The Microbial Community of Tetrodotoxin-Bearing and Non-Tetrodotoxin-Bearing Ribbon Worms (Nemertea) from the Sea of Japan


Abstract
:1. Introduction
2. Results
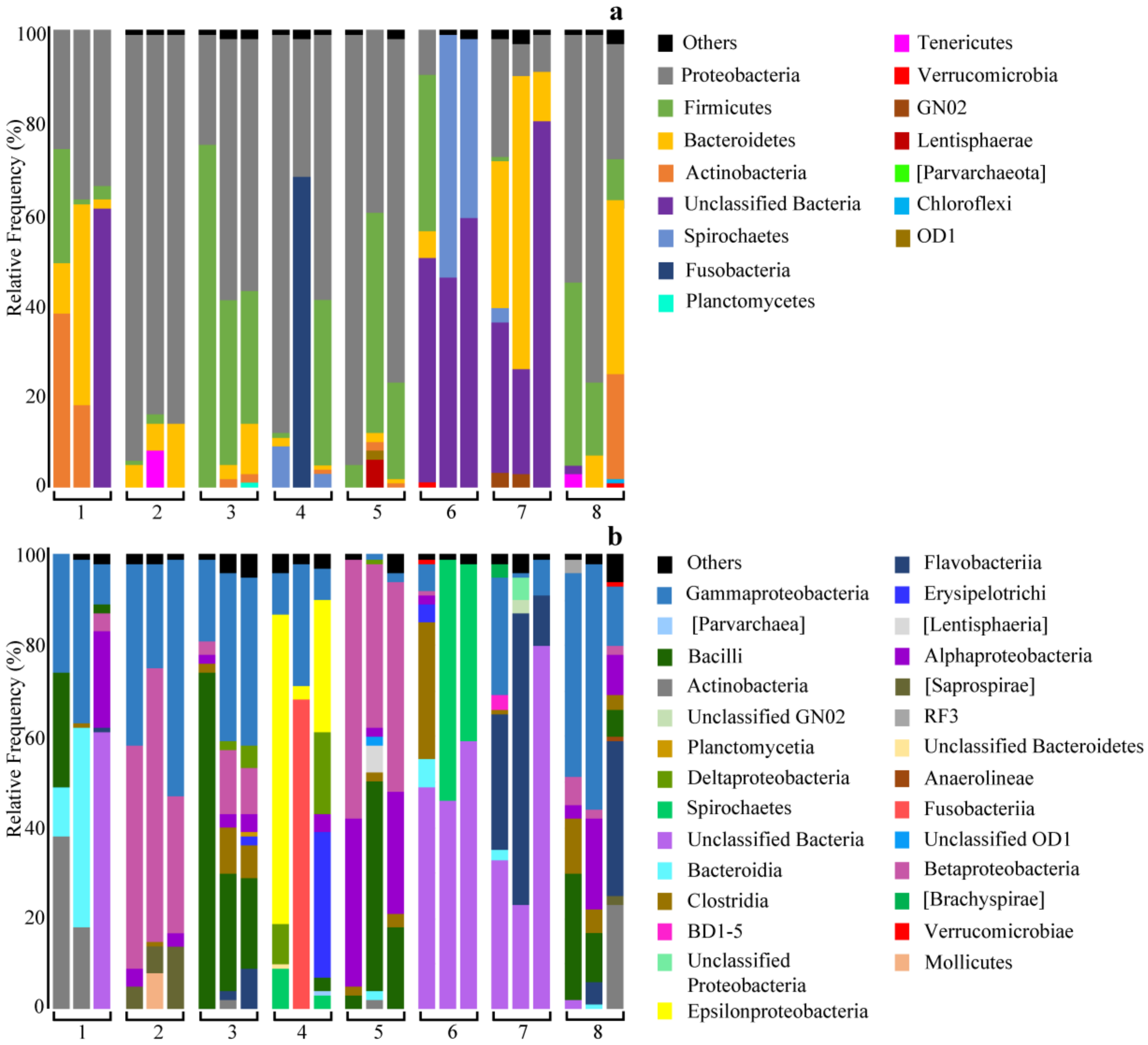
2.1. Bacterial Taxonomic Distribution in Nemertean Species
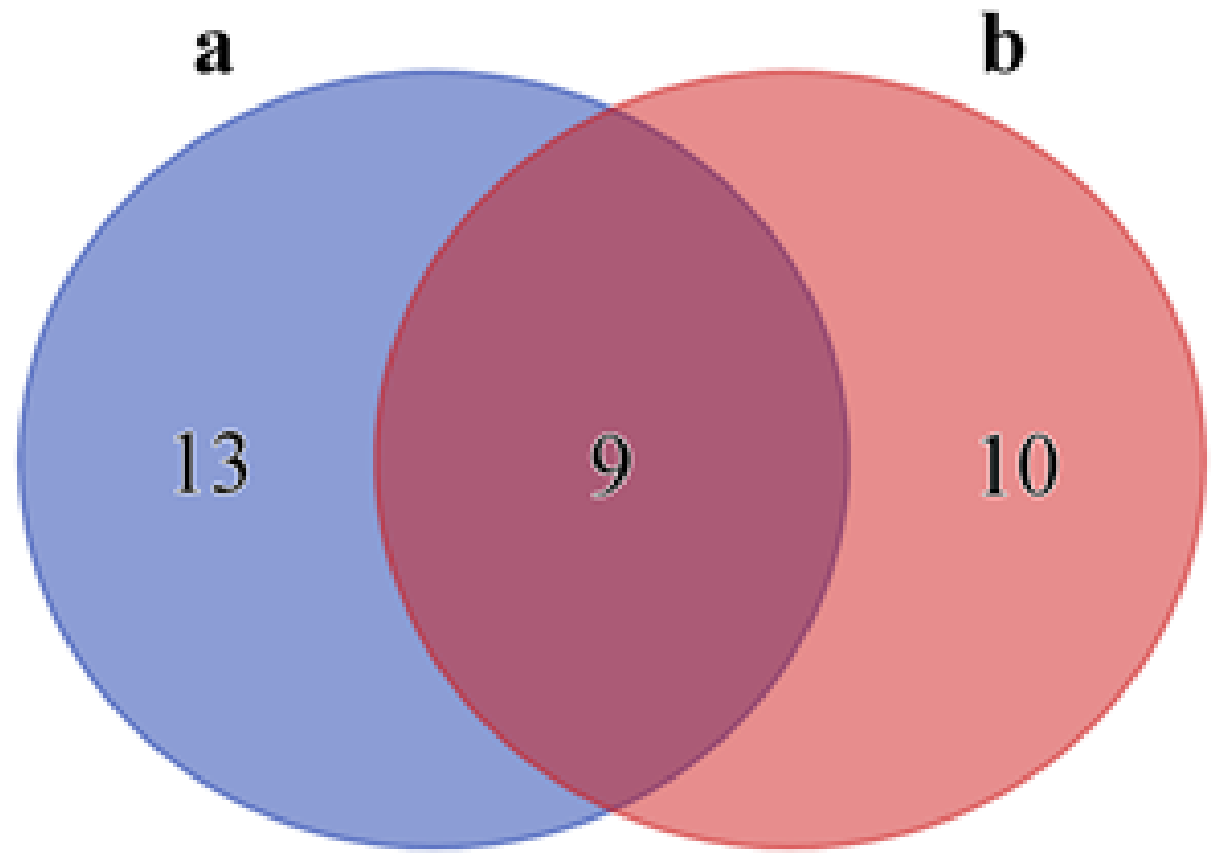
2.2. Core Microbiome of the Nemertean Species Studied
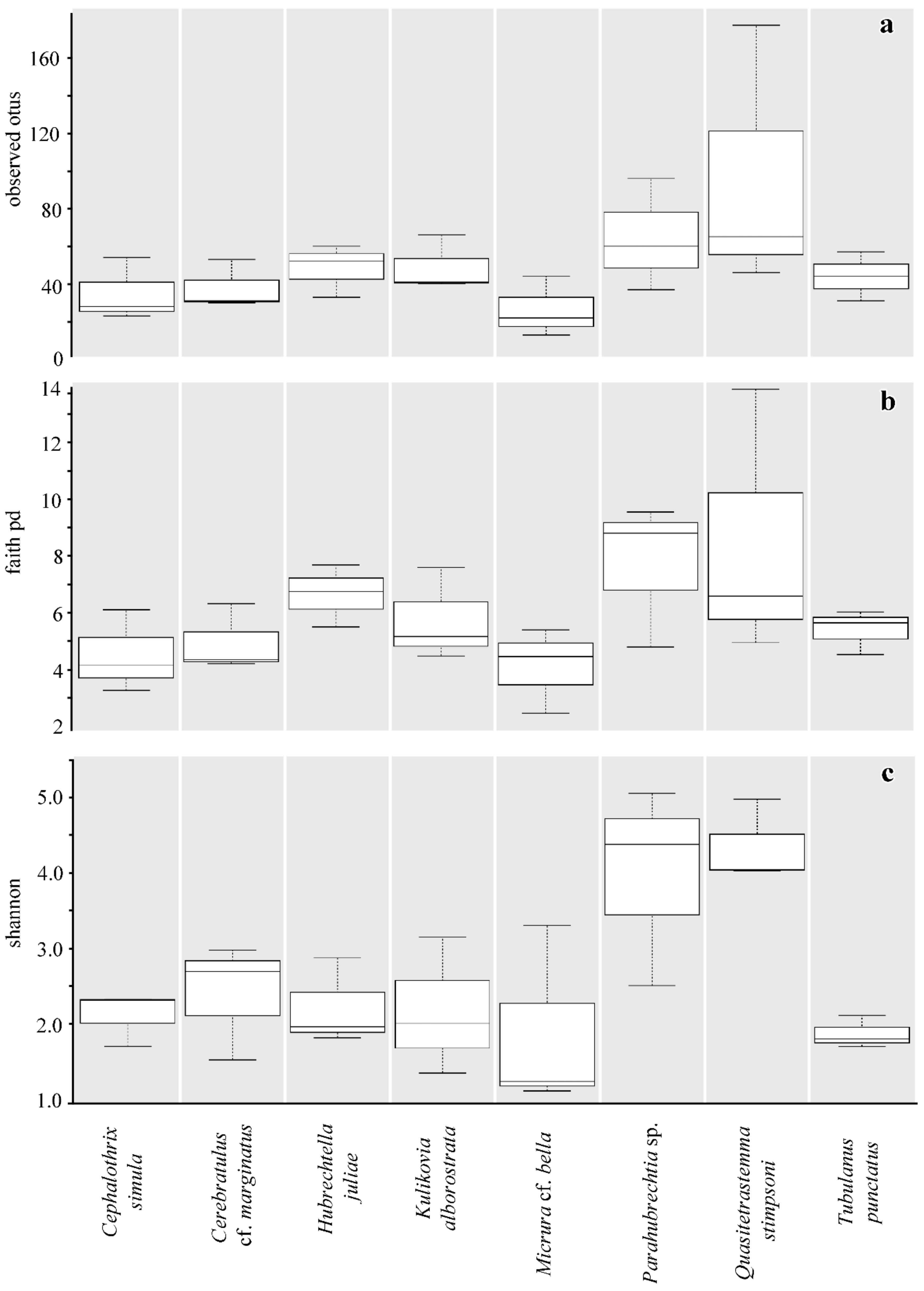
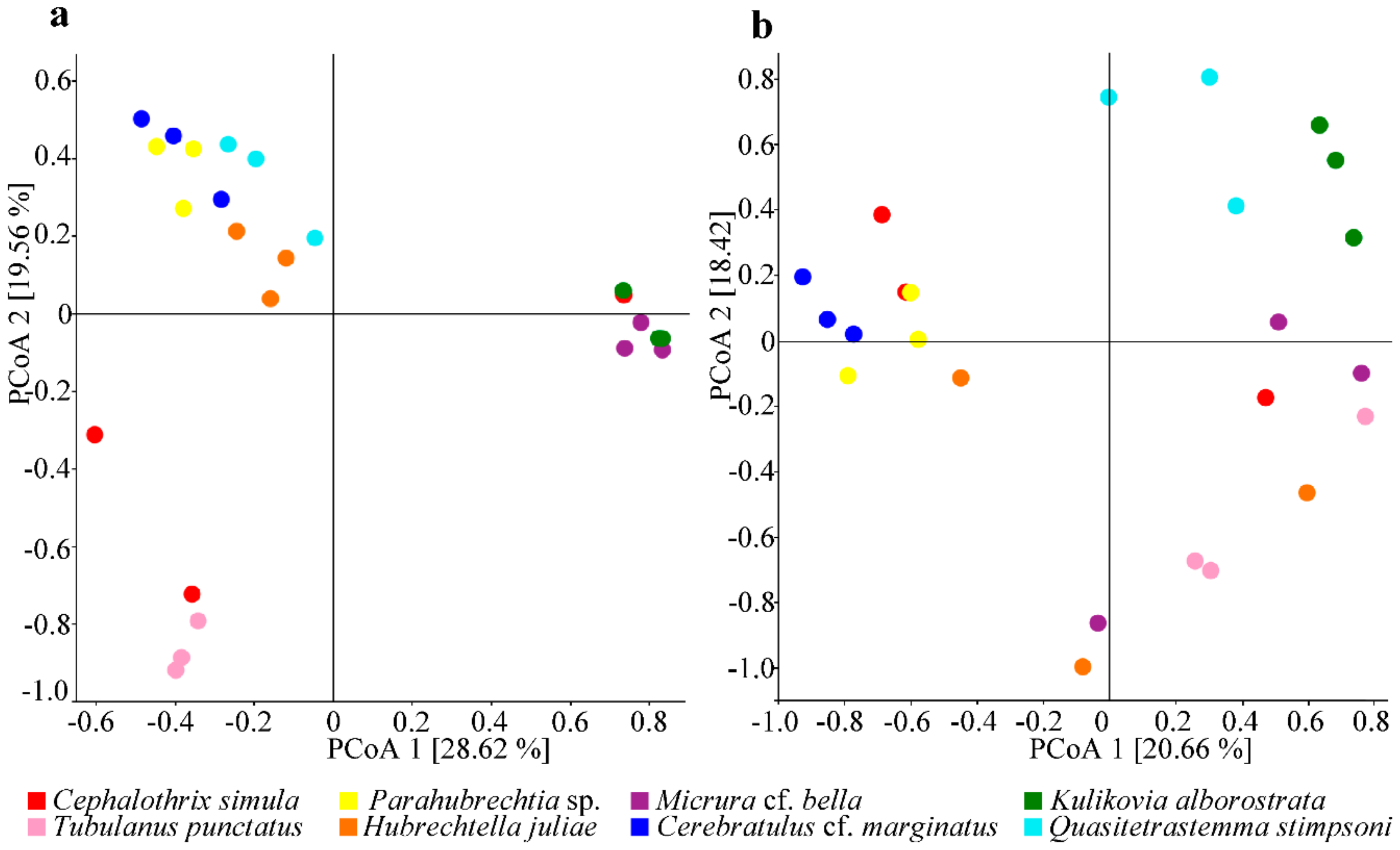
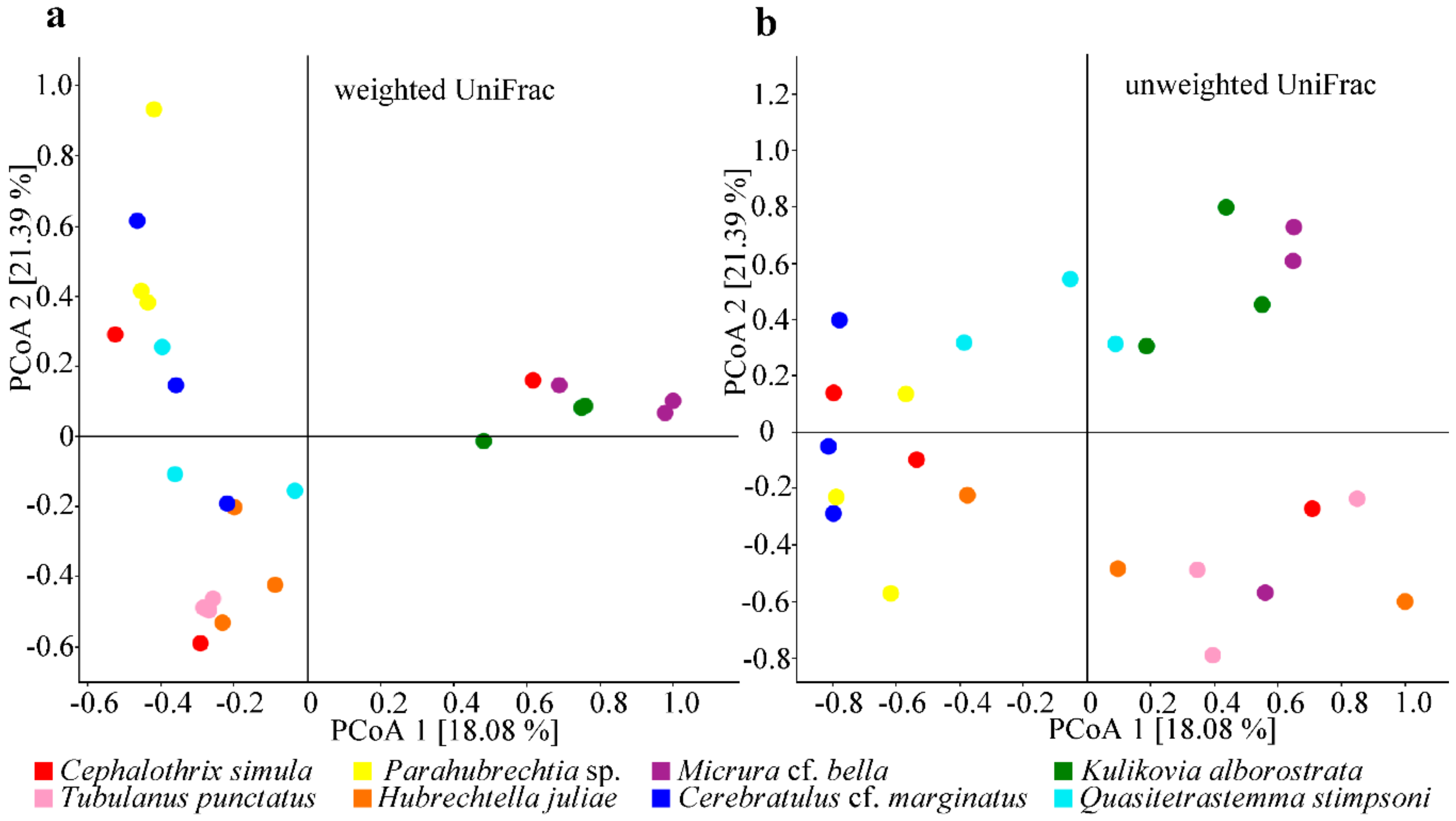
2.3. Community Diversity of the Nemertean Microbiomes Studied
2.4. TTXs Analysis in Parahubrechtia sp. and Hubrechtella juliae
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. DNA Extraction and 16S rRNA Sequence Processing
4.3. Sequence Processing and Taxonomic Assignment
4.4. Statistical Analysis
4.5. Toxin Testing in Parahubrechtia sp. and Hubrechtella juliae
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Magarlamov, T.Y.; Melnikova, D.I.; Chernyshev, A.V. Tetrodotoxin-producing bacteria: Detection, distribution and migration of the toxin in aquatic systems. Toxins (Basel) 2017, 9, 166. [Google Scholar] [CrossRef] [PubMed]
- Lago, J.; Rodriguez, L.P.; Blanco, L.; Vieites, J.M.; Cabado, A.G. Tetrodotoxin, an extremely potent marine neurotoxin: Distribution, toxicity, origin and therapeutical uses. Mar. Drugs 2015, 13, 6384–6406. [Google Scholar] [CrossRef] [PubMed]
- Vlamis, A.; Katikou, P.; Rodríguez, I.; Rey, V.; Alfonso, A.; Papazachariou, A.; Zacharaki, T.; Botana, A.M.; Botana, L.M. First detection of tetrodotoxin in greek shellfish by UPLC-MS/MS potentially linked to the presence of the dinoflagellate Prorocentrum minimum. Toxins (Basel) 2015, 7, 1779–1807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, I.; Alfonso, A.; Alonso, E.; Rubiolo, J.A.; Roel, M.; Vlamis, A.; Katikou, P.; Jackson, S.A.; Menon, M.L.; Dobson, A.D.W.; et al. The association of bacterial C 9 -based TTX-like compounds with Prorocentrum minimum opens new uncertainties about shellfish seafood safety. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kajihara, H.; Chernyshev, A.V.; Sun, S.-C.; Sundberg, P.; Crandall, F.B. Checklist of nemertean genera and species published between 1995 and 2007. Species Divers 2008, 13, 245–274. [Google Scholar] [CrossRef] [Green Version]
- Miyazawa, K.; Higashiyama, M.; Ito, K.; Noguchi, T.; Arakawa, O.; Shida, Y.; Hashimoto, K. Tetrodotoxin in two species of ribbon worm (Nemertini), Lineus fuscoviridis and Tubulanus punctatus. Toxicon 1988, 26, 867–874. [Google Scholar] [CrossRef]
- Ali, A.E.; Arakawa, O.; Noguchi, T.; Miyazawa, K.; Shida, Y.; Hashimoto, K. Tetrodotoxin and related substances in a ribbon worm Cephalothrix linearis (Nemertean). Toxicon 1990, 28, 1083–1093. [Google Scholar] [CrossRef]
- Noguchi, T.; Ali, A.E.; Arakawa, O.; Miyazawa, K.; Kanoh, S.; Shida, Y.; Nishio, S.; Hashimoto, K. Tetrodonic acid-like substance; a possible precursor of tetrodotoxin. Toxicon 1991, 29, 845–855. [Google Scholar] [CrossRef]
- Asakawa, M.; Toyoshima, T.; Shida, Y.; Noguchi, T.; Miyazawa, K. Paralytic toxins in a ribbon worm Cephalothrix species (Nemertean) adherent to cultured oysters in Hiroshima Bay, Hiroshima Prefecture, Japan. Toxicon 2000, 38, 763–773. [Google Scholar] [CrossRef]
- Asakawa, M.; Toyoshima, T.; Ito, K.; Bessho, K.; Yamaguchi, C.; Tsunetsugu, S.; Shida, Y.; Kajihara, H.; Mawatari, S.F.; Noguchi, T.; et al. Paralytic toxicity in the ribbon worm Cephalothrix species (Nemertea) in Hiroshima Bay, Hiroshima Prefecture, Japan and the isolation of tetrodotoxin as a main component of its toxins. Toxicon 2003, 41, 747–753. [Google Scholar] [CrossRef]
- Asakawa, M.; Ito, K.; Kajihara, H. Highly toxic ribbon worm Cephalothrix simula containing tetrodotoxin in Hiroshima Bay, Hiroshima Prefecture, Japan. Toxins (Basel) 2013, 5, 376–395. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.; Fenwick, D.; Powell, A.; Dhanji-Rapkova, M.; Ford, C.; Hatfield, R.G.; Santos, A.; Martinez-Urtaza, J.; Bean, T.; Baker-Austin, C.; et al. New invasive nemertean species (Cephalothrix simula) in england with high levels of tetrodotoxin and a microbiome linked to toxin metabolism. Mar. Drugs 2018, 16, 452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlasenko, A.E.; Velansky, P.V.; Chernyshev, A.V.; Kuznetsov, V.G.; Magarlamov, T.Y. Tetrodotoxin and its analogues profile in nemertean species from the sea of Japan. Toxicon 2018, 156, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.S.; Min, S.K.; Yeon, S.J.; Hwang, J.H.; Hong, J.S.; Shin, H.S. Assessment of neuronal cell-based cytotoxicity of neurotoxins from an estuarine nemertean in the han river estuary. J. Microbiol. Biotechnol. 2017, 27, 725–730. [Google Scholar] [CrossRef] [Green Version]
- Yotsu-Yamashita, M.; Abe, Y.; Kudo, Y.; Ritson-Williams, R.; Paul, V.; Konoki, K.; Cho, Y.; Adachi, M.; Imazu, T.; Nishikawa, T.; et al. First identification of 5,11-dideoxytetrodotoxin in marine animals, and characterization of major fragment ions of tetrodotoxin and its analogs by high resolution ESI-MS/MS. Mar. Drugs 2013, 11, 2799–2813. [Google Scholar] [CrossRef] [Green Version]
- Kono, M.; Matsui, T.; Furukawa, K.; Takase, T.; Yamamori, K.; Kaneda, H.; Aoki, D.; Jang, J.-H.; Yotsu-Yamashita, M. Examination of transformation among tetrodotoxin and its analogs in the living cultured juvenile puffer fish, kusafugu, Fugu niphobles by intramuscular administration. Toxicon 2008, 52, 714–720. [Google Scholar] [CrossRef]
- Ueyama, N.; Sugimoto, K.; Kudo, Y.; Onodera, K.-I.; Cho, Y.; Konoki, K.; Nishikawa, T.; Yotsu-Yamashita, M. Spiro bicyclic guanidino compounds from pufferfish: Possible biosynthetic intermediates of tetrodotoxin in marine environments. Chem. A Eur. J. 2018, 24, 7250–7258. [Google Scholar] [CrossRef]
- Carroll, S.; McEvoy, E.G.; Gibson, R. The production of tetrodotoxin-like substances by nemertean worms in conjunction with bacteria. J. Exp. Mar. Bio. Ecol. 2003, 288, 51–63. [Google Scholar] [CrossRef]
- Magarlamov, T.Y.; Beleneva, I.A.; Chernyshev, A.V.; Kuhlevsky, A.D. Tetrodotoxin-producing Bacillus sp. from the ribbon worm (Nemertea) Cephalothrix simula (Iwata, 1952). Toxicon 2014, 85, 46–51. [Google Scholar] [CrossRef]
- Melnikova, D.I.; Beleneva, I.A.; Tyunin, A.P.; Magarlamov, T.Y. The taxonomic composition, characteristics, and neurotoxic activities of ribbon worm-associated bacteria from the Sea of Japan. Russ. J. Mar. Biol. 2017, 43, 383–391. [Google Scholar] [CrossRef]
- Melnikova, D.I.; Vlasenko, A.E.; Magarlamov, T.Y. Stable tetrodotoxin production by Bacillus sp. strain 1839. Mar. Drugs 2019, 17, 704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beleneva, I.A.; Magarlamov, T.Y.; Kuhlevsky, A.D. Characterization, identification, and screening for tetrodotoxin production by bacteria associated with the ribbon worm (Nemertea) Cephalotrix simula (Ivata, 1952). Microbiology (Russian Fed) 2014, 83, 312–319. [Google Scholar] [CrossRef]
- Rizzo, C.; Lo Giudice, A. Marine invertebrates: Underexplored sources of bacteria producing biologically active molecules. Diversity 2018, 10, 52. [Google Scholar] [CrossRef] [Green Version]
- MCDermott, J.J.; Roe, P. Food, feeding behavior and reeding ecology of Nemerteans. Am. Zool. 1985, 25, 113–125. [Google Scholar] [CrossRef]
- Chernyshev, A.V. Nemertean biodiversity in the Sea of Japan and adjacent areas. In Marine Biodiversity and Ecosystem Dynamics of the Northwest Pacific Ocean; Song, S., Adrianov, A.V., Lutaenko, K.A., Xiao-Xia, S., Eds.; Publishing House of Science: Beijing, China, 2014; pp. 119–135. [Google Scholar]
- Frans, I.; Michiels, C.W.; Bossier, P.; Willems, K.A.; Lievens, B.; Rediers, H. Vibrio anguillarum as a fish pathogen: Virulence factors, diagnosis and prevention. J. Fish Dis. 2011, 34, 643–661. [Google Scholar] [CrossRef]
- Kozińska, A.; Paździor, E.; Pȩkala, A.; Niemczuk, W. Acinetobacter johnsonii and Acinetobacter iwoffii—The emerging fish pathogens. Bull. Vet. Inst. Pulawy. 2014, 58, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Moran, N.A.; Sloan, D.B. The hologenome concept: Helpful or hollow? PLoS Biol. 2015, 13, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Montalvo, N.F.; Hill, R.T. Sponge-associated bacteria are strictly maintained in two closely related but geographically distant sponge hosts. Appl. Environ. Microbiol. 2011, 77, 7207–7216. [Google Scholar] [CrossRef] [Green Version]
- Schöttner, S.; Hoffmann, F.; Cárdenas, P.; Rapp, H.T.; Boetius, A.; Ramette, A. Relationships between host phylogeny, host type and bacterial community diversity in cold-water coral reef sponges. PLoS ONE 2013, 8, 1–11. [Google Scholar] [CrossRef]
- Souza, D.T.; Genuario, D.B.; Silva, F.S.P.; Pansa, C.; Kavamura, V.N.; Moraes, F.C.; Taketani, R.G.; Melo, I.S.; Olson, J. Analysis of bacterial composition in marine sponges reveals the influence of host phylogeny and environment. FEMS Microbiol. Ecol. 2017, 93, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Strand, M.; Hedström, M.; Seth, H.; McEvoy, E.G.; Jacobsson, E.; Göransson, U.; Andersson, H.S.; Sundberg, P. The bacterial (Vibrio alginolyticus) production of tetrodotoxin in the ribbon worm Lineus longissimus—just a false positive? Mar. Drugs 2016, 14, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simidu, U.; Noguchi, T.; Hwang, D.F.; Shida, Y.; Hashimoto, K. Marine bacteria which produce tetrodotoxin. Appl. Environ. Microbiol. 1987, 53, 1714–1715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 2016, 1–22. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [Green Version]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Caporaso, J.G. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 1–17. [Google Scholar] [CrossRef]
- Chen, X.W.; Liu, H.X.; Jin, Y.B.; Li, S.F.; Bi, X.; Chung, S.; Zhang, S.S.; Jiang, Y.Y. Separation, identification and quantification of tetrodotoxin and its analogs by LC-MS without calibration of individual analogs. Toxicon 2011, 57, 938–943. [Google Scholar] [CrossRef]
- Bane, V.; Lehane, M.; Dikshit, M.; O’Riordan, A.; Furey, A. Tetrodotoxin: Chemistry, toxicity, source, distribution and detection. Toxins 2014, 6, 693–755. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nemertean Species | TTXs Presence | Reference |
|---|---|---|
| Palaeonemertea | ||
| Cephalothrix simula (Iwata, 1952) | + | [13] |
| Tubulanus punctatus Takakura, 1898 | + | [13] |
| Parahubrechtia sp. | − | Present study |
| Pilidiophora | ||
| Hubrechtella juliae (Chernyshev, 2003) | − | Present study |
| Cerebratulus cf. marginatus Renier, 1804 | − | [13] |
| Micrura cf. bella (Stimpson, 1857) | − | [13] |
| Kulikovia alborostrata (Takakura, 1898) | + | [13] |
| Hoplonemertea | ||
| Quasitetrastemma stimpsoni (Chernyshev, 1992) | + | [13] |
| Phylum | Class | Order | % of All Sequences | Found in Nemertean Species * |
|---|---|---|---|---|
| Proteobacteria | Alphaproteobacteria | Rickettsiales | 1.8 | 1,2,3,5,8 |
| Betaproteobacteria | Burkholderiales | 2.3 | All | |
| Methylophilales | 5.7 | 1,2,3,4,5,8 | ||
| Deltaproteobacteria | Sva0853 | 1.3 | 4 | |
| Gammaproteobacteria | Alteromonadales | 1.3 | All | |
| Oceanospirillales | 3.3 | All | ||
| Pseudomonadales | 2.4 | All | ||
| Vibrionales | 6 | All | ||
| Epsilonproteobacteria | Campylobacterales | 4.9 | 2,3,4,5,6,7,8 | |
| Firmicutes | Bacilli | Lactobacillales | 11.9 | All |
| Clostridia | Clostridiales | 2.8 | All | |
| Erysipelotrichi | Erysipelotrichales | 1.8 | 1,2,3,4,6,8 | |
| Bacteroidetes | Bacteroidia | Bacteroidales | 2.7 | All |
| Flavobacteria | Flavobacteriales | 6.6 | 1,2,3,4,5,7,8 | |
| Saprospirae | Saprospirales | 1 | All | |
| Actinobacteria | Actinobacteria | Actinomycetales | 3 | All |
| Fusobacteria | Fusobacteria | Fusobacteriales | 3 | 2,4,8 |
| OTU ID | Taxonomy According to Greengenes | % of Nemertean Samples | Found in Nemertean Species * |
|---|---|---|---|
| OTU_NEM_285 | Kingdom Bacteria | 92 | All |
| OTU_NEM_132 | Kingdom Bacteria; phylum OD1 | 87.5 | All |
| OTU_NEM_306 | Kingdom Bacteria; phylum Actinobacteria; class Actinobacteria; order Actinomycetales; family Corynebacteriaceae; Corynebacterium | 58 | 1,2,3,4,5,6,8 |
| OTU_NEM_332 | Kingdom Bacteria; phylum Proteobacteria; class Alphaproteobacteria; order Rhizobiales; family Bradyrhizobiaceae | 62.5 | All |
| OTU_NEM_179 | Kingdom Bacteria; phylum Proteobacteria; class Betaproteobacteria; order Burkholderiales; family Oxalobacteraceae; Cupriavidus | 100 | All |
| OTU_NEM_358 | Kingdom Bacteria; phylum Proteobacteria; class Betaproteobacteria; order Burkholderiales; family Comamonadaceae | 92 | All |
| OTU_NEM_454 | Kingdom Bacteria; phylum Proteobacteria; class Gammaproteobacteria; order Pseudomonadales; family Moraxellaceae; Acinetobacter | 71 | 1,2,3,4,5,7,8 |
| OTU_NEM_327 | Kingdom Bacteria; phylum Proteobacteria; class Gammaproteobacteria; order Pseudomonadales; family Moraxellaceae; Acinetobacter johnsonii | 62.5 | 1,2,3,4,6,7,8 |
| OTU_NEM_396 | Kingdom Bacteria; phylum Proteobacteria; class Gammaproteobacteria; order Vibrionales; family Vibrionaceae | 62.5 | 1,2,4,6,7,8 |
| OTU_NEM_95 | Kingdom Bacteria; phylum Proteobacteria; class Gammaproteobacteria; order Enterobacteriales; family Enterobacteriaceae | 75 | All |
| OTU_NEM_126 | Kingdom Bacteria; phylum Firmicutes; class Clostridia; order Clostridiales; family Lachnospiraceae | 66.7 | All |
| OTU_NEM_315 | Kingdom Bacteria; phylum Firmicutes; class Clostridia; order Clostridiales; family Lachnospiraceae; [Ruminococcus] | 54 | 1,2,4,6,7,8 |
| OTU_NEM_231 | Kingdom Bacteria; phylum Firmicutes; class Bacilli; order Lactobacillales; family Lactobacillaceae; Lactobacillus | 58 | 1,2,3,4,5,7,8 |
| OTU ID | Taxonomy According to Greengenes | % of Nemertean Samples | Found in Nemertean Species * |
|---|---|---|---|
| OTU_NEM_34 | Kingdom Bacteria; phylum Actinobacteria; class Actinobacteria; order Actinomycetales; family Propionibacteriaceae; Propionibacterium acnes | 58 | All |
| OTU_NEM_263 | Kingdom Bacteria; phylum Bacteroidetes; class Flavobacteria; order Flavobacteriales; family Flavobacteriaceae | 58 | All |
| OTU_NEM_288 | Kingdom Bacteria; phylum Proteobacteria; class Gammaproteobacteria | 67 | 1,2,4 |
| OTU_NEM_102 | Kingdom Bacteria; phylum Proteobacteria; class Gammaproteobacteria; order Alteromonadales; family Alteromonadaceae; Alteromonas | 67 | All |
| OTU_NEM_158 | Kingdom Bacteria; phylum Proteobacteria; class Gammaproteobacteria; order Alteromonadales; family Colwelliaceae | 67 | 2,3,4 |
| OTU_NEM_446 | Kingdom Bacteria; phylum Proteobacteria; class Gammaproteobacteria; order Alteromonadales; family Moritellaceae; Moritella | 58 | 2,3,4 |
| OTU_NEM_19 | Kingdom Bacteria; phylum Proteobacteria; class Gammaproteobacteria; order Alteromonadales; family Psychromonadaceae; Psychromonas | 58 | 2,3,4 |
| OTU_NEM_327 | Kingdom Bacteria; phylum Proteobacteria; class Gammaproteobacteria; order Pseudomonadales; family Moraxellaceae; Acinetobacter johnsonii | 83 | All |
| OTU_NEM_114 | Kingdom Bacteria; phylum Proteobacteria; class Gammaproteobacteria; order Oceanospirillales; family Oceanospirillaceae; Marinomonas | 58 | 2,3,4 |
| OTU_NEM_2 | Kingdom Bacteria; phylum Proteobacteria; class Gammaproteobacteria; order Vibrionales; family Pseudoalteromonadaceae; Pseudoalteromonas | 67 | All |
| OTU_NEM_396 | Kingdom Bacteria; phylum Proteobacteria; class Gammaproteobacteria; order Vibrionales; family Vibrionaceae | 92 | All |
| OTU_NEM_45 | Kingdom Bacteria; phylum Proteobacteria; class Gammaproteobacteria; order Vibrionales; family Vibrionaceae; Listonella anguillarum | 67 | All |
| OTU_NEM_315 | Kingdom Bacteria; phylum Firmicutes; class Clostridia; order Clostridiales; family Lachnospiraceae, [Ruminococcus] | 67 | All |
| OTU ID | Taxonomy According to Greengenes | % of Nemertean Samples | Found in Nemertean Species * |
|---|---|---|---|
| OTU_NEM_306 | Kingdom Bacteria; phylum Actinobacteria; class Actinobacteria; order Actinomycetales; family Corynebacteriaceae; Corynebacterium | 67 | All |
| OTU_NEM_14 | Kingdom Bacteria; phylum Bacteroidetes; class Saprospirae; order Saprospirales; family Chitinophagaceae; Sediminibacterium | 58 | All |
| OTU_NEM_103 | Kingdom Bacteria; phylum Proteobacteria; class Alphaproteobacteria; order Rhizobiales; family Xanthobacteraceae | 58 | All |
| OTU_NEM_287 | Kingdom Bacteria; phylum Proteobacteria; class Epsilonproteobacteria; order Campylobacterales; family Campylobacteraceae; Arcobacter | 58 | All |
| OTU_NEM_98 | Kingdom Bacteria; phylum Proteobacteria; class Gammaproteobacteria; order Pseudomonadales; family Pseudomonadaceae; Pseudomonas nitroreducens | 58 | All |
| OTU_NEM_94 | Kingdom Bacteria; phylum Proteobacteria; class Gammaproteobacteria; order Oceanospirillales; family Endozoicimonaceae | 58 | 2,3,4 |
| OTU_NEM_156 | Kingdom Bacteria; phylum Firmicutes; class Clostridia; order Clostridiales | 58 | All |
| OTU_NEM_134 | Kingdom Bacteria; phylum Firmicutes; class Bacilli; order Lactobacillales | 58 | 2,3,4 |
| OTU_NEM_138 | Kingdom Bacteria; phylum Firmicutes; class Bacilli; order Lactobacillales; family Leuconostocaceae | 58 | 2,3,4 |
| OTU_NEM_208 | Kingdom Bacteria; phylum Firmicutes; class Bacilli; order Lactobacillales; family Leuconostocaceae; Leuconostoc | 58 | 2,3,4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melnikova, D.I.; Magarlamov, T.Y. The Microbial Community of Tetrodotoxin-Bearing and Non-Tetrodotoxin-Bearing Ribbon Worms (Nemertea) from the Sea of Japan. Mar. Drugs 2020, 18, 177. https://doi.org/10.3390/md18030177
Melnikova DI, Magarlamov TY. The Microbial Community of Tetrodotoxin-Bearing and Non-Tetrodotoxin-Bearing Ribbon Worms (Nemertea) from the Sea of Japan. Marine Drugs. 2020; 18(3):177. https://doi.org/10.3390/md18030177
Chicago/Turabian StyleMelnikova, Daria I., and Timur Yu. Magarlamov. 2020. "The Microbial Community of Tetrodotoxin-Bearing and Non-Tetrodotoxin-Bearing Ribbon Worms (Nemertea) from the Sea of Japan" Marine Drugs 18, no. 3: 177. https://doi.org/10.3390/md18030177