Natural Polypropionates in 1999–2020: An Overview of Chemical and Biological Diversity


Abstract
:
1. Introduction
2. Isolation, Structural Features and Bioactivities of Polypropionates
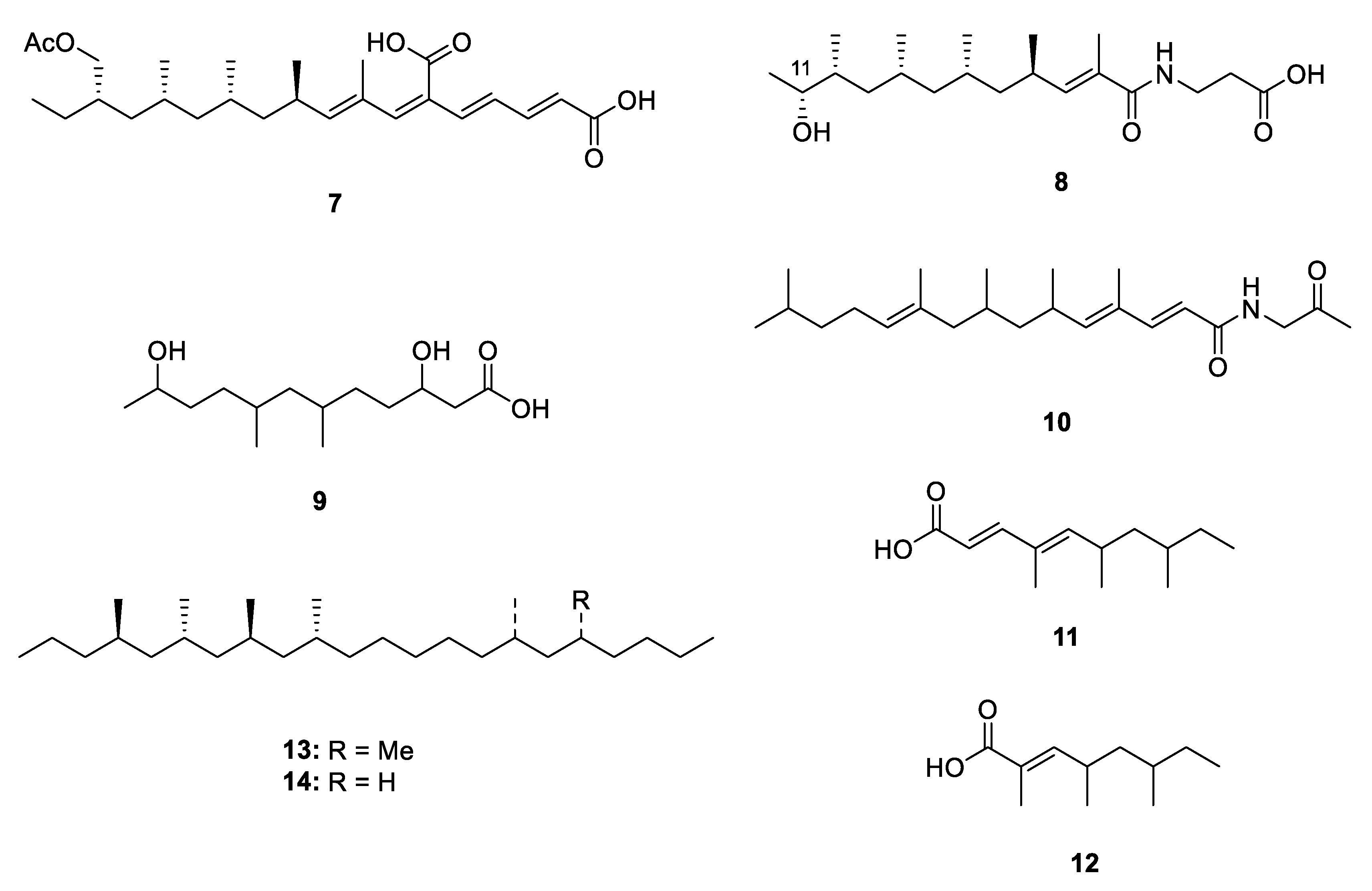
2.1. Linear Metabolites
2.2. Cyclic Metabolites
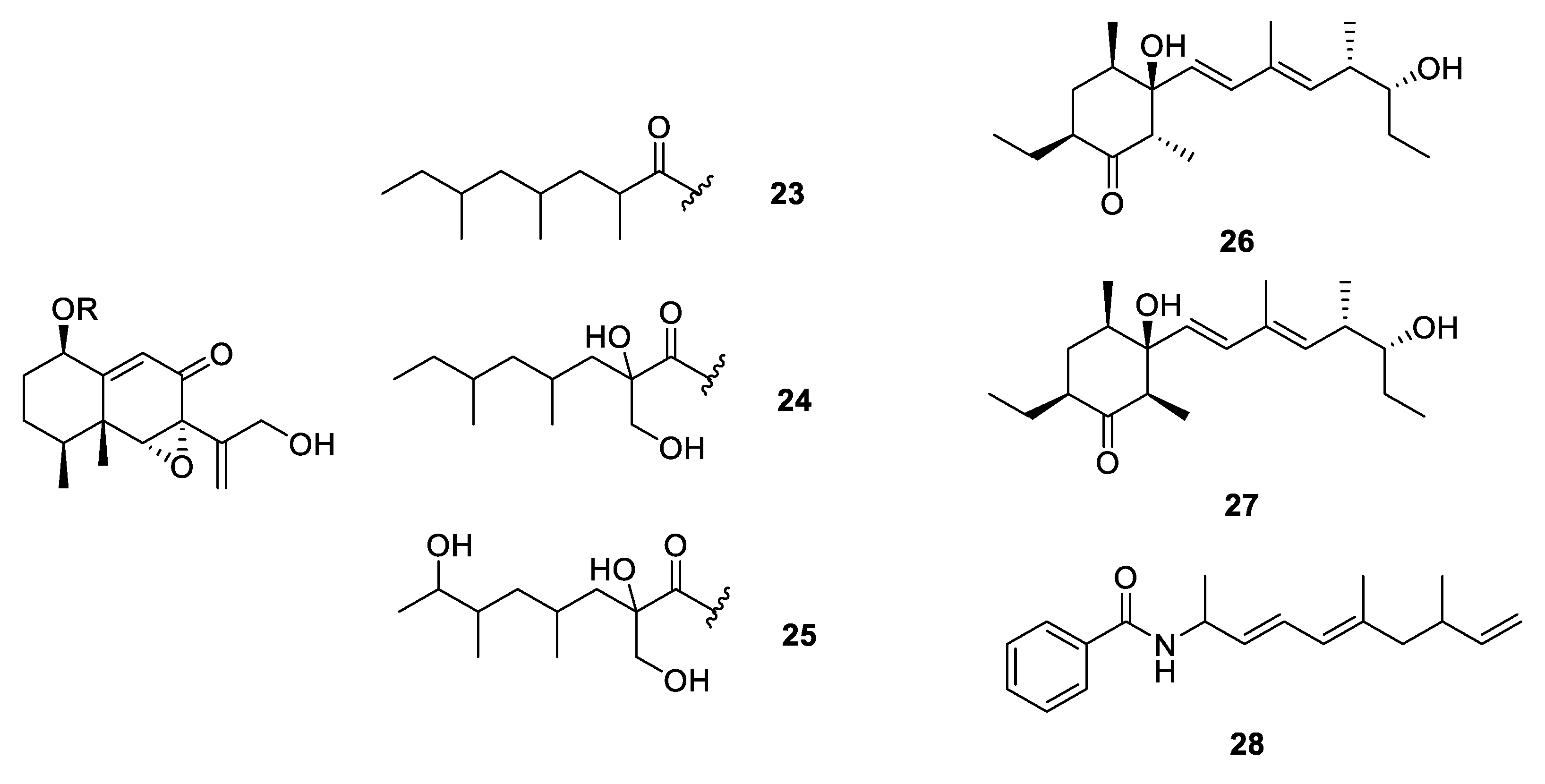
2.2.1. Carbon Homocyclic Metabolites
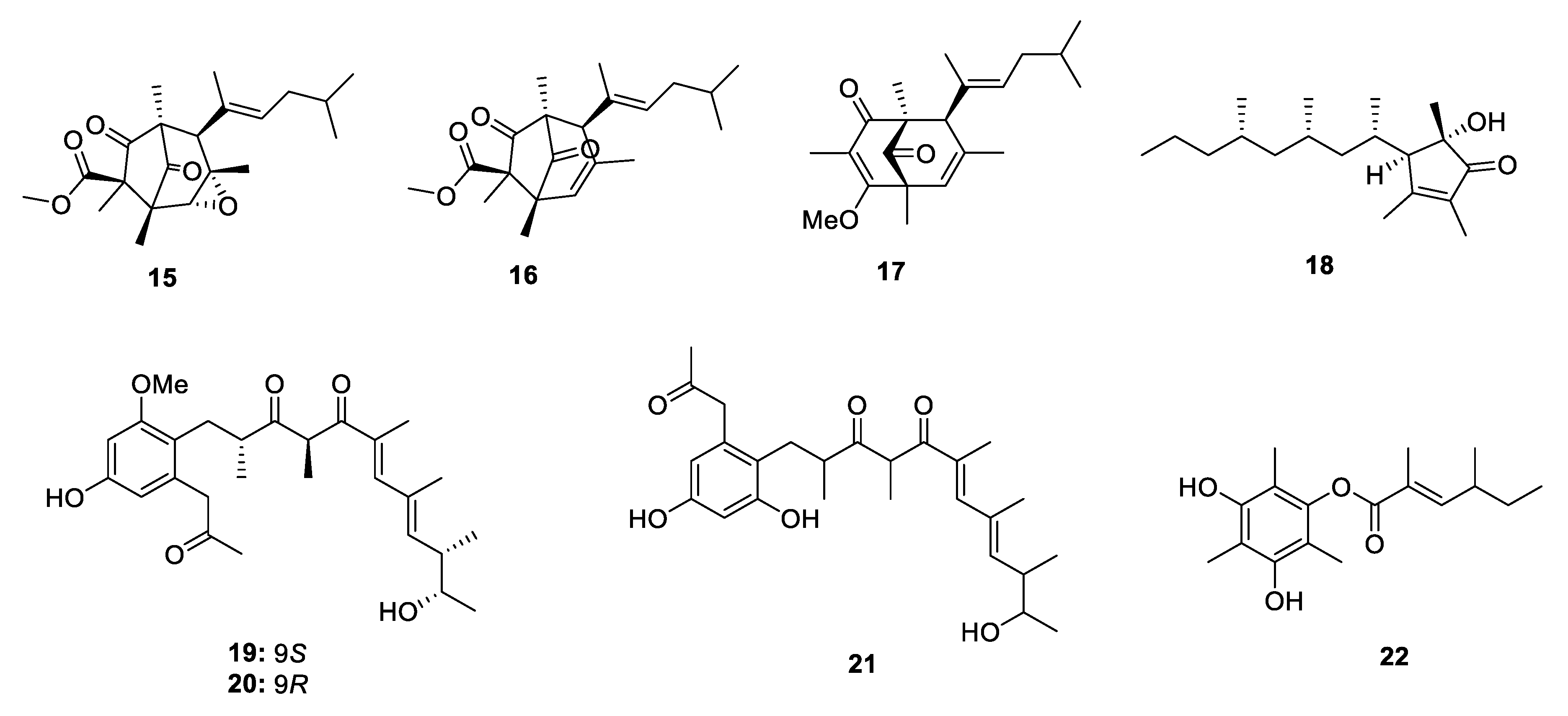
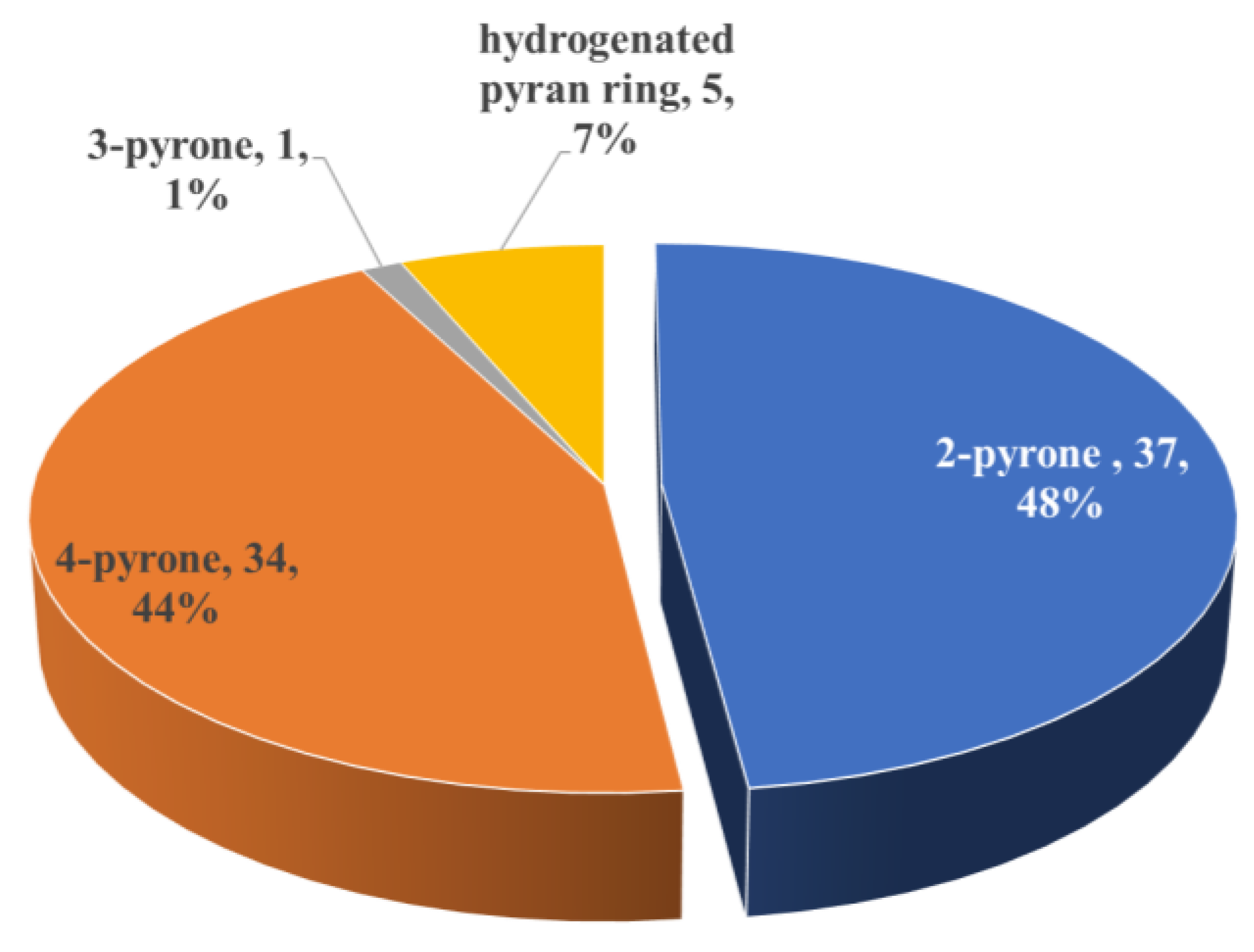
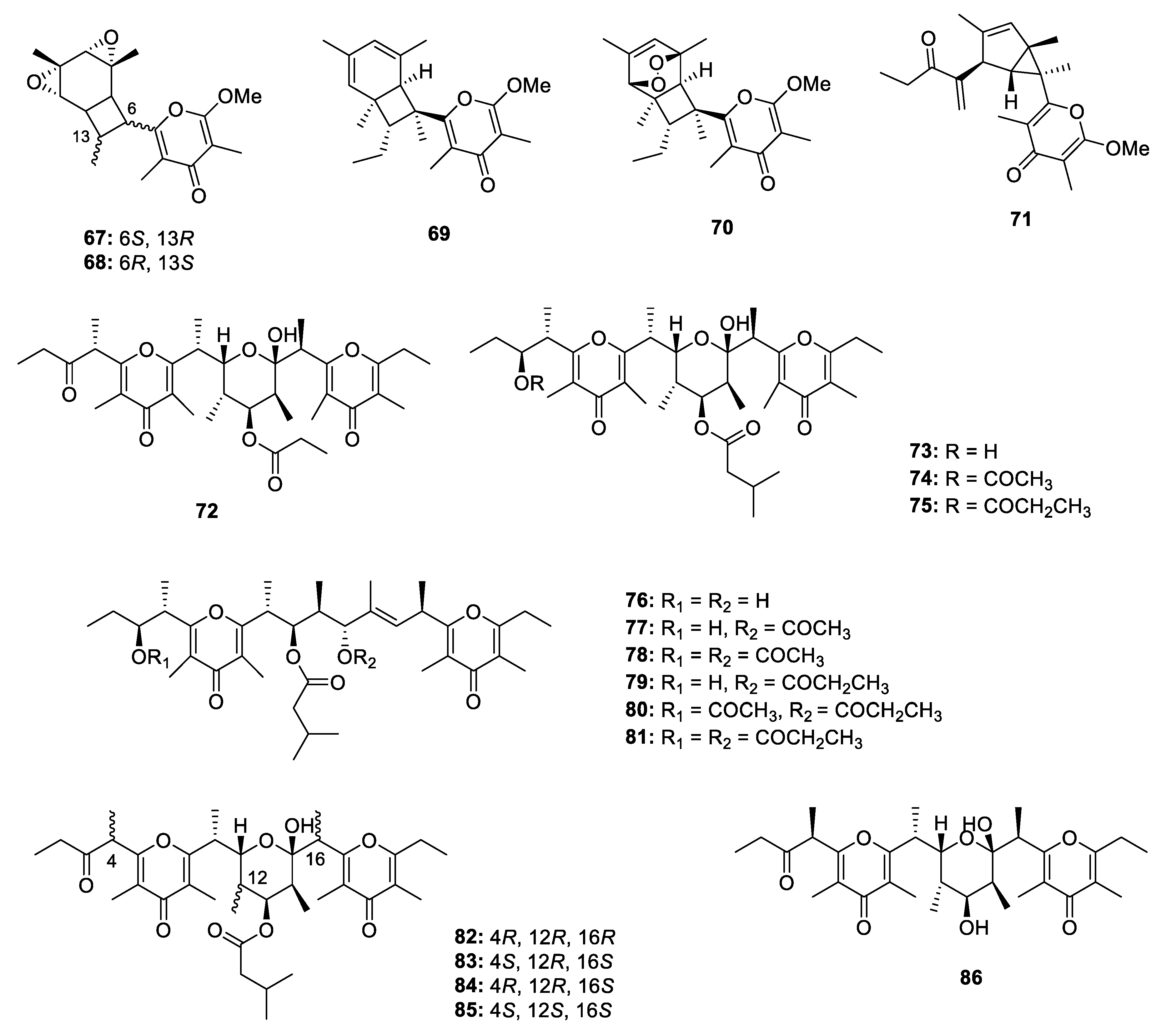
2.2.2. Pyran-Related Metabolites
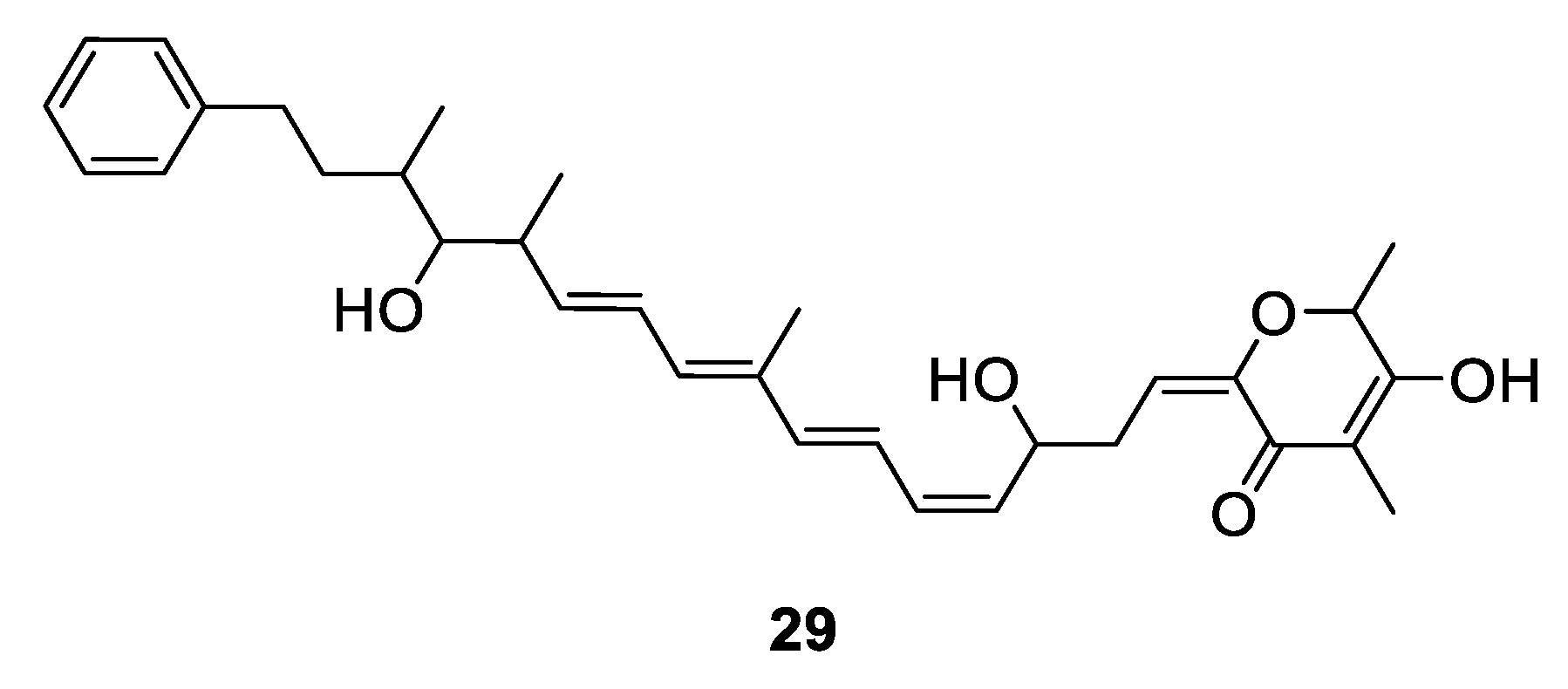
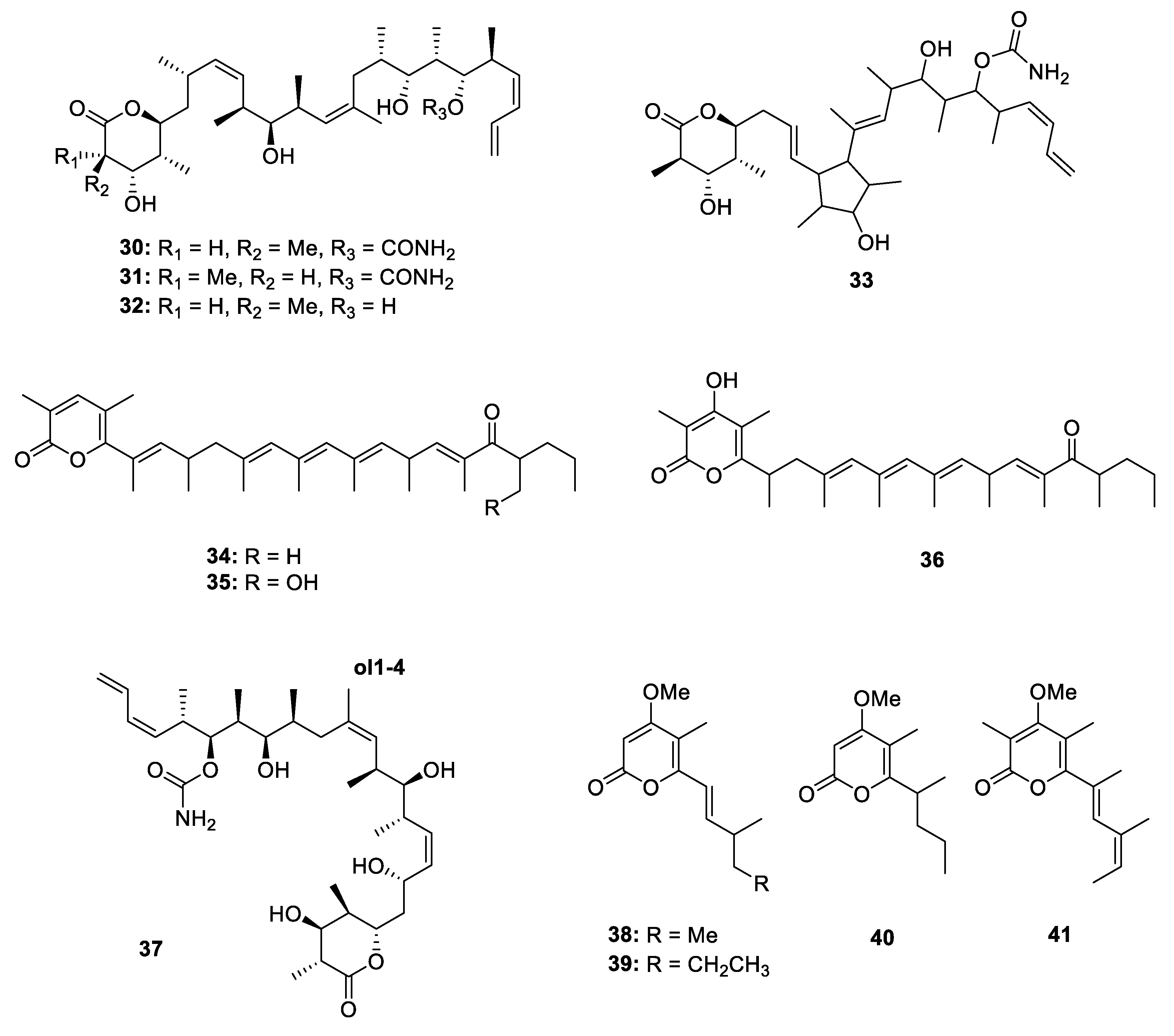
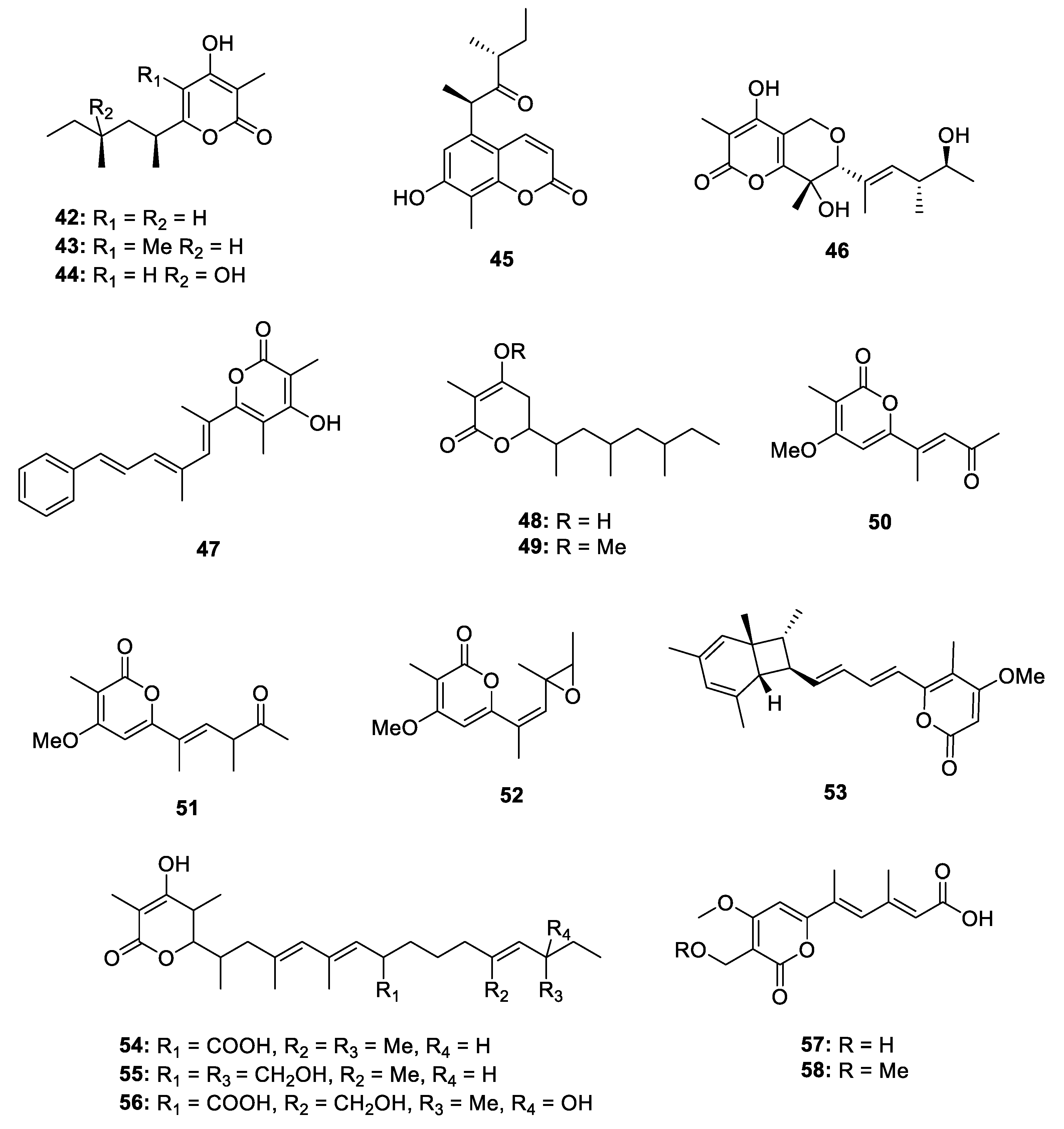
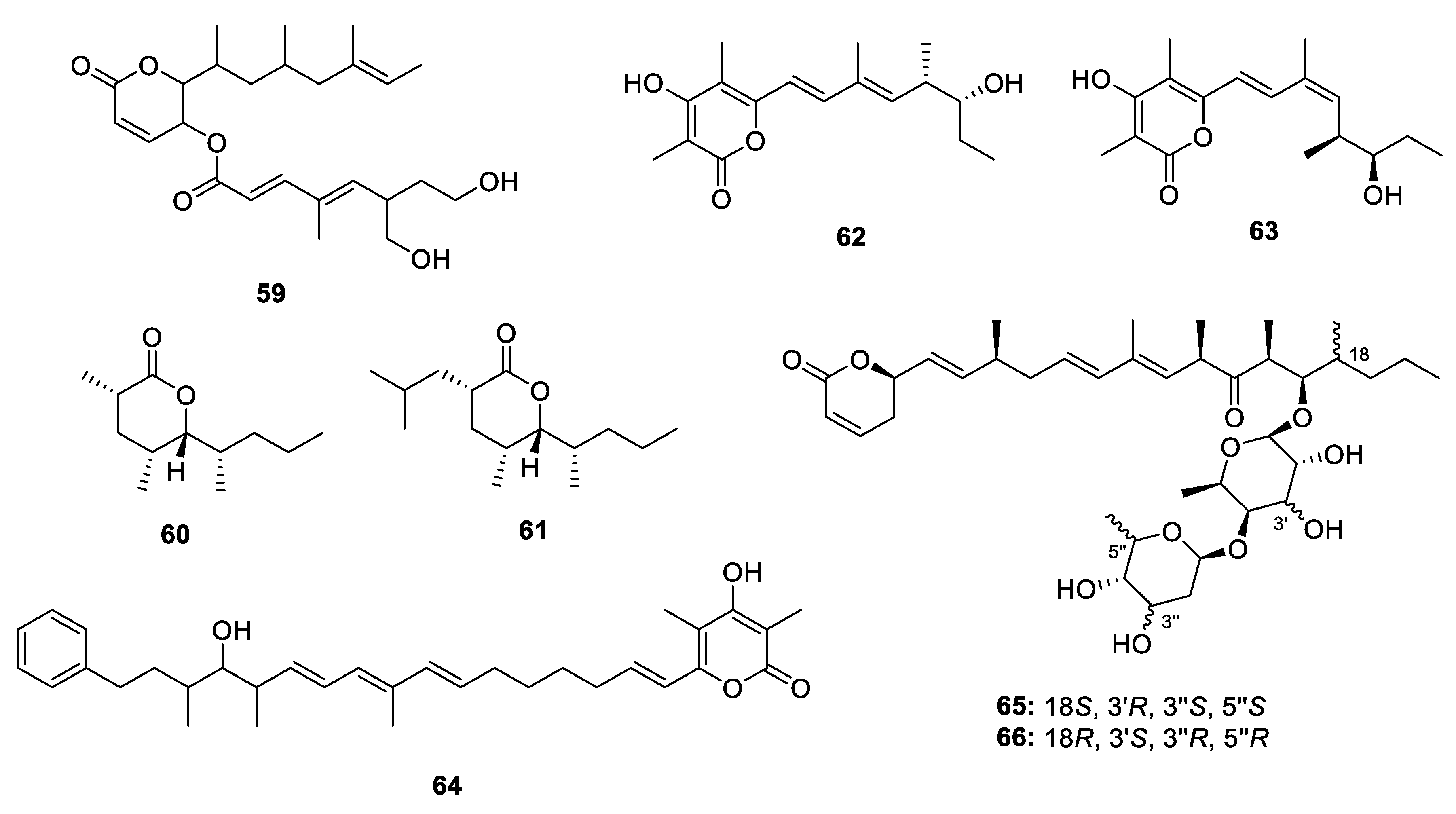
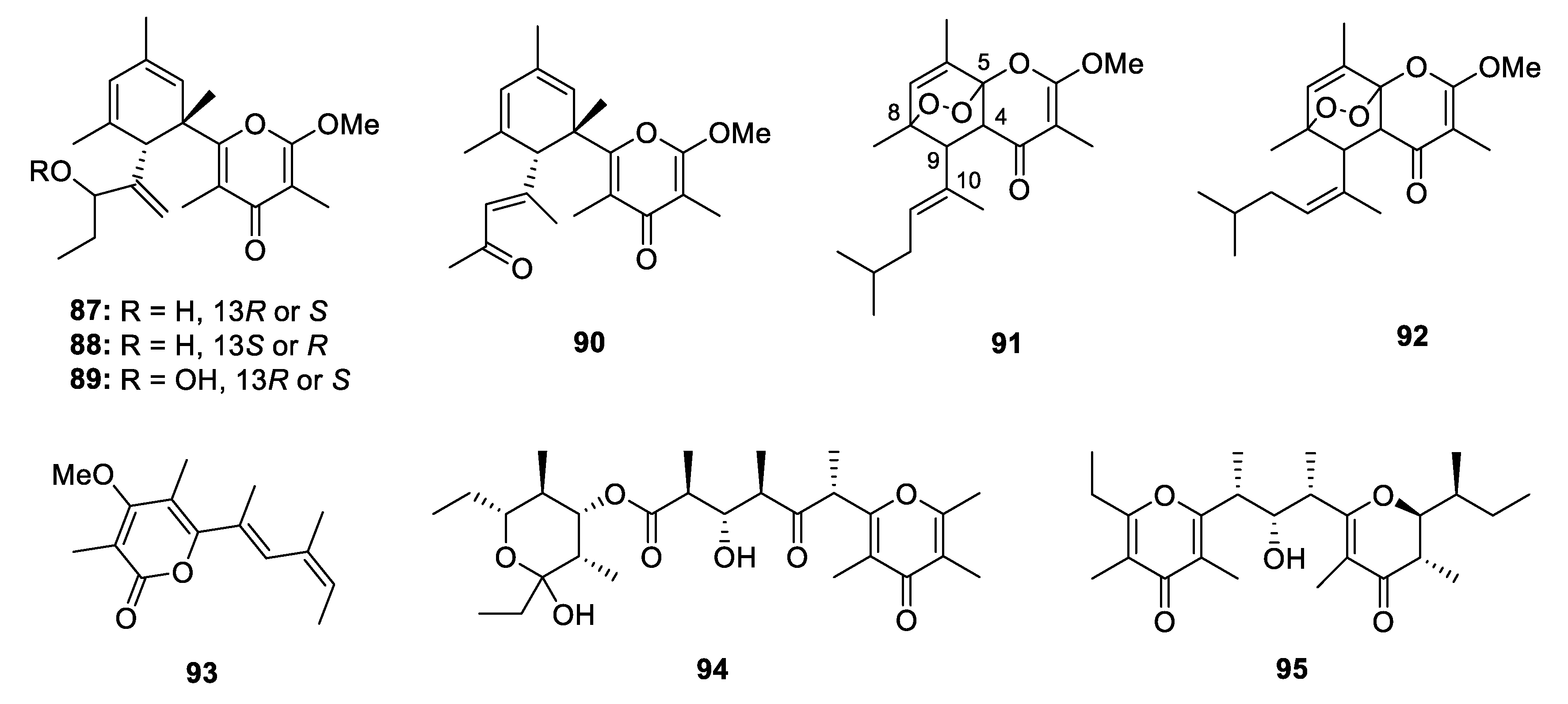
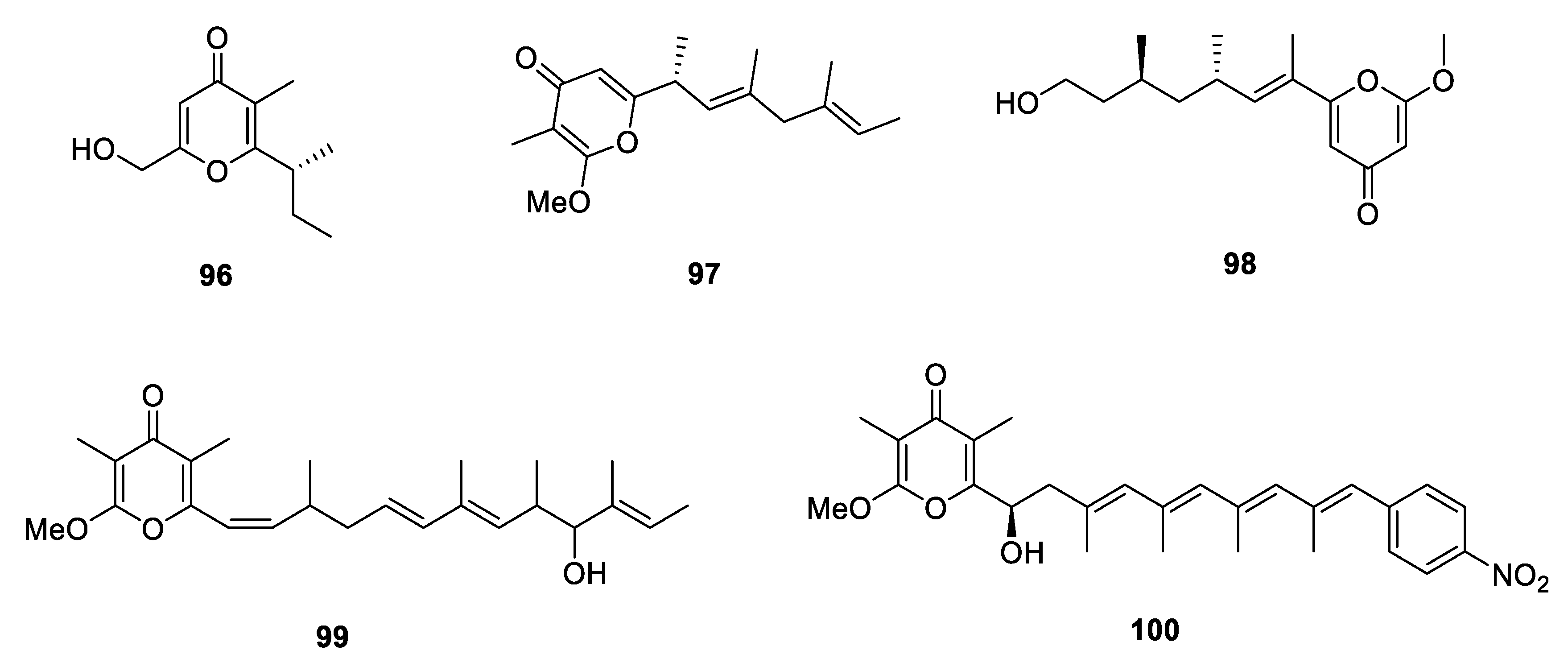
Metabolites Containing 2-Pyrone(s)
Metabolites Containing 4-Pyrone(s)
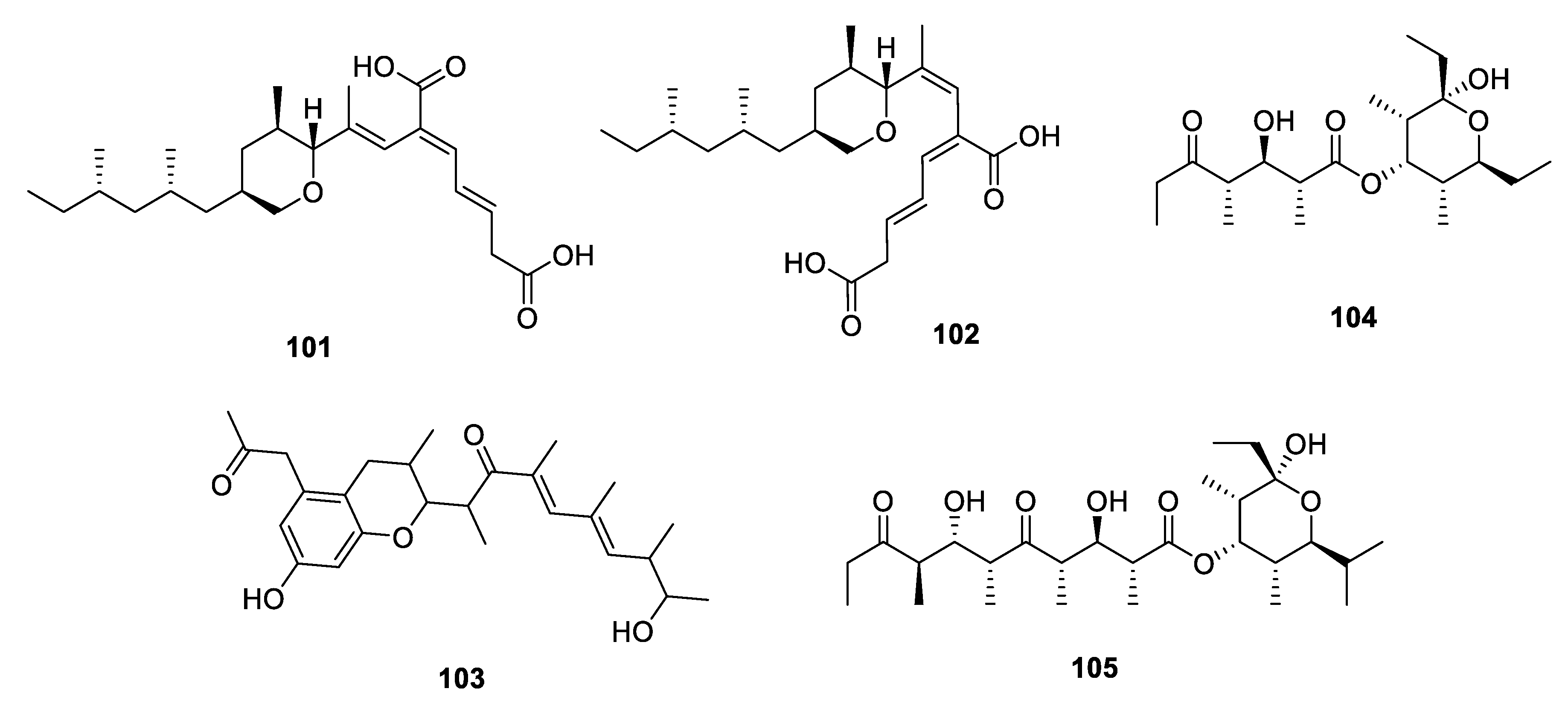
Metabolites Containing Hydrogenated Pyran(s)
2.2.3. Furan-Related Metabolites
2.2.4. Metabolites Containing both Pyran and Furan
2.2.5. Other Heterocyclic Metabolites
2.3. Macrocyclic Metabolites
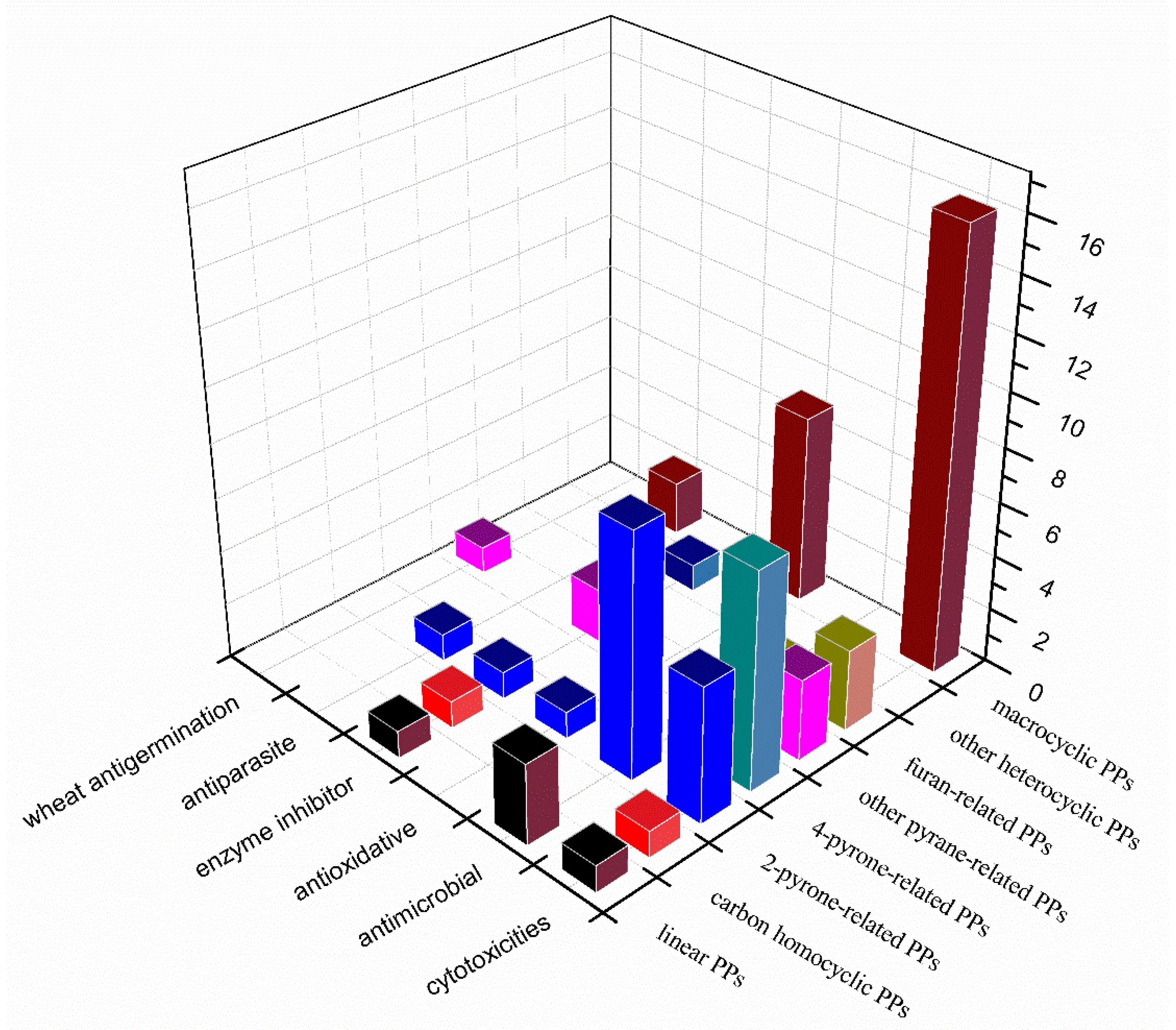
3. Summary
4. Conclusions and Outlook
Author Contributions
Funding
Conflicts of Interest
References
- Müller, W.E.G. Marine Molecular Biotechnology; Springer: Berlin, Germany, 2006; Chapter 1.2; Volume 71, pp. 570–575. [Google Scholar]
- Ireland, C.; Faulkner, D.J. Tridachione, a propionate-derived metabolite of the opisthobranch mollusc Tridachiella diomedea. J. Am. Chem. Soc. 1978, 100, 1002–1003. [Google Scholar]
- Davies-Coleman, M.T.; Garson, M.J. Marine polypropionates. Nat. Prod. Rep. 1998, 15, 477–493. [Google Scholar] [PubMed]
- Gunasekera, S.P.; Paul, G.K.; Longley, R.E.; Isbrucker, R.A.; Pomponi, S.A. Five new discodermolide analogues from the marine sponge Discodermia species. J. Nat. Prod. 2002, 65, 1643–1648. [Google Scholar] [PubMed]
- Gunasekera, S.P.; Gunasekera, M.; Longley, R.E.; Schulte, G.K. Discodermolide: A new bioactive polyhydroxylated lactone from the marine sponge Discodermia dissolute. J. Org. Chem. 1990, 55, 4912. [Google Scholar]
- Nuzzo, G.; Cutignano, A.; Moles, J.; Avila, C.; Fontana, A. Exiguapyrone and exiguaone, new polypropionates from the Mediterranean cephalaspidean mollusc Haminoea exigua. Tetrahedron Lett. 2016, 57, 71–74. [Google Scholar]
- Napolitano, J.G.; Souto, M.L.; Fernández, J.J.; Norte, M. Micromelones A and B, noncontiguous polypropionates from Micromelo undata. J. Nat. Prod. 2008, 71, 281–284. [Google Scholar]
- Bromley, C.L.; Popplewell, W.L.; Pinchuck, S.C.; Hodgson, A.N.; Davies-Coleman, M.T. Polypropionates from the South African marine mollusk Siphonaria oculus. J. Nat. Prod. 2012, 75, 497–501. [Google Scholar]
- Beukes, D.R.; Davies-Coleman, M.T. Novel polypropionates from the South African marine mollusk Siphonaria capensis. Tetrahedron 1999, 55, 4051–4056. [Google Scholar]
- Liu, Z.; Wang, Q.; Li, S.; Cui, H.; Sun, Z.; Chen, D.; Lu, Y.; Liu, H.; Zhang, W. Polypropionate derivatives with Mycobacterium tuberculosis protein tyrosine phosphatase B inhibitory activities from the deep-sea-derived fungus Aspergillus fischeri FS452. J. Nat. Prod. 2019, 82, 3440–3449. [Google Scholar]
- Lin, S.; Wu, Y.Z.; Chen, K.Y.; Ye, J.; Yang, X.W.; Zhang, W.D. Polyketides from the fungus Penicillium decumbens. J. Asian Nat. Prod. Res. 2018, 20, 445–450. [Google Scholar]
- Babadi, Z.K.; Sudarman, E.; Ebrahimipour, G.H.; Primahana, G.; Stadler, M.; Wink, J. Structurally diverse metabolites from the rare actinobacterium Saccharothrix xinjiangensis. J. Antibiot. 2020, 73, 48–55. [Google Scholar]
- Jang, Y.-W.; Lee, I.-K.; Kim, Y.-S.; Lee, S.; Lee, H.-J.; Yu, S.; Yun, B.-S. Xylarinic acids A and B, new antifungal polypropionates from the fruiting body of Xylaria polymorpha. J. Antibiot. 2007, 60, 696–699. [Google Scholar]
- Fletcher, M.T.; Chow, S.; Lambert, L.K.; Gallagher, O.P.; Cribb, B.W.; Allsopp, P.G.; Moore, C.J.; Kitching, W. 4,6,8,10,16-Penta-and 4,6,8,10,16,18-hexamethyldocosanes from the cane beetle Antitrogus parvulus-cuticular hydrocarbons with unprecedented structure and stereochemistry. Org. Lett. 2003, 26, 5083–5086. [Google Scholar]
- Chow, S.; Fletcher, M.T.; Lambert, L.K.; Gallagher, O.P.; Moore, C.J.; Cribb, B.W.; Allsopp, P.G.; Kitching, W. Novel cuticular hydrocarbons from the cane beetle Antitrogus parvulus-4,6,8,10,16-penta-and 4,6,8,10,16,18-hexamethyldocosaness-unprecedented anti-anti-anti-stereochemistry in the 4,6,8,10-methyltetrad. J. Org. Chem. 2005, 70, 1808–1827. [Google Scholar] [PubMed]
- Wu, Q.; Li, S.-W.; Xu, H.; Wang, H.; Hu, P.; Zhang, H.; Luo, C.; Chen, K.-X.; Nay, B.; Guo, Y.-W.; et al. Complex polypropionates from a South China Sea photosynthetic mollusk: Isolation and biomimetic synthesis highlighting novel rearrangements. Angew. Chem. Int. Ed. 2020, 132, 2–10. [Google Scholar]
- Gavagnin, M.; Mollo, E.; Cimino, G.; Ortea, J. A new γ-dihydropyrone-propionate from the Caribbean sacoglossan Tridachia crispate. Tetrahedron Lett. 1996, 37, 4259–4262. [Google Scholar]
- Neuhaus, G.F.; Adpressa, D.A.; Bruhn, T.; Loesgen, S. Polyketides from marine-derived Aspergillus porosus: Challenges and opportunities for determining absolute configuration. J. Nat. Prod. 2019, 82, 2780–2789. [Google Scholar]
- Jouda, J.B.; Fopossi, J.D.; Kengne, F.M.; Djama Mbazoa, C.; Golz, C.; Strohmann, C.; Fogue, S.K.; Wandji, J. Secondary metabolites from Aspergillus japonicus CAM231, an endophytic fungus associated with Garcinia preussii. Nat. Prod. Res. 2017, 31, 861–869. [Google Scholar]
- De Oliveira, C.M.; Silva, G.H.; Regasini, L.O.; Flausino, O.; Lopez, S.N.; Abissi, B.M.; Berlinck, R.G.; Sette, L.D.; Bonugli-Santos, R.C.; Rodrigues, A.; et al. Xylarenones C-E from an endophytic fungus isolated from Alibertia macrophylla. J. Nat. Prod. 2011, 74, 1353–1357. [Google Scholar]
- Oh, D.-C.; Gontang, E.A.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Salinipyrones and pacificanones, mixed-precursor polyketides from the marine actinomycete Salinispora pacifica. J. Nat. Prod. 2008, 71, 570–575. [Google Scholar]
- Sun, Y.; Tomura, T.; Sato, J.; Iizuka, T.; Fudou, R.; Ojika, M. Isolation and biosynthetic analysis of haliamide, a new PKS-NRPS hybrid metabolite from the marine myxobacterium Haliangium ochraceum. Molecules 2016, 21, 59–66. [Google Scholar] [PubMed] [Green Version]
- Okanya, P.W.; Mohr, K.I.; Gerth, K.; Kessler, W.; Jansen, R.; Stadler, M.; Muller, R. Hyafurones, hyapyrrolines, and hyapyrones: Polyketides from Hyalangium minutum. J. Nat. Prod. 2014, 77, 1420–1429. [Google Scholar] [PubMed]
- Cutignano, A.; Blihoghe, D.; Fontana, A.; Villani, G.; d’Ippolito, G.; Cimino, G. Fusaripyrones, novel polypropionates from the Mediterranean mollusc Haminoea fusari. Tetrahedron 2007, 63, 12935–12939. [Google Scholar]
- Smith, A.B.; Lamarche, M.L.; Falcone-Hindley, M. Solution structure of (+)-discodermolide. Org. Lett. 2001, 3, 695–698. [Google Scholar] [PubMed]
- Gunasekera, S.P.; Gunasekera, M.; Longley, R.E.; Schulte, G.K. Discodermolide—A new bioactive polyhydroxylated lactone from the marine sponge Discodermia-dissoluta. J. Org. Chem. 1991, 56, 1346. [Google Scholar]
- Gunasekera, S.P.; Cranick, S.; Longley, R.E. Immunosuppressive compounds from a deep water marine sponge, Agelas flabelliformis. J. Nat. Prod. 1989, 52, 757–761. [Google Scholar]
- Longley, R.E.; Caddigan, D.; Harmody, D.; Gunasekera, M.; Gunasekera, S.P. Discodermolide—A new, marine-derived immunosuppressive compound. I. In vitro studies. Transplantation 1991, 52, 650–656. [Google Scholar]
- Longley, R.E.; Caddigan, D.; Harmody, D.; Gunasekera, M.; Gunasekera, S.P.; Gunasekera, S.P. Discodermolide—A new, marine-derived immunosuppressive compound. II. In vitro studies. Transplantation 1991, 52, 656–661. [Google Scholar]
- Longley, R.E.; Gunasekera, S.P.; Faherty, D.; McLane, J.; Dumont, F. Immunosuppression by discodermolide. Ann. N. Y. Acad. Sci. 1993, 696, 94–107. [Google Scholar]
- Ding, L.; Ren, L.; Li, S.; Song, J.; Han, Z.; He, S.; Xu, S. Production of new antibacterial 4-hydroxy-alpha-pyrones by a marine fungus Aspergillus niger cultivated in solid medium. Mar. Drugs 2019, 17, 344–351. [Google Scholar]
- Li, T.X.; Meng, D.D.; Wang, Y.; An, J.L.; Bai, J.F.; Jia, X.W.; Xu, C.P. Antioxidant coumarin and pyrone derivatives from the insect-associated fungus Aspergillus versicolor. Nat. Prod. Res. 2020, 34, 1360–1365. [Google Scholar] [PubMed]
- Yamazaki, H.; Takahashi, K.; Iwakura, N.; Abe, T.; Akaishi, M.; Chiba, S.; Namikoshi, M.; Uchida, R. A new protein tyrosine phosphatase 1B inhibitory alpha-pyrone-type polyketide from Okinawan plant-associated Aspergillus sp. TMPU1623. J. Antibiot. 2018, 71, 745–748. [Google Scholar]
- Gao, H.; Li, G.; Peng, X.P.; Lou, H.X. Fupyrones A and B, two new alpha-pyrones from an endophytic fungus, Fusarium sp. F20. Nat. Prod. Res. 2020, 34, 335–340. [Google Scholar]
- Pedras, M.S.; Chumala, P.B. Phomapyrones from blackleg causing phytopathogenic fungi: Isolation, structure determination, biosyntheses and biological activity. Phytochemistry 2005, 66, 81–87. [Google Scholar]
- Asai, T.; Luo, D.; Yamashita, K.; Oshima, Y. Structures and biomimetic synthesis of novel α-pyrone polyketides of an endophytic Penicillium sp. in Catharanthus roseus. Org. Lett. 2013, 15, 1020–1023. [Google Scholar] [PubMed]
- Li, H.; Hu, J.; Wei, H.; Solomon, P.S.; Vuong, D.; Lacey, E.; Stubbs, K.A.; Piggott, A.M.; Chooi, Y.-H. Chemical ecogenomics-guided discovery of phytotoxic α-pyrones from the fungal wheat pathogen Parastagonospora nodorum. Org. Lett. 2018, 20, 6148–6152. [Google Scholar]
- Zhou, X.M.; Zheng, C.J.; Song, X.P.; Han, C.R.; Chen, W.H.; Chen, G.Y. Antibacterial alpha-pyrone derivatives from a mangrove-derived fungus Stemphylium sp. 33231 from the South China Sea. J. Antibiot. 2014, 67, 401–403. [Google Scholar]
- Tomikawa, T.; Shin-ya, K.; Furihata, K.; Kinoshita, T.; Miyajima, A.; Seto, H.; Hayakawa, Y. Rasfonin, a new apoptosis inducer in ras-dependent cells from Talaromyces sp. J. Antibiot. 2000, 53, 848–850. [Google Scholar]
- Sato, S.; Iwata, F.; Yamada, S.; Kawahara, H. 3,6,7-Tri-epi-invictolide, a diastereomer of queen recognition pheromone, and its analog from a marine-derived actinomycete. J. Antibiot. 2011, 64, 385–389. [Google Scholar]
- Řezanka, T.; Dvořáková, R. Polypropionate lactones of deoxysugars glycosides from slime mold Lycogala epidendrum. Phytochemistry 2003, 63, 945–952. [Google Scholar]
- Cueto, M.; D’Croz, L.; Maté, J.L.; San-Martín, A.; Darias, J. Elysiapyrones from Elysia diomedea. Do such metabolites evidence an enzymatically assisted electrocyclization cascade for the biosynthesis of their bicyclo[4.2.0]octane core? Org. Lett. 2005, 7, 415–418. [Google Scholar] [PubMed]
- Manzo, E.; Ciavatta, M.L.; Gavagnin, M.; Mollo, E.; Wahidulla, S.; Cimino, G. New γ-pyrone propionates from the Indian Ocean sacoglossan Placobranchus ocellatus. Tetrahedron Lett. 2005, 46, 465–468. [Google Scholar]
- Miller, A.K.; Trauner, D. Mining the tetraene manifold: Total synthesis of complex pyrones from Placobranchus ocellatus. Angew. Chem. Int. Ed. 2005, 44, 4602–4606. [Google Scholar]
- Carbone, M.; Muniain, C.; Castelluccio, F.; Iannicelli, O.; Gavagnin, M. First chemical study of the sacoglossan Elysia patagonica: Isolation of a γ-pyrone propionate hydroperoxide. Biochem. Syste. Ecol. 2013, 49, 172–175. [Google Scholar]
- Carbone, M.; Ciavatta, M.L.; Wang, J.R.; Cirillo, I.; Mathieu, V.; Kiss, R.; Mollo, E.; Guo, Y.W.; Gavagnin, M. Extending the record of bis-gamma-pyrone polypropionates from marine pulmonate mollusks. J. Nat. Prod. 2013, 76, 2065–2073. [Google Scholar]
- Zhou, Z.F.; Li, X.L.; Yao, L.G.; Li, J.; Gavagnin, M.; Guo, Y.W. Marine bis-gamma-pyrone polypropionates of onchidione family and their effects on the XBP1 gene expression. Bioorg. Med. Chem. Lett. 2018, 28, 1093–1096. [Google Scholar]
- Carbone, M.; Gavagnin, M.; Mattia, C.A.; Lotti, C.; Castelluccio, F.; Pagano, B.; Mollo, E.; Guo, Y.-W.; Cimino, G. Structure of onchidione, a bis-γ-pyrone polypropionate from a marine pulmonate mollusk. Tetrahedron 2009, 65, 4404–4409. [Google Scholar]
- Wang, J.-R.; Carbone, M.; Gavagnin, M.; Mándi, A.; Antus, S.; Yao, L.-G.; Cimino, G.; Kurtán, T.; Guo, Y.-W. Assignment of absolute configuration of bis-γ-pyrone polypropionates from marine pulmonate molluscs. Eur. J. Org. Chem. 2012, 1107–1111. [Google Scholar]
- Chen, D.L.; Zheng, W.; Feng, J.; Ma, G.X.; Liu, Y.Y.; Xu, X.D. A new bis-gamma-pyrone polypropionate from a marine pulmonate mollusc Onchidium struma. J. Asian. Nat. Prod. Res. 2019, 21, 384–390. [Google Scholar]
- Li, S.W.; Cuis, W.X.; Huan, X.J.; Gavagnin, M.; Mollo, E.; Miao, Z.H.; Yao, L.G.; Li, X.W.; Guo, Y.W. A new bis-gamma-pyrone polypropionate of onchidiol family from marine pulmonate mollusk Onchidium sp. Nat. Prod. Res. 2020, 34, 1971–1976. [Google Scholar]
- Fu, X.; Hong, E.P.; Schmitz, F.J. New polypropionate pyrones from the Philippine sacoglossan mollusk Placobranchus ocellatus. Tetrahedron 2000, 56, 8989–8993. [Google Scholar]
- Cutignano, A.; Fontana, A.; Rennzulli, L.; Cimino, G. Placidenes C-F, novel α-pyrone propionates from the Mediterranean Sacoglossan Placida dendritica. J. Nat. Prod. 2003, 66, 1399–1401. [Google Scholar] [PubMed]
- Brecknell, D.J.; Collett, L.A.; Davies-Coleman, M.T.; Garson, M.J.; Jones, D.D. New non-contiguous polypropionates from marine molluscs: A comment on their natural products status. Tetrahedron 2000, 56, 2497–2502. [Google Scholar]
- Esposito, G.; Teta, R.; Della Sala, G.; Pawlik, J.R.; Mangoni, A.; Costantino, V. Isolation of smenopyrone, a bis-gamma-pyrone polypropionate from the Caribbean sponge Smenospongia aurea. Mar. Drugs 2018, 16, 285–293. [Google Scholar]
- Peng, X.P.; Li, G.; Ji, L.X.; Li, Y.X.; Lou, H.X. Acrepyrone A, a new gamma-pyrone derivative from an endophytic fungus, Acremonium citrinum SS-g13. Nat. Prod. Res. 2020, 34, 1091–1096. [Google Scholar]
- Choi, H.G.; Song, J.H.; Park, M.; Kim, S.; Kim, C.E.; Kang, K.S.; Shim, S.H. Neuroprotective gamma-pyrones from Fusarium Solani JS-0169: Cell-based identification of active compounds and an informatics approach to predict the mechanism of action. Biomolecules 2020, 10, 91–101. [Google Scholar]
- Hayakawa, Y.; Saito, J.; Izawa, M.; Shin-ya, K. Actinopyrone D, a new downregulator of the molecular chaperone GRP78 from Streptomyces sp. J. Antibiot. 2014, 67, 831–834. [Google Scholar]
- Liu, S.H.; Xu, M.D.; Zhang, H.; Qi, H.; Zhang, J.; Liu, C.X.; Wang, J.D.; Xiang, W.S.; Wang, X.J. New cytotoxic spectinabilin derivative from ant-associated Streptomyces sp. 1H-GS5. J. Antibiot. 2016, 69, 128–131. [Google Scholar]
- Jimenez-Romero, C.; Gonzalez, K.; Rodriguez, A.D. Dolabriferols B and C, non-contiguous polypropionate esters from the tropical sea hare Dolabrifera dolabrifera. Tetrahedron Lett. 2012, 53, 6641–6645. [Google Scholar]
- Rovirosa, J.; Quezada, E. San-Martin, new polypropionates of Siphonaria-lessoni from Chilean coasts. Bol. Soc. Chil. Quim. 1991, 36, 233. [Google Scholar]
- Capon, R.J.; Faulkner, D.J. Metabolites of the pulmonate Siphonaria lessoni. J. Org. Chem. 1984, 49, 2506–2508. [Google Scholar]
- Sato, S.; Iwata, F.; Mukai, T.; Yamada, S.; Takeo, J.; Abe, A.; Kawahara, H. Indoxamycins A-F. cytotoxic tricycklic polypropionates from a marine-derived actinomycete. J. Org. Chem. 2009, 74, 5502–5509. [Google Scholar] [PubMed]
- Liu, J.T.; Wu, W.; Cao, M.J.; Yang, F.; Lin, H.W. Trienic alpha-pyrone and ochratoxin derivatives from a sponge-derived fungus Aspergillus ochraceopetaliformis. Nat. Prod. Res. 2018, 32, 1791–1797. [Google Scholar] [PubMed]
- Wang, F.; Luo, D.-Q.; Liu, J.-L. Aurovertin E, a new polyene pyrone from the basidiomycete Albatrellus confluens. J. Antibiot. 2005, 58, 412–415. [Google Scholar]
- Bu, Y.Y.; Yamazaki, H.; Takahashi, O.; Kirikoshi, R.; Ukai, K.; Namikoshi, M. Penicyrones A and B, an epimeric pair of alpha-pyrone-type polyketides produced by the marine-derived Penicillium sp. J. Antibiot. 2016, 69, 57–61. [Google Scholar]
- Choo, S.-J.; Park, H.-R.; Ryoo, J.-J.; Kim, J.-P.; Yun, B.-S.; Kim, C.-J.; Shim-ya, K.; Yoo, I.-D. Deoxyverrucosidin, a novel GRP78/BiP down-regulator, produced by Penicillium sp. J. Antibiot. 2005, 58, 210–213. [Google Scholar]
- Li, J.; Wang, Y.; Hao, X.; Li, S.; Jia, J.; Guan, Y.; Peng, Z.; Bi, H.; Xiao, C.; Cen, S.; et al. Broad-spectrum antiviral natural products from the marine-derived Penicillium sp. IMB17-046. Molecules 2019, 24, 2821–2831. [Google Scholar]
- Ueda, J.; Hashimoto, J.; Nagai, A.; Nakashima, T.; Komaki, H.; Anzai, K.; Harayama, S.; Doi, T.; Takahashi, T.; Nagasawa, K.; et al. New aureothin derivative, alloaureothin, from Streptomyces sp. MM23. J. Antibiot. 2007, 60, 321–324. [Google Scholar]
- Kawamura, T.; Fujimaki, T.; Hamanaka, N.; Torii, K.; Kobayashi, H.; Takahashi, Y.; Igarashi, M.; Kinoshita, N.; Nishimura, Y.; Tashiro, E.; et al. Isolation and structure elucidation of a novel androgen antagonist, arabilin, produced by Streptomyces sp. MK756-CF1. J. Antibiot. 2010, 63, 601–605. [Google Scholar]
- Morris, B.D.; Smyth, R.R.; Foster, S.P.; Hoffmann, M.P.; Roelofs, W.L.; Franke, S.; Francke, W. Vittatalactone, a β-lactone from the striped cucumber beetle, Acalymma vittatum. J. Nat. Prod. 2005, 68, 26–30. [Google Scholar]
- Nakashima, T.; Kamiya, Y.; Iwatsuki, M.; Takahashi, Y.; Omura, S. Mangromicins, six new anti-oxidative agents isolated from a culture broth of the actinomycete, Lechevalieria aerocolonigenes K10-0216. J. Antibiot. 2014, 67, 533–539. [Google Scholar]
- Nakashima, T.; Iwatsuki, M.; Ochiai, J.; Kamiya, Y.; Nagai, K.; Matsumoto, A.; Ishiyama, A.; Otoguro, K.; Shiomi, K.; Takahashi, Y.; et al. Mangromicins A and B: Structure and antitrypanosomal activity of two new cyclopentadecane compounds from Lechevalieria aerocolonigenes K10-0216. J. Antibiot. 2014, 67, 253–560. [Google Scholar]
- Nakashima, T.; Kamiya, Y.; Iwatsuki, M.; Sato, N.; Takahashi, Y.; Omura, S. Mangromicin C, a new analog of mangromicin. J. Antibiot. 2015, 68, 220–222. [Google Scholar]
- Zampella, A.; Giannini, C.; Debitus, C.; Roussakis, C.; D’Auria, V. New jaspamide derivatives from the marine sponge Jaspis splendans collected in Vanuatu. J. Nat. Prod. 1999, 62, 332–334. [Google Scholar]
- Gala, F.; D’Auria, M.V.; De Marino, S.; Zollo, F.; Smith, C.D.; Copper, J.E.; Zampella, A. New jaspamide derivatives with antimicrofilament activity from the sponge Jaspis splendans. Tetrahedron 2007, 63, 5212–5219. [Google Scholar]
- Gala, F.; D’Auria, M.V.; De Marino, S.; Sepe, V.; Zollo, F.; Smith, C.D.; Copper, J.E.; Zampella, A. Jaspamides H–L, new actin-targeting depsipeptides from the sponge Jaspis splendans. Tetrahedron 2008, 64, 7127–7130. [Google Scholar]
- Gala, F.; D’Auria, M.V.; De Marino, S.; Sepe, V.; Zollo, F.; Smith, C.D.; Keller, S.N.; Zampella, A. Jaspamides M–P: New tryptophan modified jaspamide derivatives from the sponge Jaspis splendans. Tetrahedron 2009, 65, 51–56. [Google Scholar]
- Sorres, J.; Martin, M.T.; Petek, S.; Levaique, H.; Cresteil, T.; Ramos, S.; Thoison, O.; Debitus, C.; Al-Mourabit, A. Pipestelides A-C: Cyclodepsipeptides from the Pacific marine sponge Pipestela candelabra. J. Nat. Prod. 2012, 75, 759–763. [Google Scholar]
- Cimino, G.; Sodano, G. Marine Natural Products−Diversity and Biosynthesis; Springer: Berlin/Heidelberg, Germany, 1993; pp. 77–115. [Google Scholar]
- Fontana, A.; Manzo, E.; Ciavatta, M.L.; Cutignano, A.; Gavagnin, M.; Cimino, G. Biosynthetic studies through feeling experiments in marine organisms. In Handbook of Marine Natural Products; Fattorusso, E., Gerwick, W.H., Taglialatela-Scafati, Eds.; Springer: Berlin, Germany, 2012; pp. 895–946. [Google Scholar]
- Pfeifer, B.A.; Khosla, C. Biosynthesis of polyketides in heterologous hosts. Micobiol. Mol. Biol. Rev. 2001, 65, 106–118. [Google Scholar]
- Inokuma, Y.; Yoshioka, S.; Ariyoshim, J.; Arai, T.; Hitora, Y.; Takada, K.; Matsunaga, S.; Rissanen, K.; Fujita, M. X-ray analysis on the nanogram to microgram scale using porous complexes. Nature 2013, 495, 461–467. [Google Scholar]
- Inokuma, Y.; Yoshioka, S.; Ariyoshi, J.; Arai, T.; Fujita, M. Preparation and guest-uptake protocol for a porous complex useful for ‘crystal-free’ crystallography. Nat. Protoc. 2014, 9, 246–252. [Google Scholar]
- Yoshioka, S.; Inokuma, Y.; Hoshino, M.; Sato, T.; Fujita, M. Absolute structure determination of compounds with axial and planar chirality using the crystalline sponge method. Chem. Sci. 2015, 6, 3765–3768. [Google Scholar]
- Turks, M.; Laclef, S.; Vogel, P. Construction of polypropionate fragments in natural product synthesis. In Stereoselective Synthesis of Drugs and Natural Products; John Wiley & Sons: Hoboken, NJ, USA, 2013; Chapter 10. [Google Scholar]

























{kind=link}
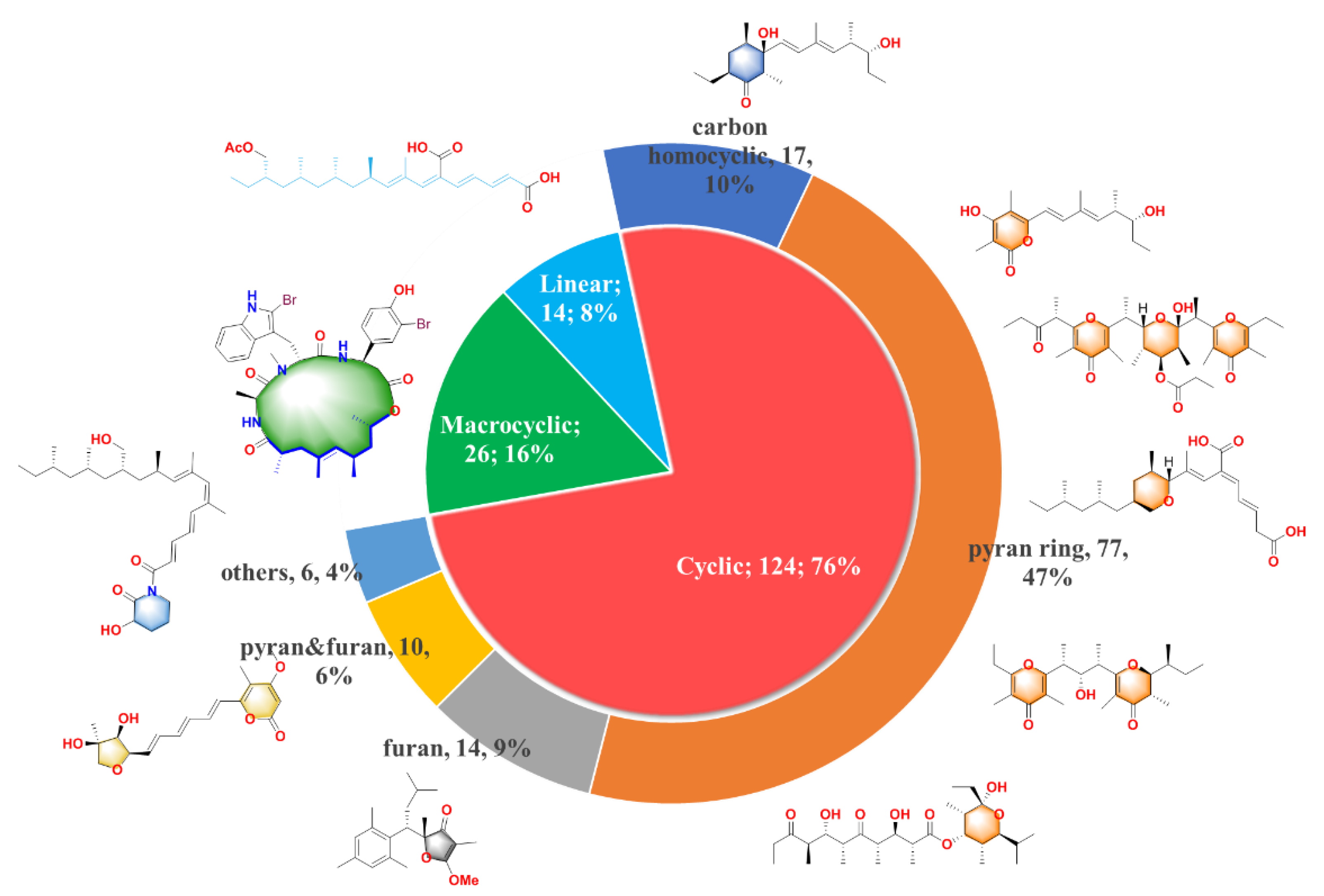{kind=link}
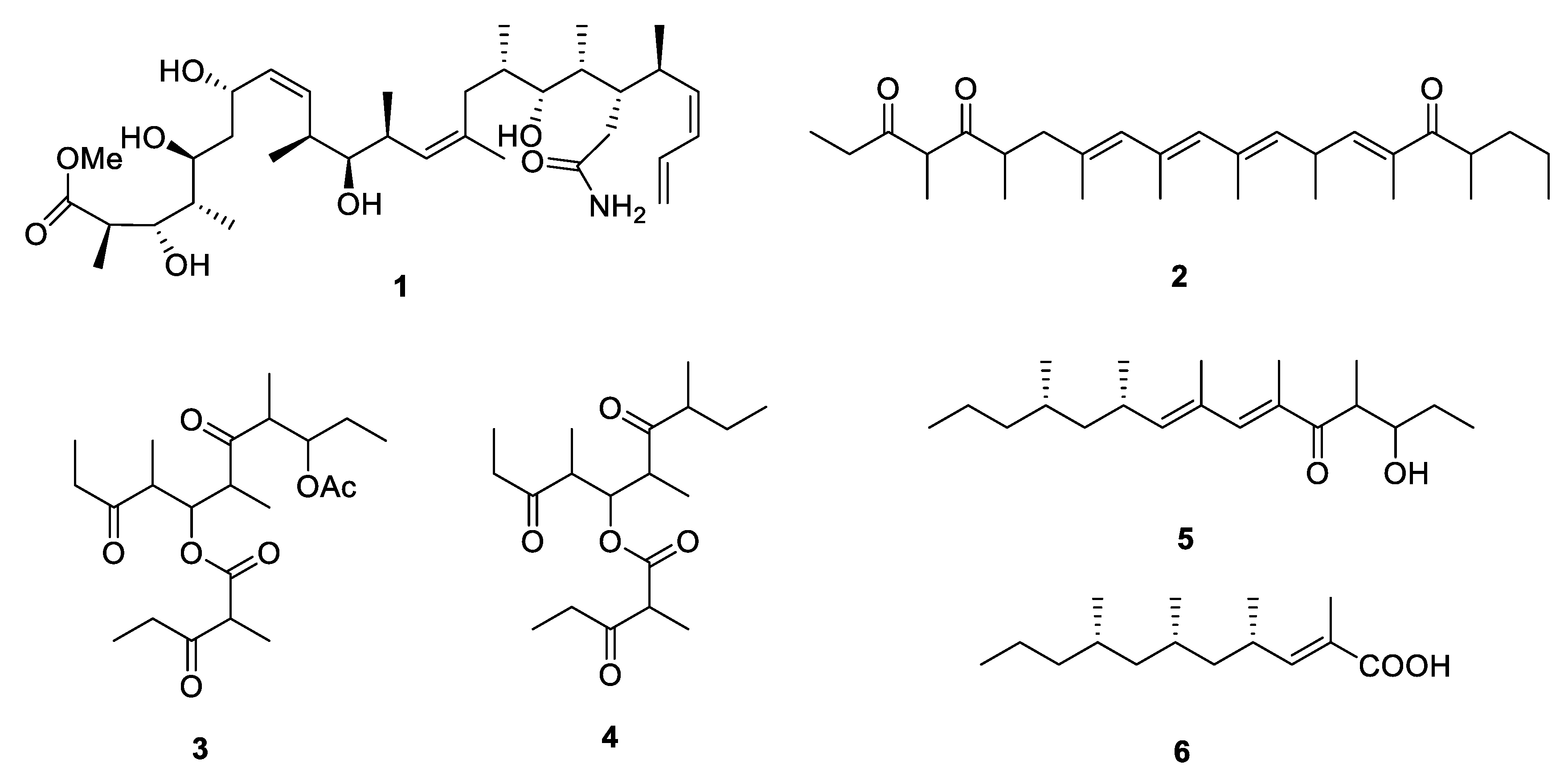{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
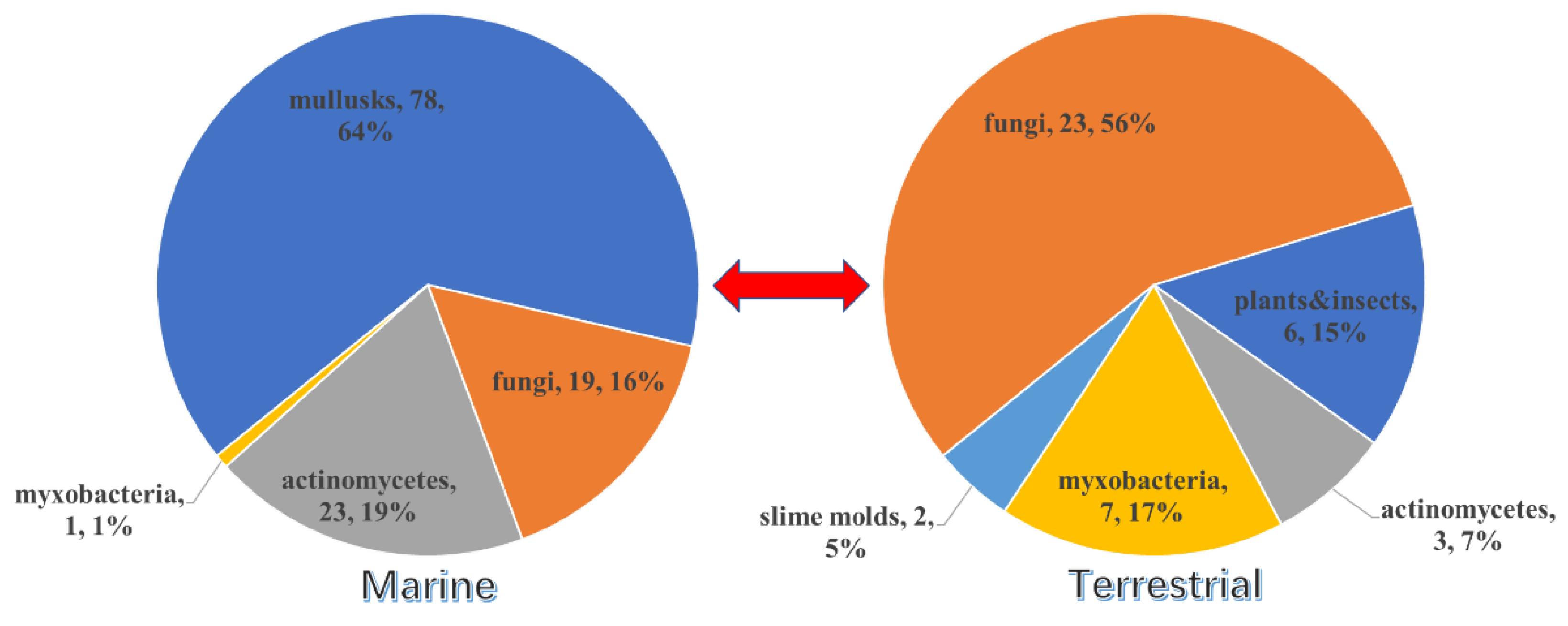
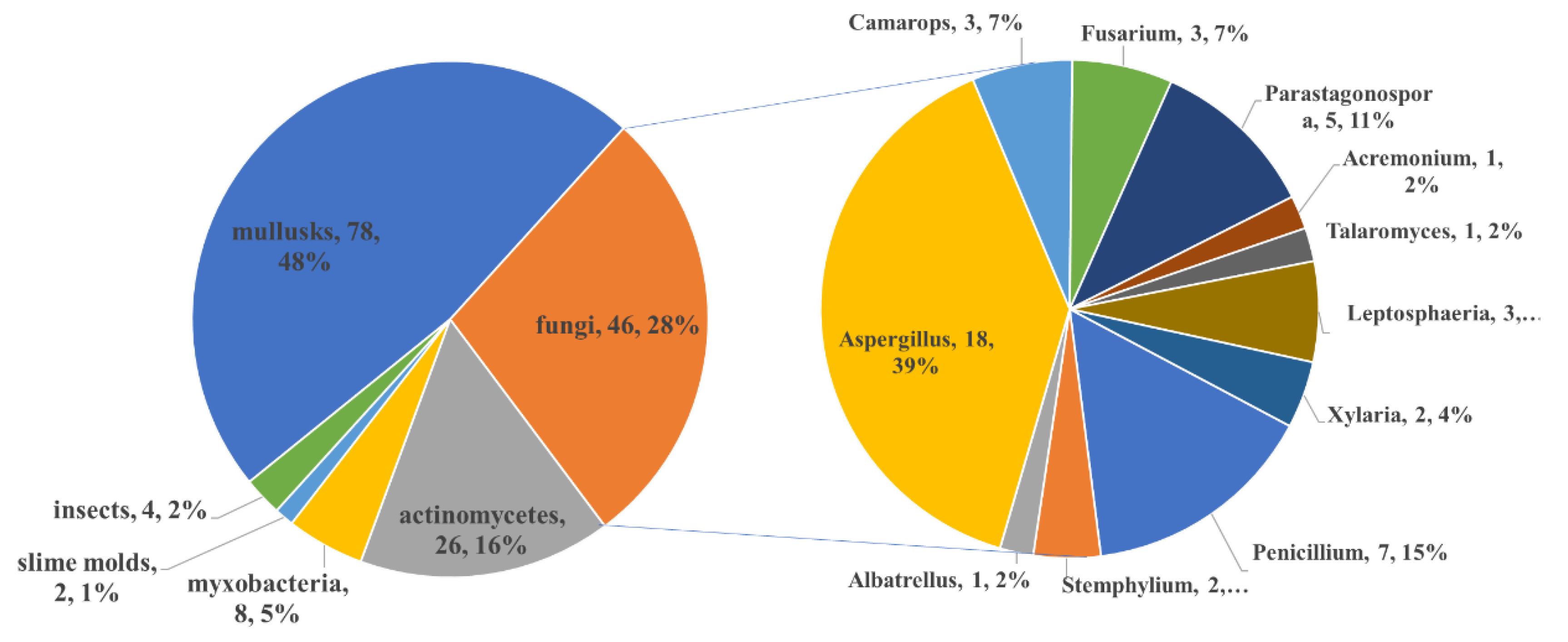
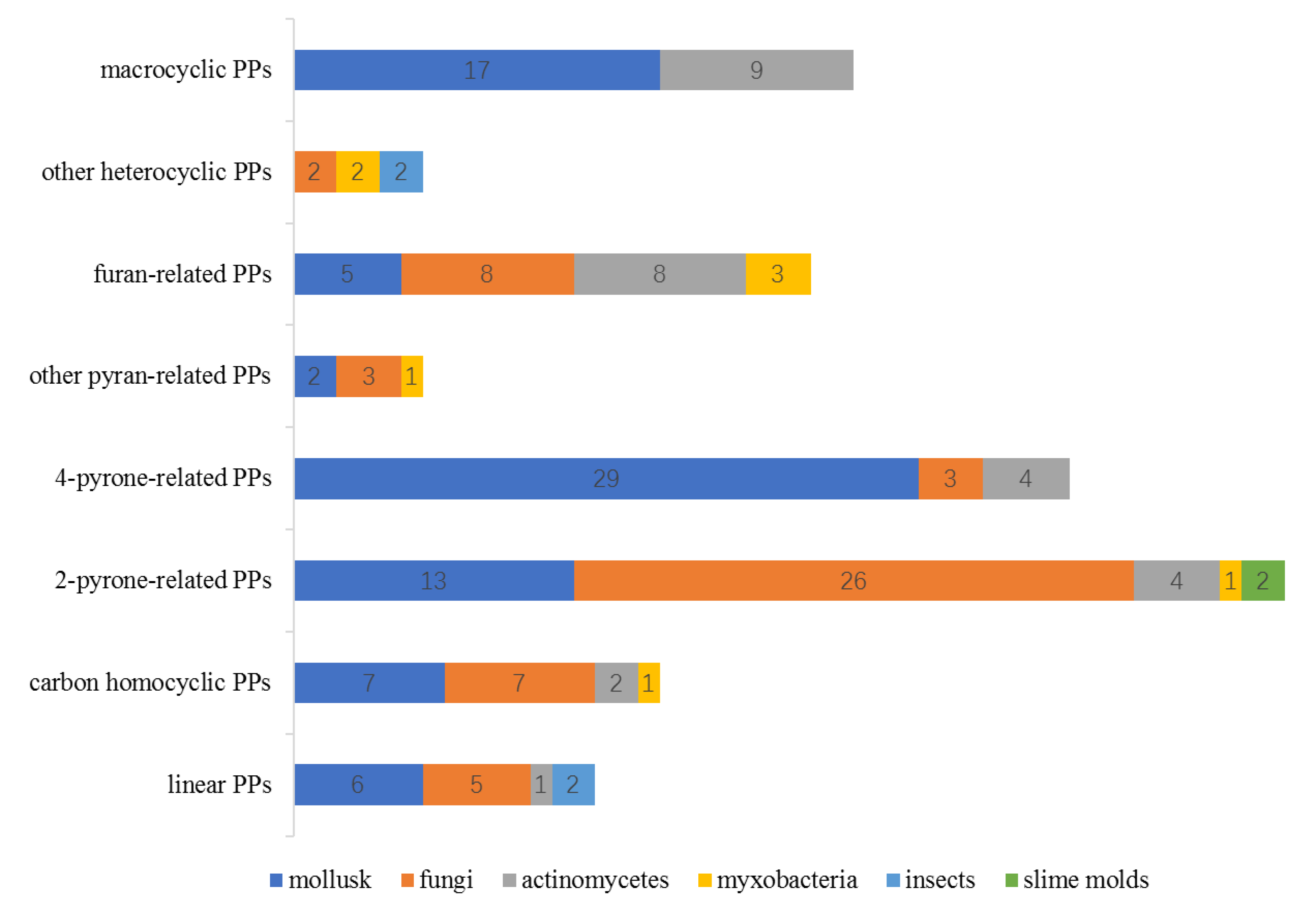
| Mollusk | Fungi | Actinomycetes | Myxobacter-ia | Insects | Slime Molds | |
|---|---|---|---|---|---|---|
| linear PPs | 6 | 5 | 1 | - | 2 | - |
| carbon homocyclic PPs | 7 | 7 | 2 | 1 | - | - |
| 2-pyrone-related PPs | 13 | 26 | 4 | 1 | - | 2 |
| 4-pyrone-related PPs | 29 | 3 | 4 | - | - | - |
| other pyran-related PPs | 2 | 3 | - | 1 | - | - |
| furan-related PPs | 5 | 8 | 8 | 3 | - | - |
| other heterocyclic PPs | - | 2 | - | 2 | 2 | - |
| macrocyclic PPs | 17 | - | 9 | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Liu, H.; Zhang, W. Natural Polypropionates in 1999–2020: An Overview of Chemical and Biological Diversity. Mar. Drugs 2020, 18, 569. https://doi.org/10.3390/md18110569
Liu Z, Liu H, Zhang W. Natural Polypropionates in 1999–2020: An Overview of Chemical and Biological Diversity. Marine Drugs. 2020; 18(11):569. https://doi.org/10.3390/md18110569
Chicago/Turabian StyleLiu, Zhaoming, Hongxin Liu, and Weimin Zhang. 2020. "Natural Polypropionates in 1999–2020: An Overview of Chemical and Biological Diversity" Marine Drugs 18, no. 11: 569. https://doi.org/10.3390/md18110569