Characterization of a New Chitosanase from a Marine Bacillus sp. and the Anti-Oxidant Activity of Its Hydrolysate

Abstract
:1. Introduction
2. Results and Discussion
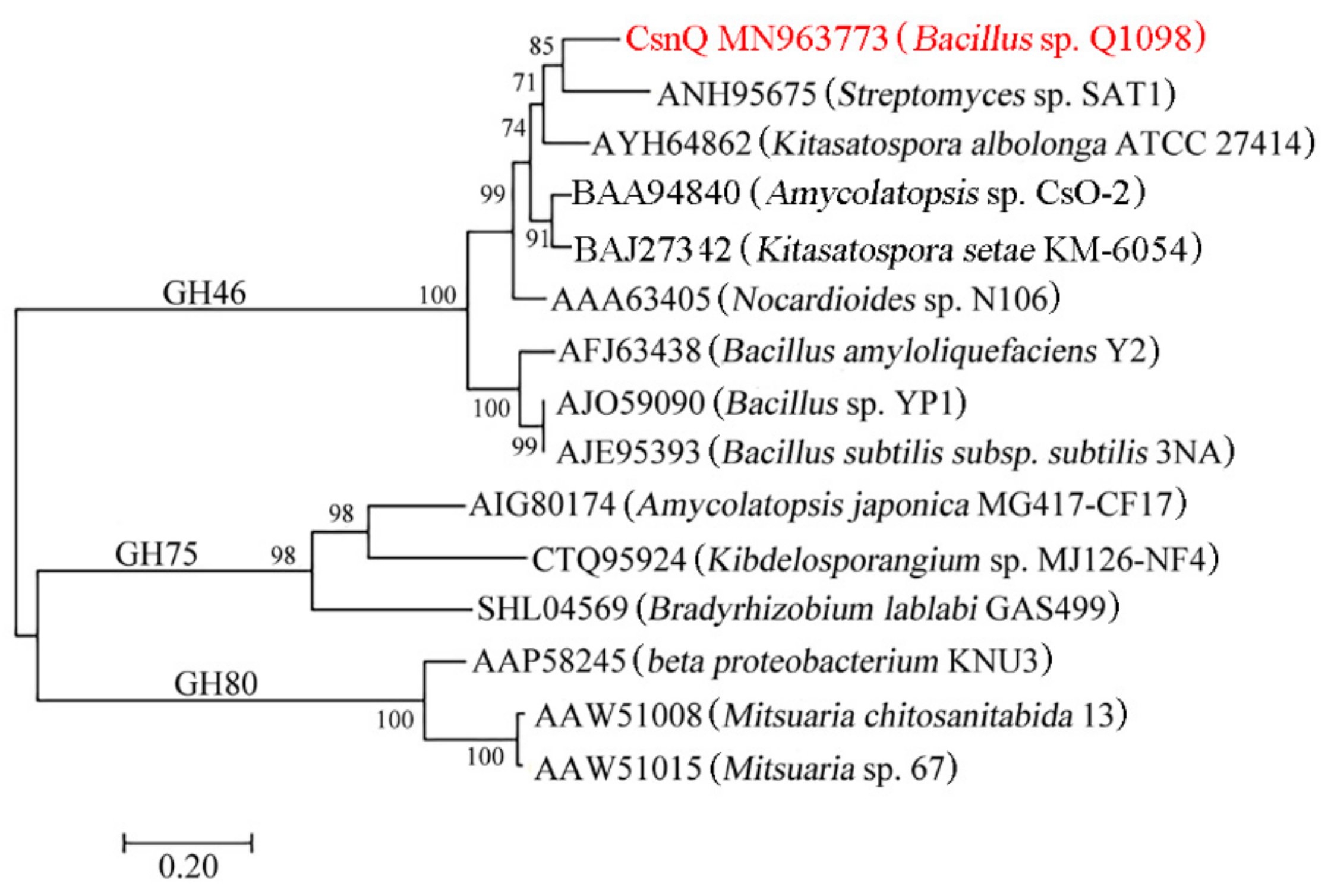
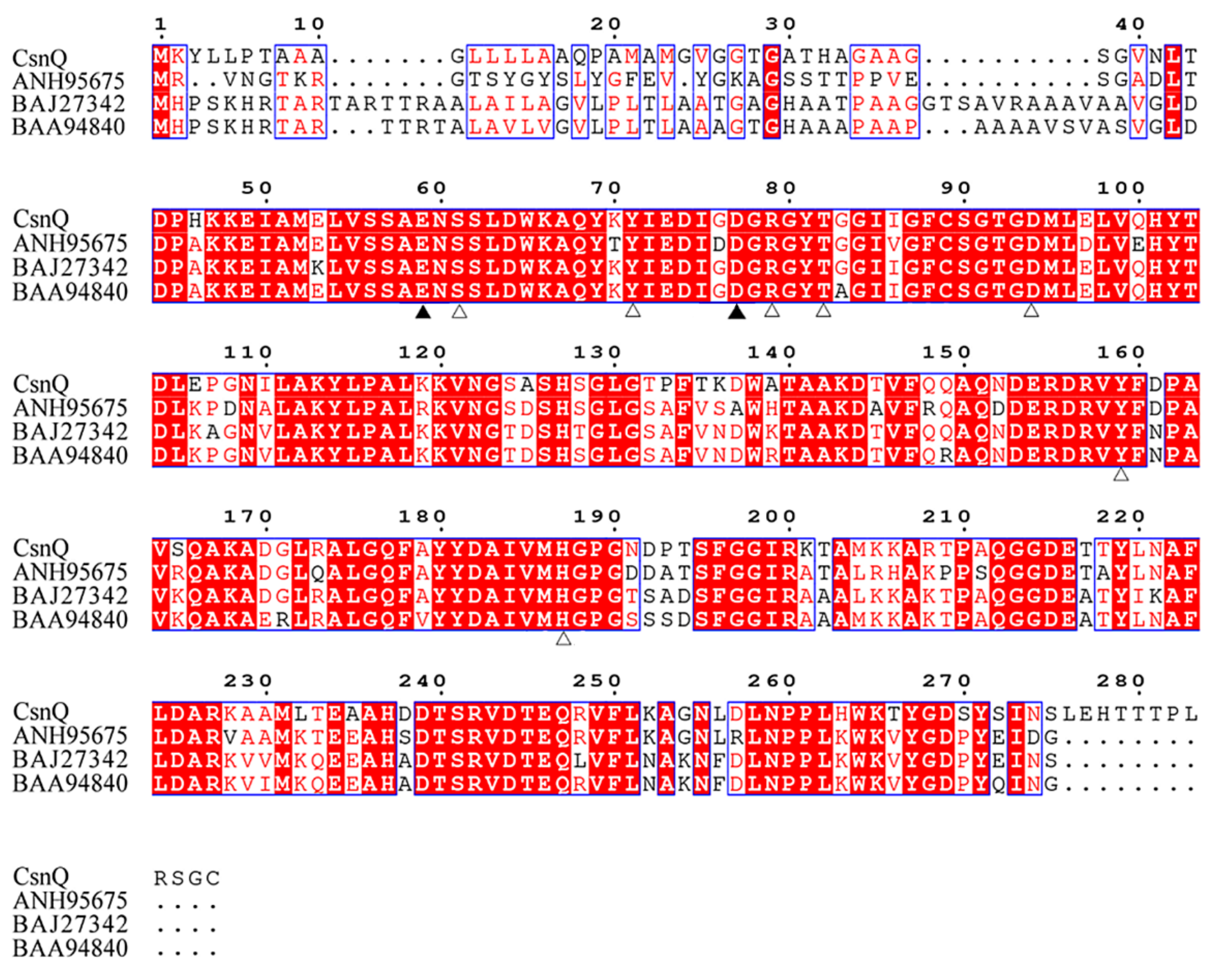
2.1. Sequence Analysis of CsnQ
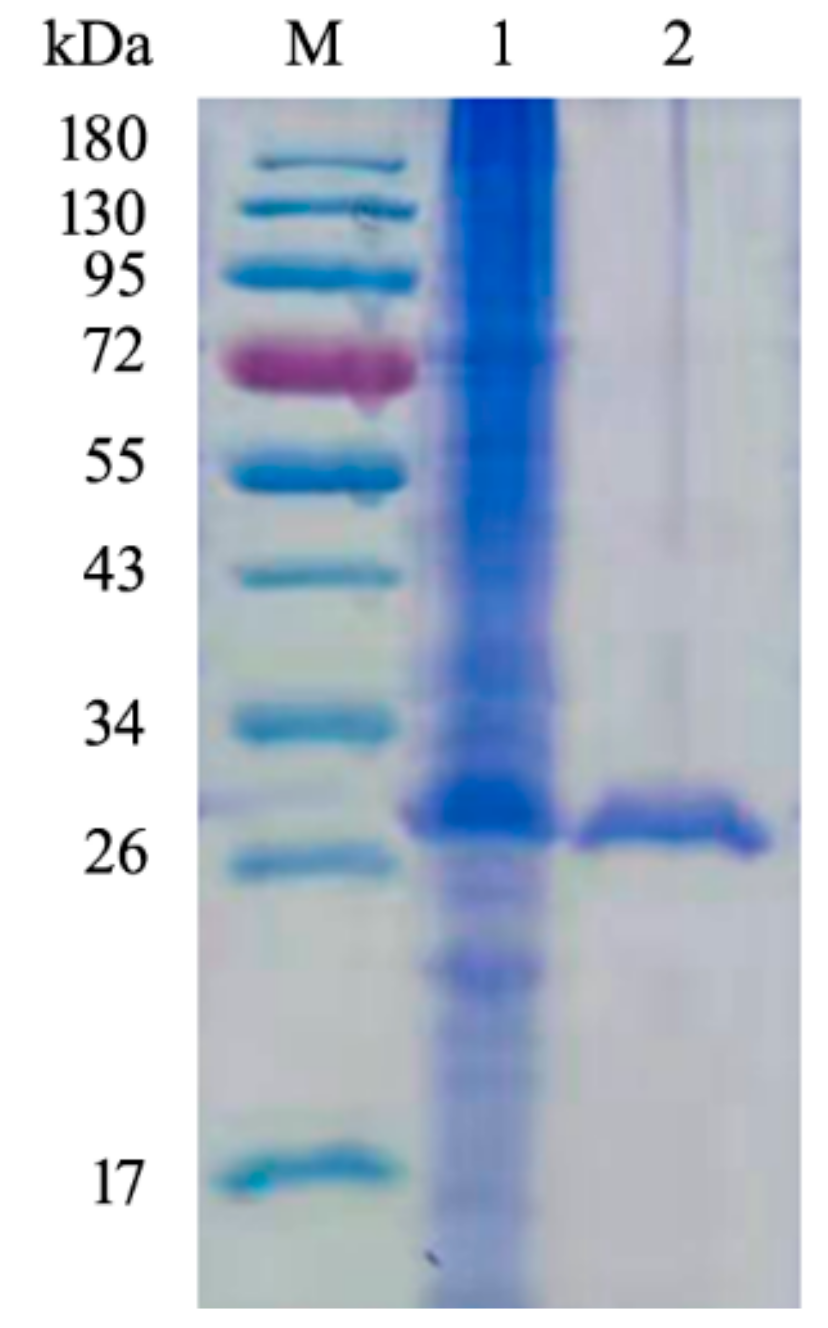
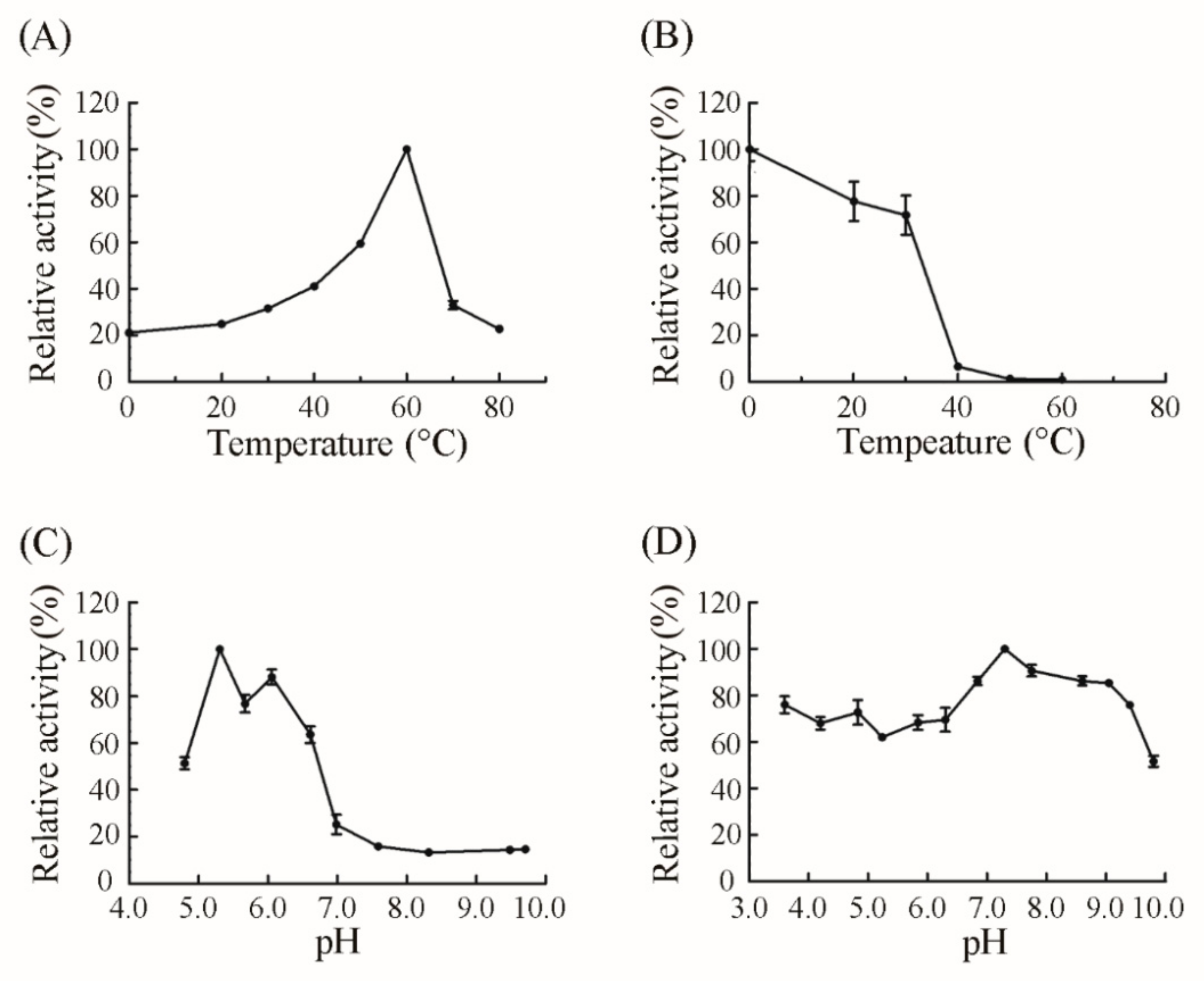
2.2. Expression, Purification and Characterization of CsnQ
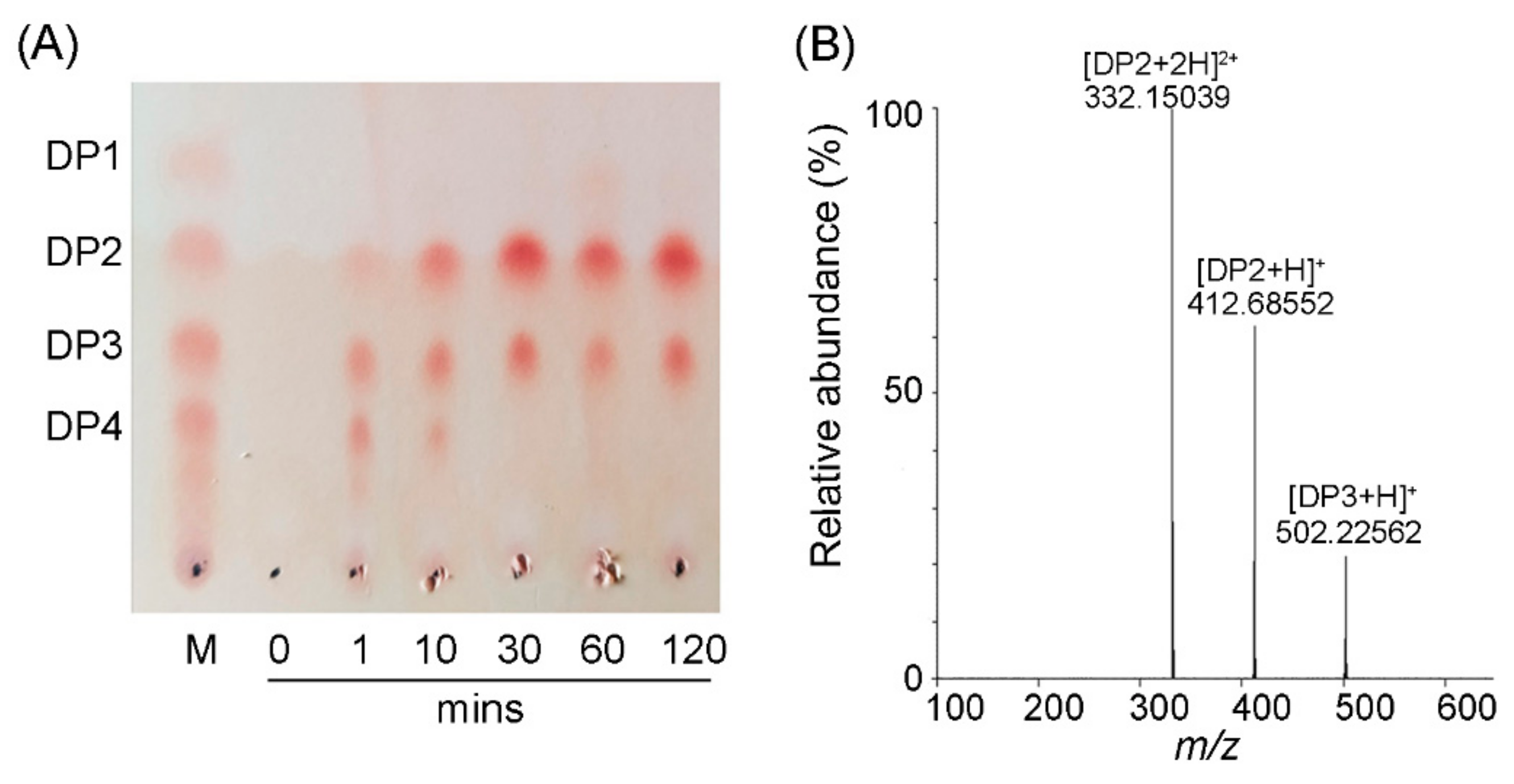
2.3. Action Mode and Reaction Product Analysis
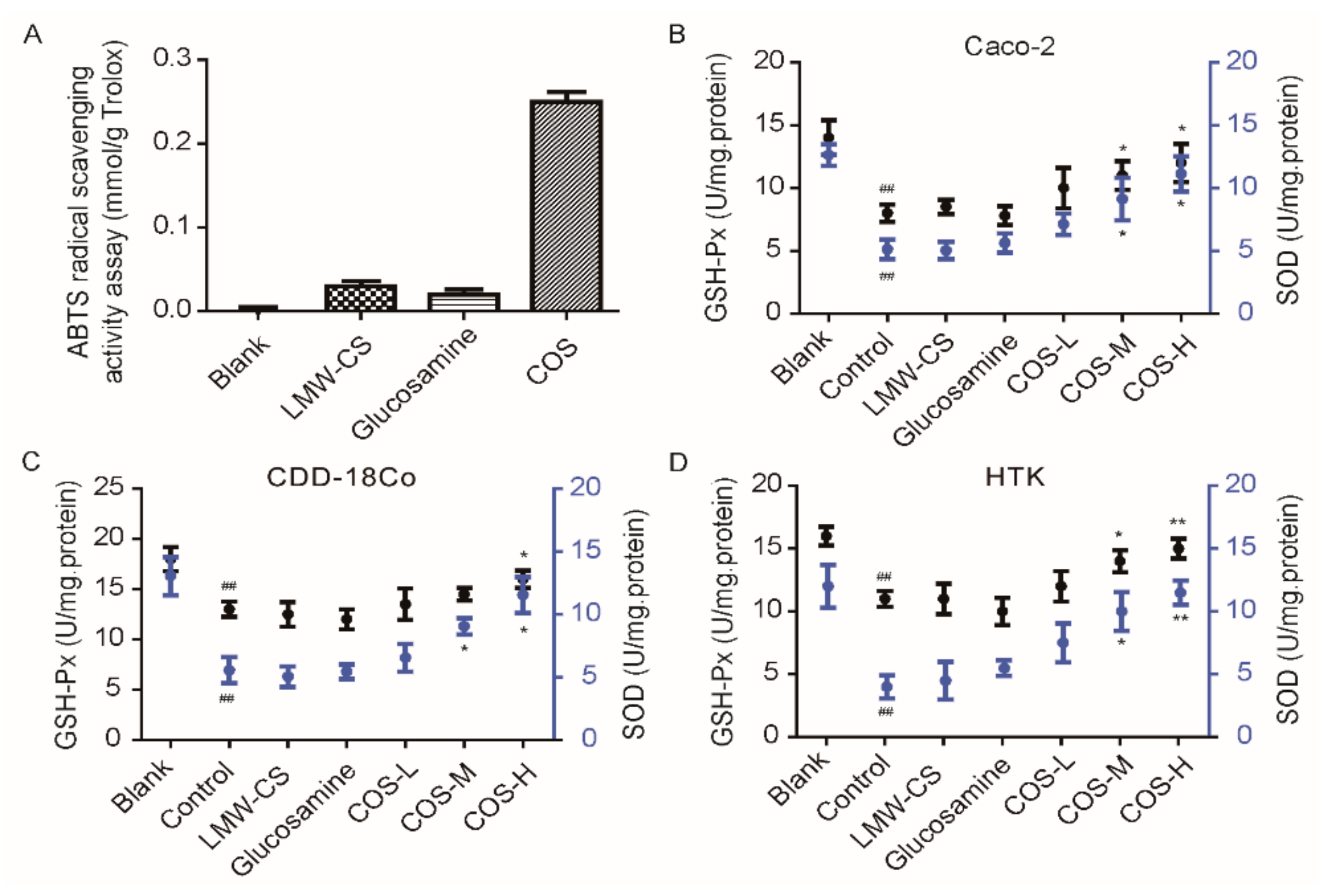
2.4. Anti-Oxidant Activity of COS
3. Materials and Methods
3.1. Strains and Materials
3.2. Sequence Analyses
3.3. Heterologous Expression of Recombinant CsnQ
3.4. Purification of Recombinant CsnQ
3.5. CsnQ Activity Assay
3.6. Effect of Temperature, pH, Metal Ions and Chemical Compounds on CsnQ Activity
3.7. Analysis of the CsnQ Reaction Products
3.8. In Vitro Analysis of the Anti-Oxidant Activity of COS
3.9. Cell Culture and Treatment
3.10. GSH-Px Activity
3.11. T-SOD Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Liaqat, F.; Eltem, R. Chitooligosaccharides and their biological activities: A comprehensive review. Carbohydr. Polym. 2018, 184, 243–259. [Google Scholar] [CrossRef]
- Harmsen, R.A.G.; Tuveng, T.R. Can we make chitosan by enzymatic deacetylation of chitin? Molecules 2019, 24, 3862. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Zheng, J.; Zhang, Y.; Zheng, P.; Tang, T.; Luo, J.K.; Cui, H.J.; Song, R.R.; Wang, Y. Chitosan hydrogel delivery system containing herbal compound functions as a potential antineuroinflammatory agent. ACS Omega 2019, 4, 10185–10191. [Google Scholar] [CrossRef] [PubMed]
- Yue, L.; Li, J.; Chen, W.; Liu, X.; Jiang, Q.; Xia, W. Geraniol grafted chitosan oligosaccharide as a potential antibacterial agent. Carbohydr. Polym. 2017, 176, 356–364. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.J.; Park, J.K.; Park, Y.I. Anti-inflammatory effects of low-molecular weight chitosan oligosaccharides in IgE-antigen complex-stimulated RBL-2H3 cells and asthma model mice. Int. Immunopharmacol. 2012, 12, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Park, P.J.; Je, J.Y.; Kim, S.K. Free radical scavenging activity of chitooligosaccharides by electron spin resonance spectrometry. J. Agric. Food Chem. 2003, 51, 4624–4627. [Google Scholar] [CrossRef] [PubMed]
- Kong, S.Z.; Li, D.D.; Luo, H.; Li, W.J.; Huang, Y.M.; Li, J.C.; Hu, Z.; Huang, N.; Guo, M.H.; Chen, Y.; et al. Anti-photoaging effects of chitosan oligosaccharide in ultraviolet-irradiated hairless mouse skin. Exp. Gerontol. 2018, 103, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Salah, R.; Michaud, P.; Mati, F.; Harrat, Z.; Lounici, H.; Abdi, N.; Drouiche, N.; Mameri, N. Anticancer activity of chemically prepared shrimp low molecular weight chitin evaluation with the human monocyte leukaemia cell line, THP-1. Int. J. Biol. Macromol. 2013, 52, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Rahimnejad, S.; Yuan, X.; Wang, L.; Lu, K.; Song, K.; Zhang, C. Chitooligosaccharide supplementation in low-fish meal diets for Pacific white shrimp (Litopenaeus vannamei): Effects on growth, innate immunity, gut histology, and immune-related genes expression. Fish Shellfish Immunol. 2018, 80, 405–415. [Google Scholar] [CrossRef]
- Kumirska, J.; Weinhold, M.X.; Sauvageau, J.C.; Thoming, J.; Kaczynski, Z.; Stepnowski, P. Determination of the pattern of acetylation of low-molecular-weight chitosan used in biomedical applications. J. Pharm. Biomed. Anal. 2009, 50, 587–590. [Google Scholar] [CrossRef]
- Mendis, E.; Kim, M.M.; Rajapakse, N.; Kim, S.K. An in vitro cellular analysis of the radical scavenging efficacy of chitooligosaccharides. Life Sci. 2007, 80, 2118–2127. [Google Scholar] [CrossRef] [PubMed]
- Artan, M.; Karadeniz, F.; Karagozlu, M.Z.; Kim, M.M.; Kim, S.K. Anti-HIV-1 activity of low molecular weight sulfated chitooligosaccharides. Carbohydr. Res. 2010, 345, 656–662. [Google Scholar] [CrossRef]
- Cho, E.J.; Rahman, M.A.; Kim, S.W.; Baek, Y.M.; Hwang, H.J.; Oh, J.Y.; Hwang, H.S.; Lee, S.H.; Yun, J.W. Chitosan oligosaccharides inhibit adipogenesis in 3T3-L1 adipocytes. J. Microbiol. Biotechnol. 2008, 18, 80–87. [Google Scholar] [PubMed]
- Lee, B.C.; Kim, M.S.; Choi, S.H.; Kim, K.Y.; Kim, T.S. In vitro and in vivo antimicrobial activity of water-soluble chitosan oligosaccharides against Vibrio vulnificus. Int. J. Mol. Med. 2009, 24, 327–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, J.C.; Tavaria, F.K.; Soares, J.C.; Ramos, O.S.; Joao Monteiro, M.; Pintado, M.E.; Xavier Malcata, F. Antimicrobial effects of chitosans and chitooligosaccharides, upon Staphylococcus aureus and Escherichia coli, in food model systems. Food Microbiol. 2008, 25, 922–928. [Google Scholar] [CrossRef]
- Johnsen, M.G.; Hansen, O.C.; Stougaard, P. Isolation, characterization and heterologous expression of a novel chitosanase from Janthinobacterium sp. strain 4239. Microb. Cell Factories 2010, 9, 5. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.; Zheng, J.; Jiao, S.; Cheng, G.; Feng, C.; Du, Y.; Liu, H. A review on the preparation of chitosan oligosaccharides and application to human health, animal husbandry and agricultural production. Carbohydr. Polym. 2019, 220, 60–70. [Google Scholar] [CrossRef]
- Fernandes, J.C.; Sereno, J.; Garrido, P.; Parada, B.; Cunha, M.F.; Reis, F.; Pintado, M.E.; Santos-Silva, A. Inhibition of bladder tumor growth by chitooligosaccharides in an experimental carcinogenesis model. Mar. Drugs 2012, 10, 2661–2675. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Wang, L.; Chen, X.; Sun, M.; Han, Y. Design and synthesis of a chitodisaccharide-based affinity resin for chitosanases purification. Mar. Drugs 2019, 17, 68. [Google Scholar] [CrossRef] [Green Version]
- Marcotte, E.M.; Monzingo, A.F.; Ernst, S.R.; Brzezinski, R.; Robertus, J.D. X-ray structure of an anti-fungal chitosanase from streptomyces N174. Nat. Struct. Biol. 1996, 3, 155–162. [Google Scholar] [CrossRef]
- Saito, J.; Kita, A.; Higuchi, Y.; Nagata, Y.; Ando, A.; Miki, K. Crystal structure of chitosanase from Bacillus circulans MH-K1 at 1.6-A resolution and its substrate recognition mechanism. J. Biol. Chem. 1999, 274, 30818–30825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukamizo, T. Chitinolytic enzymes: Catalysis, substrate binding, and their application. Curr. Protein Pept. Sci. 2000, 1, 105–124. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Q.; Shi, Y.; Wang, S.; Yang, Y.; Han, B.; Liu, W.; Jones, D.N.; Liu, W. Structural and biochemical insights into the degradation mechanism of chitosan by chitosanase OU01. Biochim. Biophys. Acta. 2015, 1850, 1953–1961. [Google Scholar] [CrossRef]
- Lyu, Q.; Wang, S.; Xu, W.; Han, B.; Liu, W.; Jones, D.N.; Liu, W. Structural insights into the substrate-binding mechanism for a novel chitosanase. Biochem. J. 2014, 461, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chen, X.; Li, X.; Han, Y.; Wang, Y.; Yao, R.; Li, S. Purification and characterization of a new cold-adapted and thermo-tolerant chitosanase from marine bacterium Pseudoalteromonas sp. SY39. Molecules 2019, 24, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Zheng, Z.; Xiao, Y.; Zhang, J.; Zhou, Y.; Li, X.; Li, S.; Yu, H. Cloning and characterization of a cold-adapted chitosanase from marine bacterium Bacillus sp. BY01. Molecules 2019, 24, 3915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Mao, X.; Guo, N.; Zhao, L.; Cao, R.; Liu, Q. Discovery and characterization of a novel chitosanase from Paenibacillus dendritiformis by phylogeny-based enzymatic product specificity prediction. J. Agric. Food Chem. 2018, 66, 4645–4651. [Google Scholar] [CrossRef]
- Masson, J.Y.; Boucher, I.; Neugebauer, W.A.; Ramotar, D.; Brzezinski, R. A new chitosanase gene from a Nocardioides sp. is a third member of glycosyl hydrolase family 46. Microbiology 1995, 141 Pt 10, 2629–2635. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.S.; Yoo, J.S.; Chung, S.Y.; Lee, Y.C.; Cho, Y.S.; Choi, Y.L. Cloning, purification, and characterization of chitosanase from Bacillus sp. DAU101. Appl. Microbiol. Biotechnol. 2006, 73, 113–121. [Google Scholar] [CrossRef]
- Yoon, H.G.; Kim, H.Y.; Lim, Y.H.; Kim, H.K.; Shin, D.H.; Hong, B.S.; Cho, H.Y. Thermostable chitosanase from Bacillus sp. strain CK4: Cloning and expression of the gene and characterization of the enzyme. Appl. Environ. Microbiol. 2000, 66, 3727–3734. [Google Scholar] [CrossRef] [Green Version]
- Liang, T.W.; Lo, B.C.; Wang, S.L. Chitinolytic bacteria-assisted conversion of squid pen and its effect on dyes and pigments adsorption. Mar. Drugs 2015, 13, 4576–4593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, R.; Mendis, E.; Kim, S.K. Factors affecting the free radical scavenging behavior of chitosan sulfate. Int. J. Biol. Macromol. 2005, 36, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Je, J.Y.; Park, P.J.; Kim, S.K. Free radical scavenging properties of hetero-chitooligosaccharides using an ESR spectroscopy. Food Chem. Toxicol. 2004, 42, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Dai, R.; Wang, Y.; Li, M.; Zhu, Y.; Chen, L.; Kang, L.; Liu, Z.; Yang, Y.; Yuan, S. A novel thermophilic exochitinase ChiEn3 from Coprinopsis cinerea exhibits a hyperhydrolytic activity toward 85% deacetylated chitosan and a significant application to preparation of chitooligosaccharides from the chitosan. Carbohydr. Polym. 2019, 207, 729–736. [Google Scholar] [CrossRef]
- Weber, H.; Roesner, J.P.; Nebe, B.; Rychly, J.; Werner, A.; Schroder, H.; Jonas, L.; Leitzmann, P.; Schneider, K.P.; Dummler, W. Increased cytosolic Ca2+ amplifies oxygen radical-induced alterations of the ultrastructure and the energy metabolism of isolated rat pancreatic acinar cells. Digestion 1998, 59, 175–185. [Google Scholar] [CrossRef]
- Xu, Q.; Ma, P.; Yu, W.; Tan, C.; Liu, H.; Xiong, C.; Qiao, Y.; Du, Y. Chitooligosaccharides protect human embryonic hepatocytes against oxidative stress induced by hydrogen peroxide. Mar. Biotechnol. 2010, 12, 292–298. [Google Scholar] [CrossRef]
- Senevirathne, M.; Ahn, C.-B.; Je, J.-Y. Hepatoprotective effect of chitooligosaccharides against tert-butylhydroperoxide-induced damage in Chang liver cells. Carbohydr. Polym. 2011, 83, 995–1000. [Google Scholar] [CrossRef]
- Karadeniz, F.; Artan, M.; Kong, C.-S.; Kim, S.-K. Chitooligosaccharides protect pancreatic β-cells from hydrogen peroxide-induced deterioration. Carbohydr. Polym. 2010, 82, 143–147. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | GenBank No. | Orgnism Source | Optimal pH | Stable pH Range | Optimal Temperature (°C) | Products (DP) | References |
|---|---|---|---|---|---|---|---|
| CsnQ | MN963773 | Bacillus sp. Q1098 | 5.31 | 6.8–9.1 | 60 | Mainly 2 | This study |
| CsnM | MH675972 | Pseudoalteromonas sp. SY39 | 5.9 | 5.8–7.9 | 40 | 2–3 | [25] |
| CsnB | MN531545 | Bacillus sp. BY01 | 5.0 | 4.6–5.8 | 35 | 2–3 | [26] |
| Csn-PD | − | Paenibacillus dendritiformis | 7.0 | 6.0–7.0 | 45 | 2–6 | [27] |
| CSN | JF950269 | Penicillium sp. D-1 | 4.0 | 3.0–5.0 | 48 | − | [28] |
| CSN-SP | DQ316095 | Bacillus sp. DAU101 | 7.5 | − | 50 | 2–6 | [29] |
| chitosanase | AF165188 | Bacillus sp. CK4 | 7.5 | − | 55 | 2–6 | [30] |
| chitosanase | GQ487532 | Janthinobacterium sp. 4239 | 5.0 | − | 45 | 1–2 | [16] |
| chitosanase | − | cereus TKU034 | 7.0 | 4.5–7.5 | 50 | 3–9 | [31] |
| Cell Types | Fragments | Concentration (μg/mL) | Anti-Oxidant Activity | References |
|---|---|---|---|---|
| Caco-2 CDD-18Co HTK | DP1 DP2 ~5 kDa | 100, 300, 500 | DP2 is the minimal fragment in free radical scavenging. | This study |
| Murine melanoma cell line | <1 kDa, 1–3 kDa. | 10, 50, 100, 500 | The effect of fragment in <1 kDa is better than in 1–3 kDa. | [11] |
| Human embryonic hepatocytes (L02 cells) | DP2–8 | 100–400 | The mixture plays effective free radical scavenging capacity. | [36] |
| Chang liver cells | 5–10 kDa, 1–5 kDa, <1 kDa. | 100, 200, 500, 1000 | The activity of anti-oxidant enzymes and the inhibition capacity of ROS are maximal in 5–10 kDa. The GSH (glutathione) content is highest in 1–5 kDa. | [37] |
| Human pancreatic β-cells | <1 kDa, 1–3 kDa, 3–5 kDa, 5–10 kDa. | 500 | The 3–5 kDa of COS possesses the highest anti-oxidant activity. | [38] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, C.; Li, X.; Yang, K.; Li, S. Characterization of a New Chitosanase from a Marine Bacillus sp. and the Anti-Oxidant Activity of Its Hydrolysate. Mar. Drugs 2020, 18, 126. https://doi.org/10.3390/md18020126
Ma C, Li X, Yang K, Li S. Characterization of a New Chitosanase from a Marine Bacillus sp. and the Anti-Oxidant Activity of Its Hydrolysate. Marine Drugs. 2020; 18(2):126. https://doi.org/10.3390/md18020126
Chicago/Turabian StyleMa, Chunrui, Xiao Li, Kun Yang, and Shangyong Li. 2020. "Characterization of a New Chitosanase from a Marine Bacillus sp. and the Anti-Oxidant Activity of Its Hydrolysate" Marine Drugs 18, no. 2: 126. https://doi.org/10.3390/md18020126