Spirulina maxima Derived Pectin Nanoparticles Enhance the Immunomodulation, Stress Tolerance, and Wound Healing in Zebrafish

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Physicochemical Properties and Toxicity of SmPNPs In Vitro and In Vivo
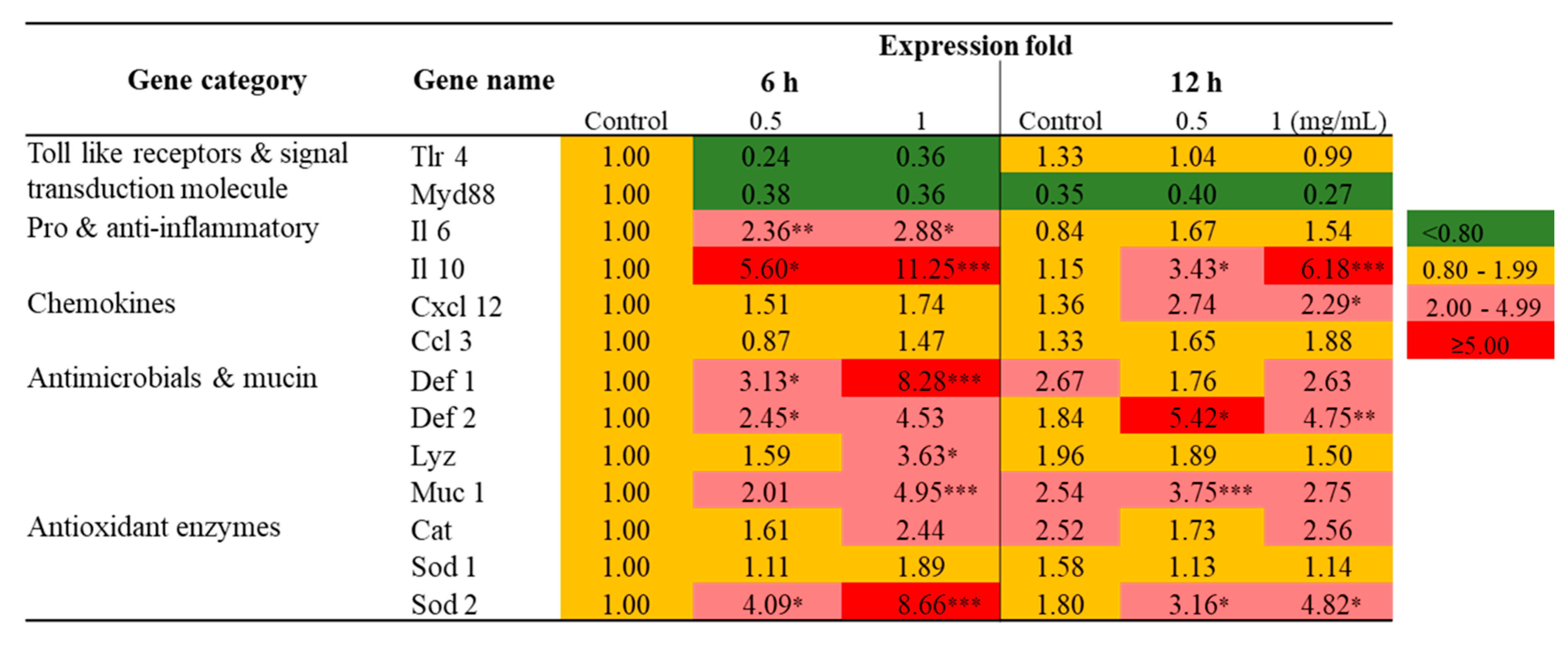
2.2. Transcriptional Profiling of Immune-Related Genes in Raw 264.7 Cells Exposed to SmPNPs
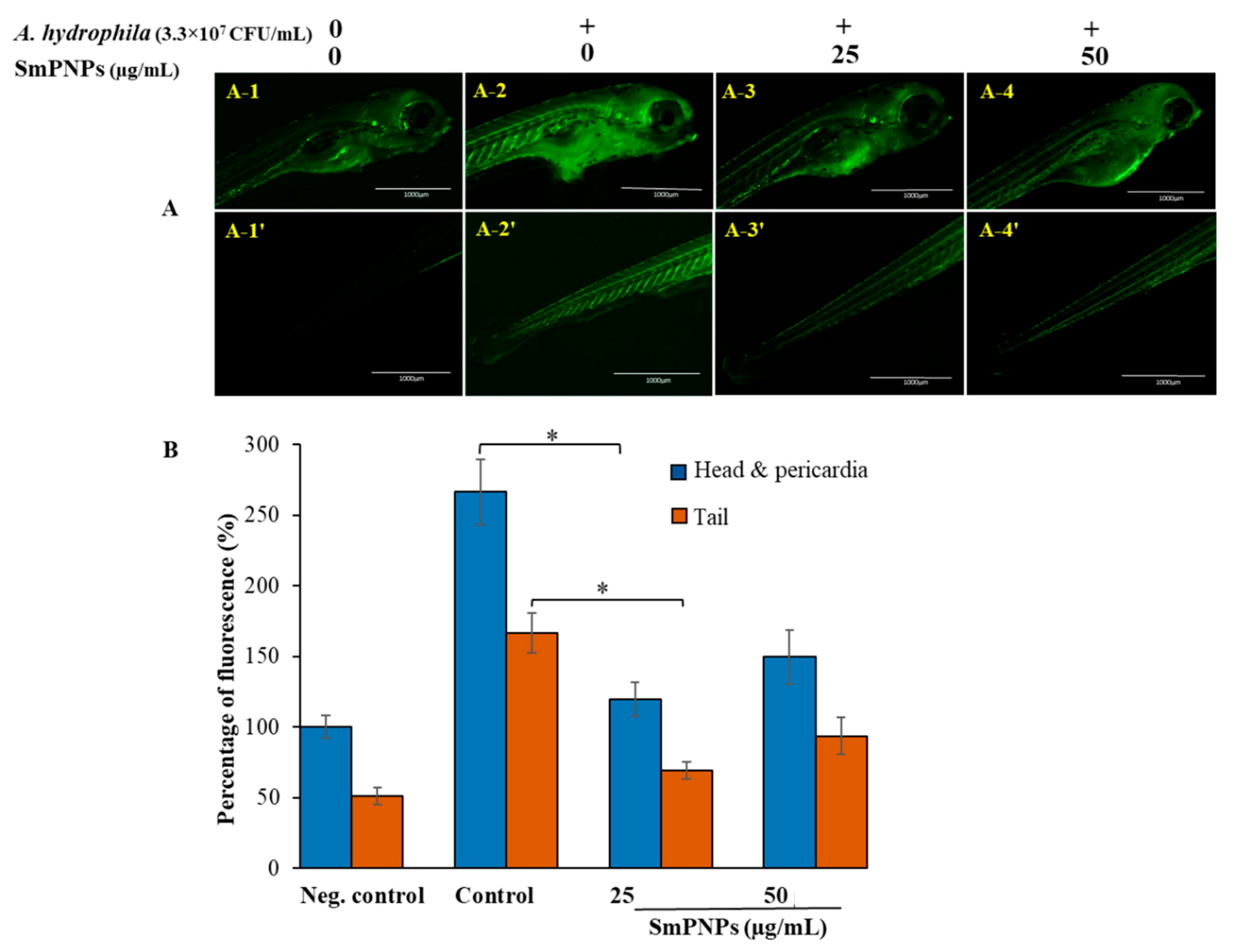
2.3. Reduction of Reactive Oxygen Species (ROS) Levels in Larvae upon Exposure to SmPNPs
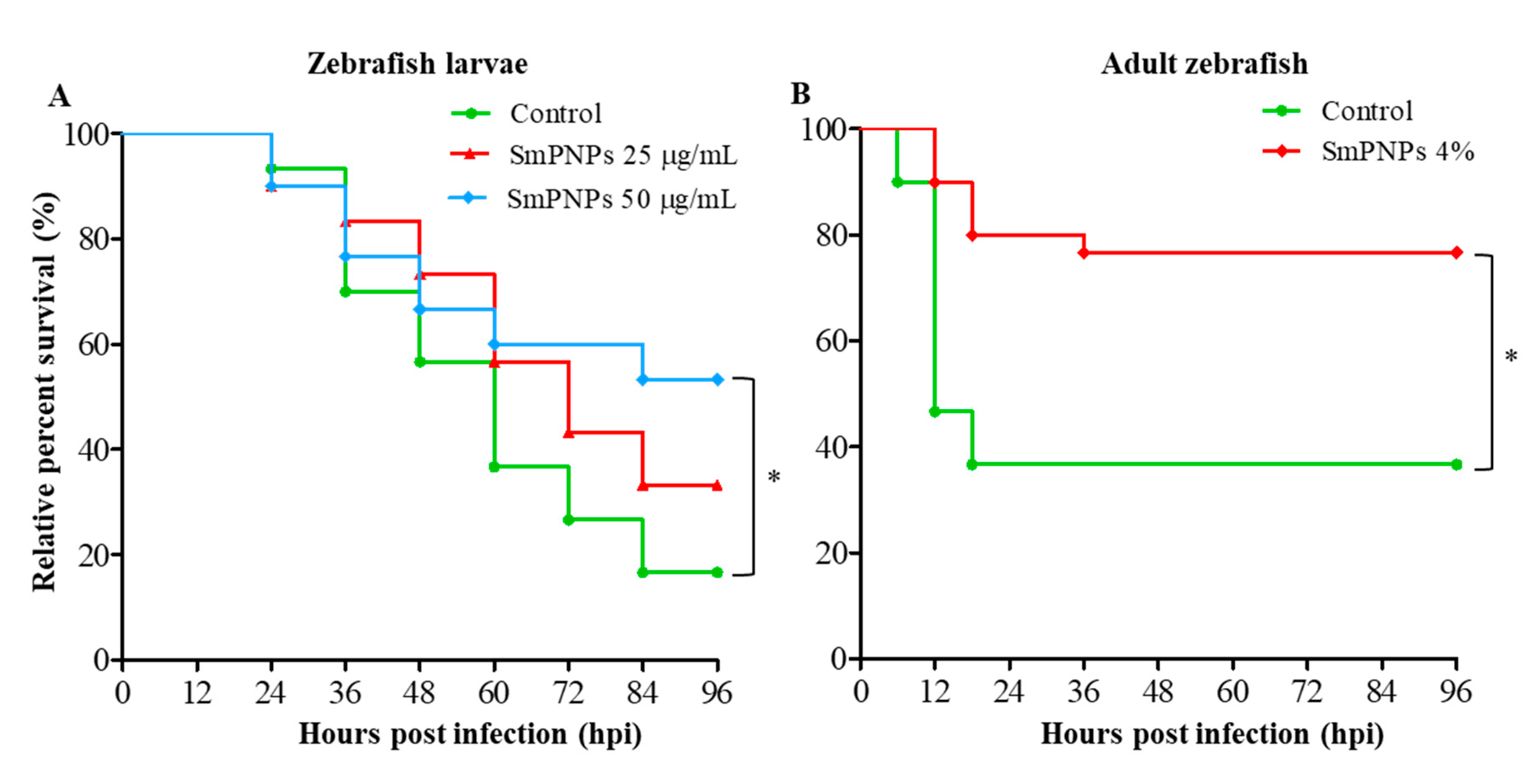
2.4. Disease Resistance of Zebrafish Larvae and Adults Exposed to SmPNPs
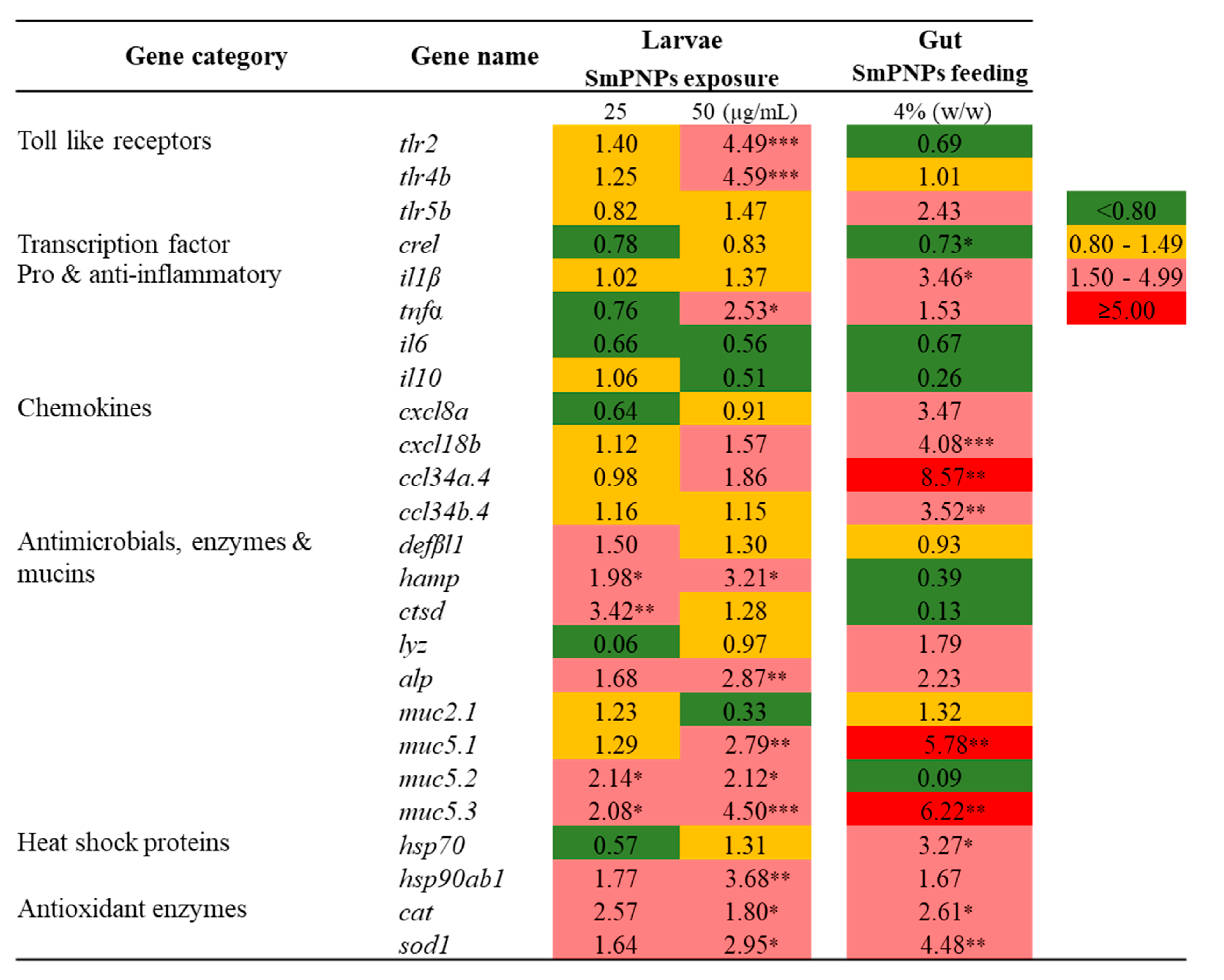
2.5. Transcriptional Profiling of Immune-Related Genes in Zebrafish upon SmPNPs Treatment
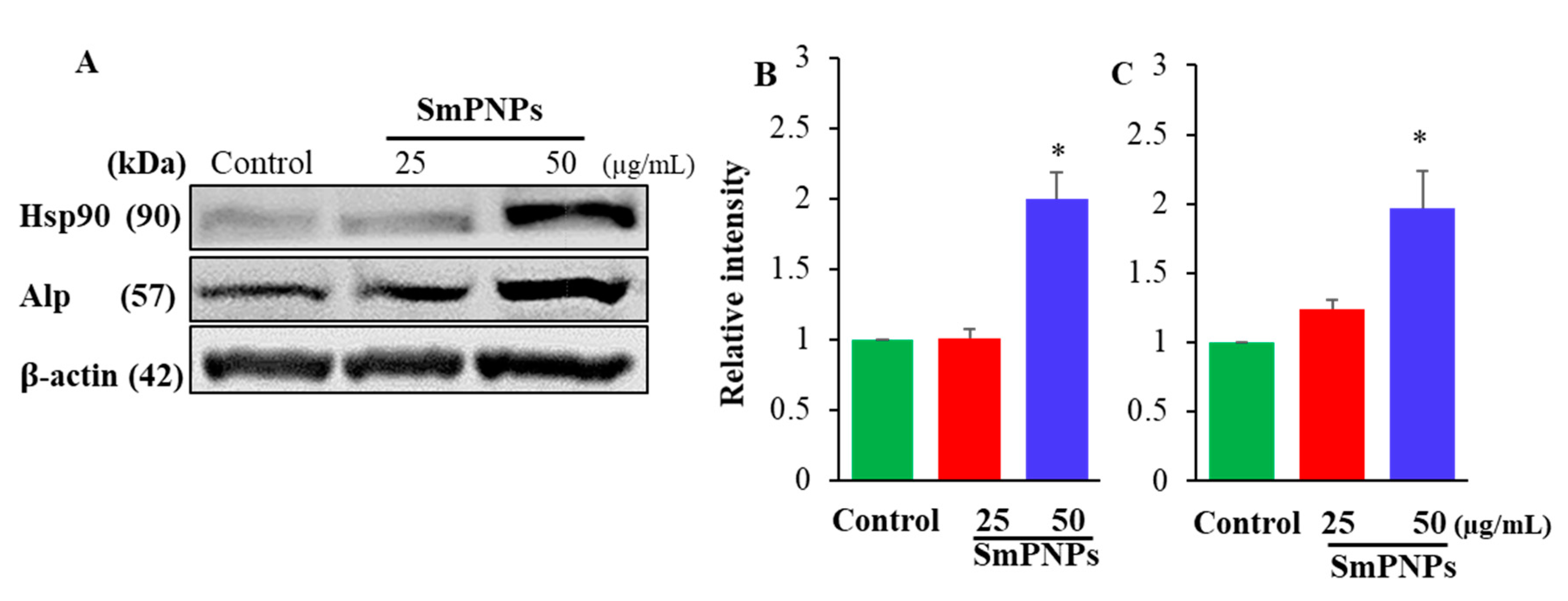
2.6. Effect of SmPNPs on the Expression of Alp and Hsp90 Proteins in Zebrafish Larave
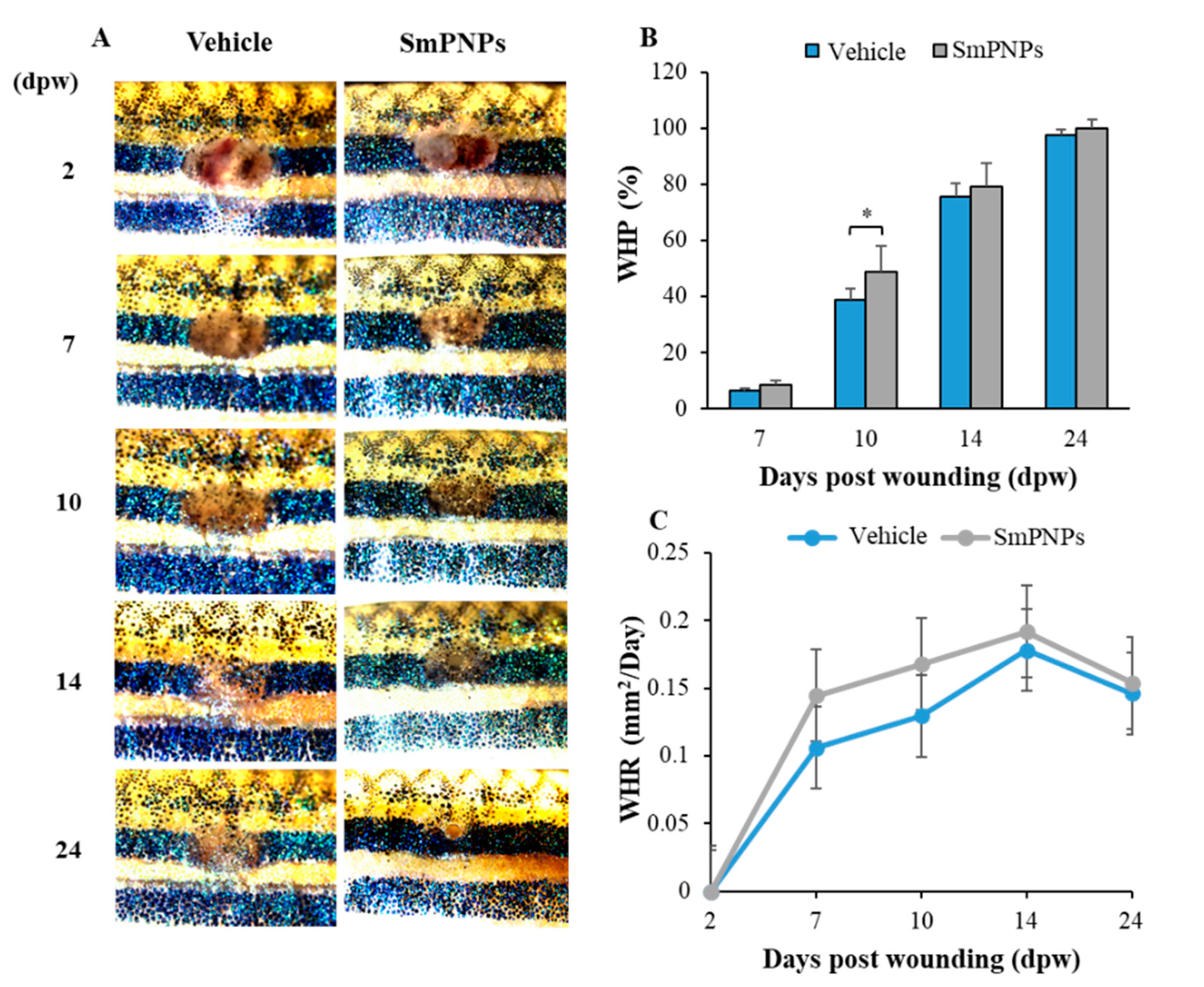
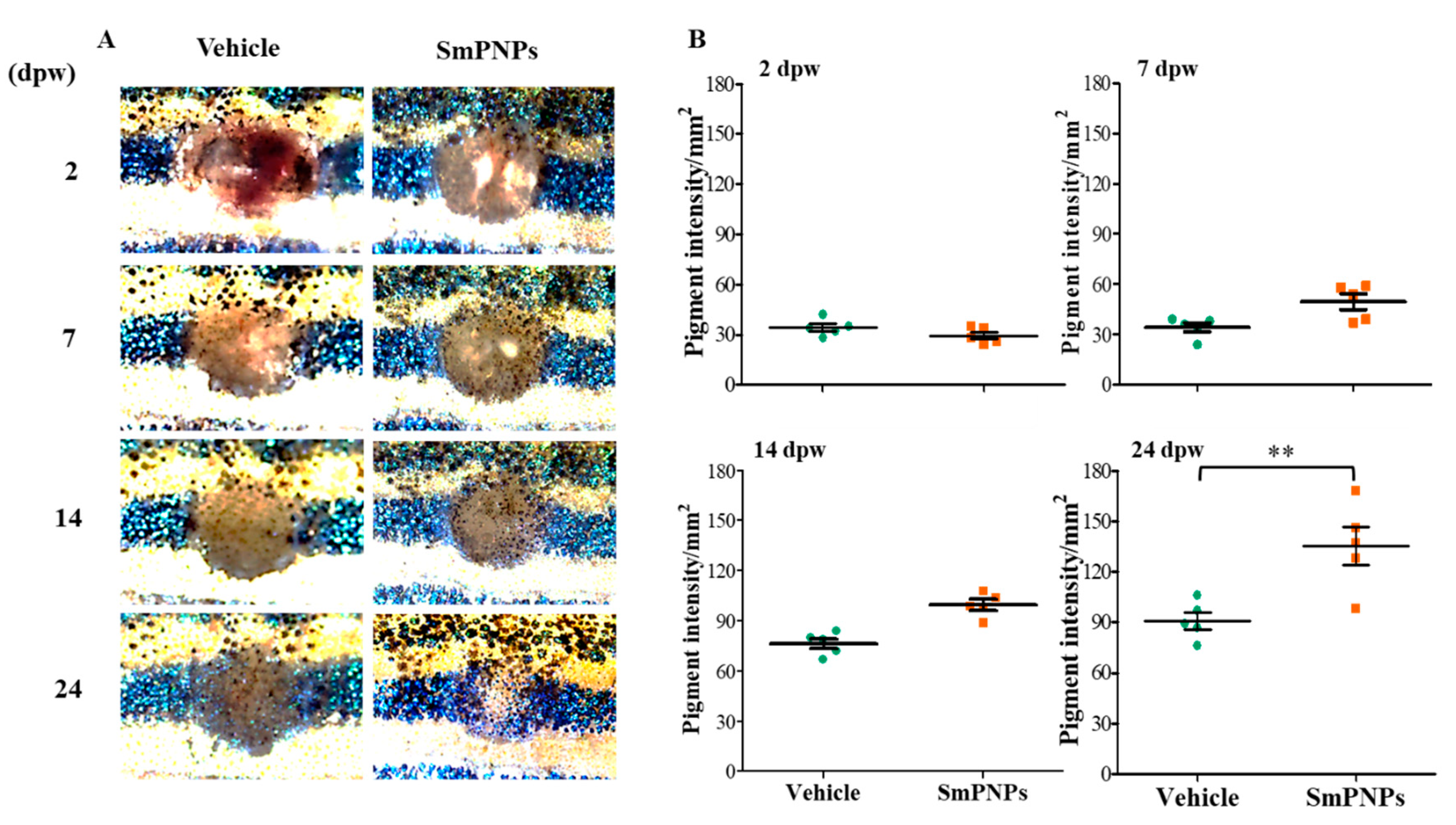
2.7. Effect of SmPNPs on Dermal Wound Healing and Pigment Restoration in Adult Zebrafish
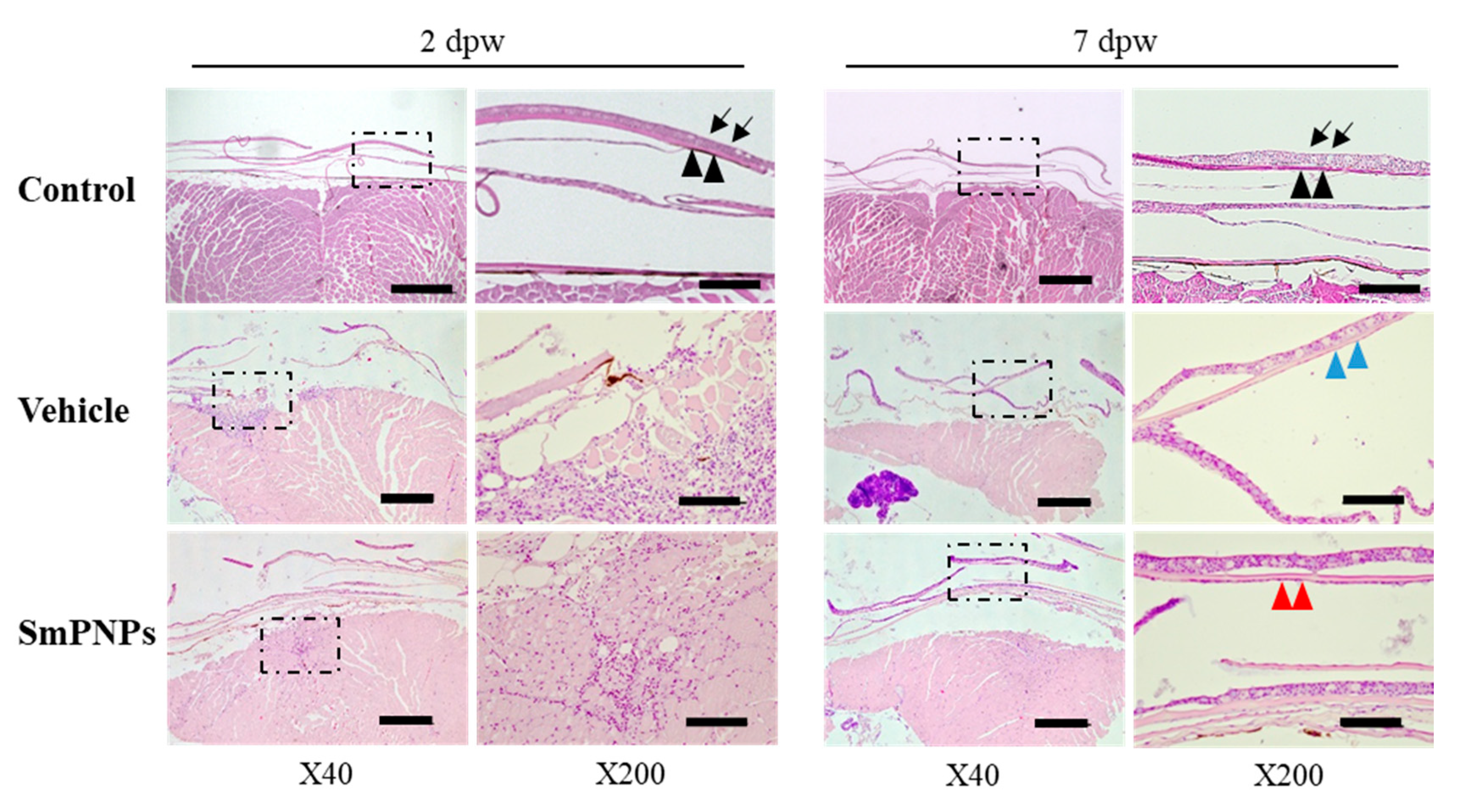
2.8. Histological Assessment of the Effect of SmPNPs on Wound Healing
2.9. Time-Course Transcriptional Analysis of Wound Healing in Adult Zebrafish upon SmPNPs Treatment
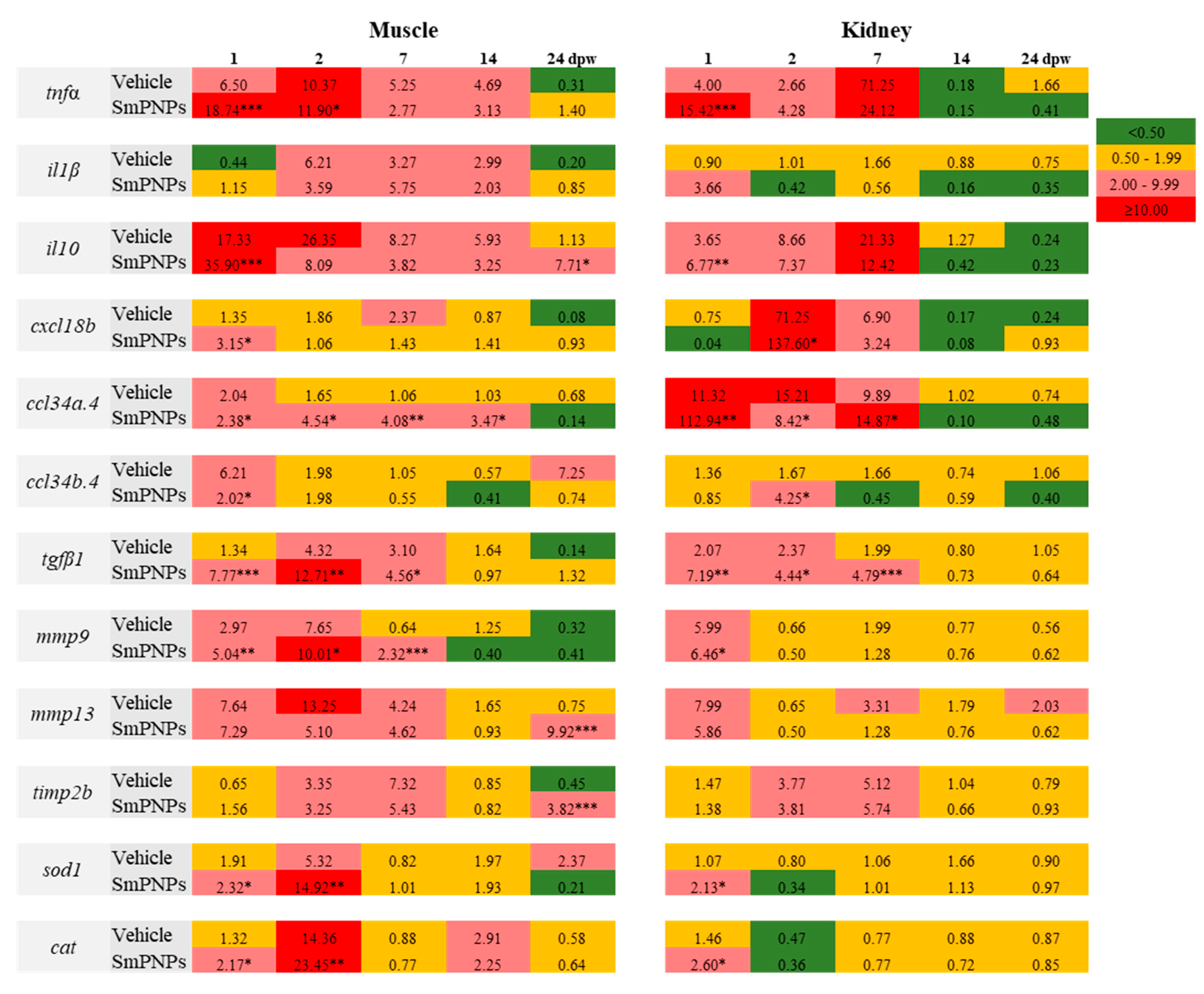
2.9.1. tnfα
2.9.2. il1β and il10
2.9.3. cxcl18b, ccl34a.4 and ccl34b.4
2.9.4. tgfβ1
2.9.5. mmp9 and mmp13
2.9.6. timp2b
2.9.7. sod1 and cat
3. Discussion
4. Materials and Methods
4.1. Preparation and Characterization of SmPNPs
4.2. Cell Culture and In Vitro Cytotoxicity of SmPNPs
4.3. Zebrafish Husbandry and Assessment of In Vivo Toxicity in Zebrafish Embryos
4.4. Determination of ROS Levels Induced by A. hydrophila Infection after Exposure to SmPNPs
4.5. SmPNPs Supplemented Diet Feeding Trial with Zebrafish
4.6. Immune Challenge of Larvae and Adult Zebrafish with A. hydrophila
4.7. Transcriptional Analysis of Immunomodulatory Genes upon SmPNPs Treatment
4.8. Immunoblot Analysis of Heat Shock Protein (Hsp90) and Alkaline Phosphatase (Alp)
4.9. Wounding and Topical Treatment of Adult Zebrafish with SmPNPs
4.10. Effect of SmPNPs on Wound Closure and Pigment Restoration
4.11. Histological Analysis of the Effect of SmPNPs on Wound Healing
4.12. Transcriptional Analysis of Wound Healing in Adult Zebrafish upon SmPNPs Treatment
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kang, H.K.; Lee, H.H.; Seo, C.H.; Park, Y. Antimicrobial and immunomodulatory properties and applications of marine-derived proteins and peptides. Mar. Drugs 2019, 17, 350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claverie, M.; McReynolds, C.; Petitpas, A.; Thomas, M.; Fernandes, S. Marine-derived polymeric materials and biomimetics: An Overview. Polymers 2020, 12, 1002. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z. The modification of natural products for medical use. Acta Pharm. Sin. B 2017, 7, 119–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minzanova, S.T.; Mironov, V.F.; Arkhipova, D.M.; Khabibullina, A.V.; Mironova, L.G.; Zakirova, Y.M.; Milyukov, V.A. Biological activity and pharmacological application of pectic polysaccharides: A review. Polymers 2018, 10, 1407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, R.; Chow, T.J. Hypolipidemic, antioxidant, and antiinflammatory activities of microalgae Spirulina. Cardiovasc. Ther. 2010, 28, 33–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michal, A.B.; Gershwin, M.E.; Amha, B. Spirulina in human nutrition and health. J. Appl. Phycol. 2009, 21, 747–748. [Google Scholar] [CrossRef]
- Karkos, P.D.; Leong, S.C.; Karkos, C.D.; Sivaji, N.; Assimakopoulos, D.A. Spirulina in clinical practice: Evidence-based human applications. Evid. Based Complement. Alternat. Med. 2011, 2011, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyoung, L.; Woon, Y.C.; Gun, H.P.; Younsik, J.; Areumi, P.; Yeonji, L.; Kang, D.H. Studies on the extraction of marine pectin from 14 Marine Algae, Its Content and Antioxidant Activity. Korean J. Food Nutr. 2020, 49, 677–685. [Google Scholar] [CrossRef]
- Popov, S.V.; Ovodov, Y.S. Polypotency of the immunomodulatory effect of pectins. Biochemistry 2013, 78, 823–835. [Google Scholar] [CrossRef]
- Heo, M.G.; Choung, S.Y. Anti-obesity effects of Spirulina maxima in high fat diet induced obese rats via the activation of AMPK pathway and SIRT1. Food Funct. 2018, 9, 4906–4915. [Google Scholar] [CrossRef]
- Brouns, F.; Theuwissen, E.; Adam, A.; Bell, M.; Berger, A.; Mensink, R.P. Cholesterol-lowering properties of different pectin types in mildly hyper-cholesterolemic men and women. Eur. J. Clin. Nutr. 2012, 66, 591–599. [Google Scholar] [CrossRef] [Green Version]
- Leclere, L.; Van Cutsem, P.; Michiels, C. Anti-cancer activities of pH-or heat-modified pectin. Front. Pharmacol. 2013, 4, 128. [Google Scholar] [CrossRef] [Green Version]
- Edirisinghe, S.L.; Dananjaya, S.H.S.; Nikapitiya, C.; Liyanage, T.D.; Lee, K.A.; Oh, C.; Do-Hyung, K.; De Zoysa, M. Novel pectin isolated from Spirulina maxima enhances the disease resistance and immune responses in zebrafish against Edwardsiella piscicida and Aeromonas hydrophila. Fish. Shellfish Immunol. 2019, 94, 558–565. [Google Scholar] [CrossRef]
- Edirisinghe, S.L.; Rajapaksha, D.C.; Nikapitiya, C.; Oh, C.; Lee, K.A.; Oh, C.; Do-Hyung, K.; De Zoysa, M. Spirulina maxima derived marine pectin promotes the in vitro and in vivo regeneration and wound healing on zebrafish. Fish. Shellfish Immunol. 2020. [Google Scholar] [CrossRef]
- Dvir, I.; Stark, A.H.; Chayoth, R.; Madar, Z.; Arad, S.M. Hypocholesterolemic effects of nutraceuticals produced from the red microalga Porphyridium sp. in rats. Nutrients 2009, 1, 156. [Google Scholar] [CrossRef] [Green Version]
- Patra, J.K.; Das, G.; Fraceto, L.F.; Campos, E.V.; Del Pilar Rodriguez-Torres, M.; Acosta-Torres, L.S.; Diaz-Torres, L.A.; Grillo, R.; Swamy, M.K.; Sharma, S.; et al. Nano based drug delivery systems: Recent developments and future prospects. J. Nanobiotechnol. 2018, 16, 71. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.; Saeed, K.; Khan, I. Nanoparticles: Properties, applications and toxicities. Arab. J. Chem. 2019, 12, 908–931. [Google Scholar] [CrossRef]
- Nikapitiya, C.; Dananjaya, S.H.S.; De Silva, B.C.J.; Heo, G.J.; Oh, C.; De Zoysa, M.; Lee, J. Chitosan nanoparticles: A positive immune response modulator as display in zebrafish larvae against Aeromonas hydrophila infection. Fish. Shellfish Immunol. 2018, 76, 240–246. [Google Scholar] [CrossRef]
- Burapapadh, K.; Takeuchi, H.; Sriamornsak, P. Development of pectin nanoparticles through mechanical homogenization for dissolution enhancement of itraconazole. Asian J. Pharm. Sci. 2016, 11, 365–375. [Google Scholar] [CrossRef] [Green Version]
- Chandrarathna, H.P.S.U.; Liyanage, T.D.; Edirisinghe, S.L.; Dananjaya, S.H.S.; Thulshan, E.H.T.; Nikapitiya, C.; Oh, C.; Do-Hyung, K.; De Zoysa, M. Marine microalgae, Spirulina maxima-derived modified pectin and modified pectin nanoparticles modulate the gut microbiota and trigger immune responses in Mice. Mar. Drugs 2020, 18, 175. [Google Scholar] [CrossRef] [Green Version]
- Fang, R.H.; Zhang, L. Nanoparticle-based modulation of the immune system. Annu. Rev. Chem. Biomol. Eng. 2016, 7, 305–326. [Google Scholar] [CrossRef]
- Jaison, J.; Barhoum, A.; Chan, Y.S.; Dufresne, A.; Danquah, M.K. Review on nanoparticles and nanostructured materials: History, sources, toxicity and regulations. Beilstein J. Nanotechnol. 2018, 9, 1050–1074. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Zhao, Y.; Nel, A.E.; Lin, S. Zebrafish: An in vivo model for nano EHS studies. Small 2013, 9, 1608–1618. [Google Scholar] [CrossRef] [Green Version]
- Drasler, B.; Sayre, P.; Steinhäuser, K.G.; Petri-Fink, A.; Rothen-Rutishauser, B. In vitro approaches to assess the hazard of nanomaterials. Nanoimpact 2017, 8, 99–116. [Google Scholar] [CrossRef]
- Formella, I.; Svahn, A.J.; Radford, R.A.; Don, E.K.; Cole, N.J.; Hogan, A.; Albert, L.; Roger, S.C.; Morsch, M. Real-time visualization of oxidative stress-mediated neurodegeneration of individual spinal motor neurons in vivo. Redox Boil. 2018, 19, 226–234. [Google Scholar] [CrossRef]
- Gilgun-Sherki, Y.; Melamed, E.; Offen, D. Oxidative stress induced-neurodegenerative diseases: The need for antioxidants that penetrate the blood brain barrier. Neuropharmacology 2001, 40, 959–975. [Google Scholar] [CrossRef]
- Fu, P.P.; Xia, Q.; Hwang, H.M.; Ray, P.C.; Yu, H. Mechanisms of nanotoxicity: Generation of reactive oxygen species. J. Food Drug Anal. 2014, 22, 64–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Oh, J.Y.; Kim, H.S.; Lee, W.; Cui, Y.; Lee, H.G.; Kim, Y.T.; Jeon, Y.J. Protective effect of polysaccharides from Celluclast-assisted extract of Hizikia fusiforme against hydrogen peroxide-induced oxidative stress in vitro in Vero cells and in vivo in zebrafish. Int. J. Biol. Macromol. 2018, 112, 483–489. [Google Scholar] [CrossRef] [PubMed]
- AlYahya, S.A.; Ameen, F.; Al-Niaeem, K.S.; Al-Sa’adi, B.A.; Hadi, S.; Mostafa, A.A. Histopathological studies of experimental Aeromonas hydrophila infection in blue tilapia, Oreochromis aureus. Saudi J. Biol. Sci. 2018, 25, 182–185. [Google Scholar] [CrossRef]
- Lall, S.P.; Dumas, A. Nutritional requirements of cultured fish: Formulating nutritionally adequate feeds. In Feed and Feeding Practices in Aquaculture; Woodhead Publishing: Cambridge, UK, 2015; pp. 53–109. [Google Scholar] [CrossRef]
- Nya, E.J.; Austin, B. Use of bacterial lipopolysaccharide (LPS) as an immunostimulant for the control of Aeromonas hydrophila infections in rainbow trout Oncorhynchus mykiss (Walbaum). J. Appl. Microbiol. 2010, 108, 686–694. [Google Scholar] [CrossRef]
- Anderson, D.P.; Siwicki, A.K. Duration of protection against Aeromonas salmonicida in brook trout immunostimulated with glucan or chitosan by injection or immersion. Progres. Fish Cult. 1994, 56, 258–261. [Google Scholar] [CrossRef]
- Dagvadorj, J.; Naiki, Y.; Tumurkhuu, G.; Noman, A.S.M.; Iftekar-E-Khuda, I.; Koide, N.; Komatsu, T.; Yoshida, T.; Yokochi, T. Interleukin (IL)-10 attenuates lipopolysaccharide-induced IL-6 production via inhibition of IκB-ζ activity by Bcl-3. Innate Immune. 2009, 15, 217–224. [Google Scholar] [CrossRef]
- Graeve, L.; Baumann, M.; Heinrich, P.C. Interleukin-6 in autoimmune diseases. Role of IL-6 in physiology and pathology of the immune defense. Clin. Investig. 1993, 71, 664–671. [Google Scholar] [CrossRef]
- Chen, H.W.; Yang, T.S.; Chen, M.J.; Chang, Y.C.; Eugene, I.; Wang, C.; Ho, C.L.; Lai, Y.J.; Yu, C.C.; Chou, J.C.; et al. Purification and immunomodulating activity of C-phycocyanin from Spirulina platensis cultured using power plant flue gas. Process Biochem. 2014, 49, 1337–1344. [Google Scholar] [CrossRef] [Green Version]
- Sokol, C.L.; Luster, A.D. The chemokine system in innate immunity. Cold Spring Harb. Perspect. Biol. 2015, 7, a016303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mühlhofer, M.; Berchtold, E.; Stratil, C.G.; Csaba, G.; Kunold, E.; Bach, N.C.; Sieber, A.A.; Haslbeck, M.; Zimmer, R.; Buchner, J. The heat shock response in yeast maintains protein homeostasis by chaperoning and replenishing proteins. Cell Rep. 2019, 29, 4593–4607. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Zhao, Y.; Xia, T.; Meng, H.; Ji, Z.; Liu, R.; George, S.; Xiong, S.; Wang, X.; Zhang, H.; et al. High content screening in zebrafish speeds up hazard ranking of transition metal oxide nanoparticles. ACS Nano 2011, 5, 7284–7295. [Google Scholar] [CrossRef]
- Mizumori, M.; Ham, M.; Guth, P.H.; Engel, E.; Kaunitz, J.D.; Akiba, Y. Intestinal alkaline phosphatase regulates protective surface microclimate pH in rat duodenum. J. Physiol. Paris 2009, 587, 3651–3663. [Google Scholar] [CrossRef]
- Singh, S.B.; Carroll-Portillo, A.; Coffman, C.; Ritz, N.L.; Lin, H.C. Intestinal alkaline phosphatase exerts anti-inflammatory effects against lipopolysaccharide by inducing autophagy. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Boateng, J.S.; Matthews, K.H.; Stevens, H.N.; Eccleston, G.M. Wound healing dressings and drug delivery systems: A review. J. Pharm. Sci. 2008, 97, 2892–2923. [Google Scholar] [CrossRef]
- Cañedo-Dorantes, L.; Cañedo-Ayala, M. Skin acute wound healing: A comprehensive review. Int. J. Inflamm. 2019, 3706315. [Google Scholar] [CrossRef]
- Tsala, D.E.; Amadou, D.; Habtemariam, S. Natural wound healing and bioactive natural products. Phytopharmacology 2013, 4, 532–560. [Google Scholar]
- Smith, A.M.; Moxon, S.; Morris, G.A. Biopolymers as wound healing materials. In Wound Healing Biomaterials; Woodhead Publishing: Cambridge, UK, 2016; pp. 261–287. [Google Scholar] [CrossRef]
- Munarin, F.A.B.I.O.L.A.; Tanzi, M.C.; Petrini, P.A.O.L.A. Advances in biomedical applications of pectin gels. Int. J. Biol. Macromol. 2012, 51, 681–689. [Google Scholar] [CrossRef]
- Seo, S.B.; Dananjaya, S.H.S.; Nikapitiya, C.; Park, B.K.; Gooneratne, R.; Kim, T.Y.; De Zoysa, M. Silver nanoparticles enhance wound healing in zebrafish (Danio rerio). Fish Shellfish Immun. 2017, 68, 536–545. [Google Scholar] [CrossRef]
- Chen, X.; Thibeault, S.L. Role of tumor necrosis factor–α in wound repair in human vocal fold fibroblasts. Laryngoscope 2010, 120, 1819–1825. [Google Scholar] [CrossRef] [Green Version]
- Roberts, A.B.; Sporn, M.B.; Assoian, R.K.; Smith, J.M.; Roche, N.S.; Wakefield, L.M.; Kehrl, J.H. Transforming growth factor type beta: Rapid induction of fibrosis and angiogenesis in vivo and stimulation of collagen formation in vitro. Proc. Natl. Acad. Sci. USA 1986, 83, 4167–4171. [Google Scholar] [CrossRef] [Green Version]
- Lichtman, M.K.; Otero-Vinas, M.; Falanga, V. Transforming growth factor beta (TGF-β) isoforms in wound healing and fibrosis. Wound Repair Regen. 2016, 24, 215–222. [Google Scholar] [CrossRef]
- An, L.; Dong, G.Q.; Gao, Q.; Zhang, Y.; Hu, L.W.; Li, J.H.; Liu, Y. Effects of UVA on TNF-α, IL-1β, and IL-10 expression levels in human keratinocytes and intervention studies with an antioxidant and a JNK inhibitor. Photodermatol. Photoimmunol. Photomed. 2010, 26, 28–35. [Google Scholar] [CrossRef]
- Hattori, N.; Mochizuki, S.; Kishi, K.; Nakajima, T.; Takaishi, H.; D’Armiento, J.; Okada, Y. MMP-13 plays a role in keratinocyte migration, angiogenesis, and contraction in mouse skin wound healing. Am. J. Pathol. 2009, 175, 533–546. [Google Scholar] [CrossRef] [Green Version]
- Beom, S.S.; Shin, S.Y.; Dananjaya, S.H.S.; De Silva, A.B.K.H.; Nikapitiya, C.; Cho, J.; De Zoysa, M. Chitosan based silver nanocomposites (CAgNCs) display antibacterial effects against Vibrio Ichthyoenteri. Scopus 2017, 34, 261–267. [Google Scholar] [CrossRef]
- Dananjaya, S.H.S.; Kulatunga, D.C.M.; Godahewa, G.I.; Nikapitiya, C.; Edussuriya, M.; De Zoysa, M. Preparation, characterization, and antimicrobial properties of chitosan–silver nanocomposites films against fish pathogenic bacteria and fungi. Indian J. Microbiol. 2017, 57, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Lakey, P.S.; Berkemeier, T.; Tong, H.; Arangio, A.M.; Lucas, K.; Pöschl, U.; Shiraiwa, M. Chemical exposure-response relationship between air pollutants and reactive oxygen species in the human respiratory tract. Sci. Rep. 2016, 6, 32916. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2—ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Liyanage, T.D.; Dahanayake, P.S.; Edirisinghe, S.L.; Nikapitiya, C.; Heo, G.J.; De Zoysa, M.; Whang, I. Biological activity of porcine gastric mucin on stress resistance and immunomodulation. Molecules 2020, 25, 2981. [Google Scholar] [CrossRef]
- Sveen, L.R.; Timmerhaus, G.; Krasnov, A.; Takle, H.; Handeland, S.; Ytteborg, E. Wound healing in post-smolt Atlantic salmon (Salmo salar L.). Sci. Rep. 2019, 9, 1–16. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajapaksha, D.C.; Edirisinghe, S.L.; Nikapitiya, C.; Dananjaya, S.; Kwun, H.-J.; Kim, C.-H.; Oh, C.; Kang, D.-H.; De Zoysa, M. Spirulina maxima Derived Pectin Nanoparticles Enhance the Immunomodulation, Stress Tolerance, and Wound Healing in Zebrafish. Mar. Drugs 2020, 18, 556. https://doi.org/10.3390/md18110556
Rajapaksha DC, Edirisinghe SL, Nikapitiya C, Dananjaya S, Kwun H-J, Kim C-H, Oh C, Kang D-H, De Zoysa M. Spirulina maxima Derived Pectin Nanoparticles Enhance the Immunomodulation, Stress Tolerance, and Wound Healing in Zebrafish. Marine Drugs. 2020; 18(11):556. https://doi.org/10.3390/md18110556
Chicago/Turabian StyleRajapaksha, Dinusha C., Shan L. Edirisinghe, Chamilani Nikapitiya, SHS Dananjaya, Hyo-Jung Kwun, Cheol-Hee Kim, Chulhong Oh, Do-Hyung Kang, and Mahanama De Zoysa. 2020. "Spirulina maxima Derived Pectin Nanoparticles Enhance the Immunomodulation, Stress Tolerance, and Wound Healing in Zebrafish" Marine Drugs 18, no. 11: 556. https://doi.org/10.3390/md18110556