A Pharmacological Comparison of Two Isomeric Nicotinic Receptor Agonists: The Marine Toxin Isoanatabine and the Tobacco Alkaloid Anatabine

Abstract
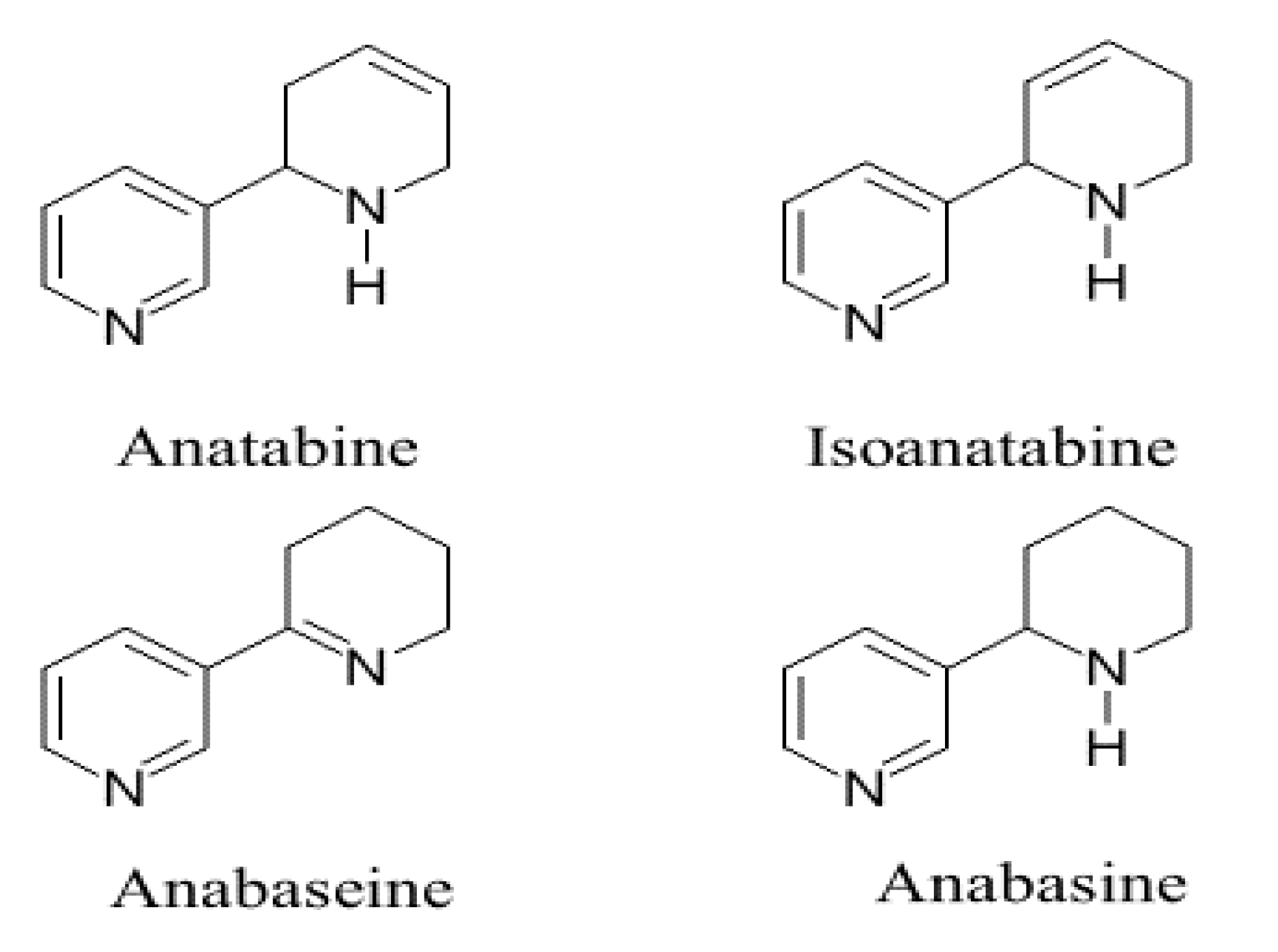
:1. Introduction
2. Results
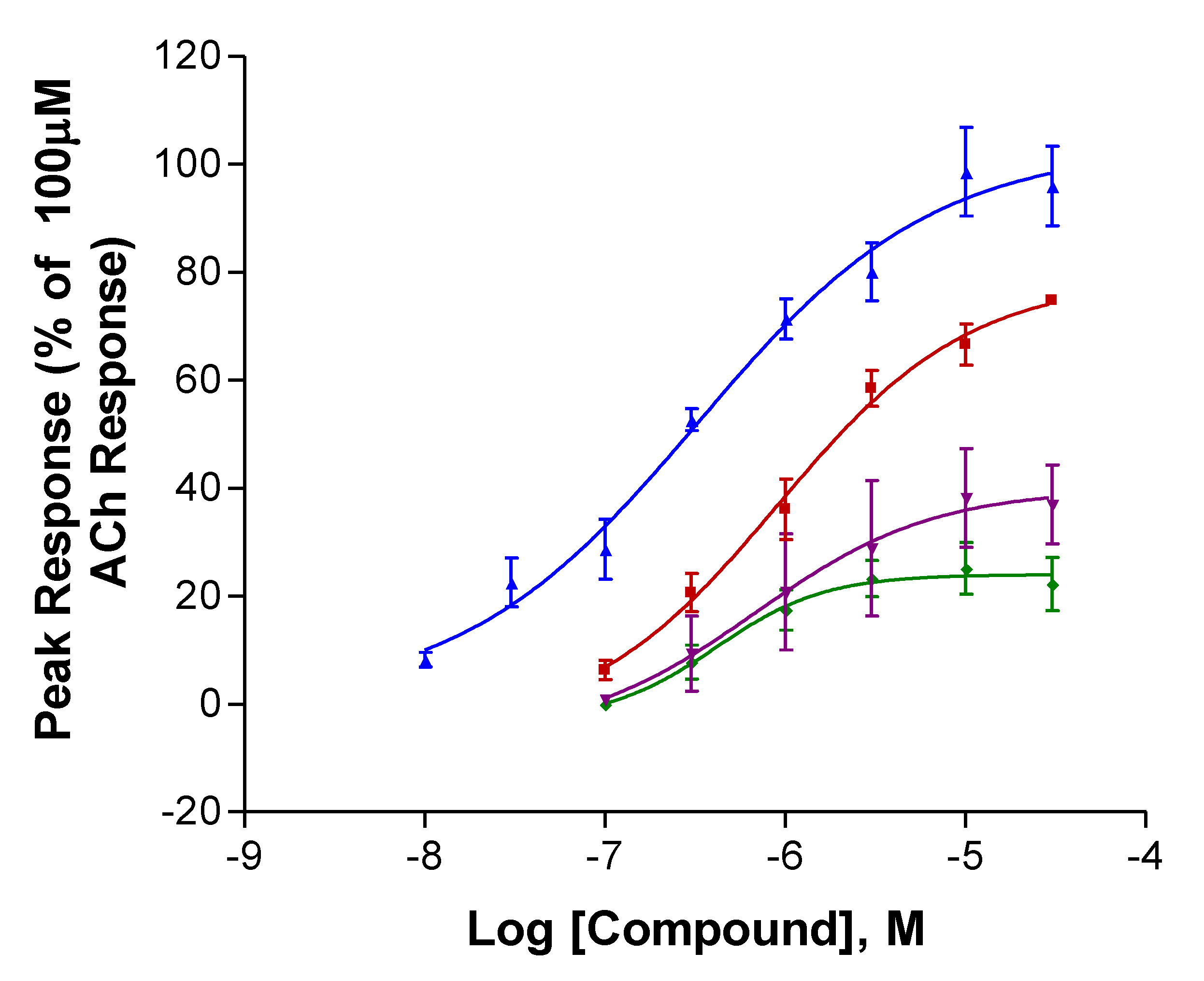
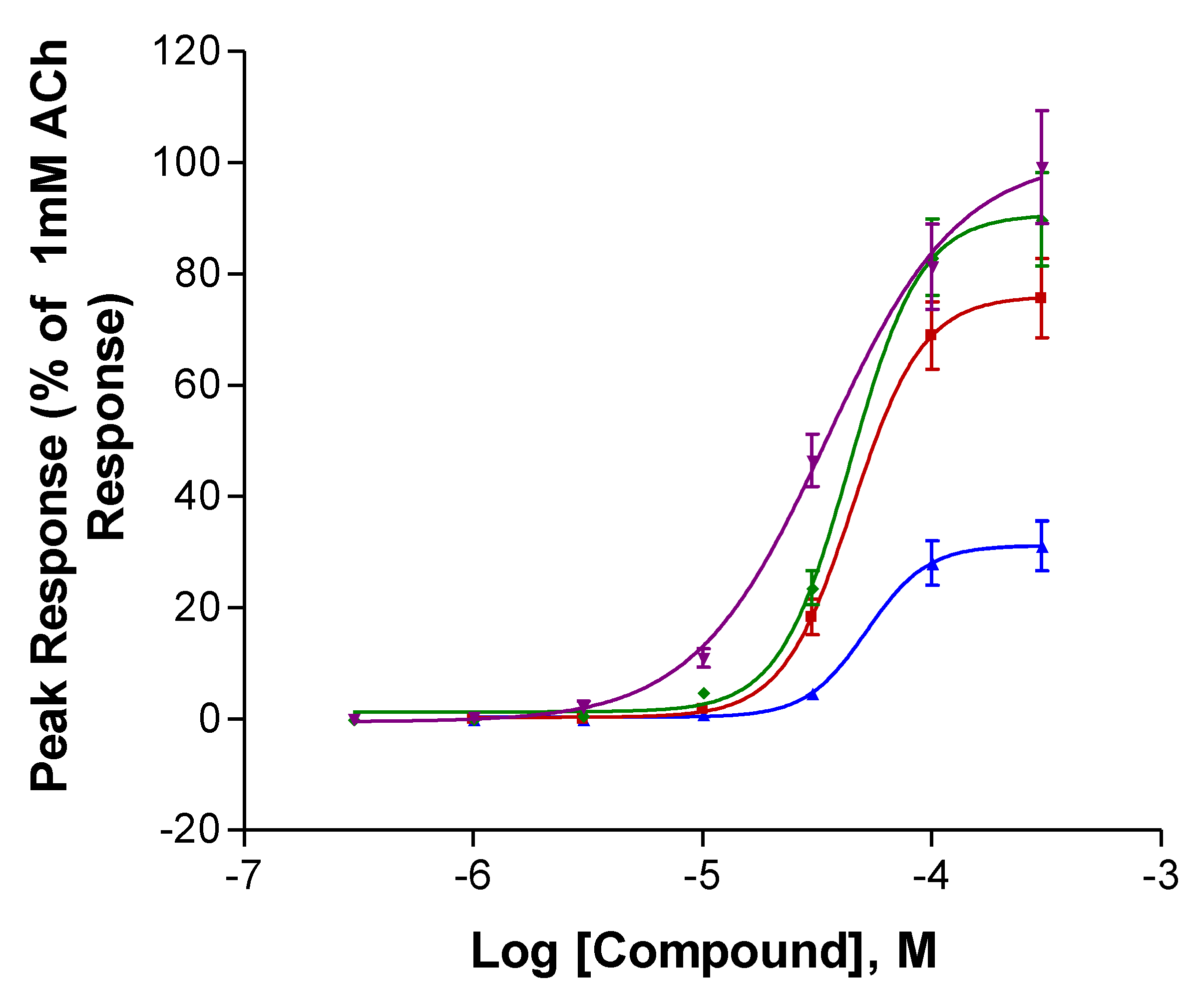
2.1. Experiments with Human nAChRs Expressed in Xenopus Oocytes
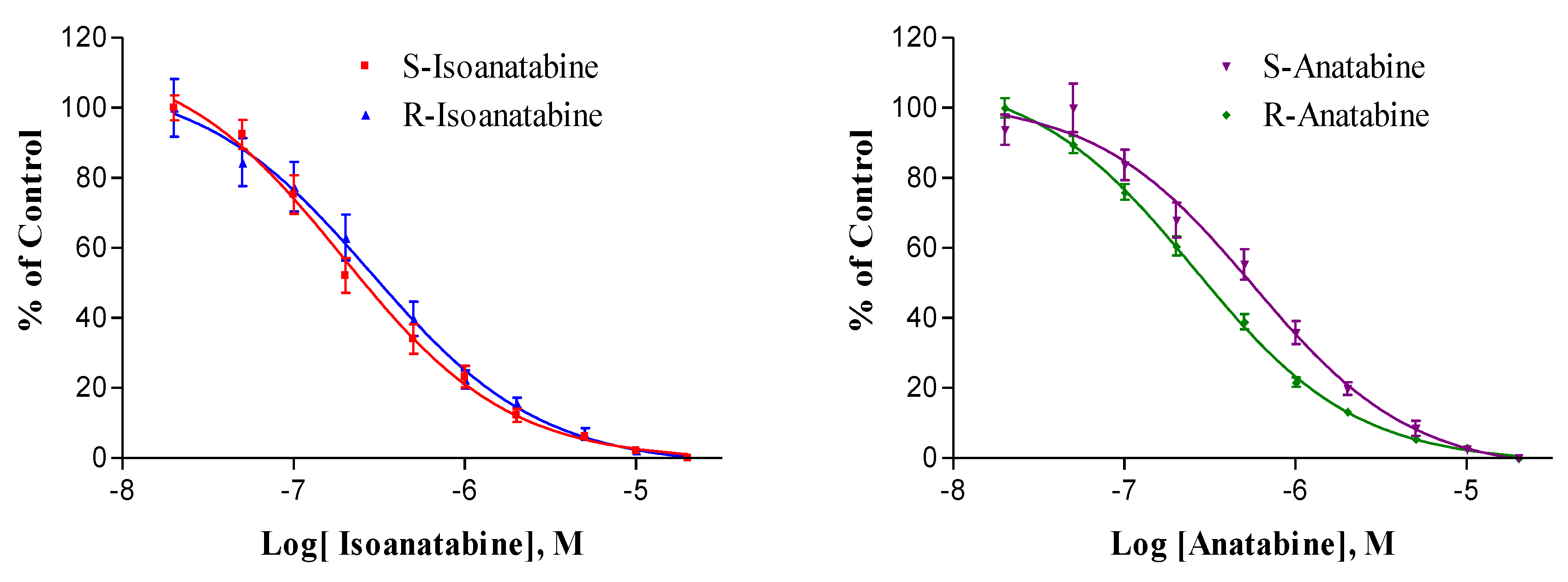
2.2. Radioligand Binding Experiments with Rat Brain nAChRs
3. Discussion
4. Materials and Methods
4.1. Isoanatabine and Anatabine Syntheses
4.2. Two-Electrode Voltage-Clamp Analysis of Human Brain nAChRs
4.3. Radioligand Binding
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wu, J.; Lukas, R.J. Naturally-expressed nicotinic receptor subtypes. Biochem. Pharmacol. 2011, 82, 800–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Changeux, J.P. The nicotinic acetylcholine receptor: The founding father of the pentameric ligand-gated ion channel superfamily. J. Biol. Chem. 2012, 287, 40207–40215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoli, M.; Pucci, S.; Vilella, A.; Gotti, C. Neuronal and extraneuronal nicotinic acetylcholine receptors. Curr. Neuropharmacol. 2018, 16, 338–349. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, D.; Lee, C.-H.; Flood, D.; Marger, F.; Donnelly-Roberts, A. Therapeutic potential of α7 nicotinic acetylcholine receptors. Pharmacol. Rev. 2015, 67, 1025–1073. [Google Scholar] [CrossRef] [Green Version]
- Bouzat, C.; Lasala, M.; Nielsen, B.E.; Corradi, J.; Esandi, M.D.C. Molecular function of α7 receptors as drug targets. J. Physiol. 2018, 596, 1847–1861. [Google Scholar] [CrossRef]
- Kem, W.R. The brain alpha7 nicotinic receptor may be an important therapeutic target for the treatment of Alzheimer’s disease: Studies with DMXBA (GTS-21). Behav. Brain Res. 2000, 113, 169–183. [Google Scholar] [CrossRef]
- Keefe, R.S.E.; Meltzer, H.A.; Dgetluck, N.; Gawryl, M.; Koenig, G.; Moebius, H.J.; Lombardo, I.; Hilt, D.C. Randomized, double-blind, placebo-controlled study of encenicline, an α7 nicotinic acetylcholine receptor agonist, as a treatment for cognitive impairment in schizophrenia. Neuropsychopharmacology 2015, 40, 3053–3060. [Google Scholar] [CrossRef]
- Haig, G.; Bain, E.; Robieson, W.; Baker, J.; Othman, A. A randomized, double-blind trial to assess the efficacy and safety of ABT-126, a selective α7 nicotinic acetylcholine receptor agonist in the treatment of cognitive impairment in subjects with schizophrenia. Am. J. Psychiat. 2016, 173, 827–835. [Google Scholar] [CrossRef] [Green Version]
- Kem, W.R.; Olincy, A.; Johnson, L.; Harris, J.; Wagner, B.D.; Buchanan, R.W.; Christians, U.; Freedman, R. Pharmacokinetic Limitations on Effects of an Alpha7-Nicotinic Receptor Agonist in Schizophrenia: Randomized Trial with an Extended-Release Formulation. Neuropsychopharmacology 2018, 43, 583–589. [Google Scholar] [CrossRef] [Green Version]
- Rollema, H.; Coe, J.W.; Chambers, L.K.; Hurst, R.S.; Stahl, S.M.; Williams, K.E. Rationale, pharmacology and clinical efficacy of partial agonists of alpha4beta2 nACh receptors for smoking cessation. Trends Pharmacol. Sci. 2007, 28, 316–325. [Google Scholar] [CrossRef]
- Bootman, M.D.; Bultynck, G. Fundamentals of cellular calcium signaling: A primer. Cold Spring Harb. Persp. Biol. 2020, 12, a038802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunzell, D.H.; McIntosh, J.M.; Papke, R.L. Diverse strategies targeting α7 homomeric and α6β2* heteromeric nicotinic acetylcholine receptors for smoking cessation. Ann. N. Y. Acad. Sci. 2014, 1327, 27–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burghaus, L.; Schutz, U.; Krempel, U.; Lindstrom, J.; Schroder, H. Loss of nicotinic acetylcholine receptor subunits alpha4 and alpha7 in the cerebral cortex of Parkinson patients. Parkinsonism Relat. Disord. 2003, 9, 243–246. [Google Scholar] [CrossRef]
- Pavlov, V.A.; Ochani, M.; Yang, L.H.; Gallowitsch-Puerta, M.; Ochani, K.; Lin, X.; Levi, J.; Parrish, W.R.; Rosas-Ballina, M.; Czura, C.J.; et al. Selective alpha7-nicotinic acetylcholine receptor agonist GTS-21 improves survival in murine endotoxemia and severe sepsis. Crit. Care Med. 2007, 35, 1139–1144. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Wang, J.; Chen, F.; Xu, Y.; Zhang, H.; Wang, Y. α7nAChR deletion aggravates myocardial infarction and enhances systemic inflammatory reaction via mTOR-signaling-related autophagy. Inflammation 2019, 42, 1190–1202. [Google Scholar] [CrossRef]
- Wonnacott, S.; Gallagher, T. The chemistry and pharmacology of anatoxin-a and related homotropanes with respect to nicotinic acetylcholine receptors. Mar. Drugs 2006, 4, 228–254. [Google Scholar] [CrossRef] [Green Version]
- Kem, W.R.; Abbott, B.C.; Coates, R.M. Isolation and structure of a hoplonemertine toxin. Toxicon 1971, 9, 15–22. [Google Scholar] [CrossRef]
- Kem, W.R.; Soti, F.; Wildeboer, K.; LeFrancois, S.; MacDougall, K.; Wei, D.-Q.; Chou, K.-C.; Arias, H. The nemertine toxin anabaseine and its derivative DMXBA (GTS-21): Chemical and pharmacological properties. Mar. Drugs 2006, 4, 255–273. [Google Scholar] [CrossRef] [Green Version]
- Badio, B.T.; Daly, J.W. Epibatidine, a potent analgetic and nicotinic agonist. Mol. Pharm. 1994, 45, 563–569. [Google Scholar]
- Harvey, S.C.; Maddox, F.N.; Luetje, C.W. Multiple determinants of dihydro-β-erythroidine sensitivity on rat neuronal nicotinic receptor αsubunits. J. Neurochem. 1996, 67, 1953–1959. [Google Scholar] [CrossRef]
- Coates, P.A.; Blagbrough, I.S.; Hardick, D.J.; Rowan, M.G.; Wonnacott, S.; Potter, B.V.L. Rapid and efficient isolation of the nicotinic receptor antagonist methyllcaconitine from delphinium—Assignment of the methylsuccinimide absolute stereochemistry. Tetrah. Lett. 1994, 35, 8701–8704. [Google Scholar] [CrossRef]
- Orechoff, A.; Menschikoff, G. Über die alkaloide von Anabasis aphylla Linnaeus. II. Mitteilung: Zur konstitution des anabasins. Ber. Dtsch. Chem Ges. 1932, 65, 232. [Google Scholar] [CrossRef]
- Lee, S.T.; Wildeboer, K.; Panter, K.E.; Kem, W.R.; Gardner, D.R.; Molyneux, R.J.; Chang, S.-W.T.; Soti, F.; Pfister, J.A. Relative toxicities and neuromuscular nicotinic receptor agonistic potencies of anabasine enantiomers and anabaseine. Neurotoxicol. Teratol. 2006, 28, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Green, B.T.; Lee, S.T.; Panter, K.E.; Welch, K.D.; Cook, D.; Pfister, J.A.; Kem, W.R. Actions of piperidine alkaloid teratogens at fetal nicotinic acetylcholine receptors. Neurotoxicol. Teratol. 2010, 32, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Kem, W.R.; Mahnir, V.M.; Papke, R.; Lingle, C. Anabaseine is a potent agonist upon muscle and neuronal alpha-bungarotoxin sensitive nicotinic receptors. J. Pharmacol. Exper. Therap. 1997, 283, 979–992. [Google Scholar]
- Andrud, A.; Xing, H.; Gabrielsen, B.; Bloom, L.; Mahnir, V.; Lee, S.; Green, B.T.; Lindstrom, J.; Kem, W.R. Investigation of the possible pharmacologically active forms of the nicotinic acetylcholine receptor agonist anabaseine. Mar. Drugs 2019, 17, 614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kem, W.R.; Scott, K.N.; Duncan, J.H. Hoplonemertine worms—A new source of pyridine neurotoxins. Experientia 1976, 32, 684–686. [Google Scholar] [CrossRef]
- Bouillon, A.; Voisin, S.S.; Robic, A.; Lancelot, J.-C.; Collot, V.; Rault, S. An efficient two-step synthesis of the quaterpyridine nemertelline. J. Org. Chem. 2003, 68, 10178–10180. [Google Scholar] [CrossRef]
- Kem, W.R.; Rocca, J.; Garraffo, H.M.; Spande, T.F.; Daly, J.W.; Soti, F. Synthesis and Spectroscopic Comparison of the Eight Methyl-2,3′-bipyridyls and Identification of a Hoplonemertine Alkaloid as 3-Methyl-2,3′-bipyridyl. Heterocycles 2009, 79, 1025–1041. [Google Scholar] [CrossRef]
- Kem, W.R.; Soti, F.; Rittschof, D. Inhibition of barnacle larval settlement and crustacean toxicity of some hoplonemertine pyridyl alkaloids. Biomol. Eng. 2003, 20, 355–361. [Google Scholar] [CrossRef]
- Rouchaud, A.; Kem, W.R. A convenient synthesis of two isomeric tetrahydropyridyl alkaloids: Isoanatabine and anatabine. J. Heterocycl. Chem. 2010, 47, 569–581. [Google Scholar] [CrossRef]
- Späth, E.; Kestler, F.L. Anatabin, ein neues tabak alkaloid. Ber. Dtsch. Chem. Ges. 1937, 239–247. [Google Scholar] [CrossRef]
- Ji, H.; Wu, Y.; Fannin, F.; Bush, L. Determination of tobacco alkaloid enantiomers using reversed phase UPLC/MS/MS. Heliyon 2019, 5, e01719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, P., III; Yu, L.; Shulgin, A.T.; Benowitz, N.L. Minor alkaloids as biomarkers for tobacco use: Comparison of users of cigarettes, smokeless tobacco, cigars, and pipes. Amer. J. Public Health 1999, 89, 731–736. [Google Scholar] [CrossRef] [Green Version]
- Parker, M.J.; Beck, A.; Luetje, C.W. Neuronal nicotinic receptor beta2 and beta4 subunits confer large differences in agonist binding affinity. Mol. Pharmacol. 1998, 54, 1132–1139. [Google Scholar] [CrossRef]
- Celie, P.H.N.; van Rossum-Fikkert, S.E.; van Kijk, W.J.; Brejc, K.; Smit, A.B.; Sixma, T.K. Nicotine and carbamylcholine binding to nicotinic acetylcholine receptors as studied in AChBP crystal structures. Neuron 2004, 41, 907–914. [Google Scholar] [CrossRef] [Green Version]
- Hibbs, R.E.; Sulzenbacher, G.; Shi, J.; Talley, T.T.; Conrod, S.; Kem, W.R.; Taylor, P.; Marchot, P.; Bourne, Y. Structural determinants for interaction of partial agonists with acetylcholine binding protein and neuronal α7 nicotinic acetylcholine receptor. EMBO J. 2009, 28, 3040–3051. [Google Scholar] [CrossRef] [Green Version]
- Paris, D.; Beaulieu-Abdelahad, D.; Abdullah, L.; Bachmeier, C.; Ait-Ghezala, G.; Reed, J.; Verman, M.; Crawford, F.; Mullan, M. Anti-inflammatory activity of anatabine via inhibition of STAT3 phosphorylation. Eur. J. Pharmacol. 2013, 698, 145–153. [Google Scholar] [CrossRef]
- Paris, D.; Beaulieu-Abdelahad, D.; Bachmeier, C.; Reed, J.; Ait-Ghezala, G.; Bishop, A.; Chao, J.; Mathura, V.; Crawford, F.; Mullan, M. Anatabine lowers Alzheimer’s Aβ production in vitro and in vivo. Eur. J. Pharmacol. 2011, 670, 384–391. [Google Scholar] [CrossRef]
- Caine, S.B.; Collins, G.T.; Thomsen, M.; Wright, C., IV; Lanier, R.K.; Mello, N.K. Nicotine-like behavioral effects of the minor tobacco alkaloids nornicotine, anabaseine, and anatabine in male rodents. Exper. Clin. Psychopharmacol. 2014, 22, 9–22. [Google Scholar] [CrossRef]
- Levin, E.D.; Hao, I.; Burke, D.; Cauley, M.; Hall, B.J.; Rezvani, A.H. Effects of tobacco smoke constituents, anabasine and anatabine, on memory and attention in female rats. J. Psychopharmacol. 2014, 28, 915–922. [Google Scholar] [CrossRef]
- Ferguson, S.; Mouzon, B.; Paris, D.; Aponte, D.; Abdullah, L.; Stewart, W.; Mujllan, M.; Crawford, F. Acute or delayed treatment with anatabine improves spatial memory and reduces pathological sequelae at late time-points after repetitive mild traumatic brain injury. J. Neurotrauma 2017, 34, 1676–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, R.I.; Doyle, M.R.; Withey, S.L.; Bergman, J. Nicotinic effects of tobacco smoke constituents in nonhuman primates. Psychopharmacology 2016, 233, 1779–1789. [Google Scholar] [CrossRef] [PubMed]
- Verma, M.; Beaulieu-Abdelahad, A.-G.; Li, R.; Crawford, F.; Mullan, M.; Paris, D. Chronic anatabine treatment reduces Alzheimer’s disease (AD)-like pathology and improves socio-behavioral deficits in a transgenic mouse model of AD. PLoS ONE 2015, 10, e0128224. [Google Scholar] [CrossRef] [Green Version]
- Paris, D.; Beaulieu-Abdelahad, D.; Mullan, M.; Ait-Ghezala, G.; Mathura, V.; Bachmeier, C.; Crawford, F.; Mullan, M.J. Amelioration of experimental autoimmune encephalomyelitis by anatabine. PLoS ONE 2013, 8, e55392. [Google Scholar] [CrossRef] [Green Version]
- Caturegli, P.; De Remigis, A.; Ferlito, M.; Landek-Salgado, M.A.; Iwama, S.; Tzou, S.-C.; Ladenson, P.W. Anatabine ameliorates experimental autoimmune thyroiditis. Endocrinology 2012, 153, 4580–4587. [Google Scholar] [CrossRef] [Green Version]
- Mello, N.K.; Fivel, P.A.; Kohut, S.J.; Caine, S.B. Anatabine significantly decreases nicotine self-administration. Exper. Clin. Psychopharmacol. 2014, 22, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Clemens, K.J.; Caille, S.; Stinus, L.; Cador, M. The addition of five minor alkaloids increases nicotine-induced hyperactivity, sensitization and intravenous self-administration in rats. Intern. J. Neuropsychopharmacol. Addict. 2009, 12, 1355–1366. [Google Scholar] [CrossRef]
- Dwoskin, L.P.; Teng, L.; Buxton, S.T.; Ravard, A.; Deo, N.; Crooks, P.A. Minor alkaloids of tobacco release[3 H]dopamine from superfused rat striatal slices. Eur. J. Pharmacol. 1995, 276, 195–199. [Google Scholar] [CrossRef]
- Harris, A.C.; Tally, L.; Muelken, P.; Banal, A.; Schmidt, C.E.; Cao, Q.; LeSage, M.G. Effects of nicotine and minor tobacco alkaloids on intracranial self-stimulation in rats. Drug Alcohol Depend. 2015, 153, 330–334. [Google Scholar] [CrossRef] [Green Version]
- Xia, W.; Velijkovic, E.; Koshibu, K.; Peitsch, M.C.; Hoeng, J. Neurobehavioral effects of selected tobacco constituents in rodents following subchronic administration. Eur. J. Pharmacol. 2019, 865, 172809. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.L.; Stokoe, S.A.; Roberts, J.P.; Sathler, M.F.; Nip, K.A.; Shou, J.; Ko, K.; Tsunoda, S.; Kim, S. Co-activation of selective nicotinic acetylcholine receptors is required to reverse beta amyloid-induced Ca2+ hyperexcitation. Neurobiol. Aging 2019, 84, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Potasiewicz, A.; Golebiowska, J.; Popik, P.; Nikiforuk, A. Procognitive effects of varenicline in the animal model of schizophrenia depend on α4β2 and α7 nAChRs nicotinic acetylcholine receptors. J. Psychopharmacol. 2019, 33, 62–63. [Google Scholar] [CrossRef] [PubMed]
- Grierson, D.S.; Harris, M.; Husson, H.P. Synthesis and chemistry of 5,6-dihydropyridinium salt adducts. J. Am. Chem. Soc. 1980, 102, 1064–1082. [Google Scholar] [CrossRef]
- Tang, Y.; Zielinski, W.L.; Bigott, H.M. Separation of nicotine and nornicotine enantiomers via normal phase HPLC on derivatized cellulose chiral stationary phases. Chirality 1998, 10, 364–369. [Google Scholar] [CrossRef]
- Papke, R.L.; Thinschmidt, J. The correction of alpha7nicotinic acetylcholine receptor concentration-response relationships in Xenopus oocytes. Neurosci. Lett. 1998, 256, 163–166. [Google Scholar] [CrossRef]
- Pabreza, L.; Dhawan, S.; Kellar, K.J. [3H]cytisine binding to nicotinic cholinergic receptors in brain. Mol. Pharmacol. 1991, 39, 9–12. [Google Scholar]
- Kem, W.R.; Mahnir, V.M.; Prokai, L.; Papke, R.L.; Cao, X.; LeFrancois, S.; Wildeboer, K.; Prokai-Tatrai, K.; Porter-Papke, J.; Soti, F. Hydroxy metabolites of the Alzheimer’s drug candidate 3-[2,4-dimethoxy)benzylidene]-anabaseine dihydrochloride (GTS-21): Their molecular properties, interactions with brain nicotinic receptors and brain penetration. Mol. Pharmacol. 2004, 65, 56–67. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | IMax (%ACh) | P | EC50 (µM) | P | N |
|---|---|---|---|---|---|
| S-isoanatabine | 78.7 ± 1.9 | 0.02 * | 1.01 ± 0.33 | 0.12 | 4 |
| R-isoanatabine | 102 ± 5.7 | 0.31 ± 0.09 | 4 | ||
| S-anatabine | 43.2 ± 4.3 | 0.029 * | 2.65 ± 1.4 | 0.3 | 3 |
| R-anatabine | 25.0 ± 5.2 | 0.74 ± 0.21 | 8 | ||
| S-isoanatabine | 78.7 ± 1.9 | 0.0061 ŧŧ | 1.01 ± 0.33 | 0.36 | 4 |
| S-anatabine | 43.2 ± 4.3 | 2.65 ± 1.4 | 3 | ||
| R-isoanatabine | 102 ± 5.7 | <0.001 ŧŧŧ | 0.31 ± 0.09 | 0.099 | 4 |
| R-anatabine | 25.0 ± 5.2 | 0.74 ± 0.21 | 8 |
| Compound | Imax (% ACh) | P | EC50 (µM) | P | n |
|---|---|---|---|---|---|
| S-Isoanatabine | 81.9 ± 9.1 | 0.0007 *** | 52.3 ± 13 | 0.61 | 9 |
| R-Isoanatabine | 35.4 ± 5.3 | 61.3 ± 12 | 9 | ||
| S-Anatabine | 113 ± 20 | 0.788 | 69.7 ± 30 | 0.569 | 9 |
| R-Anatabine | 105 ± 16 | 51.8 ± 6.5 | 8 | ||
| S-Isoanatabine | 81.9 ± 9.1 | 0.193 | 52.3 ± 13 | 0.599 | 9 |
| S-Anatabine | 113 ± 20 | 69.7 ± 30 | 9 | ||
| R-Isoanatabine | 35.4 ± 5.3 | 0.003 ŧŧ | 61.3 ± 12 | 0.50 | 9 |
| R-Anatabine | 105 ± 16 | 51.8 ± 6.5 | 8 |
| Compound | Ki (nM) | P | Hill Slope | P | n |
|---|---|---|---|---|---|
| S-Isoanatabine | 108 ± 14 | 0.13 | 1.08 ± 0.15 | 0.40 | 12 |
| R-Isoanatabine | 136 ± 11 | 0.94 ± 0.05 | 12 | ||
| S-Anatabine | 249 ± 32 | 0.001 ** | 0.98 ± 0.09 | 0.70 | 12 |
| R-Anatabine | 119 ± 16 | 0.94 ± 0.06 | 12 | ||
| S-Isoanatabine | 108 ± 14 | 0.005 ** | 1.08 ± 0.15 | 0.59 | 12 |
| S-Anatabine | 249 ± 32 | 0.98 ± 0.09 | 12 | ||
| R-Isoanatabine | 136 ± 11 | 0.39 | 0.94 ± 0.05 | 0.99 | 12 |
| R-Anatabine | 119 ± 16 | 0.94 ± 0.06 | 12 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xing, H.; Keshwah, S.; Rouchaud, A.; Kem, W.R. A Pharmacological Comparison of Two Isomeric Nicotinic Receptor Agonists: The Marine Toxin Isoanatabine and the Tobacco Alkaloid Anatabine. Mar. Drugs 2020, 18, 106. https://doi.org/10.3390/md18020106
Xing H, Keshwah S, Rouchaud A, Kem WR. A Pharmacological Comparison of Two Isomeric Nicotinic Receptor Agonists: The Marine Toxin Isoanatabine and the Tobacco Alkaloid Anatabine. Marine Drugs. 2020; 18(2):106. https://doi.org/10.3390/md18020106
Chicago/Turabian StyleXing, Hong, Sunil Keshwah, Anne Rouchaud, and William R. Kem. 2020. "A Pharmacological Comparison of Two Isomeric Nicotinic Receptor Agonists: The Marine Toxin Isoanatabine and the Tobacco Alkaloid Anatabine" Marine Drugs 18, no. 2: 106. https://doi.org/10.3390/md18020106