Laminarin Pretreatment Provides Neuroprotection against Forebrain Ischemia/Reperfusion Injury by Reducing Oxidative Stress and Neuroinflammation in Aged Gerbils


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
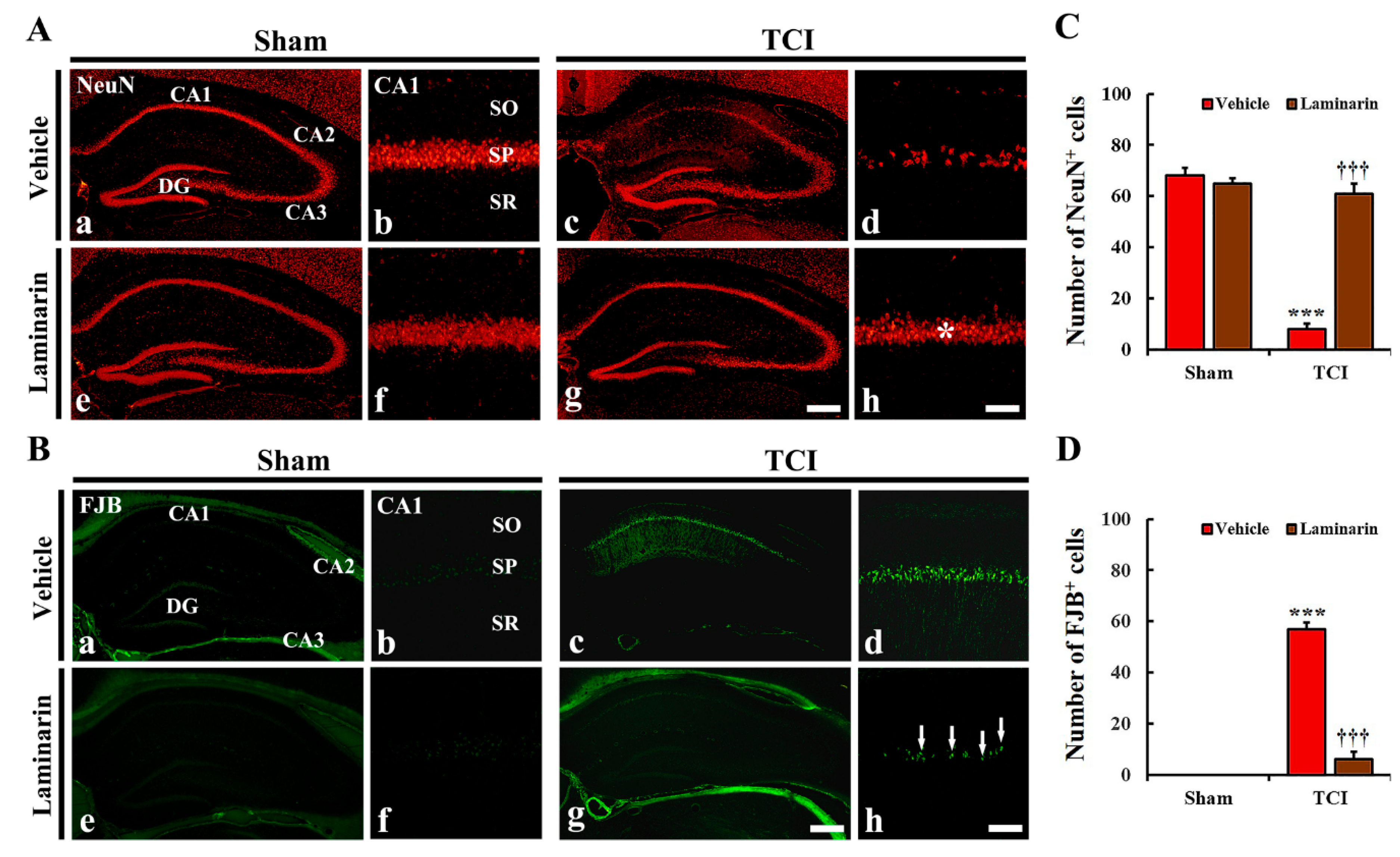
2.1. Protection of Neurons from IR Injury by Laminarin
2.1.1. Neuronal Nuclear Antigen (NeuN) Immunoreactive (+) Cells
2.1.2. Fluoro-Jade B (FJB)+ Cells
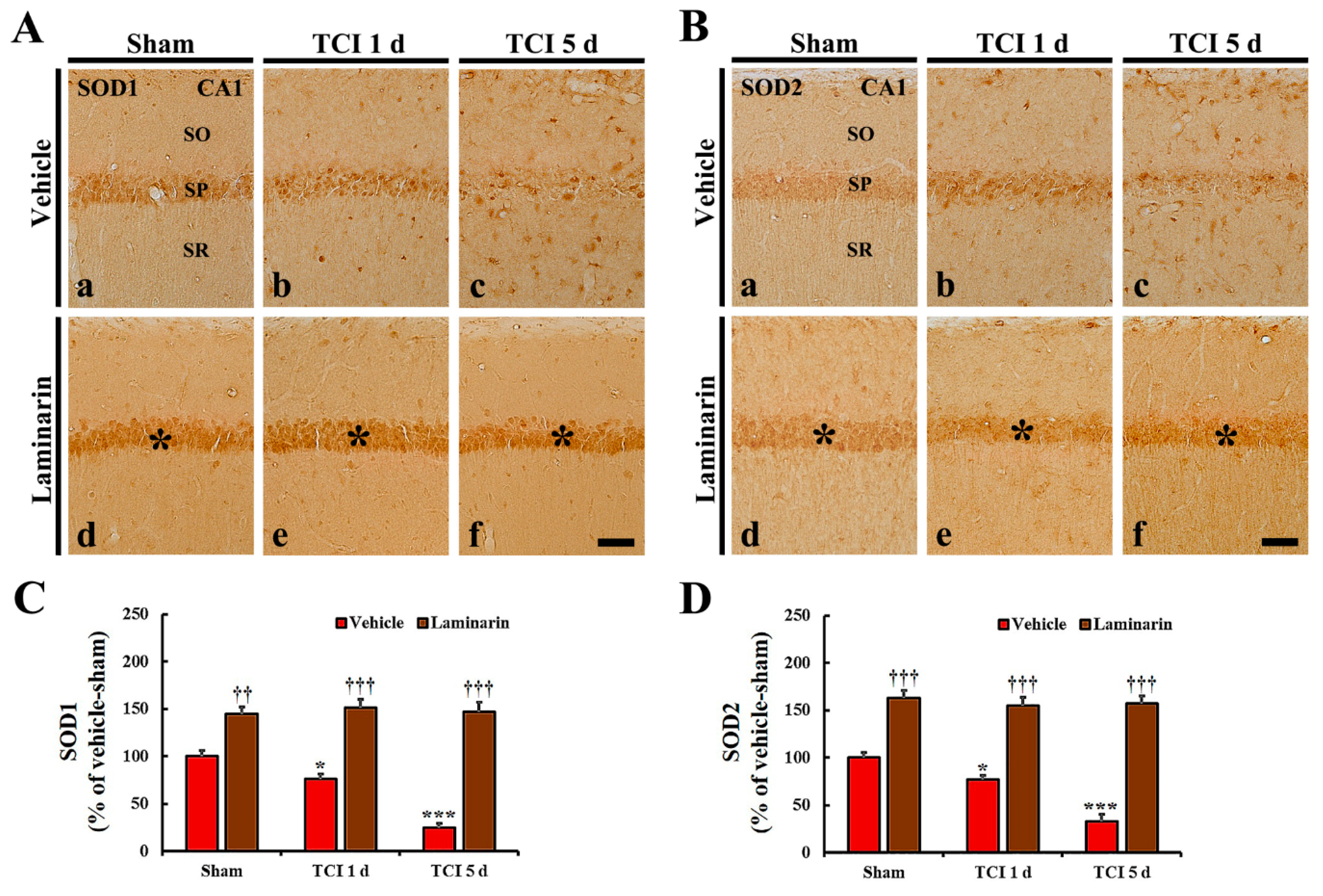
2.2. Increases of Superoxide Dismutase (SODs) Expression by Laminarin
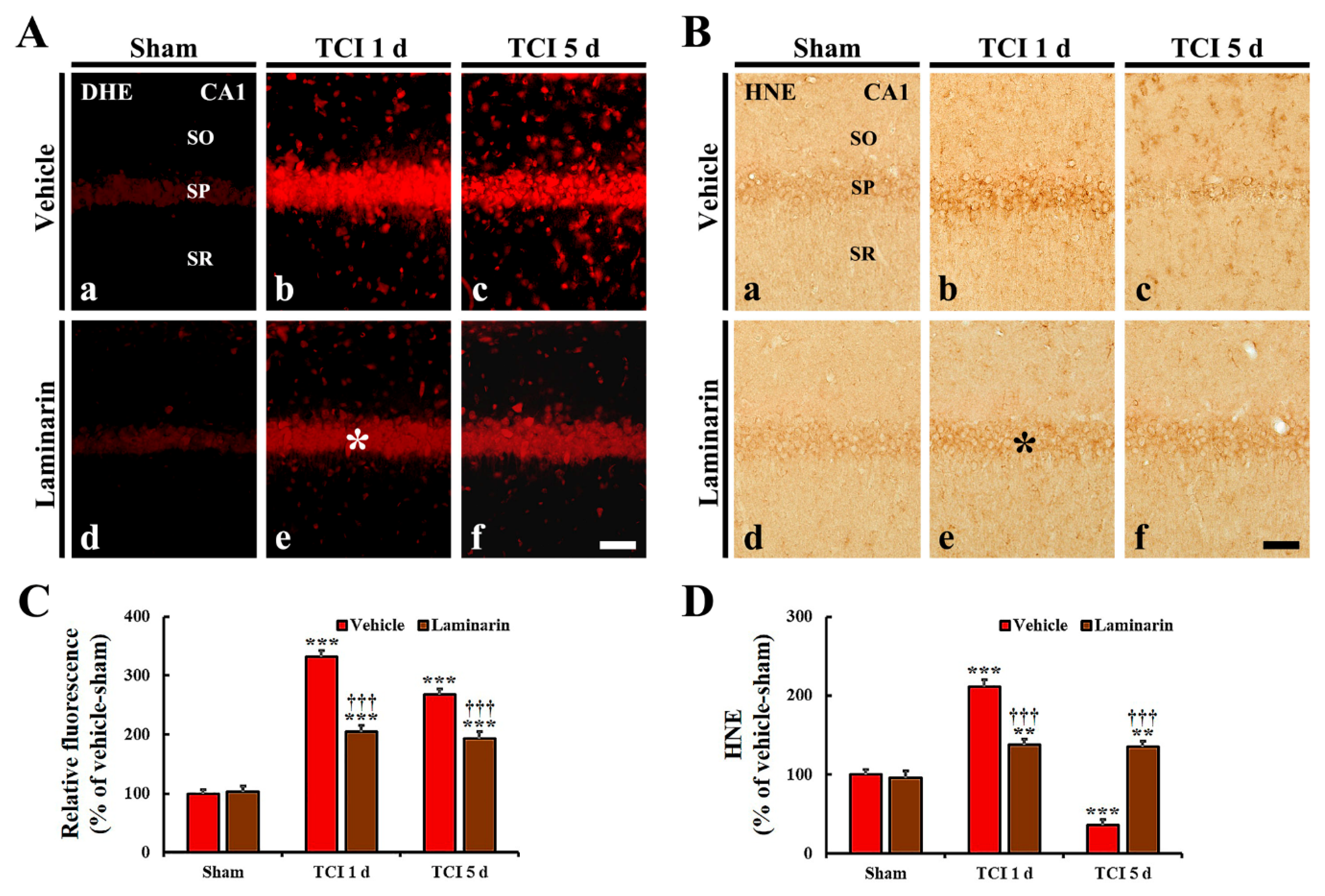
2.3. Attenuation of IR-Induced Oxidative Stress by Laminarin
2.3.1. Dihydroethidium (DHE) Fluorescence
2.3.2. 4-Hydroxy-2-Nonenal (HNE) Immunoreactivity
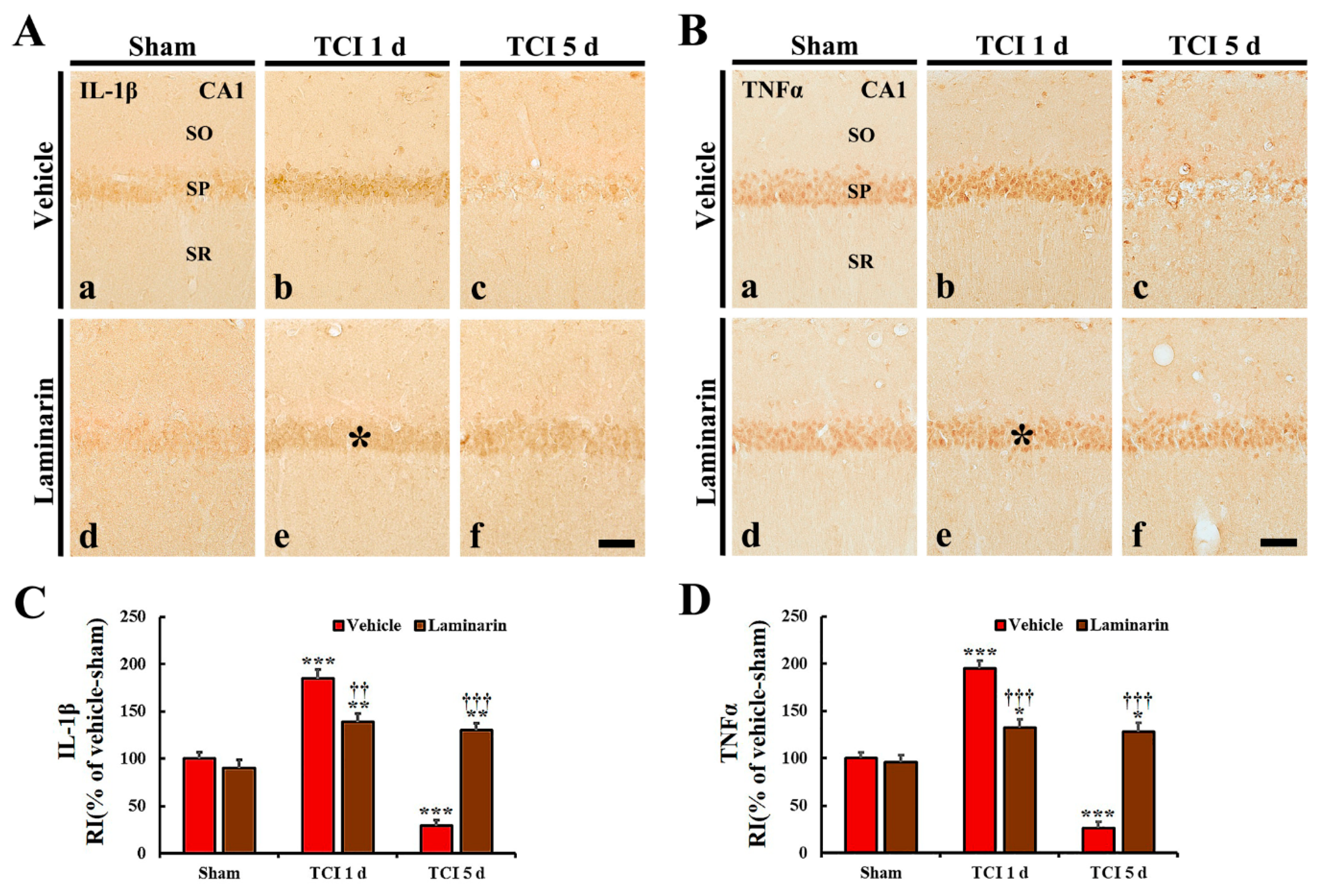
2.4. Reduction of IR-Induced Neuroinflammation by Laminarin
2.4.1. Pro-Inflammatory Cytokine Immunoreactivities
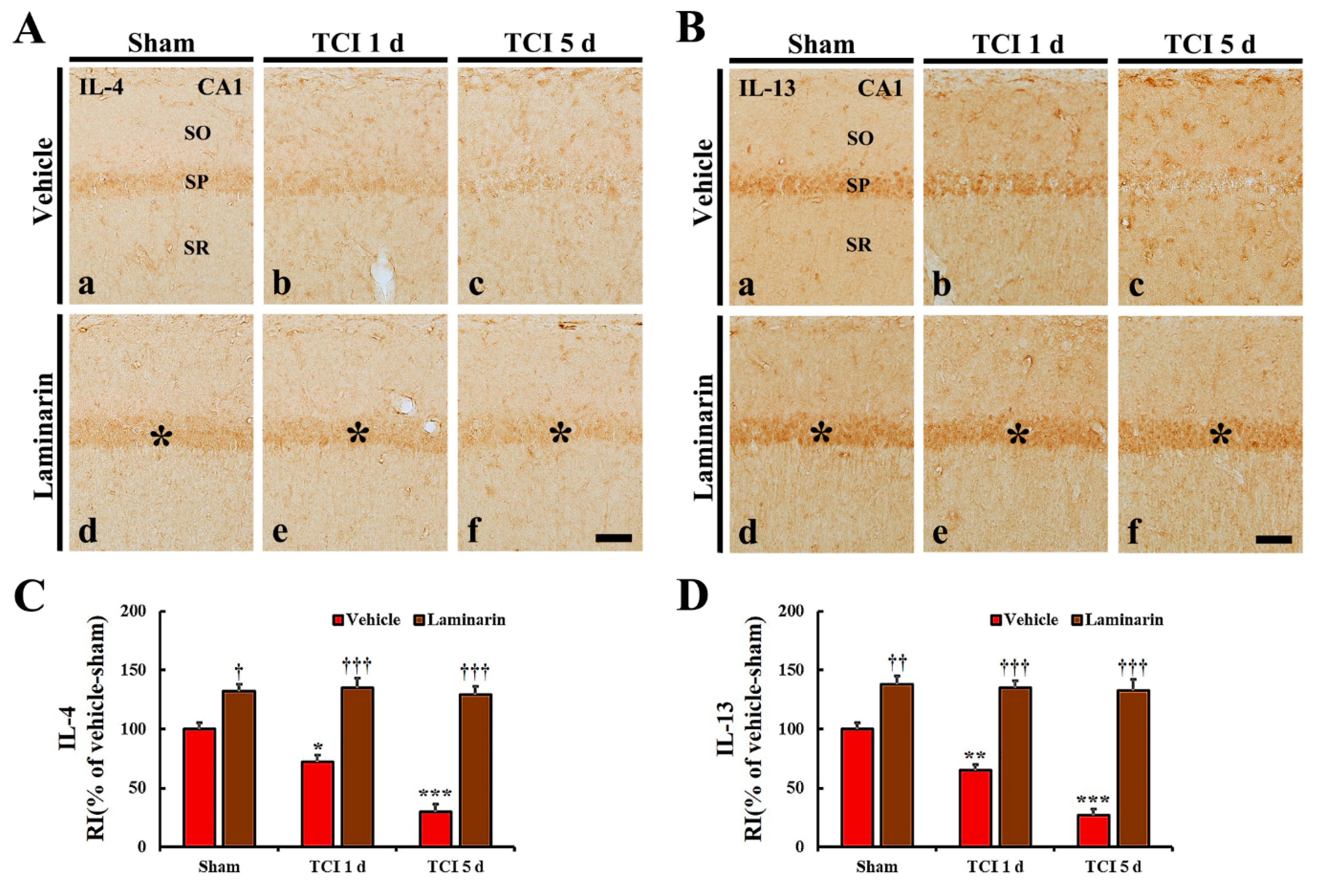
2.4.2. Anti-Inflammatory Cytokine Immunoreactivities
3. Discussion
4. Materials and Methods
4.1. Experimental Groups and Laminarin Pretreatment
4.2. Surgery of IR
4.3. Preparation of Brain Sections
4.4. NeuN Immunofluorescence and FJB Histofluorescence Staining
4.5. DHE Staining for Superoxide
4.6. Immunohistochemistry (IHC)
4.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Burad, J.; Bhakta, P.; George, J.; Kiruchennan, S. Development of acute ischemic stroke in a patient with acute respiratory distress syndrome (ARDS) resulting from H1N1 pneumonia. Acta Anaesthesiol. Taiwanica 2012, 50, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Kawai, K.; Nitecka, L.; Ruetzler, C.A.; Nagashima, G.; Joó, F.; Mies, G.; Nowak, T.S.; Saito, N.; Lohr, J.M.; Klatzo, I. Global Cerebral Ischemia Associated with Cardiac Arrest in the Rat: I. Dynamics of Early Neuronal Changes. Br. J. Pharmacol. 1992, 12, 238–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-C.; Park, J.H.; Ahn, J.H.; Kim, I.H.; Cho, J.H.; Choi, J.H.; Yoo, K.-Y.; Lee, C.H.; Hwang, I.K.; Cho, J.H.; et al. New GABAergic Neurogenesis in the Hippocampal CA1 Region of a Gerbil Model of Long-Term Survival after Transient Cerebral Ischemic Injury. Brain Pathol. 2015, 26, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Von Kummer, R.; Bourquain, H.; Bastianello, S.; Bozzao, L.; Manelfe, C.; Meier, D.; Hacke, W. Early Prediction of Irreversible Brain Damage after Ischemic Stroke at CT. Radiology 2001, 219, 95–100. [Google Scholar] [CrossRef]
- Thrift, A.G.; Cadilhac, D.A.; Thayabaranathan, T.; Howard, G.; Howard, V.J.; Rothwell, P.M.; Donnan, G.A. Global stroke statistics. Int. J. Stroke 2014, 9, 6–18. [Google Scholar] [CrossRef] [Green Version]
- Harukuni, I.; Bhardwaj, A. Mechanisms of Brain Injury after Global Cerebral Ischemia. Neurol. Clin. 2006, 24, 1–21. [Google Scholar] [CrossRef]
- Sugawara, T.; Lewén, A.; Noshita, N.; Gasche, Y.; Chan, P.H. Effects of Global Ischemia Duration on Neuronal, Astroglial, Oligodendroglial, and Microglial Reactions in the Vulnerable Hippocampal CA1 Subregion in Rats. J. Neurotrauma 2002, 19, 85–98. [Google Scholar] [CrossRef]
- Iadecola, C.; Alexander, M. Cerebral ischemia and inflammation. Curr. Opin. Neurol. 2001, 14, 89–94. [Google Scholar] [CrossRef]
- Saito, A.; Maier, C.M.; Narasimhan, P.; Nishi, T.; Song, Y.S.; Yu, F.; Liu, J.; Lee, Y.-S.; Nito, C.; Kamada, H.; et al. Oxidative Stress and Neuronal Death/Survival Signaling in Cerebral Ischemia. Mol. Neurobiol. 2005, 31, 105–116. [Google Scholar] [CrossRef]
- Shichita, T.; Sakaguchi, R.; Suzuki, M.; Yoshimura, A. Post-Ischemic Inflammation in the Brain. Front. Immunol. 2012, 3, 132. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.-H.; Li, W. Targeting oxidative stress for the treatment of ischemic stroke: Upstream and downstream therapeutic strategies. Brain Circ. 2016, 2, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Peng, T.; Jiang, Y.; Farhan, M.; Lazarovici, P.; Chen, L.; Zheng, W. Anti-inflammatory Effects of Traditional Chinese Medicines on Preclinical in vivo Models of Brain Ischemia-Reperfusion-Injury: Prospects for Neuroprotective Drug Discovery and Therapy. Front. Pharmacol. 2019, 10, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surgucheva, I.; Sharov, V.S.; Surguchov, A. γ-Synuclein: Seeding of α-Synuclein Aggregation and Transmission between Cells. Biochemistry 2012, 51, 4743–4754. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Wang, R.-H.; Wang, H.; Long, C.-L.; Wang, H. Brain protection against ischemic stroke using choline as a new molecular bypass treatment. Acta Pharmacol. Sin. 2015, 36, 1416–1425. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Park, J.H.; Maharjan, S.; Park, J.A.; Choi, K.-S.; Park, H.; Jeong, Y.; Ahn, J.H.; Kim, I.H.; Lee, J.-C.; et al. Sac-1004, a vascular leakage blocker, reduces cerebral ischemia—Reperfusion injury by suppressing blood–brain barrier disruption and inflammation. J. Neuroinflamm. 2017, 14, 122. [Google Scholar] [CrossRef]
- Zhang, W.; Song, J.; Yan, R.; Li, L.; Xiao, Z.-Y.; Zhou, W.-X.; Wang, Z.-Z.; Xiao, W.; Du, G.-H. Diterpene ginkgolides protect against cerebral ischemia/reperfusion damage in rats by activating Nrf2 and CREB through PI3K/Akt signaling. Acta Pharmacol. Sin. 2018, 39, 1259–1272. [Google Scholar] [CrossRef]
- Benjamin, E.J.; Blaha, M.J.; Chiuve, S.E.; Cushman, M.; Das, S.R.; Deo, R.; De Ferranti, S.D.; Floyd, J.; Fornage, M.; Gillespie, C.; et al. Heart Disease and Stroke Statistics—2017 Update: A Report From the American Heart Association. Circulation 2017, 135, e146–e603. [Google Scholar] [CrossRef]
- Zhou, Z.; Ji, X.; Zhang, L.; Liu, R.; Liu, Y.; Song, J.; Dong, H.; Jia, Y. Potential targets for protecting against hippocampal cell apoptosis after transient cerebral ischemia-reperfusion injury in aged rats. Neural Regen. Res. 2014, 9, 1122–1128. [Google Scholar] [CrossRef]
- Saucier, D.M.; Yager, J.Y.; Armstrong, E.A.; Keller, A.; Shultz, S.R. Enriched environment and the effect of age on ischemic brain damage. Brain Res. 2007, 1170, 31–38. [Google Scholar] [CrossRef]
- Xu, K.; Puchowicz, M.A.; Sun, X.; Lamanna, J.C. Mitochondrial Dysfunction in Aging Rat Brain Following Transient Global Ischemia. Single Mol. Single Cell Seq. 2008, 614, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.; Mendelow, A.D.; Perry, R.H.; Chambers, I.; James, O.F.W. Experimental Stroke and Neuroprotection in the Aging Rat Brain. Stroke 1995, 26, 1072–1078. [Google Scholar] [CrossRef] [PubMed]
- Pohl, F.; Kong-Thoo-Lin, P. The Potential Use of Plant Natural Products and Plant Extracts with Antioxidant Properties for the Prevention/Treatment of Neurodegenerative Diseases: In Vitro, In Vivo and Clinical Trials. Molecule 2018, 23, 3283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, P.-F.; Zhang, Z.; Wang, F.; Chen, J.-G. Natural compounds from traditional medicinal herbs in the treatment of cerebral ischemia/reperfusion injury. Acta Pharmacol. Sin. 2010, 31, 1523–1531. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.H.; Shin, M.C.; Kim, D.W.; Kim, H.; Song, M.; Lee, T.-K.; Lee, J.-C.; Kim, H.; Cho, J.H.; Kim, Y.-M.; et al. Antioxidant Properties of Fucoidan Alleviate Acceleration and Exacerbation of Hippocampal Neuronal Death Following Transient Global Cerebral Ischemia in High-Fat Diet-Induced Obese Gerbils. Int. J. Mol. Sci. 2019, 20, 554. [Google Scholar] [CrossRef] [Green Version]
- Che, N.; Ma, Y.; Xin, Y. Protective Role of Fucoidan in Cerebral Ischemia-Reperfusion Injury through Inhibition of MAPK Signaling Pathway. Biomol. Ther. 2016, 25, 272–278. [Google Scholar] [CrossRef] [Green Version]
- Kang, G.H.; Yan, B.C.; Cho, G.-S.; Kim, W.-K.; Lee, C.H.; Cho, J.H.; Kim, M.; Kang, I.-J.; Won, M.-H.; Lee, J.-C. Neuroprotective effect of fucoidin on lipopolysaccharide accelerated cerebral ischemic injury through inhibition of cytokine expression and neutrophil infiltration. J. Neurol. Sci. 2012, 318, 25–30. [Google Scholar] [CrossRef]
- Kim, H.; Ahn, J.H.; Song, M.; Kim, D.W.; Lee, T.-K.; Lee, J.-C.; Kim, Y.-M.; Kim, J.-D.; Cho, J.H.; Hwang, I.K.; et al. Pretreated fucoidan confers neuroprotection against transient global cerebral ischemic injury in the gerbil hippocampal CA1 area via reducing of glial cell activation and oxidative stress. Biomed. Pharmacother. 2019, 109, 1718–1727. [Google Scholar] [CrossRef]
- Lee, T.-K.; Ahn, J.H.; Park, C.; Kim, B.; Park, Y.; Lee, J.-C.; Park, J.H.; Yang, G.; Shin, M.; Cho, J.; et al. Pre-Treatment with Laminarin Protects Hippocampal CA1 Pyramidal Neurons and Attenuates Reactive Gliosis Following Transient Forebrain Ischemia in Gerbils. Mar. Drugs 2020, 18, 52. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.-H.; Kim, Y.-W.; Kim, H.B.; Lee, B.J.; Lee, D.S. Anti-apoptotic Activity of Laminarin Polysaccharides and their Enzymatically Hydrolyzed Oligosaccharides from Laminaria japonica. Biotechnol. Lett. 2006, 28, 439–446. [Google Scholar] [CrossRef]
- Cheng, D.; Liang, B.; Li, M.; Jin, M. Influence of Laminarin polysaccahrides on oxidative damage. Int. J. Biol. Macromol. 2011, 48, 63–66. [Google Scholar] [CrossRef]
- Neyrinck, A.M.; Mouson, A.; Delzenne, N.M. Dietary supplementation with laminarin, a fermentable marine β (1–3) glucan, protects against hepatotoxicity induced by LPS in rat by modulating immune response in the hepatic tissue. Int. Immunopharmacol. 2007, 7, 1497–1506. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Xu, L.; Zhang, W.; Cai, Y.; Jang, B.; Oh, J.; Jin, J.-O. Laminarin promotes anti-cancer immunity by the maturation of dendritic cells. Oncotarget 2017, 8, 38554–38567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Globus, M.Y.; Busto, R.; Martinez, E.; Valdés, I.; Dietrich, W.D.; Ginsberg, M.D. Comparative effect of transient global ischemia on extracellular levels of glutamate, glycine, and gamma-aminobutyric acid in vulnerable and nonvulnerable brain regions in the rat. J. Neurochem. 1991, 57, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Pulsinelli, W.A.; Brierley, J.B.; Plum, F. Temporal profile of neuronal damage in a model of transient forebrain ischemia. Ann. Neurol. 1982, 11, 491–498. [Google Scholar] [CrossRef]
- Petito, C.K.; Feldmann, E.; Pulsinelli, W.A.; Plum, F. Delayed hippocampal damage in humans following cardiorespiratory arrest. Neurology 1987, 37, 1281. [Google Scholar] [CrossRef]
- Gemma, C.; Vila, J.; Bachstetter, A.; Bickford, P.C. Oxidative Stress and the Aging Brain: From Theory to Prevention; Riddle, D.R., Ed.; Brain Aging: Boca Raton, FL, USA, 2007. [Google Scholar]
- Sparkman, N.L.; Johnson, R. Neuroinflammation associated with aging sensitizes the brain to the effects of infection or stress. Neuroimmunomodulation 2008, 15, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Olmez, I.; Ozyurt, H. Reactive oxygen species and ischemic cerebrovascular disease. Neurochem. Int. 2012, 60, 208–212. [Google Scholar] [CrossRef]
- Godinho, J.; De Sá-Nakanishi, A.B.; Moreira, L.S.; De Oliveira, R.M.W.; Huzita, C.H.; Mello, J.C.P.; Da Silva, A.O.F.; Nakamura, C.V.; Previdelli, I.S.; Ribeiro, M.H.D.M.; et al. Ethyl-acetate fraction of Trichilia catigua protects against oxidative stress and neuroinflammation after cerebral ischemia/reperfusion. J. Ethnopharmacol. 2018, 221, 109–118. [Google Scholar] [CrossRef]
- Viswanatha, G.L.; Shylaja, H.; Mohan, C. Alleviation of transient global ischemia/reperfusion-induced brain injury in rats with 1,2,3,4,6-penta-O-galloyl-β-d-glucopyranose isolated from Mangifera indica. Eur. J. Pharmacol. 2013, 720, 286–293. [Google Scholar] [CrossRef]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef]
- Davis, S.; Pennypacker, K.R. Targeting antioxidant enzyme expression as a therapeutic strategy for ischemic stroke. Neurochem. Int. 2016, 107, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.-S.; Park, E.J.; Sohn, S.; Kwon, H.J.; Shin, W.-H.; Pyo, H.K.; Jin, B.; Choi, K.; Jou, I.; Joe, E.-H. Interleukin-13 and -4 induce death of activated microglia. Glia 2002, 38, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Troup, G.; Smith, G.; Walford, R. Life span, chronologic disease patterns, and age-related changes in relative spleen weights for the mongolian gerbil (Meriones unguiculatus). Exp. Gerontol. 1969, 4, 139–143. [Google Scholar] [CrossRef]
- Park, J.H.; Ahn, J.H.; Song, M.; Kim, H.; Park, C.; Park, Y.; Lee, T.-K.; Lee, J.-C.; Kim, D.W.; Lee, C.-H.; et al. A 2-Min Transient Ischemia Confers Cerebral Ischemic Tolerance in Non-Obese Gerbils, but Results in Neuronal Death in Obese Gerbils by Increasing Abnormal mTOR Activation-Mediated Oxidative Stress and Neuroinflammation. Cells 2019, 8, 1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, M.; Ahn, J.H.; Kim, H.; Kim, D.W.; Lee, T.-K.; Lee, J.-C.; Kim, Y.-M.; Lee, C.-H.; Hwang, I.K.; Yan, B.C.; et al. Chronic high-fat diet-induced obesity in gerbils increases pro-inflammatory cytokines and mTOR activation, and elicits neuronal death in the striatum following brief transient ischemia. Neurochem. Int. 2018, 121, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Park, J.H.; Kim, I.H.; Cho, G.S.; Ahn, J.H.; Tae, H.J.; Choi, S.Y.; Cho, J.H.; Kim, D.W.; Kwon, Y.G.; et al. Neuroprotection of ischemic preconditioning is mediated by thioredoxin 2 in the hippocampal CA1 region following a subsequent transient cerebral ischemia. Brain Pathol. 2017, 27, 276–291. [Google Scholar] [CrossRef] [PubMed]
- Radtke-Schuller, S.; Schuller, G.; Angenstein, F.; Grosser, O.S.; Goldschmidt, J.; Budinger, E. Brain atlas of the Mongolian gerbil (Meriones unguiculatus) in CT/MRI-aided stereotaxic coordinates. Brain Struct. Funct. 2016, 221, 1–272. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.H.; Ahn, J.H.; Lee, T.-K.; Park, C.W.; Kim, B.; Lee, J.-C.; Kim, D.W.; Shin, M.C.; Cho, J.H.; Lee, C.-H.; et al. Laminarin Pretreatment Provides Neuroprotection against Forebrain Ischemia/Reperfusion Injury by Reducing Oxidative Stress and Neuroinflammation in Aged Gerbils. Mar. Drugs 2020, 18, 213. https://doi.org/10.3390/md18040213
Park JH, Ahn JH, Lee T-K, Park CW, Kim B, Lee J-C, Kim DW, Shin MC, Cho JH, Lee C-H, et al. Laminarin Pretreatment Provides Neuroprotection against Forebrain Ischemia/Reperfusion Injury by Reducing Oxidative Stress and Neuroinflammation in Aged Gerbils. Marine Drugs. 2020; 18(4):213. https://doi.org/10.3390/md18040213
Chicago/Turabian StylePark, Joon Ha, Ji Hyeon Ahn, Tae-Kyeong Lee, Cheol Woo Park, Bora Kim, Jae-Chul Lee, Dae Won Kim, Myoung Cheol Shin, Jun Hwi Cho, Choong-Hyun Lee, and et al. 2020. "Laminarin Pretreatment Provides Neuroprotection against Forebrain Ischemia/Reperfusion Injury by Reducing Oxidative Stress and Neuroinflammation in Aged Gerbils" Marine Drugs 18, no. 4: 213. https://doi.org/10.3390/md18040213