) hydroxyl radical scavenging activity (iii) hypochlorous acid scavenging activity | - | (i) IC50: 48–170 µg mL−1 (ii) IC50: 180–250 µg mL−1 (iii) IC50 > 1000 µg mL−1 | - | - | US + H2O then soxhlet + DCM and MeOH on extraction residue | [87] |
| Chlorella vulgaris | FRAP | TPC | 0.01–58.2 µmol TE g−1 DW | - | phenolic compounds | maceration + Hex, EtOAc + H2O (80 °C) in sequential order | [46] |
| Phaeodactylum tricornutum, Nannochloropsis gaditana, Nannochloris sp., Tetraselmis suecica | (i) DPPH (ii) reducing power (iii) FCA | TPC, flavonoids, carotenoids | (i) IC50: 356–400 µg mL−1 (ii) 24–33 AAE mL−1 (iii) IC50: 2810–12820 µg mL−1 | (i) IC50 AA = 3,7 µg mL−1 (ii) BHT = 1,4 AAE mg−1 (iii) IC50 EDTA = 10 µg mL−1 | - | Not specified | [51] |
| Dunaliella salina | TEAC | carotenoids | 11–1118 µmol TE g−1 extract | - | carotenoids | PLE Hex, EtOH or H2O | [83] |
| Dunaliella salina | TEAC | carotenoids | 115–452 µmol TE g−1 extract | - | carotenoids | sub- and super-critical CO2 | [107] |
| Chlorella vulgaris, Chlamydomonas reinhardtii | (i) DPPH (ii) TAC (iii) FRAP | TPC, flavonoids | (i) IC50: 397–423 µg mL−1 (ii) IC50: 55–73 µg mL−1 (iii) ABS700: 0.136 to 0.124 (extracts at 250 µg mL−1) | (ii) IC50 AA = 127.5 µg mL−1 (iii) ABS700 AA = 0.423 at 250 µg mL−1 | flavonoids | maceration MeOH | [47] |
| Ankistrodesmus sp., Euglena cantabrica | DPPH | - | 8–71% inhib. (extracts at 1000 µg mL−1) | conc. at 1000 µg mL−1 BHT: 26% inhib., BHA: 91% inhib. | - | maceration (40 min) + MeOH or H2O | [108] |
| Halochlorococcum porphyrae, Oltamannsiellopsis unicellularis | (i) DPPH (ii) FCA (iii) hydrogen peroxide scavenging activity (iv) superoxide radical scavenging activity (v) hydroxyl radical scavenging activity (vi) nitric oxide scavenging activity | TPC | extracts at 2000 µg mL−1 (i) 42–95% inhib. (ii) 4–72% inhib. (iii) 5–42% inhib. (iv) 5–58% inhib. (v) 4–31% inhib (vi) 1–51% inhib. | conc. at 2000 µg mL−1 (i) BHT and α-toco: 94% inhib. (ii) BHT: 11% inhib., α-toco 10% inhib. (iii) BHT 60% inhib., α-toco 62% inhib. (iv) BHT 63% inhib., α-toco 61% inhib. (v) BHT 77% inhib., α-toco 79% inhib. (vi) BHT 26% inhib., α-toco 25% inhib. | - | 80% MeOH then fractionation with Hex, CHCl3 and EtOAc or enzymatic lysis (5 carbohydrases and 5 proteases tested) | [89] |
| Chlamydomonas nivalis, Chlorella protothecoides, Chlorella pyrenoidosa, Chlorella vulgaris, Chlorella zofingiensis, Crypthecodinium cohnii, Nitzschia laevis, Schizochytrium sp., Schizochytrium mangrovei, Thraustochytrium sp. | TEAC | TPC | 0–11.4 µmol TE g−1 DW | - | - | maceration (30 min) + Hex, EtOAc and H2O (80 °C) in sequential order | [13] |
| Tetraselmis sp., Dunaliella salina, Dunaliella sp., Nannochloropsis gaditana, Chlorella sp., Navicula sp., Phaeodactylum tricurnutum, Chaetoceros sp., Isochrysis sp. | DPPH | TPC, total carotenoids, PUFA | IC50: 247–464 µg mL−1 | IC50 BHT = 6.2 µg mL−1, IC50 AA = 2.5 µg mL−1 | - | maceration (3h, dark) + EtOH | [109] |
| Isochrysis galbana | (i) DPPH (ii) TEAC | TPC, β-glucan, Co-Q10, β-carotene, fucoxanthin | (i) 0–17 mg AAE L−1 (ii) 52–56 µmol TE g−1 DW | - | - | grinding + maceration (18 h) EtOH 96% or H2O | [110] |
| Nannochloropsis gaditana | (i) DPPH (ii) β-carotene bleaching (iii) FRAP | carotenoids, tocopherols, FA | (i) 1,1–1,8 µmol TE g−1 extract (ii) 64–97% inhib. (extracts at 1000 µg mL−1) (iii) 48–86 µmol Fe(II) g−1 extract | - | carotenoids, tocopherols, FA | Supercritical CO2 | [111] |
| Dunaliella salina, Oocystis pusilla, Scenedesmus rubescens | DPPH | TPC | 0.4–17.5 µmol TE g−1 | - | phenolic compounds | maceration (30 min, 25 °C) + Hex, EtOAc and H2O (80 °C) in sequential order | [112] |
| Cymbella sp., Navicula sp., Skeletonema costatum, Isochrysis galbana, Chaetoceros calcitrans, Nannochloropsis oculata, Tetraselmis tetrathele, Scenedesmus quadricauda, Chlorella vulgaris, Oocystis sp., Trachelomonas sp. | (i) DPPH (ii) FTC (iii)TBARS | - | (i) no activity for extracts at 250–1000 µg L−1 (ii) 0–97% inhib.(extracts at 200 µg mL−1) (iii) 0–98% inhib. (extracts at 80 µg mL−1) | (i) α-toco: 85% inhib., quercetin: 65% inhib, BHT: 74% inhib. (100 µg L−1) (ii) α-toco: 84% inhib, quercetin: 92% inhib., BHT: 100% inhib. (200 µg mL−1) (iii) α-toco: 71% inhib., quercetin: 90% inhib., BHT: 98% inhib. (80 µg mL−1) | - | maceration (4 j) + MeOH | [84] |
| 2 Nannochloris sp. strains, Picochlorum sp., Desmochloris sp. | (i) DPPH (ii) FCA (iii) CCA | TPC, pigments | extracts at 1000 µg mL−1 (i) <10% inhib. (ii) <25% inhib. (iii) <30% inhib. | conc. at 1000 µg mL−1 (i) BHT: 88% inhib. (ii) EDTA: 96% inhib. (iii) EDTA: 76% inhib. | - | grinding + maceration (1 night, 20 °C) + MeOH | [113] |
| Chlorella vulgaris | (i) TEAC (ii) ORAC (iii) superoxide radical scavenging activity | TPC | (i) 146–789 µmol TE g−1 extract (ii) 243–1008 µmol TE g−1 extract (iii) IC50: 8260–10752 µg mL−1 | rosemary extract (i) 2805–2811 µmol TE g−1 (ii) 4615–4892 µmol TE g−1 (iii) IC50: 464–665 µg mL−1 | - | supercritical H2O | [81] |
| Haematococcus pluvialis | TEAC | GC-MS | 366–1974 µmol TE g−1 extract | - | α-toco., gallic acid, caramelization products and possible Maillard reaction products | supercritical H2O | [114] |
| Phaeodactylum tricornutum, Nannochloropsis salina, Nannochloropsis limnetica, Chlorella sorokiniana, Dunaliella salina, Desmodesmus sp. | (i) DPPH (ii) TEAC (iii) FCA (iv) FRAP (v) TAC | TPC, flavonoids, phenolic acids, tocopherols, carotenoids composition | (i) 8–14% inhib. (extracts at 250 µg mL−1) (ii) 2.7–24.2 TE g−1 (iii) 3–9% chelation (extracts at 250 µg mL−1) (iv) 0.1–0.5 AAE g−1 (v) 3.0–8.9 gallic acid Eq g−1 | - | phenolic compounds, carotenoids and tocopherols | US (45 min in the dark at room temp.) + MeOH | [8] |
| Tetraselmis suecica | DPPH | pigment composition | 21.1% inhib. (extract at 50 µg mL−1) | α-toco: 6% inhib. at 50 µg mL−1 | - | maceration (30 min in the dark under nitrogen atmosphere at room temp.) + EtOH/H2O (3/1) | [115] |
| Parachlorella kessleri | (i) DPPH (ii) TEAC (iii) FCA (iv) TAC | TPC, chlorophyll a and b, total carotenoids | (i) 32–69% inhib. (extracts at 100 µg mL−1) (ii) 1.4–3.0 µmol TE g−1 extract (iii) 20% inhib. (extracts at 500 µg mL−1) (iv) 2.2–4.3 mg AAE g−1 extract | - | - | grinding + maceration MeOH | [116] |
| Trentepohlia umbrina | (i) DPPH (ii) reducing power (iii) superoxide radical scavenging activity | TPC, flavonoids | (i) IC50 = 665.3 µg mL−1 (ii) ABS700 = 0.0124(extract at 125 µg mL−1) (iii) IC50 = 838.8 µg mL−1 | (i) IC50 AA = 6.4 µg mL−1 (ii) ABS700 AA = 0.0478 at 125 µg mL−1 (iii) IC50 AA = 115.6 µg mL−1 | - | maceration (72 h) + MeOH | [85] |
| Dunaliella salina | DPPH | chlorophylls, total carotenoid | 15–57% inhib. (extract at 250 µg mL−1) | AA: 95% inhib. at 250 µg mL−1 | - | US (10 min) + maceration (4 j) + EtOH | [117] |
| Skeletonema marinoi | TEAC | TPC, flavonoids, AA, β-carotene, diatoxanthin | 250–1500 fg AAE cell−1 | - | phenolic compounds, flavonoids, AA | US (1 min, in ice) + maceration (30 min, dark) MeOH | [53] |
| Chloromonas sp. | (i) DPPH (ii) TEAC | - | (i) IC50 = 1.0µg mL−1 (ii) IC50 = 0.9 µg mL−1 | (i) IC50 AA = 0.1 µg mL−1 (ii) IC50 AA = 0.2 µg mL−1 | - | maceration (24 h) + EtOH | [118] |
| Botryidiopsidaceae sp. | (i) DPPH (ii) TEAC | - | (i) IC50 = 1.5 µg mL−1 (ii) IC50 = 1.8 µg mL−1 | (i) IC50 AA = 0.2 µg mL−1 (ii) IC50 AA = 0.2 µg mL−1 | - | maceration (24 h) + EtOH | [119] |
| Crypthecodinium cohnii, Schizochytrium sp. | (i) DPPH (ii) TAC (iii) FCA (iv) reducing power | TPC, flavonoids | extracts at 500 µg mL−1 (i) 15–30% inhib. (ii) ABS695: 0,500–1,000 (iii) 10–60% inhib. (iv) ABS700: 0,050–0,300 | (ii) BHT: ABS695 = 0,500 at 500 µg mL−1 (iii) EDTA: 65% inhib. at 50 µg mL−1 (iv) BHT: ABS700 = 0,300 at 500 µg mL−1 | phenolic compounds | maceration (2 j) EtOH 70% | [54] |
| Microalgae Species | Antioxidant Assay | Composition Analyses | Antioxidant Activity | Positive Control | Molecules Involved in Antioxidant Activity | Extraction Method | Ref. |
|---|---|---|---|---|---|---|---|
| Chaetoceros calcitrans | Nitric oxide scavenging activity assay on RAW 264.7 cells (mouse macrophage) | metabolites profiling by 1H NMR + TPC | IC50: 3.5–187.7 µg mL−1 | IC50 quercetin = 4.7 µg mL−1 IC50 curcumin = 6.1 µg mL−1 | Fucoxanthin (25), astaxanthin, violaxanthin, zeaxanthin, canthaxanthin (26), and lutein (27) | US (30 min, room t °C) + MeoH, 70% EtOH, Ac, CHCl3 or Hex | [88] |
| Botryococcus braunii | (i) ROS assay and (ii) Comet assay on NIH3T3 cells (mouse embryonic fibroblast cells) | - | extract at 0.1–0.05% (i) reduction of ROS production of 35% over the control (=no microalgae extract) after stress induction (ii) no activity | (i) AA: reduction of ROS production by 64% over the control at 250 µM | - | crushing in PBS + silica sand | [90] |
| Pediastrum duplex, Halochlorococcum porphyrae, Oltmannsiellopsis unicellularis, Achnanthes longipes, Navicula sp., Amphora coffeaeformis | Comet assay on L5178 cells (mouse lymphoma cells) | Crude lipid content | extract at 25–100 µg mL−1 (i) inhibitory effect to DNA damage until 80% over the control (=no microalgae extract) after stress induction | - | - | Enzymatic extraction by 5 carbohydrases and 5 proteases | [92] |
| Cylindrotheca closterium, Coscinodiscus actinocyclus, Nitzschia closterium, 2 Pseudo-nitzschia pseudodelicatissimastrains, Tetraselmis suecica, Isochrysis galbana, Skeletonema costatum, Lauderia annulata, Leptocylindrus danicus, Chaetoceros affinis, Odontella mobiliensis, Leptocylindrus aporus, Thalassiosira rotula, Thalassiosira weissflogii, 2 Skeletonema marinoistrains, Thalassiosira rotula, Skeletonema costatum, Stephanopyxis turris, Bacteriastrum hyalinum, Guinardia striata, Proboscia alata, Guillardia theta, Rhodomonas baltica, Rhinomonas reticulata, Alexandrium tamutum, Alexandrium andersonii, Ostreopsis ovata, Alexandrium minutum, Lepidodinium viride, Prorocentrum gracile | (i) CAA and (ii) CLPAA on HepG2 cells (human liver cancer cell line) | extract at 50 µg mL−1 (i) 66–70% inhib. for Ostreopsis ovata (ii) 61–74% inhib. for Ostreopsis ovata and 100% inhib. for Alexandrium minutum but both species showed toxicity in cytotoxicity assay | - | - | US (1 min)+ H2O then addition of Ac + maceration (50 min, room temp.) then fractionation on Amberlite XAD16N resin | [91] |
| Microalgae Species | Experimental Animals | Concentration of Microalgae Tested | Experimental Time | Antioxidant Assay | Other Measure | Activity | Ref. |
|---|---|---|---|---|---|---|---|
| Schizochytrium sp. | Pacific white shrimps (Litopenaeus vannamei) | 0–75 g of dry microalgae kg−1 of feed | 12 weeks | TBARS on tail muscle | antioxidant enzymes activity (CAT, SOD), lipid composition of food and muscle | No effect of microalgae | [120] |
| Chlorella vulgaris and Amphora coffeaformis | Chickens (Cobb 500 broiler chick) | 1 g of dry microalgae kg−1 of feed | 32 days | TBARS on breast meat | SOD activity, FA and amino acids profiles of microalgae | 28–31% decrease in MDA compared to control group (feed without microalgae) | [121] |
| Schizochytrium limacinum | Chickens (Arbor Acres chick) | 1–2% of dry microalgae in feed | 42 days | In breast and thigh muscle (i) TAC (ii) TBARS | antioxidant enzymes activity of serum (SOD, GPX, CAT), FA composition of diet and muscle | Compared to control group (feed without microalgae): (i) 33–81% increase in TAC (ii) 11–35% decrease in MDA content | [122] |
| Acutodesmus obliquus | Catfish (Rhamdia quelen) | 1–3% of residual microalgae biomass (after oil extraction) in feed | 60 days | (i) TBARS in liver (ii) Comet assay in erythrocytes, liver, and brain | Antioxidant enzymes activity (SOD, CAT), pigment determination of microalgae residual biomass | (i) No effect of microalgae (ii) Decrease in DNA damage with 3% of microalgae in erythrocytes and liver, no effect in brain tissue | [123] |
| Nannochloropsis gaditana | Normal and diabetic Wistar rats | 10% of dry microalgae in feed | 8 weeks | (i) TBARS of liver mitochondria and liver tissue (ii) DNPH (protein oxidation) on liver mitochondria and liver tissue | On microalgae biomass: total carotenoids, carbohydrates, total lipids and total protein On liver mitochondria and tissue: antioxidant enzymes activity (SOD, CAT, GSH) | Compared to control group (feed without microalgae): (i) Normal rats: 0–8% decrease in MDA content Diabetic rats: 35% decrease in MDA content (ii) Normal rats: no effect. Diabetic rats: 18–25% decrease in protein oxidation | [124] |
| Nannochloropsis sp. | Juvenile turbots (Scophthalmus maximus L.) | 2.5–10% of dry microalgae in feed | 10 weeks | (i) TBARS in serum and liver (ii) TAC | Antioxidant enzyme activity (SOD, GPX) in serum and liver | Compared to control group (feed without microalgae): (i) 19–56% decrease in MDA content, (ii) 9–44% increase in TAC | [125] |
| Tetraselmis chuii | Pacific white shrimps postlarvae (Litopenaeus vannamei) | 25–100% of dry microalgae in feed | 12 days | In shrimp tissue (i) hydrogen peroxide content (ii) TBARS | Proximate analysis and antioxidant activity of the feed | (i) Decrease of about 0–25% of hydrogen peroxide content (ii) No effect of microalgae on lipid peroxidation | [126] |
| Haematococcus pluvialis, Botryococcus braunii | Wistar rats | Administration by intubation to the stomach of a single dose of one of the two microalgae biomass solubilized in olive oil as source of 200 µM equivalent of astaxanthine or lutein | 9 h | TBARS in plasma and liver | Analysis of carotenoids from plasma, liver and eyes Antioxidant enzyme activity (SOD, CAT, PX) in plasma and liver | 25–61% decrease in MDA content compared to MDA content at t0 | [127] |
| Haematococcus pluvialis, Botryococcus braunii | Wistar rats | Administration of a daily dose of one of the two microalgae biomass solubilized in olive oil as source of 200 µM equivalent of astaxanthine or lutein | 15 days | TBARS in plasma and liver | Analysis of carotenoids from plasma, liver and eyes Antioxidant enzyme activity (SOD, CAT, PX) in plasma and liver | 45–64% decrease in MDA content compared to MDA content at t0 | [128] |
| Haematococcus pluvialis | Juvenile rainbow trout (Oncorhynchus mykiss) | 1–10 g of dry microalgae kg−1 of feed | 30 days | In serum (i) FRAP (ii) TBARS | alkaline phosphatase, alanine aminotransferase, aspartate and serum total protein, glucose, triglycerides, and cholesterol | Compared to control group (feed without microalgae): (i) 36–75% increase in activity (ii) 44–69% decrease in MDA content | [129] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coulombier, N.; Jauffrais, T.; Lebouvier, N. Antioxidant Compounds from Microalgae: A Review. Mar. Drugs 2021, 19, 549. https://doi.org/10.3390/md19100549
Coulombier N, Jauffrais T, Lebouvier N. Antioxidant Compounds from Microalgae: A Review. Marine Drugs. 2021; 19(10):549. https://doi.org/10.3390/md19100549
Chicago/Turabian StyleCoulombier, Noémie, Thierry Jauffrais, and Nicolas Lebouvier. 2021. "Antioxidant Compounds from Microalgae: A Review" Marine Drugs 19, no. 10: 549. https://doi.org/10.3390/md19100549