Neutral Lipids, Glycolipids, and Phospholipids, Isolated from Sandfish (Arctoscopus japonicus) Eggs, Exhibit Anti-Inflammatory Activity in LPS-Stimulated RAW264.7 Cells through NF-κB and MAPKs Pathways


Abstract
:1. Introduction
2. Results
2.1. Fatty Acid Analysis of A. japonicus Lipid-Fractions (Neutral Lipids, Glycolipids, and Phospholipids), Which were Isolated from A. japonicus Eggs
2.2. The Cytotoxicity of A. japonicus Lipid-Fractions (Neutral Lipids, Glycolipids, and Phospholipids) against RAW264.7 Cells
2.3. The Anti-Inflammatory Effect of A. japonicus Lipid-Fractions (Neutral Lipids, Glycolipids, and Phospholipids) against NO Production in LPS-Stimulated RAW264.7 Cells
2.4. The Anti-Inflammatory Effect of A. japonicus Lipid Fractions (Neutral Lipids, Glycolipids, and Phospholipids) against Immune-Associated Gene Expression in LPS-Stimulated RAW264.7 Cells
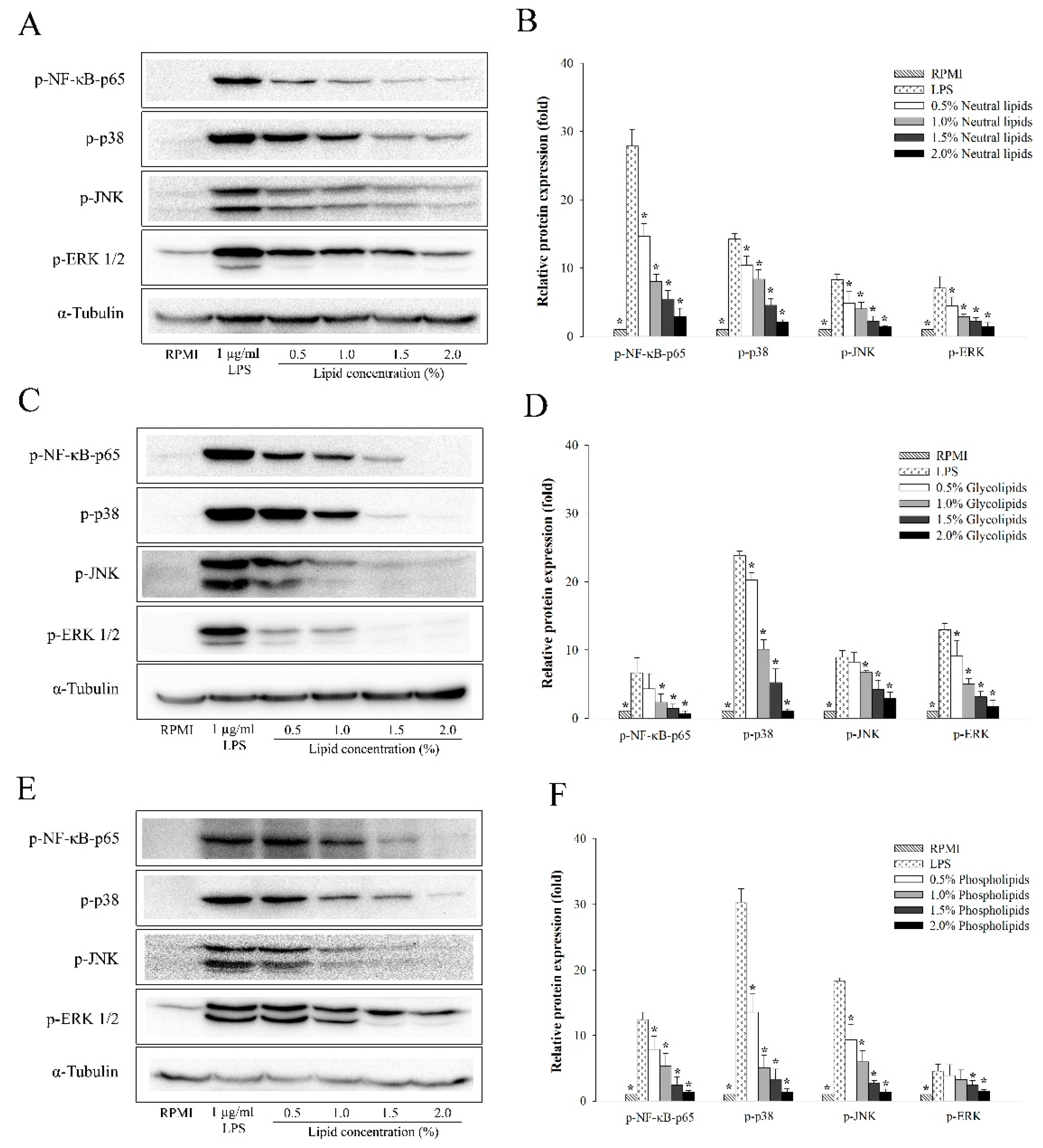
2.5. The Anti-Inflammatory Effects of A. japonicus Lipid Fractions (Neutral Lipids, Glycolipids, and Phospholipids) on the NF-κB and MAPKs Signaling Pathways of LPS-Stimulated RAW264.7 Cells
3. Discussion
4. Materials and Methods
4.1. Lipid Extraction and Separation
4.2. Fatty Acid Analysis
4.3. The Measurement of Cell Proliferation
4.4. The Measurement of Nitric Oxide (NO) Production
4.5. Qualitative Real-Time PCR
4.6. Western Blot Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sargent, J.R.; Tocher, D.R.; Bell, J.G. 4-The Lipids. In Fish Nutrition, 3rd ed.; Halver, J.E., Hardy, R.W., Eds.; Academic Press: San Diego, CA, USA, 2003; pp. 181–257. [Google Scholar]
- Tocher, D.R. Metabolism and functions of lipids and fatty acids in teleost fish. Rev. Fish Sci. 2003, 11, 107–184. [Google Scholar] [CrossRef]
- Lordan, R.; Tsoupras, A.; Zabetakis, I. Phospholipids of animal and marine origin: Structure, function, and anti-inflammatory properties. Molecules 2017, 22, 1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, R.J.; Tocher, D.R. The lipid composition and biochemistry of freshwater fish. Prog. Lipid Res. 1987, 26, 281–347. [Google Scholar] [CrossRef]
- Hutchins, P.M.; Barkley, R.M.; Murphy, R.C. Separation of cellular nonpolar neutral lipids by normal-phase chromatography and analysis by electrospray ionization mass spectrometry. J. Lipid Res. 2008, 49, 804–813. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.Y.; Park, W.K.; Suzuki, H. Analyses of glycolipids from fish, shellfish, and sea snake lipids by high-performance liquid chromatography. J. Agric. Food Chem. 1999, 47, 960–963. [Google Scholar] [CrossRef]
- Malhotra, R. Membrane glycolipids: Functional heterogeneity: A review. Biochem. Anal. Biochem. 2012, 1, 1000108. [Google Scholar] [CrossRef] [Green Version]
- Tocher, D.R.; Bendiksen, E.Å.; Campbell, P.J.; Bell, J.G. The role of phospholipids in nutrition and metabolism of teleost fish. Aquaculture 2008, 280, 21–34. [Google Scholar] [CrossRef] [Green Version]
- Wall, R.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. Fatty acids from fish: The anti-inflammatory potential of long-chain omega-3 fatty acids. Nutr. Rev. 2010, 68, 280–289. [Google Scholar] [CrossRef]
- Huynh, M.D.; Kitts, D.D.; Hu, C.; Trites, A.W. Comparison of fatty acid profiles of spawning and non-spawning Pacific herring, Clupea Harengus pallasi. Comp. Biochem. Physiol. B. Biochem. Mol. Biol. 2007, 146, 504–511. [Google Scholar] [CrossRef]
- Moriya, H.; Hosokawa, M.; Miyashita, K. Combination effect of herring roe lipids and proteins on plasma lipids and abdominal fat weight of mouse. J. Food Sci. 2007, 72, 231–234. [Google Scholar] [CrossRef]
- Tsoupras, A.; O’Keeffe, E.; Lordan, R.; Redfern, S.; Zabetakis, I. Bioprospecting for antithrombotic polar lipids from salmon, herring, and boarfish by-products. Foods 2019, 8, 416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garaffo, M.A.; Vassallo-Agius, R.; Nengas, Y.; Lembo, E.; Rando, R.; Maisano, R.; Dugo, G.; Giuffrida, D. Fatty acids profile, atherogenic (IA) and thrombogenic (IT) health lipid indices, of raw roe of Blue Fin Tuna (Thunnus thynnus L.) and their salted product “Bottarga”. Food Nutr. Sci. 2011, 2, 736–743. [Google Scholar]
- Torrecillas, S.; Montero, D.; Domínguez, D.; Robaina, L.; Izquierdo, M. Skin mucus fatty acid composition of gilthead sea bream (Sparus aurata): A descriptive study in fish fed low and high fish meal diets. Fishes 2019, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Ovissipour, M.; Rasco, B. Fatty acid and amino acid profiles of domestic and wild beluga (Huso huso) roe and impact on fertilization ratio. J. Aquac. Res. Dev. 2011, 2. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, K.A.S.; Linder, M.; Fanni, J.; Parmentier, M. Characterisation of the lipid fractions obtained by proteolytic and chemical extractions from rainbow trout (Oncorhynchus mykiss) roe. Process Biochem. 2008, 43, 376–383. [Google Scholar] [CrossRef]
- Tsoupras, A.; Lordan, R.; Demuru, M.; Shiels, K.; Saha, S.K.; Nasopoulou, C.; Zabetakis, I. Structural elucidation of Irish organic farmed salmon (Salmo salar) polar lipids with antithrombotic activities. Mar. Drugs 2018, 16, 176. [Google Scholar] [CrossRef] [Green Version]
- Tsoupras, A.; Lordan, R.; Shiels, K.; Saha, S.K.; Nasopoulou, C.; Zabetakis, I. In vitro antithrombotic properties of salmon (Salmo salar) phospholipids in a novel food-grade extract. Mar. Drugs 2019, 17, 62. [Google Scholar] [CrossRef] [Green Version]
- Wen, M.; Xu, J.; Ding, L.; Zhang, L.; Du, L.; Wang, J.; Wang, Y.; Xue, C. Eicosapentaenoic acid-enriched phospholipids improve Aβ1–40-induced cognitive deficiency in a rat model of Alzheimer’s disease. J. Funct. Foods 2016, 24, 537–548. [Google Scholar] [CrossRef]
- Holub, D.J.; Holub, B.J. Omega-3 fatty acids from fish oils and cardiovascular disease. Mol. Cell Biochem. 2004, 263, 217–225. [Google Scholar] [CrossRef]
- Molfino, A.; Amabile, M.I.; Lionetto, L.; Spagnoli, A.; Ramaccini, C.; De Luca, A.; Simmaco, M.; Monti, M.; Muscaritoli, M. DHA oral supplementation modulates serum epoxydocosapentaenoic acid (EDP) levels in breast cancer patients. Oxidative Med. Cell. Longev. 2019, 2019, 7. [Google Scholar] [CrossRef]
- Paixão, E.M.d.S.; Oliveira, A.C.d.M.; Pizato, N.; Muniz-Junqueira, M.I.; Magalhães, K.G.; Nakano, E.Y.; Ito, M.K. The effects of EPA and DHA enriched fish oil on nutritional and immunological markers of treatment naïve breast cancer patients: A randomized double-blind controlled trial. Nutr. J. 2017, 16, 71. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhang, L.; Tian, X.; Xing, J. Docosahexaenoic acid has an anti-diabetic effect in streptozotocin-induced diabetic mice. Int. J. Clin. Exp. Med. 2014, 7, 3021–3029. [Google Scholar] [PubMed]
- Vitlov Uljević, M.; Starčević, K.; Mašek, T.; Bočina, I.; Restović, I.; Kević, N.; Racetin, A.; Kretzschmar, G.; Grobe, M.; Vukojević, K.; et al. Dietary DHA/EPA supplementation ameliorates diabetic nephropathy by protecting from distal tubular cell damage. Cell Tissue Res. 2019, 378, 301–317. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.J.; Valenzuela, C.A.; De Souza, C.O.; Yaqoob, P.; Miles, E.A.; Calder, P.C. Comparative anti-inflammatory effects of plant- and marine-derived omega-3 fatty acids explored in an endothelial cell line. BBA-Mol. Cell Biol. 2020, 1865, 158662. [Google Scholar] [CrossRef] [PubMed]
- Si, T.L.; Liu, Q.; Ren, Y.F.; Li, H.; Xu, X.Y.; Li, E.H.; Pan, S.Y.; Zhang, J.L.; Wang, K.X. Enhanced anti-inflammatory effects of DHA and quercetin in lipopolysaccharide-induced RAW264.7 macrophages by inhibiting NF-kB and MAPK activation. Mol. Med. Rep. 2016, 14, 499–508. [Google Scholar] [CrossRef] [Green Version]
- Lordan, R.; Redfern, S.; Tsoupras, A.; Zabetakis, I. Inflammation and cardiovascular disease: Are marine phospholipids the answer? Food Funct. 2020, 11, 2861–2885. [Google Scholar] [CrossRef]
- Murota, K.; Takagi, M.; Watanabe, Y.; Tokumura, A.; Ohkubo, T. Roe-derived phospholipid administration enhances lymphatic docosahexaenoic acid-containing phospholipid absorption in unanesthetized rats. Prostag. Leukotr. Ess. 2018, 139, 40–48. [Google Scholar] [CrossRef]
- Guha, M.; Mackman, N. LPS induction of gene expression in human monocytes. Cell. Signal. 2001, 13, 85–94. [Google Scholar] [CrossRef]
- Babcock, T.A.; Helton, W.S.; Hong, D.; Espat, N.J. Omega-3 fatty acid lipid emulsion reduces LPS-stimulated macrophage TNF-a production. Surg. Infect. (Larchmt) 2002, 3, 145–149. [Google Scholar] [CrossRef]
- Novak, T.E.; Babcock, T.A.; Jho, D.H.; Helton, W.S.; Espat, N.J. NF-kB inhibition by omega -3 fatty acids modulates LPS-stimulated macrophage TNF-a transcription. Am. J. Physiol. Lung Cell. Mol. Physiol. 2003, 284, L84–L89. [Google Scholar] [CrossRef]
- Weldon, S.M.; Mullen, A.C.; Loscher, C.E.; Hurley, L.A.; Helen, M.; Roche, H.M. Docosahexaenoic acid induces an anti-inflammatory profile in lipopolysaccharide-stimulated human THP-1 macrophages more effectively than eicosapentaenoic acid. J. Nutr. Biochem. 2007, 18, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Etherton, T.D.; Martin, K.R.; Vanden Heuvel, J.P.; Gillies, P.J.; West, S.G.; Kris-Etherton, P.M. Anti-inflammatory effects of polyunsaturated fatty acids in THP-1 cells. Biochem. Biophys. Res. Commun. 2005, 336, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.J.; Chiu, K.C.; Fu, M.; Chu, A.; Helton, S. Fish oil modulates macrophage P44/P42 mitogen-activated protein kinase activity induced by lipopolysaccharide. J. Parenter. Enteral. Nutr. 2000, 24, 159–163. [Google Scholar] [CrossRef]
- Lee, S.I.; Yang, J.H.; Yoon, S.C.; Chun, Y.Y.; Kim, J.B.; Cha, H.K.; Choi, Y.M. Biomass estimation of sailfin sandfish, Arctoscopus japonicus, in Korean waters. Korean J. Fish. Aquat. Sci. 2009, 42, 487–493. [Google Scholar]
- Shirai, S.M.; Kuranaga, R.; Sugiyama, H.; Higuchi, M. Population structure of the sailfin sandfish, Arctoscopus japonicus (Trichodontidae), in the Sea of Japan. Ichthyol. Res. 2006, 53, 357–368. [Google Scholar] [CrossRef]
- Jang, H.L.; Liceaga, A.M.; Yoon, K.Y. Purification, characterisation and stability of an antioxidant peptide derived from sandfish (Arctoscopus japonicus) protein hydrolysates. J. Funct. Foods 2016, 20, 433–442. [Google Scholar] [CrossRef]
- Jang, H.L.; Shin, S.R.; Yoon, K.Y. Hydrolysis conditions for antioxidant peptides derived from enzymatic hydrolysates of sandfish (Arctoscopus japonicus). Food Sci. Biotechnol. 2017, 26, 1191–1197. [Google Scholar] [CrossRef]
- Jang, H.L.; Liceaga, A.M.; Yoon, K.Y. Isolation and characteristics of anti-inflammatory peptides from enzymatic hydrolysates of sandfish (Arctoscopus japonicus) protein. J. Aquat. Food. Prod. Technol. 2017, 26, 234–244. [Google Scholar] [CrossRef]
- Jang, H.L.; Young Yoon, K. Optimal conditions of enzymatic hydrolysis for producing anti-inflammatory peptides from sandfish (Arctoscopus japonicus) hydrolysate. Korean J. Food Sci. Technol. 2018, 50, 203–208. [Google Scholar]
- Ishihara, Y.; Watanabe, F. Lipid content and fatty acid composition of Japanese sandfish Arctoscopus japonicus caught offshore of Tottori Prefecture, Japan. Nippon Suisan Gakk. 2013, 79, 229–231. [Google Scholar] [CrossRef]
- Rod-In, W.; Monmai, C.; Lee, S.M.; Jung, S.K.; You, S.; Park, W.J. Anti-inflammatory effects of lipids extracted from Arctoscopus japonicus eggs on LPS-stimulated RAW264.7 cells. Mar. Drugs 2019, 17, 580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prabhakara Rao, P.; Balaswamy, K.; Narsing Rao, G.; Jyothirmayi, T. Lipid classes, fatty acid and phospholipid composition of roe lipids from Catla catla and Cirrhinus mrigala. Int. Food Res. J. 2013, 20, 275–279. [Google Scholar]
- Prabhakara Rao, P.G.; Jyothirmayi, T.; Karuna, M.S.L.; Prasad, R.B.N. Studies on lipid profiles and fatty acid composition of roe from rohu (Labeo rohita) and murrel (Channa striatus). J. Oleo Sci. 2010, 59, 515–519. [Google Scholar] [PubMed] [Green Version]
- Shirai, N.; Higuchi, T.; Suzuki, H. Analysis of lipid classes and the fatty acid composition of the salted fish roe food products, Ikura, Tarako, Tobiko and Kazunoko. Food Chem. 2006, 94, 61–67. [Google Scholar] [CrossRef]
- Majumder, A.; Misra, K.K.; Homechoudhuri, S. Lipid and fatty acid composition in the flesh of an edible marine fish: Amadi (Coilia reynaldi). Proc. Zool. Soc. 2013, 66, 86–91. [Google Scholar] [CrossRef]
- Oxley, A.; Torstensen, B.E.; Rustan, A.C.; Olsen, R.E. Enzyme activities of intestinal triacylglycerol and phosphatidylcholine biosynthesis in Atlantic salmon (Salmo salar L.). Comp. Biochem. Physiol. B 2005, 141, 77–87. [Google Scholar] [CrossRef]
- Calder, P.C. n-3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. Am. J. Clin. Nutr. 2006, 83, 1505S–1519S. [Google Scholar] [CrossRef]
- Nasopoulou, C.; Tsoupras, A.B.; Karantonis, H.C.; Demopoulos, C.A.; Zabetakis, I. Fish polar lipids retard atherosclerosis in rabbits by down-regulating PAF biosynthesis and up-regulating PAF catabolism. Lipids Health Dis. 2011, 10, 213. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Song, C.; Koh, W.; Yon, G.; Kim, Y.; Ryu, S.; Kwon, H.; Lee, K. Anti-inflammatory effects of the Zingiber officinale roscoe constituent 12-dehydrogingerdione in lipopolysaccharide-stimulated Raw264.7 Cells. Phytother Res. 2013, 27, 1200–1205. [Google Scholar] [CrossRef]
- Ahmad, T.B.; Rudd, D.; Kotiw, M.; Liu, L.; Benkendorff, K. Correlation between fatty acid profile and anti-inflammatory activity in common Australian seafood by-products. Mar. Drugs 2019, 17, 155. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.K.; Rasmussen, H.E.; Ehlers, S.J.; Blobaum, K.R.; Lu, F.; Schlegal, V.L.; Carr, T.P.; Lee, J.Y. Repression of proinflammatory gene expression by lipid extract of Nostoc commune var sphaeroides Kutzing, a blue-green alga, via inhibition of nuclear factor-kB in RAW264.7 macrophages. Nutr. Res. 2008, 28, 83–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, D.H.; Kim, K.B.W.R.; Kim, M.J.; Kang, B.K.; Ahn, D.H. Skipjack tuna (Katsuwonus pelamis) eyeball oil exerts an anti-inflammatory effect by inhibiting NF-kB and MAPK activation in LPS-induced RAW264.7 cells and croton oil-treated mice. Int. Immunopharmacol. 2016, 40, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Shen, X.; Chen, G.; Cao, X.; Yang, J. A comparative study of the effects upon LPS induced macrophage RAW264.7 inflammation in vitro of the lipids of Hippocampus trimaculatus Leach. J. Oleo Sci. 2015, 64, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Fernando, I.P.S.; Sanjeewa, K.K.A.; Kim, H.-S.; Kim, S.-Y.; Lee, S.-H.; Lee, W.W.; Jeon, Y.-J. Identification of sterols from the soft coral Dendronephthya gigantea and their anti-inflammatory potential. Environ. Toxicol. Pharmacol. 2017, 55, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Banskota, A.H.; Stefanova, R.; Sperker, S.; Lall, S.P.; Craigie, J.S.; Hafting, J.T.; Critchley, A.T. Polar lipids from the marine macroalga Palmaria palmata inhibit lipopolysaccharide-induced nitric oxide production in RAW264.7 macrophage cells. Phytochemistry 2014, 101, 101–108. [Google Scholar] [CrossRef]
- Lopes, G.; Daletos, G.; Proksch, P.; Andrade, P.B.; Valentão, P. Anti-inflammatory potential of monogalactosyl diacylglycerols and a monoacylglycerol from the edible brown seaweed Fucus spiralis Linnaeus. Mar. Drugs 2014, 12, 1406–1418. [Google Scholar] [CrossRef]
- Ishii, M.; Nakahara, T.; Araho, D.; Murakami, J.; Nishimura, M. Glycolipids from spinach suppress LPS-induced vascular inflammation through eNOS and NK-κB signaling. Biomed. Pharmacother. 2017, 91, 111–120. [Google Scholar] [CrossRef]
- Tak, P.P.; Firestein, G.S. NF-κB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef]
- McGrath, M.A.; Harnett, M.M.; Thalhamer, T. MAPKs and their relevance to arthritis and inflammation. Rheumatology 2008, 47, 409–414. [Google Scholar]
- Zhu, J.; Luo, C.; Wang, P.; He, Q.; Zhou, J.; Peng, H. Saikosaponin A mediates the inflammatory response by inhibiting the MAPK and NF-κB pathways in LPS-stimulated RAW264.7 cells. Exp. Ther. Med. 2013, 5, 1345–1350. [Google Scholar] [CrossRef] [Green Version]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Phys. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garces, R.; Mancha, M. One-step lipid extraction and fatty acid methyl esters preparation from fresh plant tissues. Anal. Biochem. 1993, 211, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Park, W.J.; Kothapalli, K.S.D.; Lawrence, P.; Tyburczy, C.; Brenna, J.T. An alternate pathway to long-chain polyunsaturates: The FADS2 gene product Delta8-desaturates 20:2n-6 and 20:3n-3. J. Lipid Res. 2009, 50, 1195–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.K.; Cho, M.L.; Karnjanapratum, S.; Shin, I.S.; You, S.G. In vitro and in vivo immunomodulatory activity of sulfated polysaccharides from Enteromorpha prolifera. Int. J. Biol. Macromol. 2011, 49, 1051–1058. [Google Scholar] [CrossRef]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15N]nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid | Neutral Lipids | Glycolipids | Phospholipids |
|---|---|---|---|
| Saturated fatty acid (SFA) | |||
| 16:0 | 33.17 ± 0.28 aA | 26.30 ± 0.27 aB | 19.01 ± 1.27 bC |
| 18:0 | 3.82 ± 0.05 fB | 6.23 ± 0.06 gA | 6.23 ± 0.42 eA |
| Total SFAs | 37.00 ± 0.31 | 32.53 ± 0.33 | 25.24 ± 1.68 |
| Monounsaturated fatty acid (MUFA) | |||
| 16:1n7 | 9.874 ± 0.28 cA | 7.51 ± 0.05 fB | - |
| 18:1n9 | 25.94 ± 0.50 bA | 22.18 ± 0.23 bB | 15.83 ± 1.00 cC |
| 18:1n7 | 10.07 ± 0.14 cB | 8.98 ± 0.02 eC | 12.07 ± 0.73 dA |
| 20:1 | 1.05 ± 0.06 hA | - | - |
| Total MUFAs | 46.80 ± 0.79 | 38.68 ± 0.26 | 27.89 ± 1.73 |
| Polyunsaturated fatty acid (PUFA) | |||
| 18:2n6 (LA) | 1.14 ± 0.02 hA | - | - |
| 18:3n3 (ALA) | 0.44 ± 0.02 iA | - | - |
| 20:3n3 | 1.99 ± 0.08 gC | 3.49 ± 0.06 hB | 6.21 ± 0.25 eA |
| 20:5n3 (EPA) | 7.83 ± 0.35 dC | 12.07 ± 0.21 dB | 18.96 ± 1.27 bA |
| 22:6n3 (DHA) | 4.80 ± 0.65 eC | 13.23 ± 0.33 cB | 21.70 ± 1.92 aA |
| Total PUFAs | 16.20 ± 0.70 | 28.79 ± 0.58 | 46.86 ± 3.40 |
| Target Gene | Sequence (from 5′ to 3′) | |
|---|---|---|
| IL-1β | Forward Reverse | GGGCCTCAAAGGAAAGAATC TACCAGTTGGGGAACTCTGC |
| iNOS | Forward Reverse | TTCCAGAATCCCTGGACAAG TGGTCAAACTCTTGGGGTTC |
| IL-6 | Forward Reverse | AGTTGCCTTCTTGGGACTGA CAGAATTGCCATTGCACAAC |
| COX-2 | Forward Reverse | AGAAGGAAATGGCTGCAGAA GCTCGGCTTCCAGTATTGAG |
| TNF-α | Forward Reverse | ATGAGCACAGAAAGCATGATC TACAGGCTTGTCACTCGAATT |
| β-Actin | Forward Reverse | CCACAGCTGAGAGGAAATC AAGGAAGGCTGGAAAAGAGC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rod-in, W.; Monmai, C.; Shin, I.-s.; You, S.; Park, W.J. Neutral Lipids, Glycolipids, and Phospholipids, Isolated from Sandfish (Arctoscopus japonicus) Eggs, Exhibit Anti-Inflammatory Activity in LPS-Stimulated RAW264.7 Cells through NF-κB and MAPKs Pathways. Mar. Drugs 2020, 18, 480. https://doi.org/10.3390/md18090480
Rod-in W, Monmai C, Shin I-s, You S, Park WJ. Neutral Lipids, Glycolipids, and Phospholipids, Isolated from Sandfish (Arctoscopus japonicus) Eggs, Exhibit Anti-Inflammatory Activity in LPS-Stimulated RAW264.7 Cells through NF-κB and MAPKs Pathways. Marine Drugs. 2020; 18(9):480. https://doi.org/10.3390/md18090480
Chicago/Turabian StyleRod-in, Weerawan, Chaiwat Monmai, Il-sik Shin, SangGuan You, and Woo Jung Park. 2020. "Neutral Lipids, Glycolipids, and Phospholipids, Isolated from Sandfish (Arctoscopus japonicus) Eggs, Exhibit Anti-Inflammatory Activity in LPS-Stimulated RAW264.7 Cells through NF-κB and MAPKs Pathways" Marine Drugs 18, no. 9: 480. https://doi.org/10.3390/md18090480