Application-Oriented Marine Isomerases in Biocatalysis


Institute of Biomolecular Chemistry, National Research Council, Via Campi Flegrei, 34, 80078 Pozzuoli, Italy
Mar. Drugs 2020, 18(11), 580; https://doi.org/10.3390/md18110580
Submission received: 3 November 2020
/
Revised: 16 November 2020
/
Accepted: 20 November 2020
/
Published: 21 November 2020
(This article belongs to the Special Issue Marine Enzymes: Sources, Biochemistry and Bioprocesses for Marine Biotechnology – II)
Abstract
:The class EC 5.xx, a group of enzymes that interconvert optical, geometric, or positional isomers are interesting biocatalysts for the synthesis of pharmaceuticals and pharmaceutical intermediates. This class, named “isomerases,” can transform cheap biomolecules into expensive isomers with suitable stereochemistry useful in synthetic medicinal chemistry, and interesting cases of production of l-ribose, d-psicose, lactulose, and d-phenylalanine are known. However, in two published reports about potential biocatalysts of marine origin, isomerases are hardly mentioned. Therefore, it is of interest to deepen the knowledge of these biocatalysts from the marine environment with this specialized in-depth analysis conducted using a literature search without time limit constraints. In this review, the focus is dedicated mainly to example applications in biocatalysis that are not numerous confirming the general view previously reported. However, from this overall literature analysis, curiosity-driven scientific interest for marine isomerases seems to have been long-standing. However, the major fields in which application examples are framed are placed at the cutting edge of current biotechnological development. Since these enzymes can offer properties of industrial interest, this will act as a promoter for future studies of marine-originating isomerases in applied biocatalysis.
1. Introduction
In 2010, one of the first comprehensive review articles about enzymes of marine origin especially suitable for future applications in biocatalysis reported an account of the knowledge in the field. As stated in the conclusion, the enormous pool of marine biodiversity is an excellent natural reservoir for acquiring an inventory of enzymes that is one of the focal points of the potential of blue biotechnology. The importance of the examples reported, picked up from the different classes of enzymes, supported the view that the marine environment is to be considered as an additional source of new enzymes for the biocatalysis. Important examples are among oxidoreductases and carbohydrate-active enzymes. Their novel chemical and stereochemical properties are included in the list of useful habitat-related characteristics possessed by these enzymes, adding value to the often-observed usual resistance of these proteins to high salt concentration and/or organic solvent. Other enzymes, characterized by a potent chemical action on nonactivated carbon atoms are also of extreme interest. Details on the characteristics of other marine representatives belonging to other classes of enzymes (lipid active hydrolases, novel esterases, and other hydrolytic activities) further supported the conclusion [1]. Isomerases are included only with a few cases, in particular, an alanine racemase from the hepatopancreas of the black tiger prawn, Penaeus mondon [2] and a fatty acid isomerase from marine alga Ptilota filicina [3].
Moreover, in a subsequent review, dated 2016 [4], on the advances in marine enzymes used in food and pharmaceutical applications, although the list of enzymes discussed included oxidoreductases, hydrolases, transferases, ligases, lyases, and explicitly mentioned isomerases, no details on examples of the latter class were reported.
Among the seven major groups of the international classification of enzymes, the class EC 5.xx and subclasses include isomerases, a group of biocatalysts that interconvert optical, geometric, or positional isomers. These enzymes, involved in the central metabolism of most living organisms, also have trivial names, such as epimerase, racemase, cis-trans isomerases, cycloisomerase, and tautomerase, according to the specific reaction types they catalyze. One of the illustrative examples is triose phosphate isomerase (5.3.1.1) that catalyzes the interconversion of dihydroxyacetone phosphate and d-glyceraldehyde 3-phosphate. In practical examples reported for the applications of isomerase in the field of biocatalysis, some important molecules are prepared. In a recent review for the biocatalyzed synthesis of pharmaceuticals and pharmaceutical intermediates [5], the cases reported concern the transformation of cheap biomolecules into expensive isomers possessing suitable stereochemistry for applications in synthetic medicinal chemistry, as described for production of l-ribose (l-arabinose isomerase from Paenibacillus polymyxa), d-psicose (cascade reactions with thermophilic enzymes), lactulose (cellobiose 2-epimerase from Caldicellulosiruptor saccharolyticus), and d-phenylalanine (cascade reaction using d-succinylase from Cupriavidus and N-succinylamino acid racemase from Geobacillus stearothermophilus). In addition, the use of racemases at the industrial level for chiral resolution of racemates is mentioned. In these processes, the racemases are usually part of a cascade reaction and other enzymes are responsible for the chiral resolution [6]. In their conclusions, the authors recognized the widespread distribution of isomerases among species with high biodiversity, indicating that isomerases suitable for many synthetic problems can be identified in particular habitats.
Economically successful biocatalytic processes depend on robust performance, high selectivity, increased stability, increased activity, and broader availability of biocatalysts. Marine-originating enzymes can offer most of these features. Environmental concern is the main factor to drive the growth of the industrial enzymes market globally (projected to reach USD 8.7 billion by 2026). In this, growing attention to green technologies for the conversion of biomass and saccharification of carbohydrate polymers isomerases are included for starch and dairy industries.
The current interest for application-oriented isomerases in scientific literature is demonstrated in the field of rare sugars applied as sweeteners and building blocks as in the study of ribose-5-phosphate isomerase of an Ochrobactrum sp. [7] to increase reaction rate in isomerizing l-rhamnose to l-rhamnulose.
Prompted by this importance and by the rare mentions of biocatalysts of marine origin in specialized reviews, it seems of interest to deepen the knowledge of isomerases from this habitat dedicating a specialized in-depth analysis to them. Indeed, during this literature search, it became evident that curiosity-driven scientific interest has been long-standing as in the case of phosphoglucose isomerases in marine mollusks [8]. This review is dedicated mainly to examples of the applications of marine isomerases, discussing general scientific interest from a historical perspective and in tabulated forms.
2. Literature Search
The search for articles in this review has been preliminarily conducted using the words “isomerase” and “marine” in titles, abstracts, and keywords without interval time limit in the Science Direct database with access to 3800 scientific journals in major scientific disciplines. However, the limited range of articles retrieved (31 hits) prompted us to use the more generic PubMed database with the same words thus retrieving 172 hits. Besides, trivial names of these enzymes (as epimerase, racemase, cis-trans isomerases, cycloisomerase, and tautomerase) were also coupled with “marine” in an accessory search using Web of Science database (Table 1). Science alert mailing lists were used, up to the manuscript submission to update the analysis with the most recent results. In this review, an analysis of reports about enzymatic activities already oriented to applications is firstly compiled in the central Section 3. In Section 4, Section 5 and Section 6, tabulated lists of the remaining articles according to types of molecules on which these isomerases can catalyze their reactions are discussed. Isomerases acting on carbohydrate molecules (Table 2) [9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38] are listed in the first of three tables following with lipids (Table 3) [39,40,41,42,43,44,45,46] and amino acids and peptides (Table 4) [47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74]. Each chronologically ordered line reports the name of the enzyme, as indicated in the reference, the reaction catalyzed, and a short description of the intent and scientific field of the work.
3. Application-Oriented Biocatalysts
A few applicative examples of works using marine isomerase are collected in this paragraph.
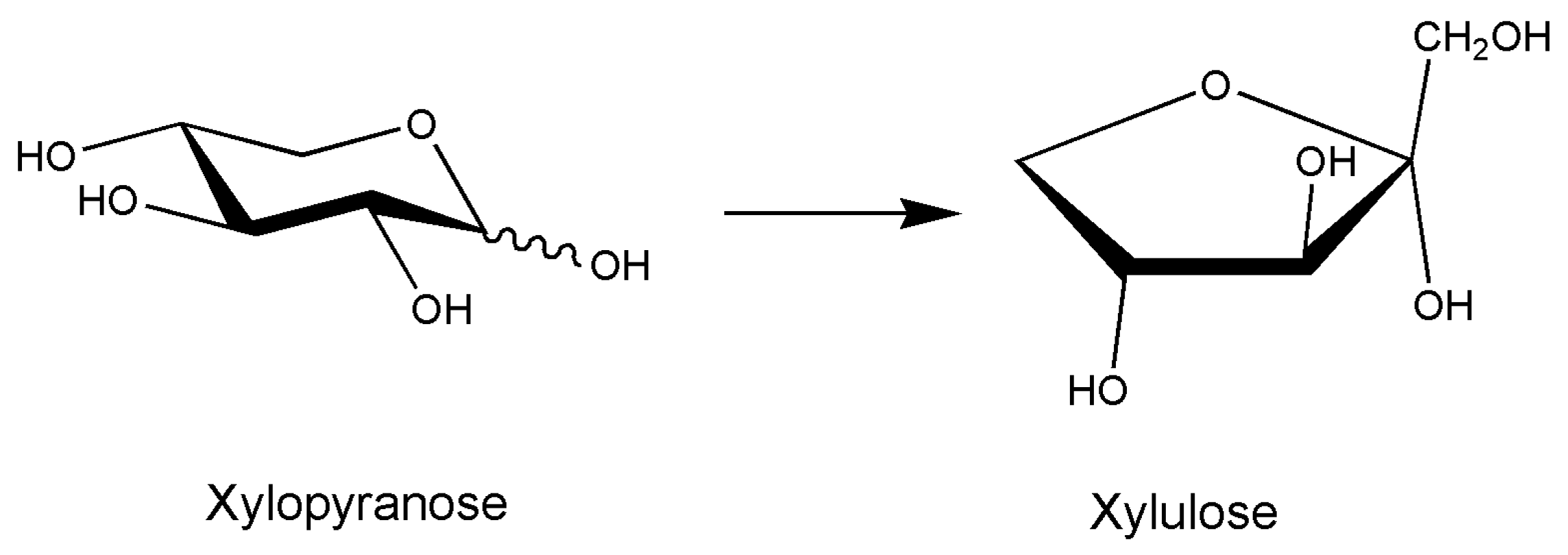
A gene encoding for d-xylose isomerase from a marine bacterium, Vibrio sp. strain XY-214, has been expressed in E. coli, and the production of d-xylulose from β-1,3-xylan was carried out. This paper concerns the growing attention to green technologies for the conversion of biomass and saccharification of carbohydrate polymers, in particular, the polysaccharide β-1,3-xylan of the invasive green alga Caulerpa taxifolia. Marine Vibrio sp. strain can grow on β-1,3-xylan as a sole carbon source; the rationale for the work is based on the synergistic action of two types of enzymes enabling the complete degradation of β-1,3-xylan to d-xylose, i.e., 1,3-β- d-xylan xylanohydrolase and a β-1,3-xylosidase. d-xylulose is then formed by the marine d-xylose isomerase. d-xylulose can be used for ethanol fermentation thus allowing the use of the algal polymer β-1,3-xylan of C. taxifolia as a base for ethanol production. The article is a preliminary study for a possible real application of the total saccharification of the polymer. The work has been conducted by purified enzymes, and 2.62 g/L reducing sugars were released by the action of the two β-1,3-xylan degrading activities, the subsequent isomerization of d-xylose thus producing d-xylulose. Time-course experiments analyzing reaction mixtures by HPLC were reported, and different reaction conditions were analyzed also in presence of sodium tetraborate for possible complexes with xylulose-borate shifting the equilibrium [75].
Yarrowia lipolytica is a marine microorganism of industrial interest for the physiological ability to utilize different substrates for growth (polyalcohols, organic acids, and long-chain hydrocarbons). In a recent short communication [76], the isomaltulose production using an engineered Yarrowia lipolytica strain is reported. Sucrose isomerase catalyzes the enzymatic rearrangement of the α-1,2 linkage between glucose and fructose to an α-1,6 linkage (producing isomaltulose) or α-1,4 linkage (producing trehalulose). Marine origin examples of sucrose isomerase and its use for biological isomaltulose production were not known up to a review of 2014 [77]. In fact, the sucrose isomerase overexpressed from Pantoea dispersa but the high and efficient process of isomaltulose production was based on enzyme production and enzymatic catalysis during fermentation, thus reducing costs and simplifying the bioprocess. The maximum isomaltulose production was 572.1 g/L, with a yield of 0.96 g/g of sucrose.
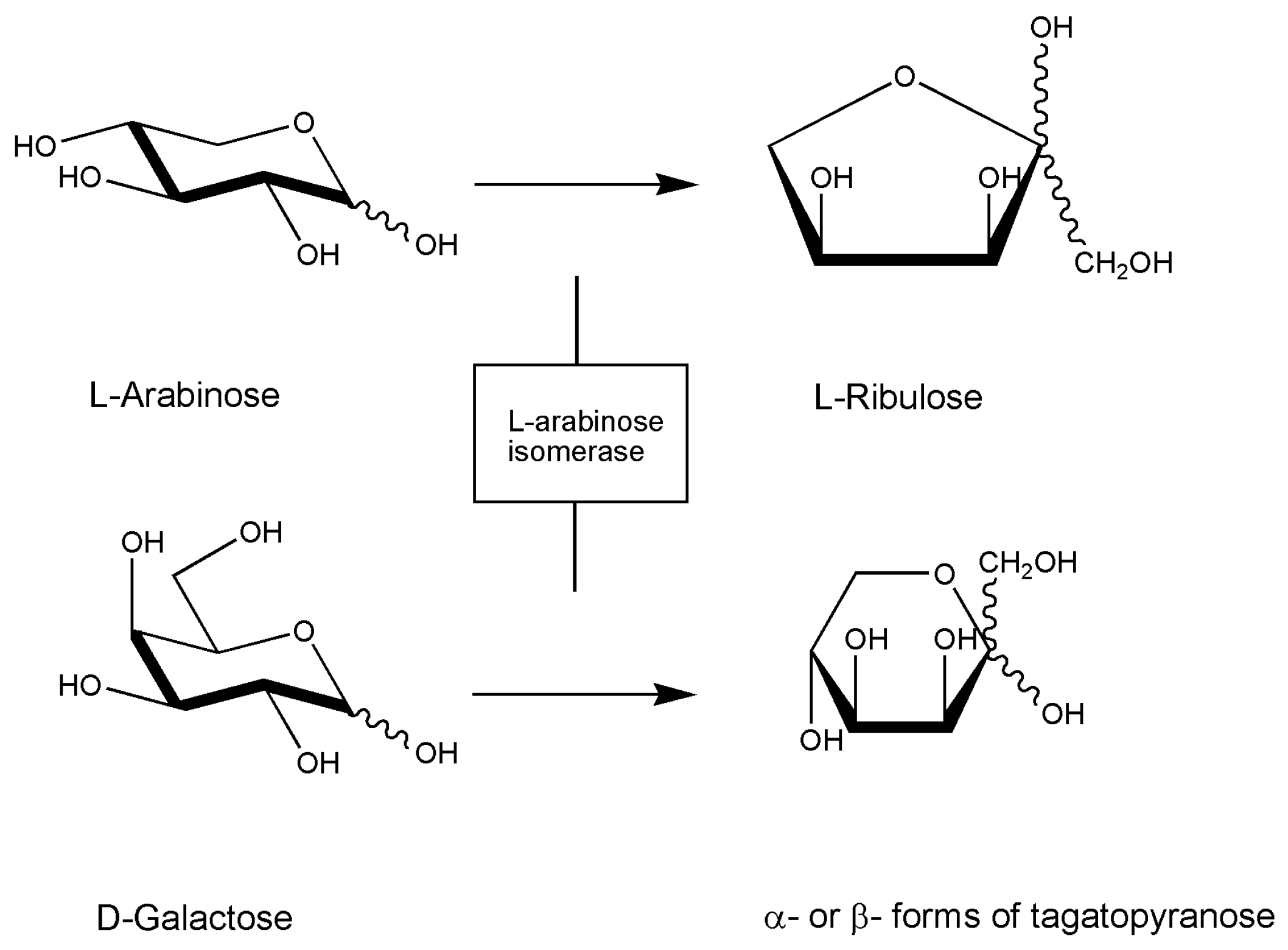
A very recent report demonstrates the use of l-arabinose isomerase for production of D-tagatose, a rare sugar of importance in the food industry that has been approved as GRAS drug by the US Food and Drug Administration and used as a substitute for sucrose in low-calorie sweeteners. As in the above case, although the enzyme is from Lactobacillus sakei 23K and converts d-galactose from agar into d-tagatose, it is worth to mention this research effort here to show the interest for the profitable application of red algae carbohydrate polymers as a substrate for d-tagatose production (Figure 1) [78].
Ribose-5-phosphate isomerase is another enzyme of interest in the field of rare sugars that are used as sweeteners and for production of interesting building blocks for fine chemistry as reported in the study of ribose-5-phosphate isomerase from an Ochrobactrum sp. [7] to increase reaction rate in isomerizing l-rhamnose to l-rhamnulose already mentioned. Although the microorganism was isolated from soil samples, marine examples are also known and could be potential alternatives. Substrate specificity and reaction properties were explored, the results encouraged for the application of ribose-5-phosphate isomerase as a biocatalyst in preparation of rare sugars.
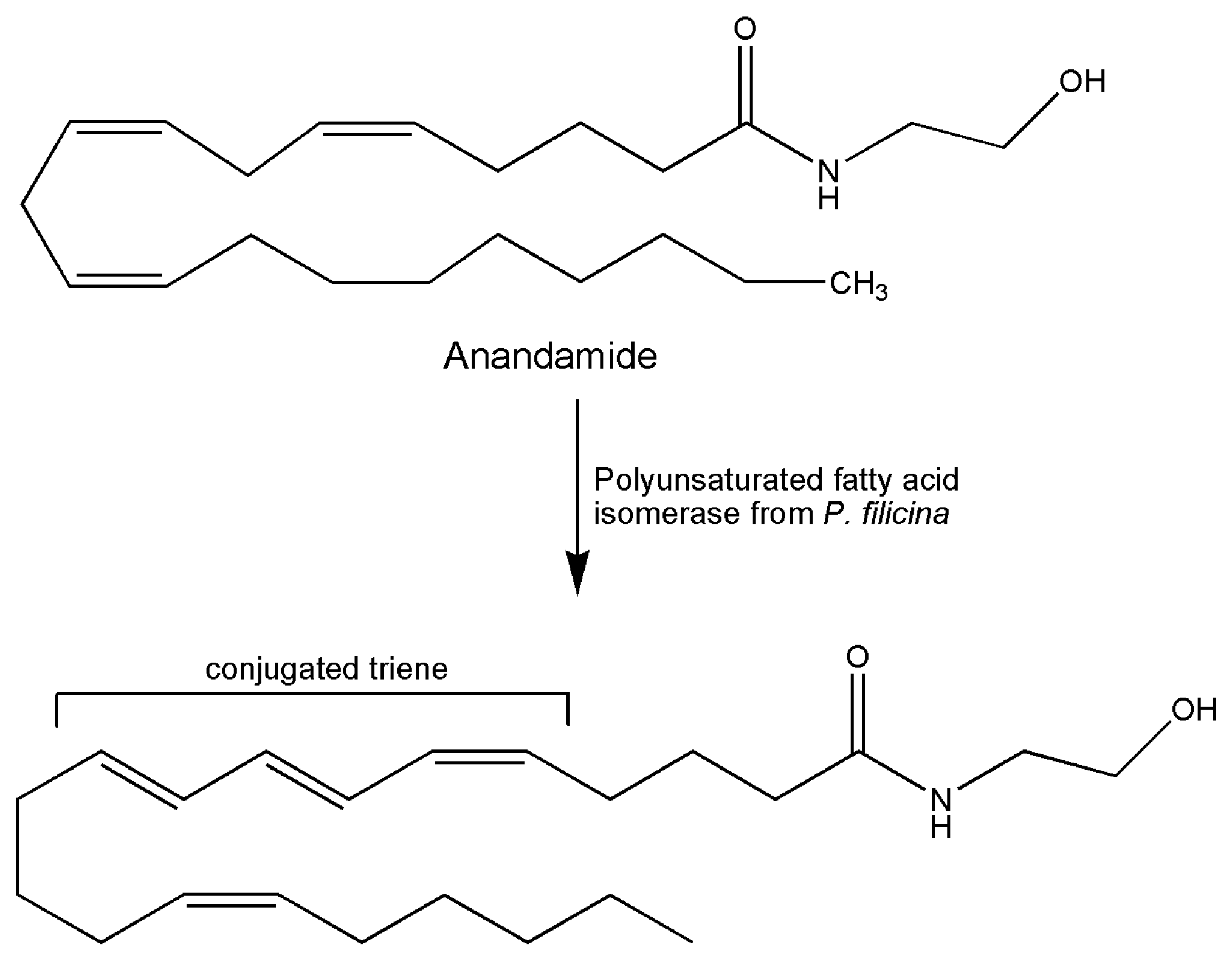
A report already present in literature in 1994 was the first describing a novel isomerase for the biosynthesis of conjugated triene-containing fatty acids in the red alga Ptilota filicina [79]. At the time, in fact, many hypotheses and studies on the biosynthetic pathway of conjugated polyenes in marine organisms were already present in literature, coming from studies centered on natural products of marine origin. The main interest is focused on pharmacological studies indicating a role for these bioactive molecules in the treatment of tumors, against weight gain, and as enhancers of the immune system. In particular, the biosynthesis of a conjugated triene (4,5Z,7E,9E,14Z,17Z)-eicosapentaenoic acid from eicosapentaenoic acid was indicated. The product is present among natural products of the red alga Ptilota filicina. The enzyme was isolated from alga tissues and assayed with arachidonic acid forming a triene structure, evidenced by UV absorption. The product of arachidonate incubation was also identified as the corresponding conjugated triene metabolite. A substrate specificity investigation revealed that the eicosapentaenoic structure was the best substrate for the enzyme. Incorporation of deuterium at C11 position of arachidonate was demonstrated by 1H NMR spectroscopy and mass spectrometry for the reaction conducted in the deuterated buffer. Intramolecular and intermolecular hydrogen transfers using stereospecifically deuterated substrates and oxygen sensibility of reaction were also studied. These authors were able to show that unlike the well-characterized aerobic reaction of lipoxygenases, molecular oxygen was not required by their isomerase with no net desaturation occurring during the reaction, thus providing useful insights for the use of these biocatalysts as tools for the synthesis of novel compounds. Later in time, the P. filicina enzyme was purified to electrophoretic homogeneity, and the cloning and functional expression including the study of other important characteristics such as molecular weight, subunit structure, and glycosylation were reported [3]. The ability of this enzyme in the isomerization of methylene interrupted olefins led the authors to try the reaction with anandamide, the well-known N-ethanolamide of arachidonic acid, the first endogenous ligand of cannabinoid receptor. The conjugated triene anandamide product was shown to possess high-affinity binding for the receptor [80] (Figure 2). A 33% yield was obtained in preparative experiments for the full spectroscopic chemical characterization of the reaction product. As the same authors speculated, they were able to show the use of these marine enzymes in the synthetic production of novel compounds for pharmacological probes. The same group already studied, in 1991, the oxylipin metabolism. In this work, the conversion of arachidonic acid into the vicinal diol fatty acid 12R,13S-dihydroxy-5Z,8Z,10E,14Z-eicosatetraenoic acid by an acetone powder of the marine red alga, Gracilariopsis lemaneiformis, occurred via intermediate formation of hydroperoxide 12S-hydroperoxy-5Z,8Z,10E,14Z-eicosatetraenoic acid, postulating the existence of a hydroperoxide isomerase in this red alga. The broad substrate specificity and the high stereospecificity of the product formed in the step of oxygen insertion were of interest in the application in biocatalysis [81].
By studying how cultured fish cells derived from turbot (Scophthalmus maximus), gilthead seabream (Sparus aurata), and Atlantic salmon (Salmo salar) metabolize all-cis octadecapentaenoic acid, some authors discovered the action of an isomerase acting on all-cis 18:5n-3 acids producing 2-trans 18:5n-3 acids thought to be common intermediates in the β-oxidation of these acids by marine animals [82]. Similar isomerases acting on double bonds of different compounds have been hypothesized in a study on biodegradation of alkenones and related compounds of the marine microalgae Emiliania huxleyi by microbial mats collected in large ponds. Among products, authors found cis/trans or trans/cis alkene and alkenone isomers and suggested that the formation of these isomeric compounds is likely due to extracellular bacterial cis/trans isomerases [83].
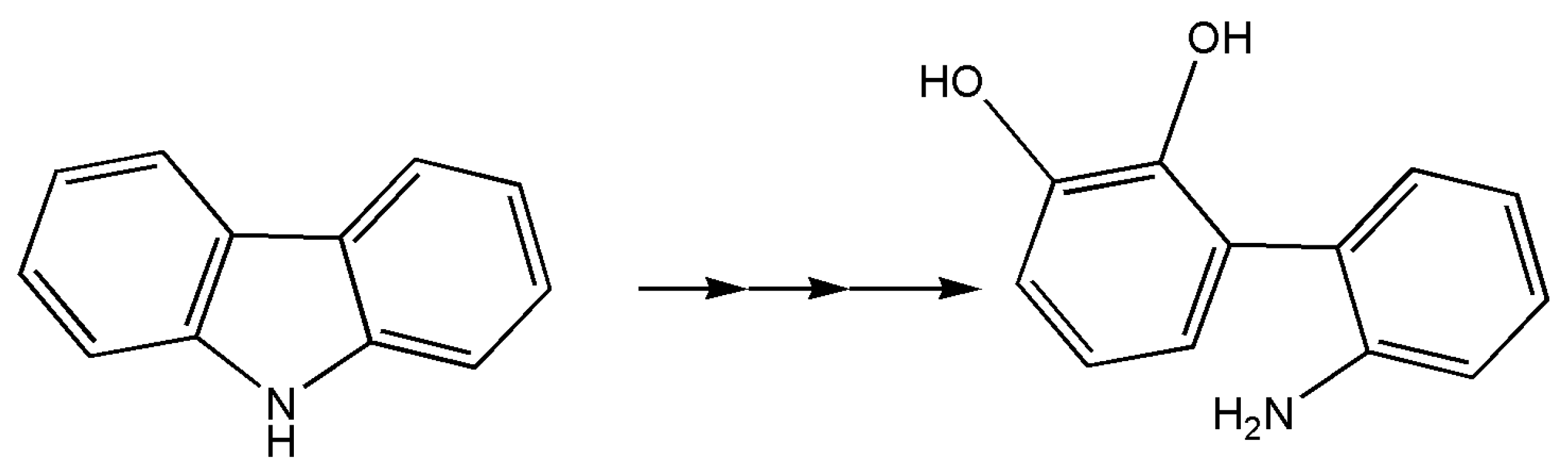
In a study centered on searching for marine microorganisms capable of carbazole remediation schematically represented in Figure 3, the marine bacterium Neptuniibacter sp. strain CAR-SF has been found utilizing carbazole as its sole carbon and nitrogen sources [84]. Among the enzymes involved in the degradation pathway, 4-oxalocrotonate tautomerase (and others) is indicated, and Escherichia coli cells transformed in this work required ferredoxin and ferredoxin reductase for necessary initial dioxygenation of carbazole. The authors indicated that this was the first report of genes involved in carbazole degradation isolated from a marine bacterium, however, only the conversion product of carbazole through dioxygenation by dioxygenase was found (2′-aminobiphenyl-2,3-diol).
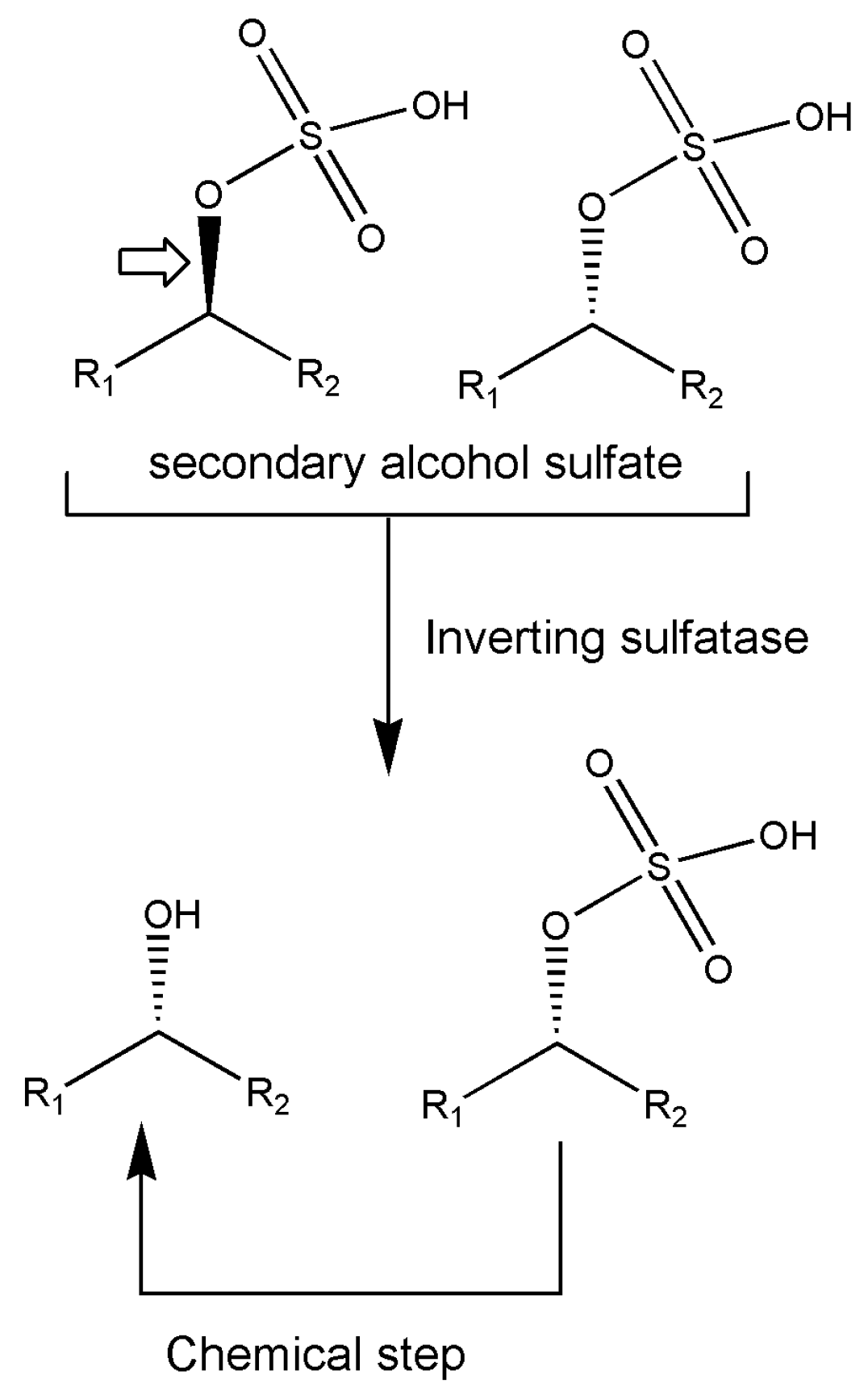
Various enzymes have been analyzed in an interesting report [85] on strategies for the deracemization of a racemate into a single stereoisomeric product; these include mandelate racemase, lactate racemase, or alkyl sulfatases from the actinomycete Rhodococcus ruber DSM 44541, the marine planctomycete Rhodopirellula baltica DSM 10,527, and others, known to possess a rich inorganic sulfur metabolism. Although not belonging to isomerases, it is worth mentioning that alkyl sulfatases display not only enantioselectivity but also stereoselectivity for retention or inversion of the configuration of the formed product during sulfate hydrolysis and these authors report about a scheme devised to produce single stereoisomer from racemate. The (±)-sec-alkyl sulfate ester is subjected to inverting alkyl sulfatase producing a mixture of hydrolyzed ester with the same configuration of the remaining unreacted ester. The latter is then hydrolyzed in a chemical step with retention of configuration producing the alcohol in this enantioconvergent process (Figure 4). The importance of marine-originating biocatalysts is clearly assessed by this example.
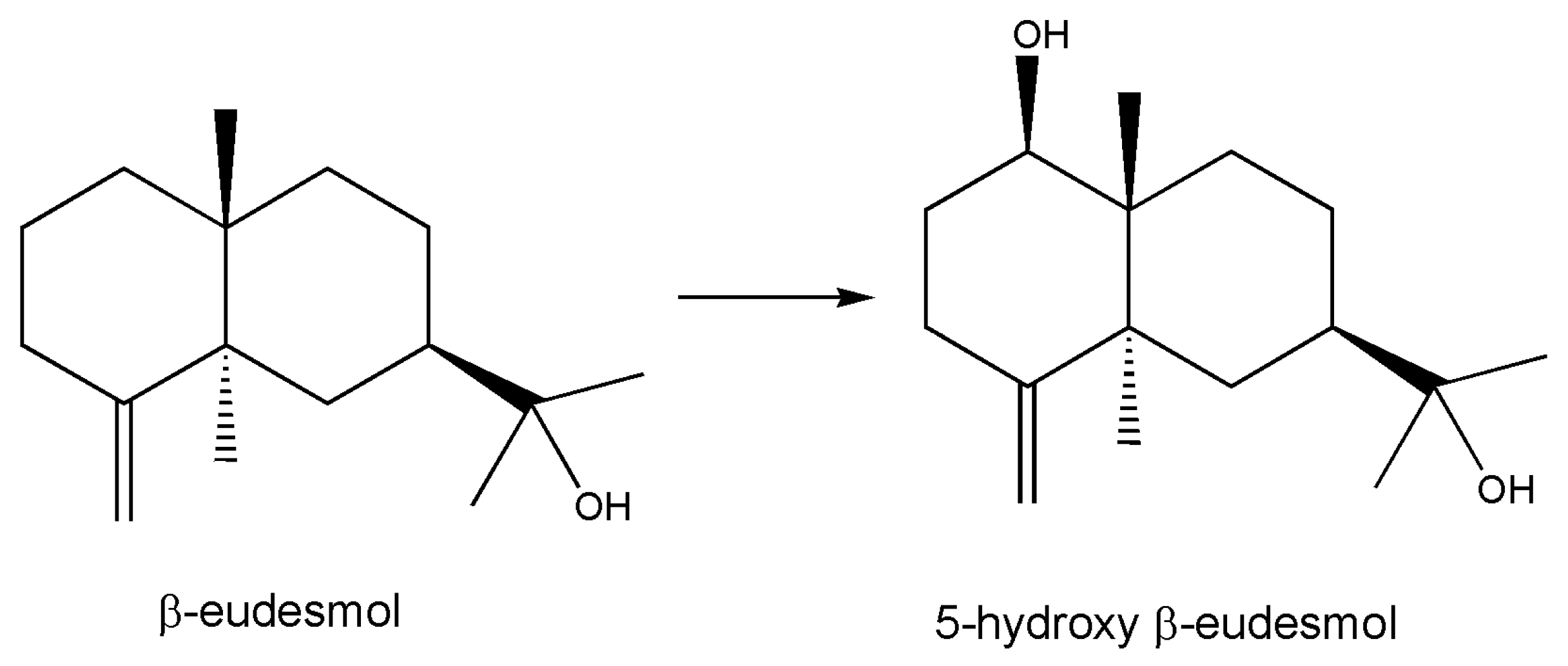
Cytochromes P450 are important biocatalysts performing hydroxylation reactions in regio- and stereospecific manners operating on inactive carbon atoms; they are useful for the bioassisted synthesis of organic molecules. In an interesting paper [86], authors constructed a fusion protein of a peptidyl-prolyl cis-trans isomerase isolated from the hyperthermophilic archaeon Thermococcus sp. with the cytochrome P450 BM3 derived from Bacillus megaterium and evaluate its stability in E. coli cells in a series of bioconversion experiments with various substituted naphthalenes. It is known that peptidyl-prolyl cis-trans isomerases catalyze the cis-trans isomerization of the proline imide bond in polypeptides, which may affect the folding rate of proteins. This fusion protein exists as the predominant soluble protein and more stable than the unfused P450. Various substituted naphthalenes were converted to their monohydroxylated derivatives, and the reaction was also tested on a sesquiterpene (Figure 5) that has physiological functions such as β-eudesmol that was found to be hydroxylated in a regio- and stereo-specific manner.
4. Marine Isomerases Acting on Sugar Molecules
In Table 2, the articles [9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38] found on isomerases acting on carbohydrate pathways are briefly listed. As cited in the first entry about Pelvetia canaliculata in 1973 [9], the scientific interest was early generally present in literature since 1956 in investigations conducted on different organisms. For an algal polymannuronic-5-epimerase, converting polymannuronic acid to a mixed polymer containing guluronic acid, the preparation of ammonium sulfate precipitation was reported [9]. Scientific interests for the articles listed in Table 2 and experimental methodologies used reflected the successes in biochemistry, molecular biology, and genetics achieved during the last century. All these listed articles did not contain applicative results that are detailed above in the paragraph on application-oriented biocatalysts. Very often, a basic study of the marine biocatalyst is reported as in the case [28] of xylose isomerase from Fulvimarina pelagi. It was identified by sequence analysis of the F. pelagi genome, i.e., PCR amplified, cloned, and expressed in E. coli, while the aim of the work is framed into the field of biofuel production (Figure 6).
In some cases, the interest of the article is focused on biomedical field as for the two reports [32,33] for the study of allergen function of triosephosphate isomerases from Octopus fangsiao and freshwater crayfish Procambarus clarkii. The consumption of seafood products, in fact, can be related to the high frequency of food-induced immune responses, and these studies are important to develop therapeutic and diagnostic approaches to these issues. In the case of freshwater crayfish Procambarus clarkii, the increased production and consumption can result, in fact, in allergic reactions, including life-threatening anaphylaxis. Another interesting article showing modern scientific interest is the one related to GDP-l-galactose mutase in Marinactinospora thermotolerans [34], an article dealing with a rare and interesting molecule, l-Galf, hardly found in the environment; the biosynthetic action and roles and relevant enzymes acting on l-Galf are of interest as this moiety is inserted in an aminonucleoside antibiotic possessing medicinal potential.
5. Marine Isomerases Acting on Lipid Molecules
Table 3 contains a few listed articles concerning isomerases acting on lipid molecules. The scientific interest was present as early as the beginning of the 1990s as indicated by the genetic study [39] related to the steroidogenic enzyme involved in the production of 17α-hydroxyprogesterone in the trout Oncorhynchus mykiss. The enzyme, produced after expression of cDNA in COS-1 cells, was capable of converting dehydroepiandrosterone to androstenedione. The article, aimed at the finding of necessary probes for identification of steroidogenic enzyme genes in fish species, is in the frame of investigations on the molecular evolution of vertebrate steroidogenic enzymes. As for industrial production of carotenoid pigments such as β-carotene and astaxanthin utilized as food or feed supplements, the interest for the marine bacterium Agrobacterium aurantiacum is well documented [40] in the reported review discussing the advances achieved in the field of metabolic engineering for the microbial production of these compounds at the end of 1990s’. In 1999 [42], from the same bacterium, a gene cluster was introduced into E. coli to produce astaxanthin at a value 50 times higher than previously achieved. All the aspects about the enzymes of interest from the marine alga Ptilota filicina were already discussed in the previous paragraph, and recent interest for Dunaliella salina [46] in this field is also present; the latter study provides an insight for induction of β-carotene production in optimized cultivation systems. The case of Schizochytrium, the marine fungus producing significant amounts of docosahexaenoic acid (DHA) is of very interest due to positive effects on atherosclerosis, hypertriglyceridemia, hypertension, and cancers of the compound. The paper listed reports of the mechanisms of DHA biosynthesis in Schizochytrium constructing and analyzing cDNA library with the possible interesting prospect for new tools to engineer the production of PUFAs [43]. The sterol composition and the related biosynthetic genes were also studied in Chromera velia [44], a marine alveolate although the work is framed in the field of basic studies about sterol composition for deriving chemotaxonomic relationships.
6. Marine Isomerases Acting on Amino Acids and Peptides
In Table 4, many articles reported concern alanine racemase found in marine organisms. This enzyme, previously discovered in 1951 in other sources was found [47] in the bivalve Corbicula japonica in 1985. The authors partially purified the protein evaluating biochemical properties in relation to those of bacterial origin and linked enzyme role to the osmoregulation in these marine organisms. Many other works related to this enzyme are present [2,48,51,52,53] always concerning the osmoregulation action, as in the crayfish Procambarus clarkii and in the hepatopancreas of black-tiger prawn, Penaeus monodon, up to recent interest for salinipeptins, a group of natural peptides in halotolerant Streptomyces isolated from the Great Salt Lake. Salinipeptins, natural products containing D-amino acids, are subjected to extensive enzymatic post-translational modifications during biogenesis [72]. They are substrates of potentially new epimerases of interest during these bioprocesses. A study related to genomic analysis of serine racemase is also found [61]; d-serine, besides frequently found in the bacterial cell walls, lipopeptides and siderophores, also exists as a free molecule in the marine environment with Roseobacter litoralis being a special producer. The cases related to disulfide isomerase are also numerous in this Table. They are mostly related to the studies on conotoxins or conopeptides, disulfide-rich peptides found in cone snails that found application in research and possible therapy. The studies mainly based on genetics focus the attention on post-translational reactions catalyzed by these enzymes for diversification of conopeptides structures and folding. Other interests are related to mechanisms of the immune response. Another topic of great interest among these isomerases is related to the cyclophilins (peptidyl-prolyl cis-trans isomerase, PPIase activity) that catalyze the isomerization of peptide bonds from trans to cis at proline residues and facilitate protein folding. Their expression is usually enhanced in response to inflammation or malignancy and are involved in functions related to cell metabolism and energy homeostasis and are of therapeutic importance for these and other actions (antifungal, antiviral, and antioxidant activities); they are also of economic importance in oysters cultivation for their involvement in the oyster immune response against infections of Crassostrea ariakensis by pathogen rickettsia-like organisms [66].
7. Other Enzymes
There are other reports on different enzymatic activities belonging to isomerases that do not fit well into the sections above and are mentioned in this paragraph. The first is an interesting review on chitin metabolism in the marine environment [87]. Authors hypothesized the presence of a mutase in the chitin catabolic cascade, in a more complex system with respect to the usually simplistic accepted hydrolytic pathway based on a chitinase producing the disaccharide N,N′-diacetylchitobiose, and on a beta-N-acetylglucosaminidase producing the final product GlcNAc. The mutase, as described in this review, could represent the activity that converts GlcNAc-1-P, generated from small chitin oligosaccharides and chitobiose for entering the cell membrane, into the 6-P.
Various other articles are present dealing with dopachrome tautomerases involved in the final step of the enzymically regulated melanin biogenesis for the conversion of dopachrome into dihydroxyindoles. In marine organisms, especially in bivalves, the enzymes involved in the biogenesis of melanin are recognized as the general class of phenoloxidases while less is known about the existence and functional role of dopachrome tautomerase genes [88] in mollusk or other organisms [89,90]. However, this field of investigation is quite active due to the role of D-dopachrome tautomerase as cytokine, member of the macrophage migration inhibitory factor protein superfamily. They are associated to important physiological processes such as cell recruitment and migration, tumorigenesis and cancer progress, and inflammatory and autoimmune diseases. Many of these studies on immune system of marine fish may contribute to develop better disease management strategies for fish aquaculture as for Japanese sea bass (Lateolabrax japonicus) [91] or for the clam Ruditapes philippinarum [92].
8. Conclusions
The study of biocatalysts on a global scale from marine environment is just starting and possesses huge potential for the development of applications with industrial benefits due to marine biological diversity and to the specificity of biological marine metabolisms. This knowledge constitutes the core of marine biotechnology and only a deep understanding of the complexity of this ecosystem will enable human beings to protect the oceans and organisms populating them and pave the way for the sustainable exploitation of marine resources. Among the many fields covered that are highly relevant to societal challenges the biorefinery value-chain, food industries and fine chemicals are included among others. However, many challenges remain, generally speaking a deep comprehension of the “marine biotechnology landscape” and a multidisciplinary approach, in education and training [93].
In two comprehensive reports on examples of the application of marine-originating biocatalysts in 2010 and 2016 abovementioned [1,4], marine isomerases were hardly discussed although other classes of enzymes cited are used in food and pharmaceutical applications. After the analysis of literature articles, a first undoubted conclusion of this in-depth review is that curiosity-driven scientific interest for these enzymes seems to be present for a long time. Most of the literature found, tabulated according to the type of molecules on which these enzymes act, indicated a general scientific interest in historical perspective in different fields. As more recent examples, the biomedical field for allergen function of triosephosphate isomerases for seafood consumption, or efforts for the elucidation of the biosynthetic action of GDP-L-galactose mutase acting on interesting and rare L-Galf, must be mentioned. As for isomerases acting on lipid molecules, both basic interest for investigations on the molecular evolution of vertebrate steroidogenic enzymes or more oriented studies for carotenoid pigments production, are present. Similar situation for isomerases acting on protein molecules was noted, e.g., of alanine racemases, in the studies related to the role of the osmoregulation in marine organisms and for new epimerases catalyzing interesting bioprocesses during post-translational modifications of natural peptides known as salinipeptins.
On the other hand, application-oriented examples of marine isomerases already applied in biocatalysis are a few confirming the general result reported in previously published reviews [1,4]. However, major fields in which these few papers are framed are depicted in a better manner in this review. Works are placed at the cutting edge of biotechnological development such as the conversion of biomasses and saccharification of carbohydrate polymers (d-xylose isomerase), in biomedicine and nutraceuticals (isomaltulose production, l-arabinose isomerase for production of d-tagatose and ribose-5-phosphate isomerase), and in bioremediation field (cytochromes P450, carbazole remediation, etc.). Therefore, despite the scarcity of direct applicative examples found, novel stability features and chemical/stereochemical properties found in general examples of marine biocatalysts will be present in the numerous studied isomerases as well. These enzymes in fact can offer properties related to the habitat, which are greatly appreciated under a general biotechnological perspective. As last conclusion, it can be said that these properties will surely act as a promoter for future studies of these marine-originating isomerases in applied biocatalysis.
Funding
This research received no external funding.
Conflicts of Interest
The author declares no conflict of interest
References
- Trincone, A. Potential biocatalysts originating from sea environments. J. Mol. Catal. B Enzym. 2010, 66, 241–256. [Google Scholar] [CrossRef]
- Uo, T.; Ueda, M.; Nishiyama, T.; Yoshimura, T.; Esaki, N. Purification and characterization of alanine racemase from hepatopancreas of black-tiger prawn, Penaeus monodon. J. Mol. Catal. B Enzym. 2001, 12, 137–144. [Google Scholar] [CrossRef]
- Zheng, W.; Wise, M.L.; Wyrick, A.; Metz, J.G.; Yuan, L.; Gerwick, W.H. Polyenoic fatty acid isomerase from the marine alga Ptilota filicina: Protein characterization and functional expression of the cloned cDNA. Arch. Biochem. Biophys. 2002, 401, 11–20. [Google Scholar] [CrossRef]
- Lima, R.N.; Porto, A.L.M. Recent advances in marine enzymes for biotechnological processes. In Marine Enzymes Biotechnology: Production and Industrial Applications, Part I—Production of Enzymes; Kim, S.-K., Toldra, F., Eds.; Academic Press Elsevier: New York, NY, USA, 2016; pp. 153–192. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, H.; Ang, E.L.; Zhao, H. Biocatalysis for the synthesis of pharmaceuticals and pharmaceutical intermediates. Bioorg. Med. Chem. 2018, 26, 1275–1284. [Google Scholar] [CrossRef]
- Femmer, C.; Bechtold, M.; Roberts, T.M.; Panke, S. Exploiting racemases. Appl. Microbiol. Biotechnol. 2016, 100, 7423–7436. [Google Scholar] [CrossRef] [PubMed]
- Ju, X.; Xu, X.; Shen, M.; Mo, X.; Fan, H.; Liangzhi Li, L. Biochemical and structural insights into an Ochrobactrum sp. CSL1 ribose-5-phosphate isomerase A and its roles in isomerization of rare sugars. Enz. Microb. Technol. 2020, 140, 109604. [Google Scholar] [CrossRef]
- Noel, P.; Wilkins, P. Phosphoglucose isomerase in marine molluscs. In Isozymes; Markert, C.L., Ed.; Academic Press: Cambridge, MA, USA, 1975; pp. 931–943. [Google Scholar] [CrossRef]
- Madgwick, J.; Haug, A.; Larsen, B. Polymannuronic acid 5-epimerase from the marine alga Pelvetia canaliculata (L.) Dcne. et Thur. Acta Chem. Scand. 1973, 27, 3592–3594. [Google Scholar] [CrossRef] [Green Version]
- Sawyer, M.H.; Baumann, P.; Baumann, L. Pathways of D-fructose and D-glucose catabolism in marine species of Alcaligenes, Pseudomonas marina, and Alteromonas communis. Arch. Microbiol. 1977, 112, 169–172. [Google Scholar] [CrossRef]
- Lavie, B.; Nevo, E.; Zoller, U. Differential viability of phosphoglucose isomerase allozyme genotypes of marine snails in nonionic detergent and crude oil-surfactant mixtures. Environ. Res. 1984, 35, 270–276. [Google Scholar] [CrossRef]
- Hall, J.G. The adaptation of enzymes to temperature: Catalytic characterization of glucosephosphate isomerase homologues isolated from Mytilus edulis and Isognomon alatus, bivalve molluscs inhabiting different thermal environments. Mol. Biol. Evol. 1985, 2, 251–269. [Google Scholar] [CrossRef] [Green Version]
- Adler, E.; Knowles, J. A Thermolabile triosephosphate isomerase from the psychrophile Vibrio sp. strain ANT-300. Arch. Biochem. Biophys. 1995, 321, 137–139. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Ragan, M.A. Cloning and characterization of the nuclear gene and cDNAs for triosephosphate isomerase of the marine red alga Gracilaria verrucosa. Curr. Genet. 1995, 28, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Manchenko, G.P. Unusual isozyme patterns of glucose-6-phosphate isomerase in Polydora brevipalpa (Polychaeta: Spionidae). Biochem. Genet. 2001, 39, 285–288. [Google Scholar] [CrossRef] [PubMed]
- Iwahara, K.; Takahashi, R.; Naomi, T.; Kida, M.; Miyamoto, R.; Tokuyama, T. Purification and comparison of triosephosphate isomerases from ammonia-oxidizing bacteria isolated from terrestrial and marine environments. J. Biosci. Bioeng. 2001, 91, 603–606. [Google Scholar] [CrossRef]
- Goulard, F.; Pondaven, P.; Diouris, M.; Deslandes, E.; Floch, J.Y. Partial Purification and Characterization of UDP-Glucose-4-Epimerase from Solieria chordalis (Rhodophyceae). Bot. Mar. 2003, 46, 107–111. [Google Scholar] [CrossRef]
- Caponera, J.A.; Rawson, P.D. Highly divergent duplicate mannose-6-phosphate isomerase (Mpi) genes in the blue mussel, Mytilus edulis. Mar. Genom. 2008, 1, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Fitriani, D.; Saksono, B. Cloning of araA Gene Encoding L-Arabinose Isomerase from Marine Geobacillus stearothermophilus Isolated from Tanjung Api, Poso, Indonesia. HAYATI J. Biosci. 2010, 17, 58–62. [Google Scholar] [CrossRef] [Green Version]
- Schoville, S.D.; Flowers, J.M.; Burton, R.S. Diversifying Selection Underlies the Origin of Allozyme Polymorphism at the Phosphoglucose Isomerase Locus in Tigriopus californicus. PLoS ONE 2012, 7, e40035. [Google Scholar] [CrossRef] [Green Version]
- Rodionova, I.A.; Scott, D.A.; Grishin, N.V.; Osterman, A.L.; Rodionov, D.A. Tagaturonate-fructuronate epimerase UxaE, a novel enzyme in the hexuronate catabolic network in Thermotoga maritima. Environ. Microbiol. 2012, 14, 2920–2934. [Google Scholar] [CrossRef] [Green Version]
- Chung, S.-K.; Ryu, S.-I.; Lee, S.-B. Characterization of UDP-glucose 4-epimerase from Pyrococcus horikoshii: Regeneration of UDP to produce UDP-galactose using two-enzyme system with trehalose. Bioresour. Technol. 2012, 110, 423–429. [Google Scholar] [CrossRef]
- Raedts, J.; Lundgren, M.; Kengen, S.W.M.; Li, J.-P.; Van Der Oost, J. A Novel Bacterial Enzyme with d-Glucuronyl C5-epimerase Activity. J. Biol. Chem. 2013, 288, 24332–24339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Wang, X.; Zhang, Y.; Yu, J.; Liu, T.; Chen, S.; Chi, S. Origin and evolution of alginate-c5-mannuronan-epimerase gene based on transcriptomic analysis of brown algae. Acta Oceanol. Sin. 2014, 33, 73–85. [Google Scholar] [CrossRef]
- Yun, E.J.; Lee, S.; Kim, H.T.; Pelton, J.G.; Kim, S.; Ko, H.-J.; Choi, I.-G.; Kim, K.H. The novel catabolic pathway of 3,6-anhydro-L-galactose, the main component of red macroalgae, in a marine bacterium. Environ. Microbiol. 2014, 17, 1677–1688. [Google Scholar] [CrossRef] [PubMed]
- Akutsu, J.-I.; Zhang, Z.; Morita, R.; Kawarabayasi, Y. Identification and characterization of a thermostable bifunctional enzyme with phosphomannose isomerase and sugar-1-phosphate nucleotidylyltransferase activities from a hyperthermophilic archaeon, Pyrococcus horikoshii OT3. Extremophiles 2015, 19, 1077–1085. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Zavala, A.A.; Carrasco-Miranda, J.S.; Ramirez-Aguirre, C.D.; López-Hidalgo, M.; Benítez-Cardoza, C.G.; Ochoa-Leyva, A.; Cardona-Felix, C.S.; Diaz-Quezada, C.; Rudiño-Piñera, E.; Sotelo-Mundo, R.R.; et al. Structural insights from a novel invertebrate triosephosphate isomerase from Litopenaeus vannamei. Biochim. et Biophys. Acta (BBA) Bioenerg. 2016, 1864, 1696–1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lajoie, C.A.; Kitner, J.B.; Potochnik, S.J.; Townsend, J.M.; Beatty, C.C.; Kelly, C.J. Cloning, expression and characterization of xylose isomerase from the marine bacterium Fulvimarina pelagiin in Escherichia coli. Biotechnol. Prog. 2016, 32, 1230–1237. [Google Scholar] [CrossRef]
- Fischl, R.; Bertelsen, K.; Gaillard, F.; Coelho, S.; Michel, G.; Klinger, M.; Boyen, C.; Czjzek, M.; Hervé, C. The cell-wall active mannuronan C5-epimerases in the model brown alga Ectocarpus: From gene context to recombinant protein. Glycobiology 2016, 26, 973–983. [Google Scholar] [CrossRef] [Green Version]
- Inoue, A.; Satoh, A.; Morishita, M.; Tokunaga, Y.; Miyakawa, T.; Tanokura, M.; Ojima, T. Functional heterologous expression and characterization of mannuronan C5-epimerase from the brown alga Saccharina japonica. Algal Res. 2016, 16, 282–291. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Yun, E.J.; Kim, K.H.; Kim, H.-Y.; Choi, I.-G. 3,6-Anhydro-L-galactonate cycloisomerase from Vibrio sp. strain EJY3: Crystallization and X-ray crystallographic analysis. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2017, 73, 511–514. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, Z.-W.; Hurlburt, B.K.; Li, G.-L.; Zhang, Y.-X.; Fei, D.-X.; Shen, H.-W.; Cao, M.-J.; Liu, G.-M. Identification of triosephosphate isomerase as a novel allergen in Octopus fangsiao. Mol. Immunol. 2017, 85, 35–46. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, Y.-X.; Liu, M.; Maleki, S.J.; Zhang, M.-L.; Liu, Q.-M.; Cao, M.-J.; Su, W.-J.; Liu, G.-M. Triosephosphate isomerase and filamin C share common epitopes as novel allergens of Procambarus clarkii. J. Agric. Food Chem. 2017, 65, 950–963. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Chen, Q.; Song, Y.; Huang, H.; Li, J.; Ma, J.; Li, Q.; Ju, J. Deciphering the sugar biosynthetic pathway and tailoring steps of nucleoside antibiotic A201A biosynthesis unveils GDP-l-galactose mutase. Proc. Natl. Acad. Sci. USA 2017, 114, 4948–4953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merkx-Jacques, A.; Rasmussen, H.; Muise, D.M.; Benjamin, J.J.R.; Kottwitz, H.; Tanner, K.; Milway, M.T.; Purdue, L.M.; Scaife, M.A.; Armenta, R.E.; et al. Engineering xylose metabolism in thraustochytrid T18. Biotechnol. Biofuels 2018, 11, 248. [Google Scholar] [CrossRef] [PubMed]
- Xia, F.; Li, M.-S.; Liu, Q.-M.; Liu, M.; Yang, Y.; Cao, M.-J.; Chen, G.-X.; Jin, T.; Liu, G.-M. Crystal Structure Analysis and Conformational Epitope Mutation of Triosephosphate Isomerase, a Mud Crab Allergen. J. Agric. Food Chem. 2019, 67, 12918–12926. [Google Scholar] [CrossRef]
- Hu, Y.; Du, Q.; Mi, P.; Shang, E.; Sui, Z. Gene cloning and expression regulation in the pathway of agar and floridean starch synthesis of Gracilariopsis lemaneiformis (Rhodophyta). Environ. Boil. Fishes 2019, 31, 1889–1896. [Google Scholar] [CrossRef]
- Rengasamy, S.; Subramanian, M.R.; Perumal, V.; Ganeshan, S.; Al Khulaifi, M.M.; Al-Shwaiman, H.A.; Elgorban, A.M.; Syed, A.; Thangaprakasam, U. Purification and kinetic behavior of glucose isomerase from Streptomyces lividans RSU26. Saudi J. Biol. Sci. 2020, 27, 1117–1123. [Google Scholar] [CrossRef]
- Sakai, N.; Tanaka, M.; Takahashi, M.; Adachi, S.; Nagahama, Y. Isolation and expression of rainbow trout (Oncorhynchus mykiss) ovarian cDNA encoding 3β-hydroxysteroid dehydrogenase/Δ5−4-isomerase. Fish Physiol. Biochem. 1993, 11, 273–279. [Google Scholar] [CrossRef]
- Misawa, N.; Shimada, H. Metabolic engineering for the production of carotenoids in non-carotenogenic bacteria and yeasts. J. Biotechnol. 1998, 59, 169–181. [Google Scholar] [CrossRef]
- Wise, M.L.; Rossi, J.; Gerwick, W.H. Characterization of the substrate binding site of polyenoic fatty acid isomerase, a novel enzyme from the marine alga Ptilota filicina. Biochemistry 1997, 36, 2985–2992. [Google Scholar] [CrossRef]
- Wang, C.W.; Oh, M.K.; Liao, J.C. Engineered isoprenoid pathway enhances astaxanthin production in Escherichia coli. Biotechnol. Bioeng. 1999, 62, 235–241. [Google Scholar] [CrossRef]
- Huang, J.; Jiang, X.; Zhang, X.; Chen, W.; Tian, B.; Shu, Z.; Hu, S. Expressed sequence tag analysis of marine fungus Schizochytrium producing docosahexaenoic acid. J. Biotechnol. 2008, 138, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Leblond, J.D.; Dodson, J.; Khadka, M.; Holder, S.; Seipelt, R.L. Sterol composition and biosynthetic genes of the recently discovered photosynthetic alveolate, Chromera velia (chromerida), a close relative of apicomplexans. J. Eukaryot. Microbiol. 2012, 59, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Liu, M.; He, M.; Ye, Y.; Huang, J. Illustrating and Enhancing the Biosynthesis of Astaxanthin and Docosahexaenoic Acid in Aurantiochytrium sp. SK4. Mar. Drugs 2019, 17, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Q.-L.; Zheng, J.-L.; Liu, J. Transcription activation of β-carotene biosynthetic genes at the initial stage of stresses as an indicator of the increased β-carotene accumulation in isolated Dunaliella salina strain GY-H13. Aquat. Toxicol. 2020, 222, 105472. [Google Scholar] [CrossRef] [PubMed]
- Omura, Y.; Hayashi, Y.; Matsushima, O.; Katayama, H.; Yamada, K. Partial purification and characterization of alanine racemase from the brackish-water bivalve Corbicula japonica. J. Exp. Mar. Biol. Ecol. 1985, 94, 281–289. [Google Scholar] [CrossRef]
- Yamada, A.; Matsushima, O. The relation of d-alanine and alanine racemase activity in molluscs. Comp. Biochem. Physiol. Part B 1992, 103, 617–621. [Google Scholar] [CrossRef]
- Oren, A.; Gurevich, P. Diversity of lactate metabolism in halophilic archaea. Can. J. Microbiol. 1995, 41, 302–307. [Google Scholar] [CrossRef]
- Iida, T.; Furutani, M.; Iwabuchi, T.; Maruyama, T. Gene for a cyclophilin-type peptidyl–prolyl cis–trans isomerase from a halophilic archaeum, Halobacterium cutirubrum. Gene 1997, 204, 139–144. [Google Scholar] [CrossRef]
- Shibata, K.; Shirasuna, K.; Motegi, K.; Kera, Y.; Abe, H.; Yamada, R.-H. Purification and properties of alanine racemase from crayfish Procambarus clarkii. Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol. 2000, 126, 599–608. [Google Scholar] [CrossRef]
- Yokoyama, T.; Tanaka, Y.; Sato, M.; Kan-No, N.; Nakano, T.; Yamaguchi, T.; Nagahisa, E. Alanine racemase activity in the microalga Thalassiosira sp. Fish. Sci. 2005, 71, 924–930. [Google Scholar] [CrossRef]
- Yokoyama, T.; Mimura, Y.; Sato, M.; Nagahisa, E. Purification and some properties of alanine racemase from the marine gastropod Cellana grata. Fish. Sci. 2006, 72, 195–201. [Google Scholar] [CrossRef]
- Safavi-Hemami, H.; Siero, W.A.; Gorasia, D.G.; Young, N.D.; Macmillan, D.; Williamson, N.A.; Purcell, A.W. Specialisation of the Venom Gland Proteome in Predatory Cone Snails Reveals Functional Diversification of the Conotoxin Biosynthetic Pathway. J. Proteome Res. 2011, 10, 3904–3919. [Google Scholar] [CrossRef]
- Sha, Z.-X.; Liu, H.; Wang, Q.-L.; Liu, Y.; Lu, Y.; Li, M.; Chen, S.-L. Channel catfish (Ictalurus punctatus) protein disulphide isomerase, PDIA6: Molecular characterization and expression regulated by bacteria and virus inoculation. Fish Shellfish. Immunol. 2012, 33, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Li, L.-C.; Hsu, Y.-T.; Chang, H.-L.; Wu, T.-M.; Sung, M.-S.; Cho, C.-L.; Lee, T.-M. Polyamine effects on protein disulfide isomerase expression and implications for hypersalinity stress in the marine algaUlva lactucaLinnaeus1. J. Phycol. 2013, 49, 1181–1191. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, D.; Wang, L.; Gai, Y.; Zhou, Z.; Zhang, H.; Song, L. The molecular characterization of a cyclophilin A from Chinese mitten crab Eriocheir sinensis and the antifungal activity of its recombinant protein. Electron. J. Biotechnol. 2013, 16. [Google Scholar] [CrossRef]
- Kim, S.-H.; Lee, J.M.; Kang, D.S.; Kim, D.-G.; Ahn, S.-H.; Kong, I.-S. Expression, purification and characterization of soluble recombinant peptidyl-prolyl cis/trans isomerase from Vibrio anguillarum. Protein Expr. Purif. 2014, 101, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tang, W.; Wang, X.; Chen, Y.; Wu, Y.; Qiang, Y.; Feng, Y.; Ren, Z.; Chen, S.; Xu, A. PPIase is associated with the diversity of conotoxins from cone snail venom glands. Biochimie 2015, 112, 129–138. [Google Scholar] [CrossRef]
- Jakob, R.P.; Schmidpeter, P.A.M.; Koch, J.R.; Schmid, F.X.; Maier, T. Structural and Functional Characterization of a Novel Family of Cyclophilins, the AquaCyps. PLoS ONE 2016, 11, e0157070. [Google Scholar] [CrossRef] [Green Version]
- Ekubota, T.; Shimamura, S.; Kobayashi, T.; Nunoura, T.; Deguchi, S. Distribution of eukaryotic serine racemases in the bacterial domain and characterization of a representative protein in Roseobacter litoralis Och 149. Microbiology 2016, 162, 53–61. [Google Scholar] [CrossRef]
- Subin, C.S.; Pradeep, M.A.; Vijayan, K.K. FKBP-type peptidyl-prolyl cis-trans isomerase from thermophilic microalga, Scenedesmus sp.: Molecular characterisation and demonstration of acquired salinity and thermotolerance in E. coli by recombinant expression. Environ. Boil. Fishes 2016, 28, 3307–3315. [Google Scholar] [CrossRef]
- Hoppstock, L.; Trusch, F.; Lederer, C.; Van West, P.; Könneke, M.; Bayer, P. NmPin from the marine thaumarchaeote Nitrosopumilus maritimus is an active membrane associated prolyl isomerase. BMC Biol. 2016, 14, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safavi-Hemami, H.; Li, Q.; Jackson, R.L.; Song, A.S.; Boomsma, W.; Bandyopadhyay, P.K.; Gruber, C.W.; Purcell, A.W.; Yandell, M.; Olivera, B.M.; et al. Rapid expansion of the protein disulfide isomerase gene family facilitates the folding of venom peptides. Proc. Natl. Acad. Sci. USA 2016, 113, 3227–3232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueroa-Montiel, A.; Ramos, M.A.; Mares, R.E.; Dueñas, S.; Pimienta, G.; Ortiz, E.; Possani, L.D.; Licea-Navarro, A.F. In Silico Identification of Protein Disulfide Isomerase Gene Families in the De Novo Assembled Transcriptomes of Four Different Species of the Genus Conus. PLoS ONE 2016, 11, e0148390. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Xie, J.; Yang, S.; Ye, S.; Luo, M.; Wu, X. First characterization of three cyclophilin family proteins in the oyster, Crassostrea ariakensis Gould. Fish Shellfish. Immunol. 2016, 55, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, X.; Ren, Z.; Tang, W.; Zou, Q.; Wang, J.; Chen, S.; Zhang, H.; Xu, A. Oxidative Folding of Conopeptides Modified by Conus Protein Disulfide Isomerase. Protein J. 2017, 36, 407–416. [Google Scholar] [CrossRef]
- Muhammad, F.; Zhi-Feng, Z.; Ming-Yu, S.; Shafi, M. cDNA Cloning and expression of cyclophilin A (LvCypA) in white leg shrimp, Litopenaeus vannamei. Pak. J. Zoöl. 2017, 49, 935–941. [Google Scholar] [CrossRef]
- Lee, H.; Kim, S.H.; Han, Y.-J.; Im, S. PsCYP1 of marine red alga Pyropia seriata (Bangiales, Rhodophyta) confers salt and heat tolerance in Chlamydomonas. J. Appl. Phycol. 2017, 29, 617–625. [Google Scholar] [CrossRef]
- Morinaka, B.I.; Vagstad, A.L.; Piel, J. Radical S-Adenosylmethionine Peptide Epimerases: Detection of Activity and Characterization of d-Amino Acid Products. Methods Enzymol. 2018, 604, 237–257. [Google Scholar] [CrossRef]
- Qiu, X.; Zhu, W.; Wang, W.; Jin, H.; Zhu, P.; Zhuang, R.; Yan, X. Structural and functional insights into the role of a cupin superfamily isomerase in the biosynthesis of Choi moiety of aeruginosin. J. Struct. Biol. 2019, 205, 44–52. [Google Scholar] [CrossRef]
- Shang, Z.; Winter, J.M.; Kauffman, C.A.; Yang, I.; Fenical, W. Salinipeptins: Integrated genomic and chemical approaches reveal unusual D-amino acid-containing ribosomally synthesized and post-translationally modified peptides (RiPPs) from a Great Salt Lake Streptomyces sp. ACS Chem. Biol. 2019, 14, 415–425. [Google Scholar] [CrossRef]
- Company, R.; Antúnez, O.; Cosson, R.P.; Serafim, A.; Shillito, B.; Cajaraville, M.; Bebianno, M.J.; Torreblanca, A. Protein expression profiles in Bathymodiolus azoricus exposed to cadmium. Ecotoxicol. Environ. Saf. 2019, 171, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Ulagesan, S.; Choi, J.-W.; Nam, T.-J.; Choi, Y.H. Peptidyl-prolyl isomerase and the biological activities of recombinant protein cyclophilin from Pyropia yezoensis (PyCyp). Protein Expr. Purif. 2020, 172, 105636. [Google Scholar] [CrossRef] [PubMed]
- Umemoto, Y.; Shibata, T.; Araki, T. d-Xylose Isomerase from a Marine Bacterium, Vibrio sp. Strain XY-214, and d-Xylulose Production from β-1,3-Xylan. Mar. Biotechnol. 2011, 14, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wang, Z.-P.; Sheng, J.; Zheng, Y.; Ji, X.-F.; Zhou, H.-X.; Liu, X.-Y.; Chi, Z.-M. High and efficient isomaltulose production using an engineered Yarrowia lipolytica strain. Bioresour. Technol. 2018, 265, 577–580. [Google Scholar] [CrossRef]
- Mu, W.; Li, W.; Wang, X.; Zhang, T.; Jiang, B. Current studies on sucrose isomerase and biological isomaltulose production using sucrose isomerase. Appl. Microbiol. Biotechnol. 2014, 98, 6569–6582. [Google Scholar] [CrossRef]
- Jeong, D.W.; Hyeon, J.E.; Shin, S.K.; Han, S.O. Trienzymatic Complex System for Isomerization of Agar-Derived d-Galactose into d-Tagatose as a Low-Calorie Sweetener. J. Agric. Food Chem. 2020, 68, 3195–3202. [Google Scholar] [CrossRef]
- Wise, M.L.; Hamberg, M.; Gerwick, W.H. Biosynthesis of conjugated triene-containing fatty acids by a novel isomerase from the red marine alga Ptilota filicina. Biochemistry 1994, 33, 15223–15232. [Google Scholar] [CrossRef]
- Wise, M.L.; Söderström, K.; Murray, T.F.; Gerwick, W.H. Synthesis and cannabinoid receptor binding activity of conjugated triene anandamide, a novel eicosanoid. Cell. Mol. Life Sci. 1996, 52, 88–92. [Google Scholar] [CrossRef]
- Gerwick, W.H.; Moghaddam, M.; Hamberg, M. Oxylipin metabolism in the red alga Gracilariopsis lemaneiformis: Mechanism of formation of vicinal dihydroxy fatty acids. Arch. Biochem. Biophys. 1991, 290, 436–444. [Google Scholar] [CrossRef]
- Ghioni, C.; Porter, A.E.A.; Sadler, I.H.; Tocher, D.R.; Sargent, J.R. Cultured fish cells metabolize octadecapentaenoic acid (all-cis Δ3,6,9,12,15–18:5) to octadecatetraenoic acid (all-cis Δ 6,9,12,15–18:4) via its 2-trans intermediate (trans Δ2, all-cis δ6,9,12,15–18:5). Lipids 2001, 36, 145–153. [Google Scholar] [CrossRef] [Green Version]
- Rontani, J.-F.; Bonin, P.; Jameson, I.; Volkman, J.K. Degradation of alkenones and related compounds during oxic and anoxic incubation of the marine haptophyte Emiliania huxleyi with bacterial consortia isolated from microbial mats from the Camargue, France. Org. Geochem. 2005, 36, 603–618. [Google Scholar] [CrossRef]
- Nagashima, H.; Zulkharnain, A.; Maeda, R.; Fuse, H.; Iwata, K.; Omori, T. Cloning and nucleotide sequences of carbazole degradation genes from marine bacterium Neptuniibacter sp. strain CAR-SF. Curr. Microbiol. 2009, 61, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Glueck, S.; Gadler, P.; Kroutil, W.; Nestl, B.M.; Larissegger-Schnell, B.; Ueberbacher, B.; Wallner, S.; Faber, K. Biocatalytic approaches for the quantitative production of single stereoisomers from racemates. Biochem. Soc. Trans. 2006, 34, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Misawa, N.; Nodate, M.; Otomatsu, T.; Shimizu, K.; Kaido, C.; Kikuta, M.; Ideno, A.; Ikenaga, H.; Ogawa, J.; Shimizu, S.; et al. Bioconversion of substituted naphthalenes and β-eudesmol with the cytochrome P450 BM3 variant F87V. Appl. Microbiol. Biotechnol. 2010, 90, 147–157. [Google Scholar] [CrossRef]
- Jung, B.-O.; Roseman, S.; Park, J.K. The central concept for chitin catabolic cascade in marine bacterium, Vibrios. Macromol. Res. 2008, 16, 1–5. [Google Scholar] [CrossRef]
- Rosani, U.; Domeneghetti, S.; Maso, L.; Wegner, K.M.; Venier, P. An Evolutionary Perspective of Dopachrome Tautomerase Enzymes in Metazoans. Genes 2019, 10, 495. [Google Scholar] [CrossRef] [Green Version]
- Waisiel, A.A.; Rozeboom, H.J.; Hauke, D.; Baas, B.-J.; Zandvoort, E.; Quax, W.J.; Thunnissen, A.-M.W.H.; Poelarends, G.J. Structural and functional characterization of macrophage migration inhibiotry factor homologue from the marine cyanobacterium Prochlorocuccus marinus. Biochemitry 2010, 49, 7572–7581. [Google Scholar] [CrossRef] [Green Version]
- Zuasti, A.; Martínez-Liarte, J.H.; Solano, F.; Ferrer, C. Melanization stimulating factors in the integument of the Mugil cephalus and Dicertranchus labrax. Histol. Histopathol. 2000, 15, 1145–1150. [Google Scholar]
- Xu, F.; Li, M.-Y.; Chen, J.; Feng, X.; Ming-Yun, L.; Chen, J. D-dopachrome tautomerase from Japanese sea bass (Lateolabrax japonicus) is a chemokine-like cytokine and functional homolog of macrophage migration inhibitory factor. Zool Res. 2020, 41, 39–50. [Google Scholar] [CrossRef]
- Wang, D.; Yang, D.; Wang, Q.; Zhao, Y.; Li, C.; Wei, Q.; Han, Y.; Zhao, J. Two macrophage migration inhibitory factors (MIFs) from the clam Ruditapes philippinarum: Molecular characterization, localization and enzymatic activities. Fish Shellfish. Immunol. 2018, 78, 158–168. [Google Scholar] [CrossRef]
- Trincone, A. Enzymatic Processes in Marine Biotechnology. Mar. Drugs 2017, 15, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Reaction scheme of l-arabinose isomerase. The enzyme can also convert d-galactose to d-tagatose with lower efficiency. The enzyme is also present in the marine Geobacillus stearothermophilus (see Table 2).
Figure 1.
Reaction scheme of l-arabinose isomerase. The enzyme can also convert d-galactose to d-tagatose with lower efficiency. The enzyme is also present in the marine Geobacillus stearothermophilus (see Table 2).

Figure 2.
Reaction scheme of formation of conjugated triene anandamide catalyzed by P. filicina enzyme.
Figure 2.
Reaction scheme of formation of conjugated triene anandamide catalyzed by P. filicina enzyme.

Figure 3.
Carbazole degradation to 2′-aminobiphenyl-2,3-diol. Remediation study is present for this compound by the marine bacterium Neptuniibacter sp. A tautomerase could be involved in the lower degradation pathway as in total cleavage pathway for the degradation of phenols, modified phenols, and catechols.
Figure 3.
Carbazole degradation to 2′-aminobiphenyl-2,3-diol. Remediation study is present for this compound by the marine bacterium Neptuniibacter sp. A tautomerase could be involved in the lower degradation pathway as in total cleavage pathway for the degradation of phenols, modified phenols, and catechols.

Figure 4.
The reaction scheme for the enantioconvergent chemoenzymatic hydrolysis of sulfate esters by inverting marine Rhodococcus sulfatase. Big arrow indicated the preferred substrate.
Figure 4.
The reaction scheme for the enantioconvergent chemoenzymatic hydrolysis of sulfate esters by inverting marine Rhodococcus sulfatase. Big arrow indicated the preferred substrate.

Figure 5.
β-eudesmol hydroxylation. Regio- and stereo-specific actions of cytochrome P450 [86] were studied in depth with using NMR spectroscopy; 80% yield in preparative experiments was obtained.
Figure 5.
β-eudesmol hydroxylation. Regio- and stereo-specific actions of cytochrome P450 [86] were studied in depth with using NMR spectroscopy; 80% yield in preparative experiments was obtained.

Figure 6.
Xylose isomerase reaction; the enzyme catalyzes the interconversion of aldose and ketose sugars with broad substrate specificity; in the case of glucose, fructose is formed, and xylose isomerases are used extensively in the high-fructose corn syrup industry. The hemicellulose fraction of cellulosic biomass can be converted to xylose by xylanases, the need of xylose isomerase is based on the possibility of fermenting d-xylulose by Saccharomyces cerevisiae completing biomass utilization, being xylose not fermented.
Figure 6.
Xylose isomerase reaction; the enzyme catalyzes the interconversion of aldose and ketose sugars with broad substrate specificity; in the case of glucose, fructose is formed, and xylose isomerases are used extensively in the high-fructose corn syrup industry. The hemicellulose fraction of cellulosic biomass can be converted to xylose by xylanases, the need of xylose isomerase is based on the possibility of fermenting d-xylulose by Saccharomyces cerevisiae completing biomass utilization, being xylose not fermented.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Searches in literature.
| Databases | Search Statement | Hits |
|---|---|---|
| Science Direct | Isomerase * and marine in titles, abstracts and keywords | 31 |
| PubMed | Isomerase * and marine | 172 |
| WoS 1 | Marine epimerase * in All fields | 53 |
| WoS | Marine racemase * in All fields | 44 |
| WoS | Marine cis–trans isomerase * in All fields | 56 |
| WoS | Marine cycloisomerase * in All fields | 3 |
| WoS | Marine tautomerase * in All fields | 26 |
| WoS | Marine mutase * in All fields | 45 |
1 Web of Science, last date accessed June 2020 with updates up to submission; * indicated the plural.
Table 2.
Marine isomerases acting on sugar molecules.
| Reference/Year | Organism | Enzyme | Reaction | Note |
|---|---|---|---|---|
| [9] 1973 | Alga Pelvetia canaliculata | Polymannuronic-5-epimerase | Conversion of polymannuronic acid to a mixed polymer containing guluronic acid | Preparation of ammonium sulfate fraction of the enzyme |
| [10] 1977 | Marine species of Alcaligenes, Pseudomonas marina, and Alteromonas communis | P-hexose isomerase | Glycolytic pathway | Entner–Doudoroff pathway |
| [11] 1984 | Marine snails | Phosphoglucose isomerase | Glycolytic enzyme | Tolerance to detergents as monitoring tool |
| [12] 1985 | Bivalve mollusks: Mytilus edulis and Isognomon alatus | Glucose phosphate isomerase | Glycolytic enzyme | Biochemical-based study of adaptation of enzyme to temperature |
| [13] 1995 | Psychrophilic marine eubacterium Vibrio sp. strain ANT-300 | Triosephosphate isomerase | Interconversion dihydroxyacetone phosphate and d-glyceraldehyde-3-phosphate | Thermolability study |
| [14] 1995 | Marine red alga Gracilaria verrucosa | Triosephosphate isomerase | Interconversion dihydroxyacetone phosphate and d-glyceraldehyde-3-phosphate | Genetic study |
| [15] 2001 | Polychaeta Polydora brevipalpa | Glucose-6-phosphate isomerase | Glycolytic enzyme | Study of isozyme pattern |
| [16] 2001 | Marine ammonia-oxidizing bacteria Nitrosomonas | Triosephosphate isomerases | Interconversion dihydroxyacetone phosphate and d-glyceraldehyde-3-phosphate | Purification and characterization |
| [17] 2003 | Macroalga Solieria chordalis | UDP-glucose-4-epimerase | Catalyzing both the synthesis of UDP-Gal and UDP-Glc | Characterization of the enzyme |
| [18] 2008 | Blue mussel Mytilus edulis | Mannose-6-phosphate isomerase | Glycolytic enzyme | Genetic study |
| [19] 2010 | Marine Geobacillus stearothermophilus | l-Arabinose Isomerase | Converting d-galactose to d-tagatose | Clone and sequence araA gene |
| [20] 2012 | Marine copepod Tigriopus californicus | Phosphoglucose isomerase | Glycolytic enzyme | Genetic variability study |
| [21] 2012 | Thermotoga maritima | Tagaturonate-fructuronate epimerase UxaE | Epimerization of tagaturonate to fructuronate | Study of metabolism of galacturonate and glucuronate from pectin and xylan |
| [22] 2012 | Pyrococcus horikoshii | UDP-glucose 4-epimerase | Catalyzing both the synthesis of UDP-Gal and UDP-Glc | Characterization study of the enzyme that could be coupled with trehalose synthase |
| [23] 2013 | Marine bacterium Bermanella marisrubri sp. RED65 | d-glucuronyl C5-epimerase | Epimerization of d-glucuronic acid to its C5-epimer l-iduronic acid | Recombinant protein expressed in Escherichia coli showed epimerization activity |
| [24] 2014 | Brown algae | Alginate-C5-mannuronan-epimerase | Catalyze the conversion of mannuronate to guluronate and determine the M/G ratio of alginate | Genetic study: predicted 94 algG genes open reading frame (ORF) sequences of brown algae |
| [25] 2015 | Marine bacterium Vibrio sp. | 3,6-Anhydro-l-galactonate cycloisomerase | Converts 3,6-anhydro-l-galactonate into 2-keto-3-deoxygalactonate | Identification of intermediate products of 3,6-anhydro-l-galactose |
| [26] 2015 | Pyrococcus horikoshii | Phosphomannose isomerase | Mannosylglycerate biosynthetic pathway | Recombinant protein expressed in E. coli with double activity (Man-1-P GTase activity) |
| [27] 2016 | Marine Pacific whiteleg shrimp Litopenaeus vannamei | Triosephosphate isomerase | Interconversion dihydroxyacetone phosphate and d-glyceraldehyde-3-phosphate | Structural and mechanistic study and insights into glycolysis regulation in crustaceans |
| [28] 2016 | Marine bacterium Fulvimarina pelagi | Xylose isomerase | Interconversion of d-xylose and d-xylulose | Cloning, expression, and characterization for use in biofuels’ production |
| [29] 2016 | Brown alga Ectocarpus | Mannuronan C5-epimerase | Control the distribution pattern of (1-4) linked β-d-mannuronic acid (M) and alpha-l-guluronic acid (G) residues in alginates | Transcript expression |
| [30] 2016 | Alga Saccharina japonica | Mannuronan C5-epimerase | Control of the distribution pattern of (1-4) linked β-d-mannuronic acid (M) and alpha-l-guluronic acid (G) residues in alginates | Functional recombinant expression of protein in insect-cell system revealing alternate epimerization of beta-d-mannuronic acid to alpha-l-guluronic acid |
| [31] 2017 | Vibrio sp. strain EJY3 | 3,6-Anhydro-l-galactonate cycloisomerase | Converts 3,6-anhydro-l-galactonate into 2-keto-3-deoxygalactonate | Crystallization and X-ray analysis of recombinant protein |
| [32] 2017 | Octopus fangsiao | Triosephosphate isomerase | Interconversion dihydroxyacetone phosphate and d-glyceraldehyde-3-phosphate | Study of allergen function |
| [33] 2017 | Freshwater crayfish Procambarus clarkii | Triosephosphate isomerase | Interconversion dihydroxyacetone phosphate and d-glyceraldehyde-3-phosphate | Study of allergen function |
| [34] 2017 | Marinactinospora thermotolerans | GDP-l-galactose mutase | Conversion of pyranose form to furanose structure | Study of the sugar biosynthetic pathway |
| [35] 2018 | Marine fungus-like thraustochytrids | Xylose isomerase | Interconversion of d-xylose and d-xylulose | Identification and characterization of xylose metabolism |
| [36] 2019 | Scylla paramamosain | Triosephosphate isomerase | Interconversion dihydroxyacetone phosphate and d-glyceraldehyde-3-phosphate | Crystal structure |
| [37] 2019 | Gracilariopsis lemaneiformis | Mannose-6-phosphate isomerase, GDP-mannose-3,5-epimerase | Pathways of floridean starch | Transcriptomic study for the study of the mechanism of substrate competition of synthesis pathways of floridean starch |
| [38] 2020 | Marine Streptomyces lividans RSU26 | Glucose isomerase | Fructose to glucose conversion | Characterization study and optimization of enzyme production |
Table 3.
Marine isomerases acting on lipid molecules.
| Reference/Year | Organism | Enzyme | Reaction | Note |
|---|---|---|---|---|
| [39] 1993 | Rainbow trout Oncorhynchus mykiss | 3β-hydroxysteroid dehydrogenase/Δ(5-4)-isomerase | Steroidogenic enzymes involved in the production of 17α-hydroxyprogesterone | Genetic study |
| [40] 1997 | Marine bacterium Agrobacterium aurantiacum | Carotenoid gene cluster | β-carotene biosynthesis | Metabolic engineering study |
| [41] 1997 | Alga Ptilota filicina | Polyenoic fatty acid isomerase | Assay by conversion of arachidonic acid to a conjugated triene | Biochemical study of binding site characteristics |
| [42] 1999 | Marine bacterium Agrobacterium aurantiacum | Isopentenyl diphosphate (IPP) isomerase and gene cluster (crtBIYZW) | Isoprenoid pathway | Study to enhance astaxanthin production by engineering isoprenoid pathway |
| [3] 2002 | Marine alga Ptilota filicina | Polyenoic fatty acid isomerase | Assay by conversion of arachidonic acid to a conjugated triene | Study of protein characterization and functional expression |
| [43] 2008 | Marine fungus Schizochytrium | Enzymes involved in biosynthesis of fatty acid via polyketide synthases | Confirmation PKS pathway | Genetic study of docohexanoic acid biosynthesis |
| [44] 2012 | Marine alveolate Chromera velia | Isopentenyl diphosphate Δ-isomerase | Sterol biosynthesis | Study of sterol composition of Chromera velia for chemotaxonomic relationships |
| [45] 2019 | Marine thraustochytrid Aurantiochytrium | Isopentenyl pyrophosphate isomerase | Biosynthetic pathways of docosahexaenoic acid (DHA) and ketocarotenoid astaxanthin | Analyses of the genome, transcriptome, key enzymes, and pathway products |
| [46] 2020 | Dunaliella salina | 15-cis-Z-carotene isomerase, prolycopene isomerase | β-carotene biosynthesis | Study of β-carotene biosynthesis: seven full length cDNA sequences cloned |
Table 4.
Marine isomerases acting on amino acids and peptides.
| Reference/Year | Organism | Enzyme | Reaction | Note |
|---|---|---|---|---|
| [47] 1985 | Bivalve Corbicula japonica | Alanine racemase | l to d alanine | Partial purification and characterization |
| [48] 1992 | Eighteen molluscan species | Alanine racemase | l to d alanine | Comparative study and distribution |
| [49] 1995 | Haloferax volcanii and Haloarcula species | Lactate racemase | l to d lactate | Study of enzymatic diversity among species |
| [50] 1997 | Halobacterium cutirubrum | Peptidyl-prolyl cis/trans isomerase | Isomerization of peptide bonds (trans-cis) at Pro residues; facilitates protein folding | Genetic study and expression in E. coli |
| [51] 2000 | Crayfish Procambarus clarkii | Alanine racemase | l to d alanine | Isolation, kinetic properties, substrate specificity, structural characteristics |
| [2] 2001 | Black-tiger prawn, Penaeus monodon | Alanine racemase | l to d alanine | Kinetic properties and substrate specificity |
| [52] 2005 | Microalga Thalassiosira sp. | Alanine racemase | l to d alanine | Kinetic properties and substrate specificity |
| [53] 2006 | Marine gastropod Cellana grata | Alanine racemase | l to d alanine | First purification study and kinetic assessment in gastropod |
| [54] 2011 | Marine cone snails | Disulfide isomerase | Oxidation, isomerization, and reduction of S–S bonds | Proteomic study showing presence of multitude of isoform of the enzyme |
| [55] 2012 | Channel catfish Ictalurus punctatus | Disulfide isomerase | Oxidation, isomerization and reduction of S–S bonds | Genetic study |
| [56] 2013 | Marine alga Ulva lactuca | Disulfide isomerase | Oxidation, Isomerization, and reduction of S–S bonds | Study of cloning and expression |
| [57] 2013 | Crab Eriocheir sinensis | Peptidyl-prolyl cis/trans isomerase | Isomerization of peptide bonds (trans-cis) at Pro residues; facilitates protein folding | Purification of recombinant protein and assessment of antifungal properties |
| [58] 2014 | Marine bacterium Vibrio anguillarum | Peptidyl-prolyl cis/trans isomerase | Isomerization of peptide bonds (trans-cis) at Pro residues; facilitates protein folding | Changes in protein expression of V. anguillarum, gene expression in E. coli and biochemical characterization |
| [59] 2015 | Core snails | Disulfide isomerase | Oxidation, Isomerization, and reduction of S–S bonds | Proteomic study |
| [60] 2016 | Marine Alphaproteobacteria | A novel family of peptidyl-prolyl isomerase | Isomerization of peptide bonds (trans-cis) at Pro residues; facilitates protein folding | Structural and functional characterization |
| [61] 2016 | Marine heterotrophic bacterium Roseobacter litoralis | Serine racemase | Racemization and minor dehydration of serine | Genomic analysis |
| [62] 2016 | Thermophilic chlorophycean microalga, Scenedesmus sp. | Peptidyl-prolyl cis/trans isomerase | Isomerization of peptide bonds (trans-cis) at Pro residues; facilitates protein folding | Cloning and expression of the enzyme in E. coli and indication of role in stress-tolerance mechanisms |
| [63] 2016 | Marine thaumarchaeote Nitrosopumilus maritimus | Peptidyl-prolyl cis/trans isomerase | Isomerization of peptide bonds (trans-cis) at Pro residues; facilitates protein folding | A protein structure study |
| [64] 2016 | Superfamily Conoidea | Disulfide isomerase | Oxidation, Isomerization, and reduction of S–S bonds | Study of diversification of enzymatic protein folding correlated with diversity of conotoxins |
| [65] 2016 | Marine snails belonging to Conus | Disulfide isomerase | Oxidation, Isomerization, and reduction of S–S bonds | Transcriptomic and in silico analysis and characterization of the group of PDI protein sequences |
| [66] 2016 | Oyster Crassostrea ariakensis Gould | Peptidyl-prolyl isomerase (cyclophilins) | Isomerization of peptide bonds (trans-cis) at Pro residues; facilitates protein folding | Enzymatic tissue distribution and role of the three enzymes identified and involvement in in oyster immune response |
| [67] 2017 | Cone snail species | Disulfide isomerase | Oxidation, Isomerization, and reduction of S–S bonds | Cloned 12 disulfide isomerase genes and study of reaction on conopeptides |
| [68] 2017 | Shrimp, Litopenaeus vannamei | Peptidyl-prolyl isomerase (cyclophilins) | Isomerization of peptide bonds (trans-cis) at Pro residues; facilitates protein folding | Cloning and tissue distribution of the enzyme |
| [69] 2017 | Red alga Pyropia seriata | Peptidyl-prolyl isomerase (cyclophilins) | Isomerization of peptide bonds (trans-cis) at Pro residues; facilitates protein folding | Transcriptomic study |
| [70] 2018 | Cyanobacterial genomes | PoyD, a member of the radical S-adenosylmethionine superfamily | Introducing d-amino acids into a ribosomally synthesized peptide | Heterologous expression in E. coli, detection of epimerase activity, and localization of epimerization sites |
| [71] 2019 | Cyanobacteria | AerE, a cupin superfamily enzyme | 1,3-allylic isomerization | Study of the biosynthesis of aeruginosins trapeptides possessing antithrombotic activity |
| [72] 2019 | Halotolerant Streptomyces sp. strain GSL-6C | Inferring new epimerases | Conversion of l- to d-amino acids | Genome analysis integrating a study on salinipeptins |
| [73] 2019 | Hydrothermal vent mussel Bathymodiolus azoricus | Peptidyl-prolyl cis/trans isomerase | Isomerization of peptide bonds (trans-cis) at Pro residues; facilitates protein folding | Study of proteome changes upon Cd exposure for bioindicator identification |
| [74] 2020 | Marine red algae Pyropia yezoensis | Peptidyl-prolyl isomerase (cyclophilins) | Isomerization of peptide bonds (trans-cis) at Pro residues; facilitates protein folding | Analysis of the biological activity of recombinant cyclophilin |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Trincone, A. Application-Oriented Marine Isomerases in Biocatalysis. Mar. Drugs 2020, 18, 580. https://doi.org/10.3390/md18110580
AMA Style
Trincone A. Application-Oriented Marine Isomerases in Biocatalysis. Marine Drugs. 2020; 18(11):580. https://doi.org/10.3390/md18110580
Chicago/Turabian StyleTrincone, Antonio. 2020. "Application-Oriented Marine Isomerases in Biocatalysis" Marine Drugs 18, no. 11: 580. https://doi.org/10.3390/md18110580
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.