Improvement of Psoriasis by Alteration of the Gut Environment by Oral Administration of Fucoidan from Cladosiphon Okamuranus

 and
and
Abstract
:
1. Introduction
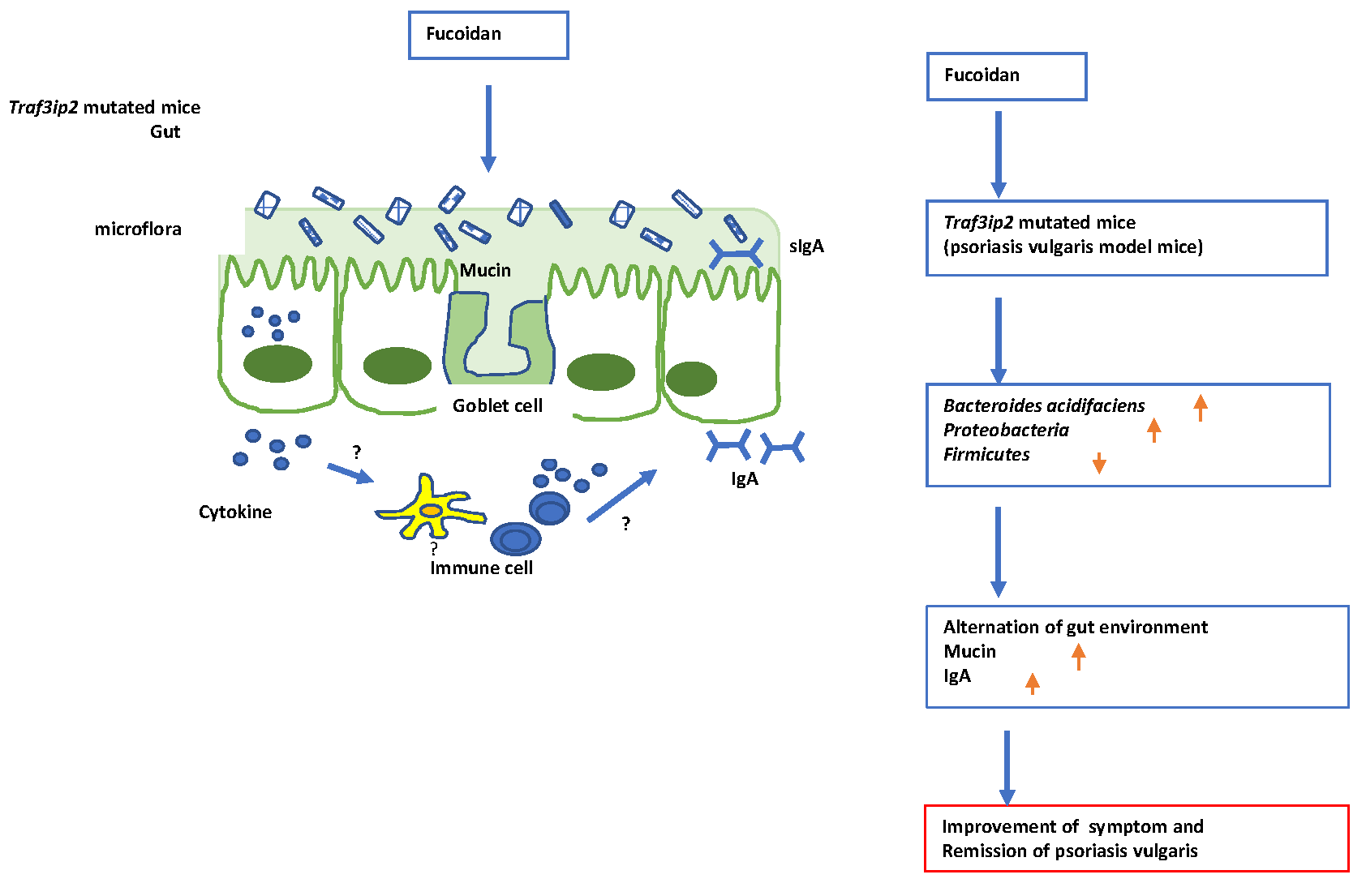
2. Results
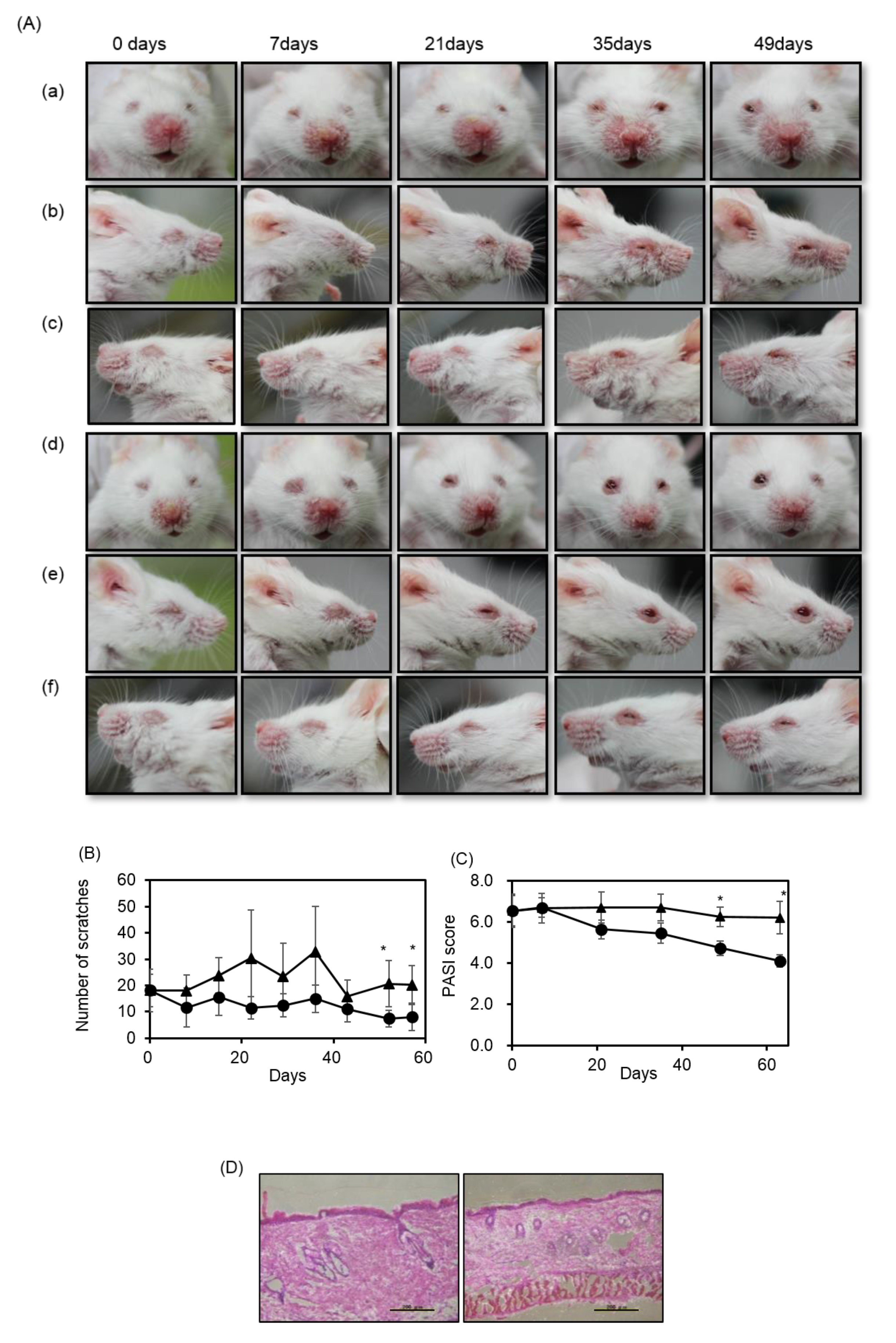
2.1. Fucoidan Diet Rescued Psoriasis Symptoms on m-Traf3ip2 Mouse Faces and Reduced Scratching
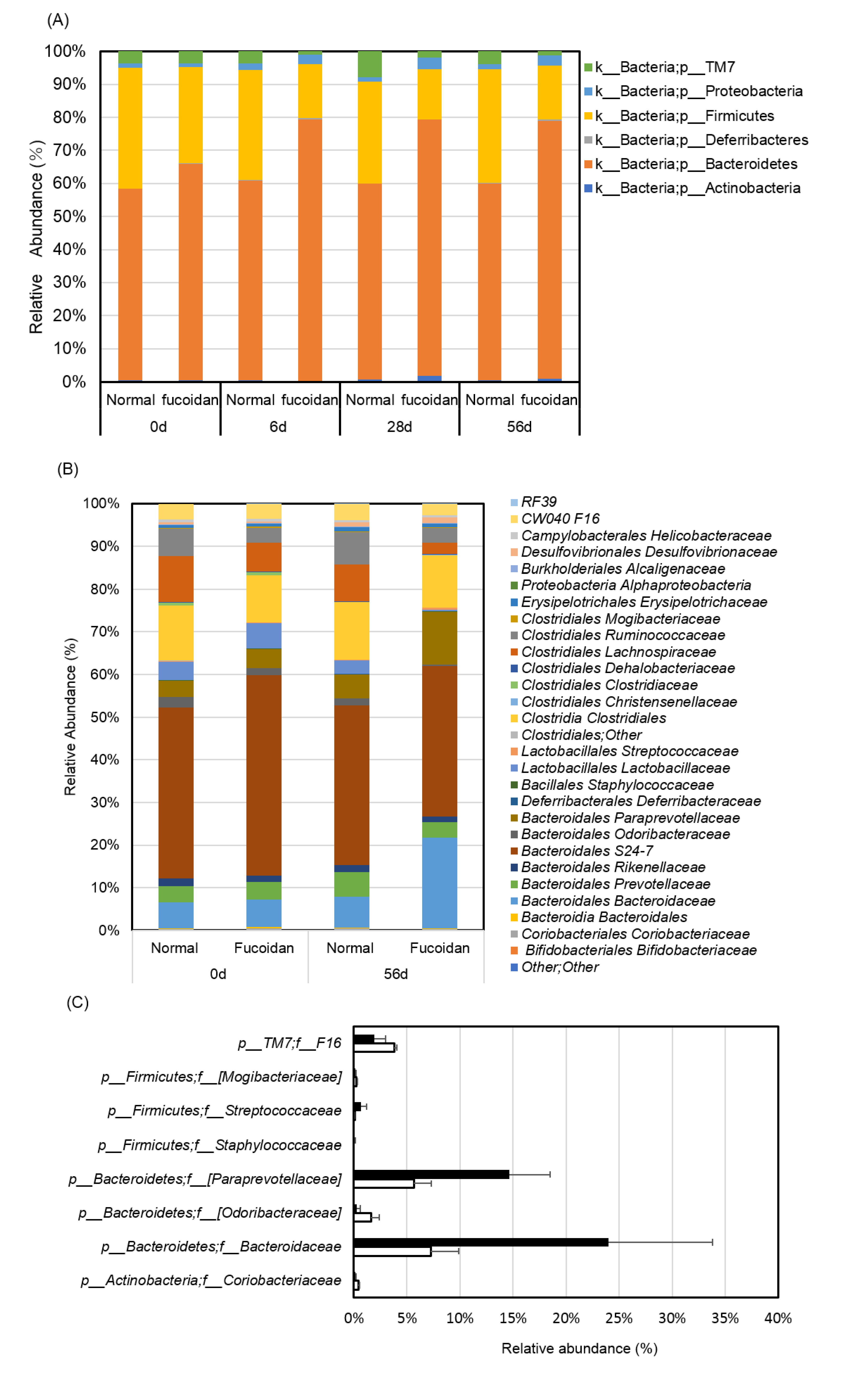
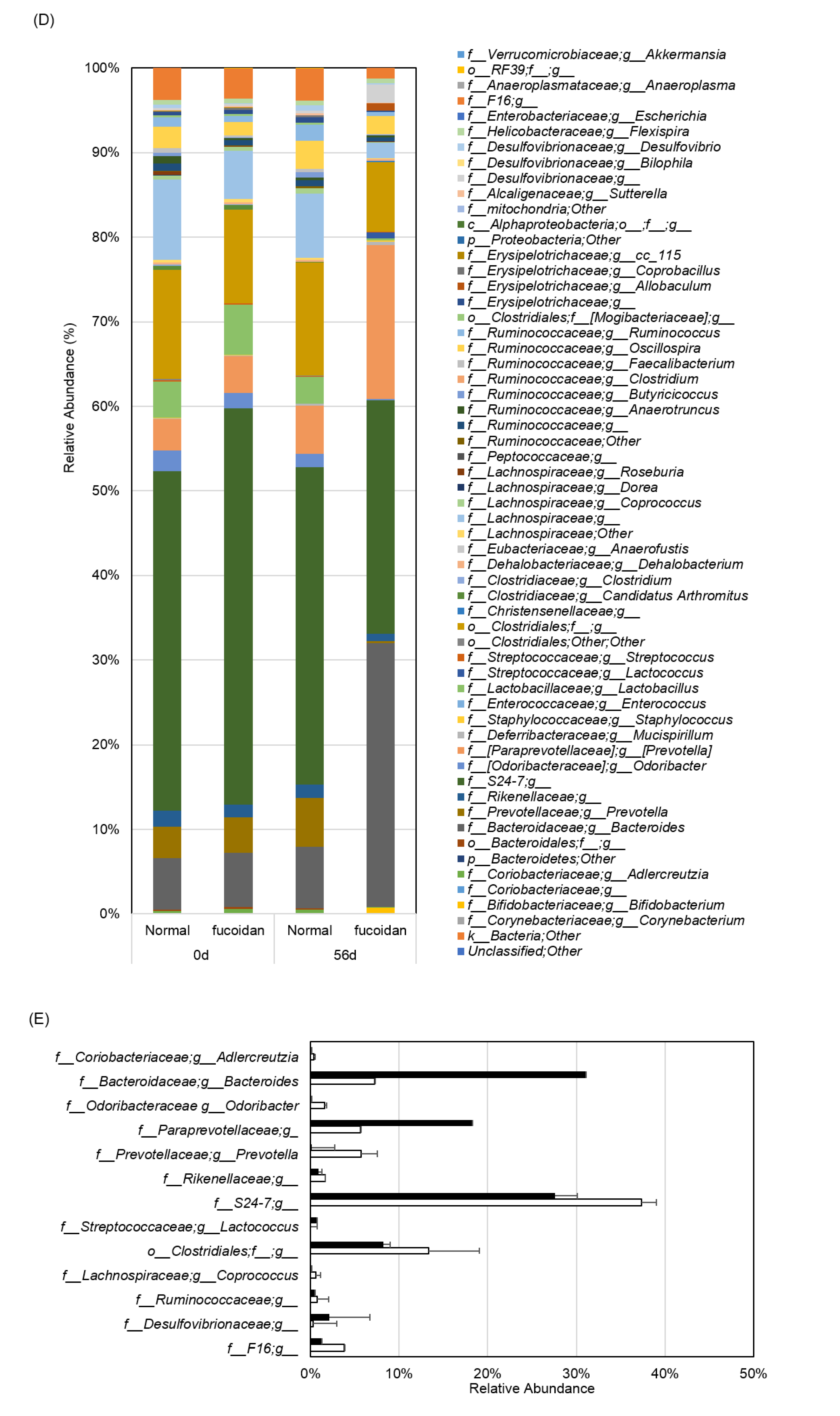
2.2. Fucoidan Drastically Changed Microbiota in the Small Intestines of m-Traf3ip2 Mice
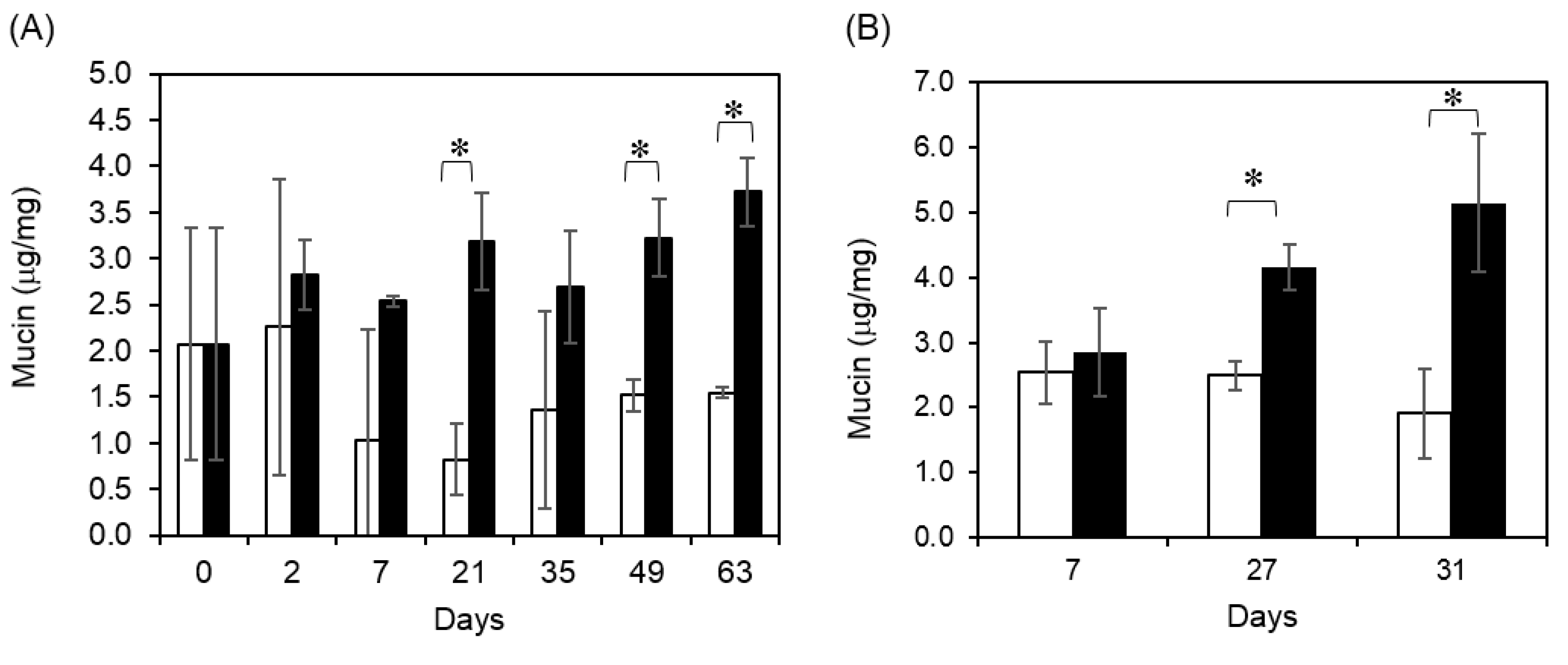
2.3. Fucoidan Diet Promoted Fecal Mucin Production
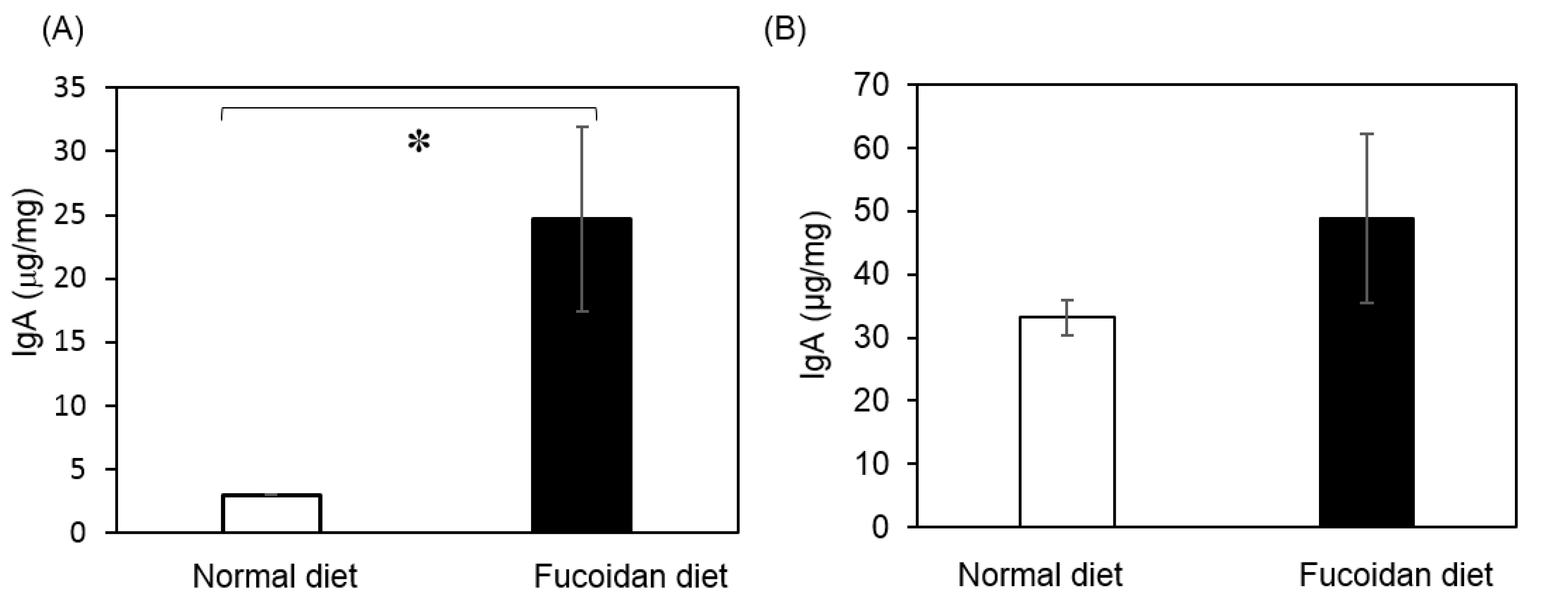
2.4. Production of IgA in Feces and Cecum
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Animals and Diets
4.3. Analysis of Phenotype and Scratch Test
4.4. Histochemical Analysis of Facial Skin Dissection
4.5. Microbiota Analysis in Feces
4.6. Quantification of Fecal Mucin and Cecum IgA
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Report on Psoriasis. Available online: https://apps.who.int/iris/bitstream/handle/10665/204417/9789241565189_eng.pdf.psoriasis;jsessionid=54912784D28C9F36ECCD45471AC5775B?sequence=1 (accessed on 10 March 2020).
- Genetic Analysis of Psoriasis Consortium & the Wellcome Trust Case Control Consortium; Strange, A.; Capon, F.; Spencer, C.C.A.; Knight, J.; Weale, M.E.; Allen, M.H.; Barton, A.; Band, G.; Bellenguez, C.; et al. A genome-wide association study identifies new psoriasis susceptibility loci and an interaction between HLA-C and ERAP1. Nat. Genet. 2010, 42, 985–990. [Google Scholar] [PubMed] [Green Version]
- Hayashi, M.; Hirota, T.; Saeki, H.; Nakagawa, H.; Ishiuji, Y.; Matsuzaki, H.; Tsunemi, Y.; Kato, T.; Shibata, S.; Sugaya, M.; et al. Genetic polymorphism in the TRAF3IP2 gene is associated with psoriasis vulgaris in a Japanese population. J. Dermatol. Sci. 2014, 73, 264–265. [Google Scholar] [CrossRef] [PubMed]
- Hüffmeier, U.; Uebe, S.; Ekici, A.B.; Bowes, J.; Giardina, E.; Korendowych, E.; Juneblad, K.; Apel, M.; McManus, R.; Ho, P.; et al. Common variants at TRAF3IP2 are associated with susceptibility to psoriatic arthritis and psoriasis. Nat. Genet. 2010, 42, 996–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsushima, Y.; Kikkawa, Y.; Takada, T.; Matsuoka, K.; Seki, Y.; Yonekawa, H.; Minegishi, Y.; Karsuyama, H.; Yonekawa, H. An atopic dermatitis-like skin disease with hyper-IgE-emia develops in mice carrying a spontaneous recessive point mutation in the Traf3ip2 (Act1/CIKS) gene. J. Immunol. 2010, 185, 2340–2349. [Google Scholar] [CrossRef]
- Scher, J.U.; Ubeda, C.; Artacho, A.; M, A.; Isaac, S.; Reddy, S.M.; Marmon, S.; Neimann, A.; Brusca, S.; Patel, T.; et al. Decreased bacterial diversity characterizes the altered gut microbiota in patients with psoriatic arthritis, resembling dysbiosis in inflammatory bowel disease. Arthritis Rheumatol. 2015, 67, 128–139. [Google Scholar] [CrossRef] [Green Version]
- Becattini, S.; Taur, Y.; Pamer, E.G. Antibiotic-Induced Changes in the Intestinal Microbiota and Disease. Trends Mol. Med. 2016, 22, 458–478. [Google Scholar] [CrossRef] [Green Version]
- Chiodini, R.J.; Dowd, S.E.; Chamberlin, W.M.; Galandiuk, S.; Davis, B.; Glassing, A. Microbial Population Differentials between Mucosal and Submucosal Intestinal Tissues in Advanced Crohn’s Disease of the Ileum. PLoS ONE 2015, 10, e0134382. [Google Scholar] [CrossRef]
- Ford, A.; Siegel, M.; Bagel, J.; Cordoro, K.; Garg, A.; Gottlieb, A.B.; Green, L.J.; Gudjonsson, J.E.; Koo, J.; Lebwohl, M.; et al. Dietary Recommendations for Adults With Psoriasis or Psoriatic Arthritis From the Medical Board of the National Psoriasis Foundation. JAMA Dermatol. 2018, 154, 934–950. [Google Scholar] [CrossRef]
- Szabó-Fodor, J.; Bónai, A.; Bóta, B.; Egyed, L.S.; Lakatos, F.; Pápai, G.; Zsolnai, A.; Glávits, R.; Horvatovich, K.; Kovács, M. Physiological Effects of Whey- and Milk-Based Probiotic Yogurt in Rats. Pol. J. Microbiol. 2017, 66, 483–490. [Google Scholar] [CrossRef] [Green Version]
- O’Shea, C.J.; O’Doherty, J.V.; Callanan, J.J.; Doyle, D.; Thornton, K.; Sweeney, T. The effect of algal polysaccharides laminarin and fucoidan on colonic pathology, cytokine gene expression and Enterobacteriaceae in a dextran sodium sulfate-challenged porcine model. J. Nutr. Sci. 2016, 5, 15. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, K.; Ichimura, M.; Tsuneyama, K.; Moritoki, Y.; Tsunashima, H.; Omagari, K.; Hara, M.; Yasuda, I.; Miyakawa, H.; Kikuchi, K. Fructo-oligosaccharides and intestinal barrier function in a methionine-choline-deficient mouse model of nonalcoholic steatohepatitis. PLoS ONE 2017, 12, e0175406. [Google Scholar] [CrossRef] [PubMed]
- Masahata, K.; Umemoto, E.; Kayama, H.; Kotani, M.; Takeda, K.; Kurakawa, T.; Kikuta, J.; Gotoh, K.; Motooka, D.; Sato, S.; et al. Generation of colonic IgA-secreting cells in the cecal patch. Nat. Commun. 2014, 5, 3704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’Incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I.; et al. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teruya, T.; Tatemoto, H.; Konishi, T.; Tako, M. Structural characteristics and in vitro macrophage activation of acetyl fucoidan from Cladosiphon okamuranus. Glycoconj. J. 2009, 26, 1019–1028. [Google Scholar] [CrossRef] [PubMed]
- Fukahori, S.; Yano, H.; Akiba, J.; Ogasawara, S.; Momosaki, S.; Sanada, S.; Kuratomi, K.; Ishizaki, Y.; Moriya, F.; Yagi, M.; et al. Fucoidan, a major component of brown seaweed, prohibits the growth of human cancer cell lines in vitro. Mol. Med. Rep. 2011, 1, 537–542. [Google Scholar] [CrossRef] [Green Version]
- Mori, N.; Nakasone, K.; Tomimori, K.; Ishikawa, C. Beneficial effects of fucoidan in patients with chronic hepatitis C virus infection. World J. Gastroenterol. 2012, 18, 2225–2230. [Google Scholar] [CrossRef]
- Yang, J.-H. Topical Application of Fucoidan Improves Atopic Dermatitis Symptoms in NC/Nga Mice. Phytother. Res. 2012, 26, 1898–1903. [Google Scholar] [CrossRef]
- Xue, M.; Ji, X.; Liang, H.; Liu, Y.; Wang, B.; Sun, L.; Li, W. The effect of fucoidan on intestinal flora and intestinal barrier function in rats with breast cancer. Food Funct. 2018, 9, 1214–1223. [Google Scholar] [CrossRef]
- Gill, S.R.; Pop, M.; DeBoy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.; Gordon, J.I.; Relman, D.A.; Fraser, C.M.; Nelson, K.E. Metagenomic Analysis of the Human Distal Gut Microbiome. Science 2006, 312, 1355–1359. [Google Scholar] [CrossRef] [Green Version]
- Satchithanandam, S.; Vargofcak-Apker, M.; Calvert, R.J.; Leeds, A.R.; Cassidy, M.M. Alteration of Gastrointestinal Mucin by Fiber Feeding in Rats. J. Nutr. 1990, 120, 1179–1184. [Google Scholar] [CrossRef]
- Shi, H.; Zheng, R.; Wu, J.; Zuo, T.; Xue, C.; Tang, Q.J. The Preventative Effect of Dietary Apostichopus japonicus on Intestinal Microflora Dysregulation in Immunosuppressive Mice Induced by Cyclophosphamide. J. Biosci. Med. 2016, 4, 24–35. [Google Scholar]
- Bäckhed, F. Host-Bacterial Mutualism in the Human Intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Shan, X.; Cai, C.; Hao, J.; Li, G.; Yu, G. Dietary fucoidan modulates the gut microbiota in mice by increasing the abundance ofLactobacillusandRuminococcaceae. Food Funct. 2016, 7, 3224–3232. [Google Scholar] [CrossRef]
- Guevarra, R.B.; Hong, S.H.; Cho, J.H.; Kim, B.-R.; Shin, J.; Lee, J.H.; Na Kang, B.; Kim, Y.H.; Wattanaphansak, S.; Isaacson, R.; et al. The dynamics of the piglet gut microbiome during the weaning transition in association with health and nutrition. J. Anim. Sci. Biotechnol. 2018, 9, 54. [Google Scholar] [CrossRef] [Green Version]
- Palm, N.W.; De Zoete, M.R.; Cullen, T.W.; Barry, N.A.; Stefanowski, J.; Hao, L.; Degnan, P.H.; Hu, J.; Peter, I.; Zhang, W.; et al. Immunoglobulin A coating identifies colitogenic bacteria in inflammatory bowel disease. Cell 2014, 158, 1000–1010. [Google Scholar] [CrossRef] [Green Version]
- Duan, M.; Sun, X.; Ma, N.; Liu, Y.; Luo, T.; Song, S.; Xu, Y. Polysaccharides from Laminaria japonica alleviated metabolic syndrome in BALB/c mice by normalizing the gut microbiota. Int. J. Boil. Macromol. 2019, 121, 996–1004. [Google Scholar] [CrossRef]
- Tanoue, T.; Honda, K. Regulation of intestinal Th17 and Treg cells by gut microbiota. Inflamm. Regen. 2015, 35, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Sawin, E.A.; De Wolfe, T.J.; Aktas, B.; Stroup, B.M.; Murali, S.G.; Steele, J.L.; Ney, D.M. Glycomacropeptide is a prebiotic that reduces Desulfovibrio bacteria, increases cecal short-chain fatty acids, and is anti-inflammatory in mice. Am. J. Physiol. Liver Physiol. 2015, 309, G590–G601. [Google Scholar] [CrossRef] [Green Version]
- Yanagibashi, T.; Hosono, A.; Oyama, A.; Tsuda, M.; Suzuki, A.; Hachimura, S.; Takahashi, Y.; Momose, Y.; Itoh, K.; Hirayama, K.; et al. IgA production in the large intestine is modulated by a different mechanism than in the small intestine: Bacteroides acidifaciens promotes IgA production in the large intestine by inducing germinal center formation and increasing the number of IgA+ B cells. Immunobiology 2013, 218, 645–651. [Google Scholar] [CrossRef]
- Graf, J. The Family Rikenellaceae. In The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2014; pp. 857–859. [Google Scholar]
- Nakano, V. Intestinal Bacteroides and Parabacteroides Species Producing Antagonistic Substances. Current Trends in Microbiology. Available online: http://www.icb.usp.br/~bmm/mariojac/arquivos/nakano%20et%20al.%202013.pdf (accessed on 10 March 2020).
- Soltanian, S.; Stuyven, E.; Cox, E.; Sorgeloos, P.; Bossier, P. Beta-glucans as immunostimulant in vertebrates and invertebrates. Crit. Rev. Microbiol. 2009, 35, 109–138. [Google Scholar] [CrossRef] [PubMed]
- Goodridge, H.S.; Wolf, A.J.; Underhill, D.M. β-Glucan recognition by the innate immune system. Immunol. Rev. 2009, 230, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Richter, J.; Svozil, V.; Král, V.; Dobiášová, L.R.; Vetvicka, V. β-glucan affects mucosal immunity in children with chronic respiratory problems under physical stress: Clinical trials. Ann. Transl. Med. 2015, 3, 2305–5839. [Google Scholar]
- Malin, E.V.J.; Gunnar, C.H. Immunological aspects of intestinal mucus and mucins. Nat. Rev. Immunol. 2016, 16, 639–649. [Google Scholar]
- Fredriksson, T.; Pettersson, U. Severe Psoriasis—Oral Therapy with a New Retinoid. Dermatologica 1978, 157, 238–244. [Google Scholar] [CrossRef]
- Inoue, R.; Ohue-Kitano, R.; Tsukahara, T.; Tanaka, M.; Masuda, S.; Inoue, T.; Yamakage, H.; Kusakabe, T.; Hasegawa, K.; Shimatsu, A.; et al. Prediction of functional profiles of gut microbiota from 16S rRNA metagenomic data provides a more robust evaluation of gut dysbiosis occurring in Japanease type 2 diabitic patients. J. Clin. Biochem. Nutr. 2017, 61, 217–221. [Google Scholar] [CrossRef] [Green Version]
- Krampis, K.; Booth, T.; Chapman, B.; Tiwari, B.; Bicak, M.; Field, D.; Nelson, K.E. Cloud BioLinux: Pre-configured and on-demand bioinformatics computing for the genomics community. BMC Bioinform. 2012, 13, 42. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Administration Days (D) | 0 D | 6 D | 28 D | 56 D | ||||
|---|---|---|---|---|---|---|---|---|
| Diet | Norml | Fucoidan | Normal | Fucoidan | Normal | Fucoidan | Normal | Fucoidan |
| Relative abundance (%) | ||||||||
| Actinobacteria | 0.43 ± 0.15 | 0.48 ± 0.15 | 0.38 ± 0.04 | 0.25 ± 0.05 (b, d) | 0.70 ± 0.19 | 1.88 ± 1.52 (b, c) | 0.45 ± 0.15 | 0.83 ± 0.72 |
| Bacteroidetes | 60.0 ± 7.69 | 65.4 ± 5.57 | 60.2 ± 2.90 | 79.2 ± 1.74 (c, a) | 59.1 ± 4.18 | 77.4 ± 2.78 | 59.4 ± 9.69 | 78.2 ± 6.42 (b, d) |
| Deferribacteres | 0.10 ± 0.07 | 0.08 ± 0.04 | 0.25 ± 0.11 | 0.33 ± 0.33 | 0.05 ± 0.05 | 0.10 ± 0.00 (a, c) | 0.20 ± 0.12 | 0.40 ± 0.31 |
| Firmicutes | 36.3 ± 8.56 | 29.2 ± 4.43 | 33.1 ± 2.18 | 16.3 ± 2.25 (b, c) | 30.8 ± 2.27 | 15.2 ± 1.9 (a, c) | 34.3 ± 9.05 | 16.3 ± 4.98 (b, d) |
| Proteobacteria | 1.35 ± 0.27 | 1.10 ± 0.16 | 1.95 ± 0.30 | 3.03 ± 0.33 (c, a) | 1.40 ± 0.27 | 3.50 ± 0.87 (a, c) | 1.73 ± 0.53 | 3.05 ± 0.62 (a, d) |
| TM7 | 3.75 ± 1.51 | 3.58 ± 1.29 | 3.73 ± 0.62 | 0.98 ± 0.26 (b, c) | 7.78 ± 1.92 | 1.98 ± 0.19 (c) | 3.80 ± 0.24 | 1.23 ± 0.11 (b, c) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takahashi, M.; Takahashi, K.; Abe, S.; Yamada, K.; Suzuki, M.; Masahisa, M.; Endo, M.; Abe, K.; Inoue, R.; Hoshi, H. Improvement of Psoriasis by Alteration of the Gut Environment by Oral Administration of Fucoidan from Cladosiphon Okamuranus. Mar. Drugs 2020, 18, 154. https://doi.org/10.3390/md18030154
Takahashi M, Takahashi K, Abe S, Yamada K, Suzuki M, Masahisa M, Endo M, Abe K, Inoue R, Hoshi H. Improvement of Psoriasis by Alteration of the Gut Environment by Oral Administration of Fucoidan from Cladosiphon Okamuranus. Marine Drugs. 2020; 18(3):154. https://doi.org/10.3390/md18030154
Chicago/Turabian StyleTakahashi, Masanobu, Kento Takahashi, Sunao Abe, Kosuke Yamada, Manami Suzuki, Mai Masahisa, Mari Endo, Keiko Abe, Ryo Inoue, and Hiroko Hoshi. 2020. "Improvement of Psoriasis by Alteration of the Gut Environment by Oral Administration of Fucoidan from Cladosiphon Okamuranus" Marine Drugs 18, no. 3: 154. https://doi.org/10.3390/md18030154