Antioxidant and Cytotoxic Effects on Tumor Cells of Exopolysaccharides from Tetraselmis suecica (Kylin) Butcher Grown Under Autotrophic and Heterotrophic Conditions

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Adaptation of Autotrophic to Heterotrophic Culture of T. suecica
2.2. Elemental Analysis of Autotrophic and Heterotrophic Biomass Cultures of T. suecica
2.3. Biochemical Composition of Autotrophic and Heterotrophic Biomass Cultures of T. suecica
2.4. Phenolic Compounds and Antioxidant Activity of Autotrophic and Heterotrophic Biomass Cultures of T. suecica
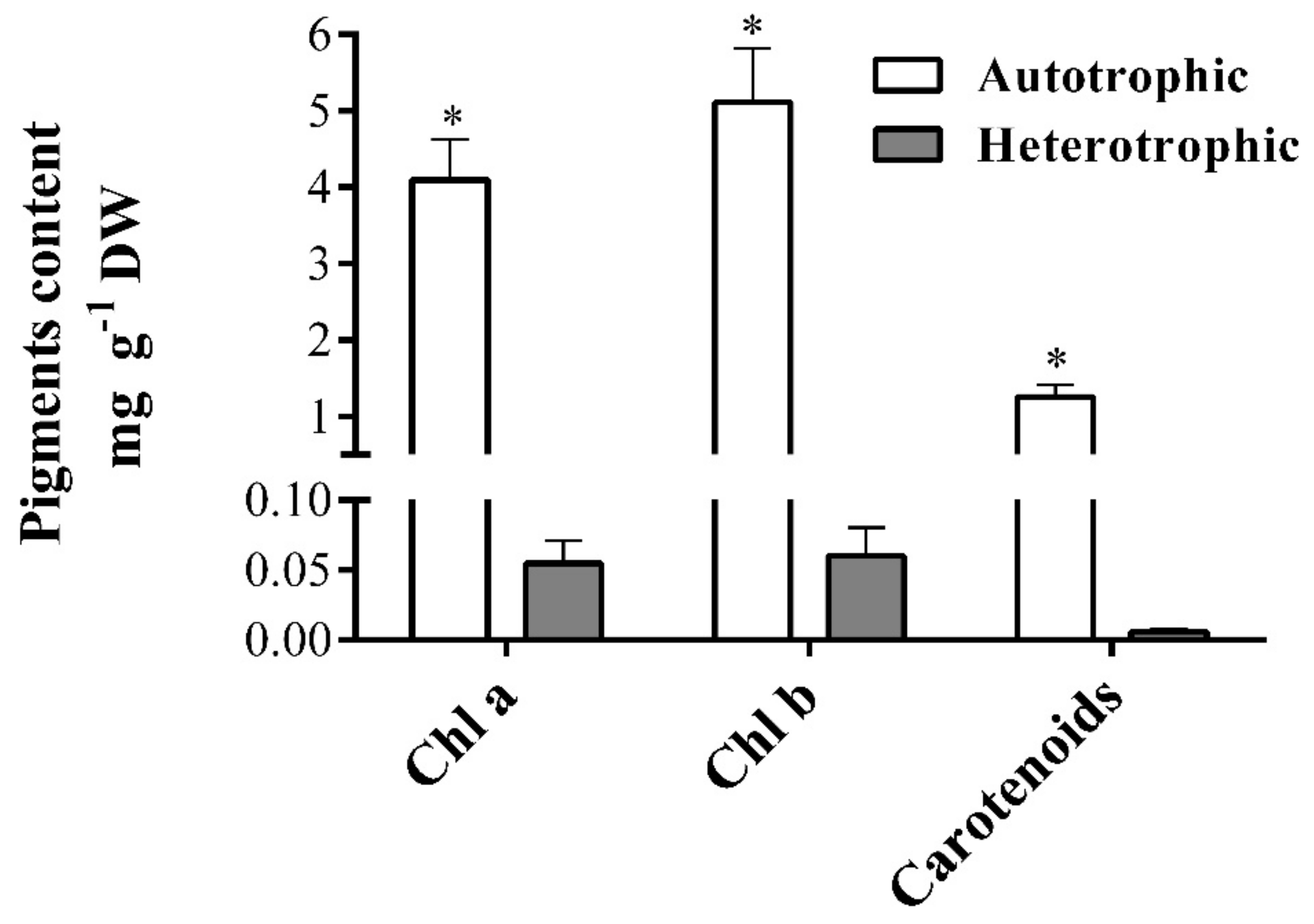
2.5. Pigment Content of Autotrophic and Heterotrophic Biomass Cultures of T. suecica
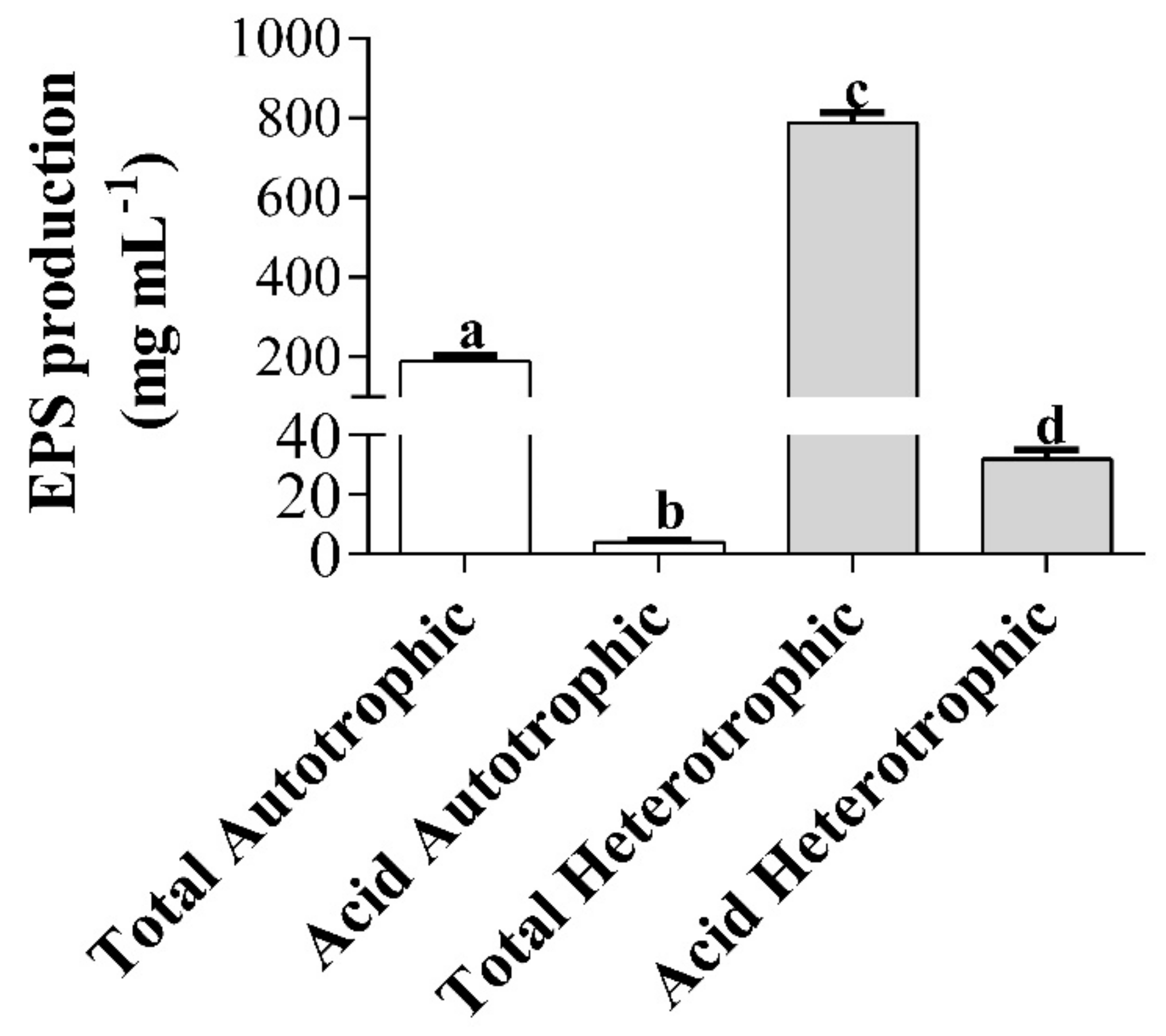
2.6. Production and Extraction of Exopolysaccharides (EPS) of Autotrophic and Heterotrophic Biomass Cultures of T. suecica
2.7. Elemental Analysis of Exopolysaccharides (EPS) of Autotrophic and Heterotrophic Biomass Cultures of T. suecica
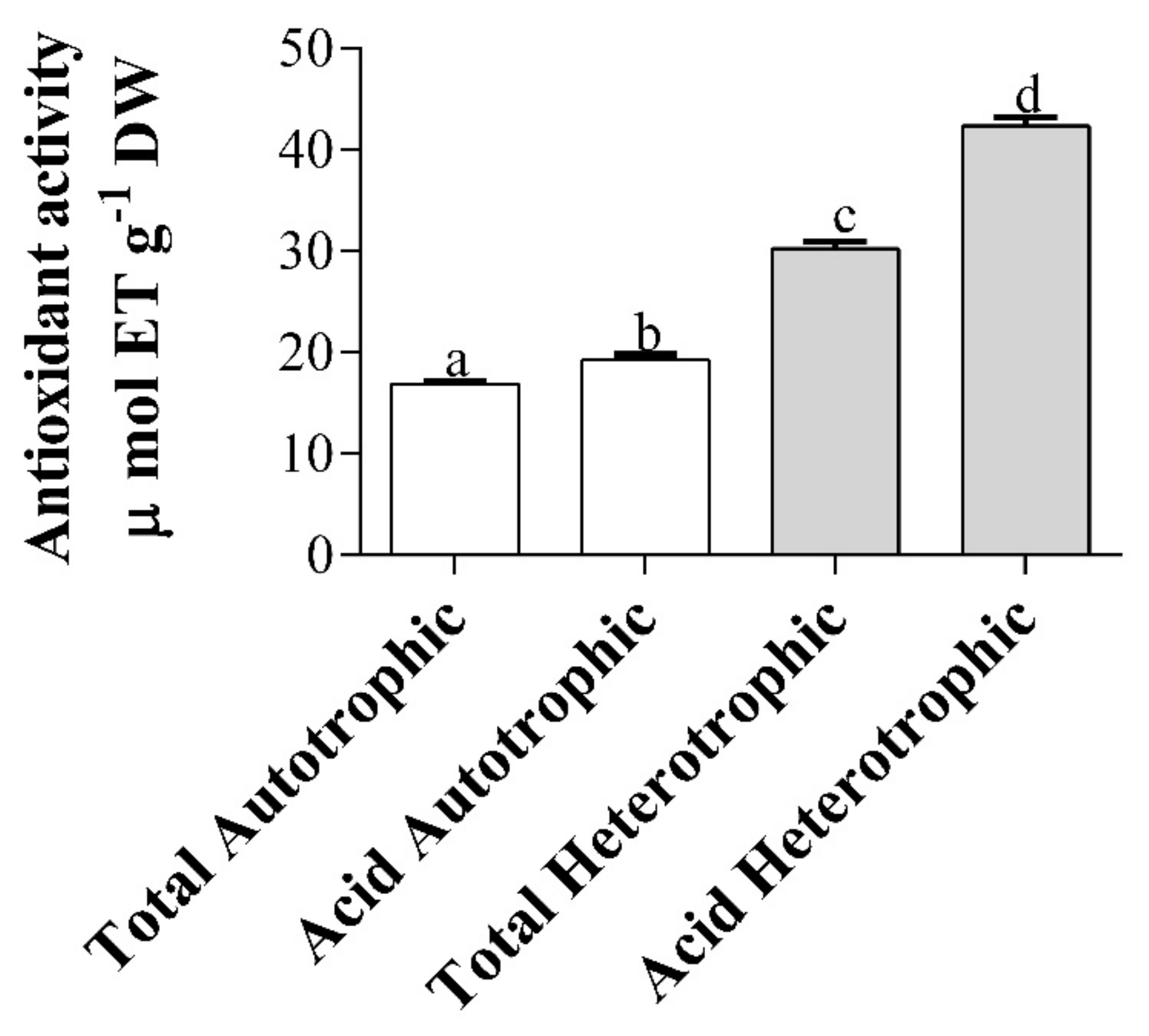
2.8. Antioxidant Activity of Exopolysaccharides (EPS) of Autotrophic and Heterotrophic Biomass Cultures of T. suecica
2.9. Fourier-Transform Infrared Spectroscopy (FTIR) of Exopolysaccharides (EPS) of Autotrophic and Heterotrophic Biomass Cultures of T. suecica
2.10. Gas Chromatography—Mass Spectrometry (GC-MS) of Exopolysaccharides (EPS) of Autotrophic and Heterotrophic Biomass Cultures of T. suecica
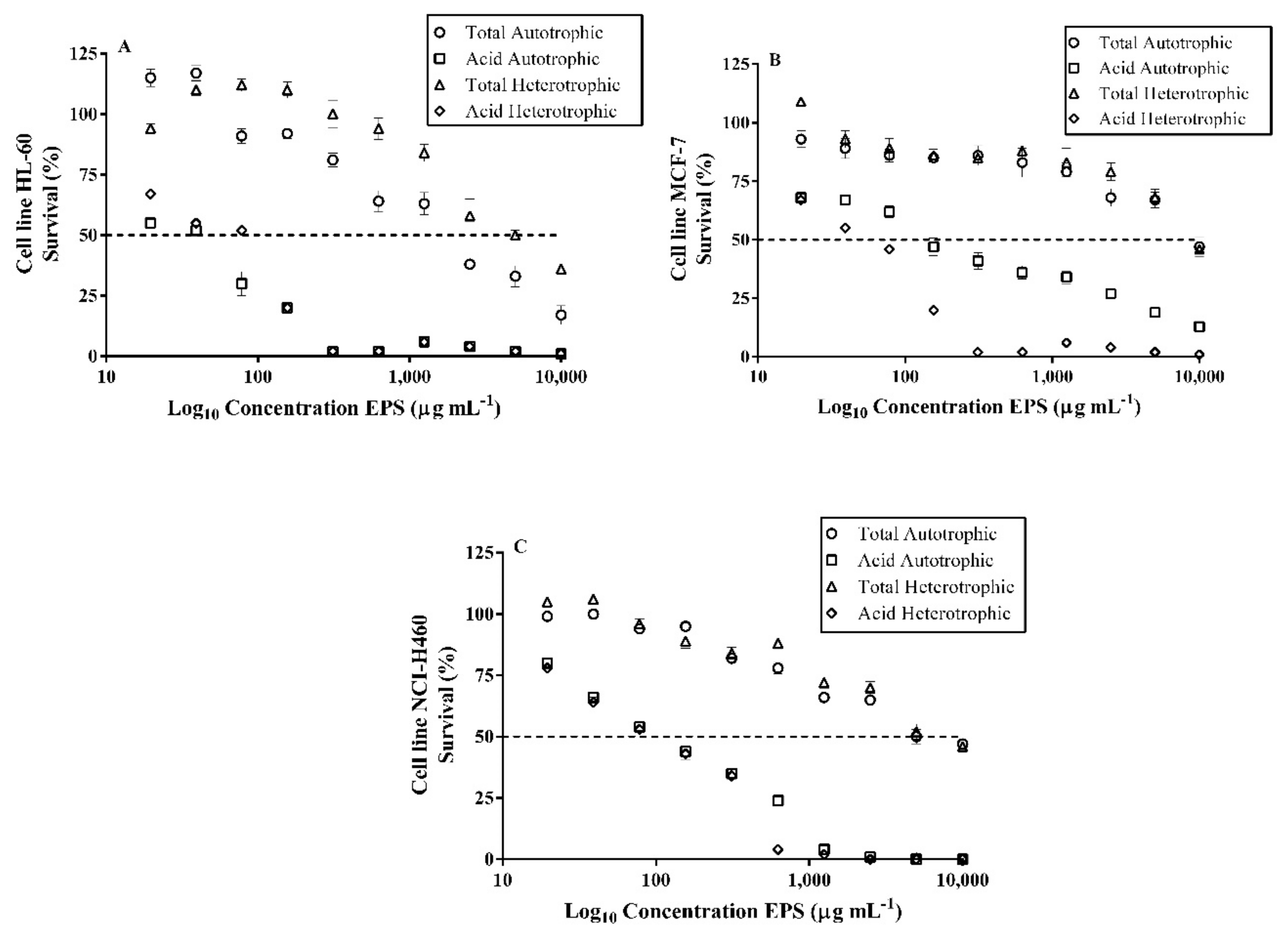
2.11. Cytotoxic Effects on Tumor Cells of Exopolysaccharides (EPS) of Autotrophic and Heterotrophic Biomass Cultures of T. suecica
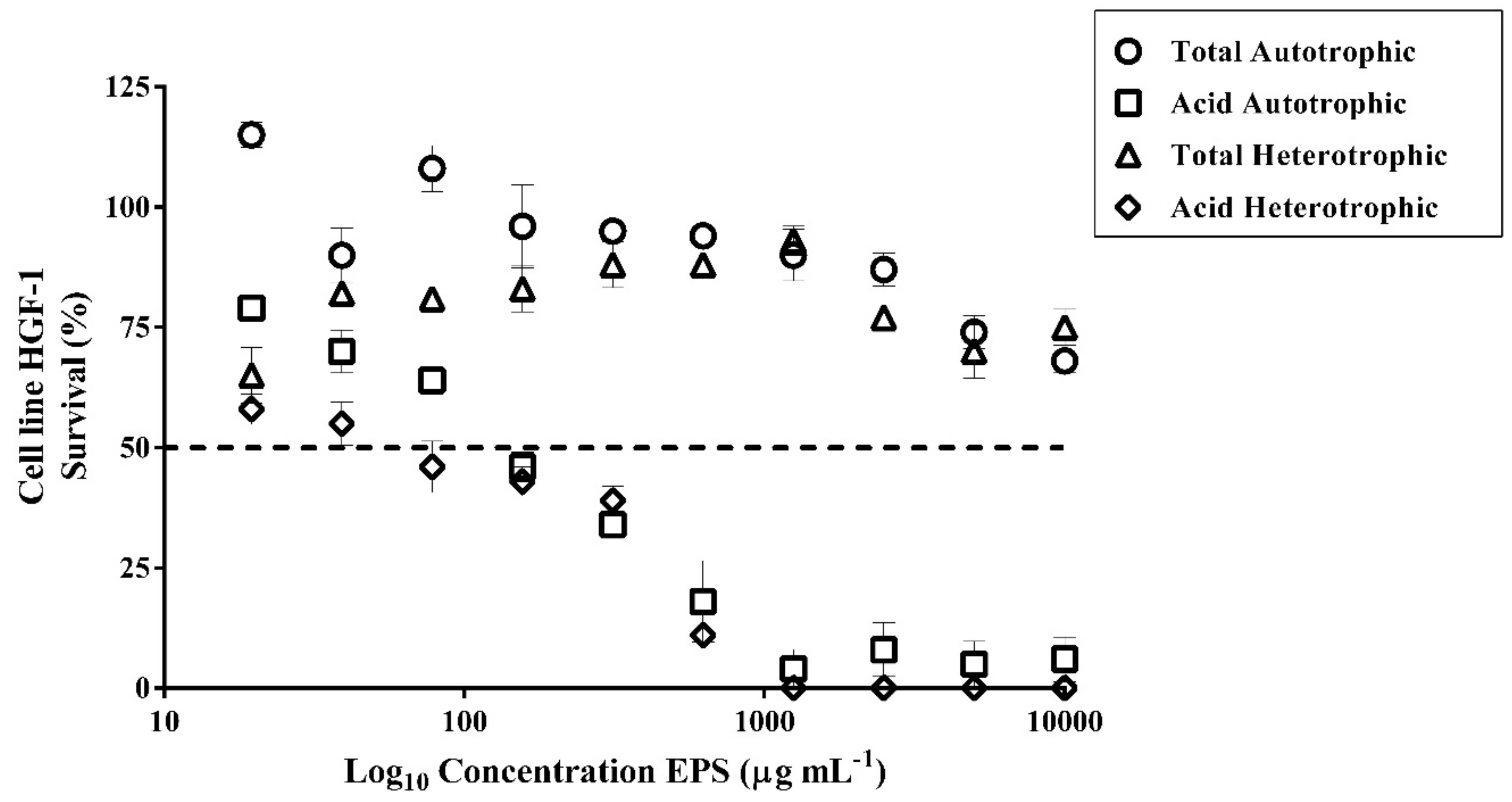
2.12. Cytotoxic of Exopolysaccharides (EPS) of Autotrophic and Heterotrophic Biomass Cultures of T. suecica
3. Materials and Methods
3.1. Biological Material
3.2. Culture Conditions
3.2.1. Autotrophic Culture of T. suecica
3.2.2. Heterotrophic Culture of T. suecica
3.3. Extraction Conditions
3.3.1. Biomass Extraction
3.3.2. Exopolysaccharides Extraction
3.4. Population Parameters (Cell Density, Cell Concentration, Specific Growth Rate, Biovolume and Cell Volume)
3.5. Total Carbon (C), Hydrogen (H), Nitrogen (N) and Sulfur (S) of Dry Biomass and Exopolysaccharides
3.6. Biochemical Composition of Autotrophic and Heterotrophic Biomass Cultures of T. suecica
3.7. Determination of Phenolic Compounds
3.8. Determination of Pigments
3.9. Lipopolysaccharides (LPS) Contamination Assay
3.10. Antioxidant Capacity
3.10.1. ABTS Assay Scavenging of Free Radical in Exopolysaccharides and Biomass
3.10.2. DPPH Free-Radical Method in Biomass from T. suecica
3.11. Fourier-Transform Infrared Spectroscopy (FTIR)
3.12. Gas chromatography–Mass Spectrometry (GC-MS)
3.12.1. Hydrolysis and Derivatization of EPS
3.12.2. Gas Chromatography/Mass Spectrometry (GC-MS) Analysis
3.13. Cell Line Cultures
3.14. Cytotoxic Effects on Tumor Cells Assay
3.15. Cytotoxicity Assay in Healthy Cell Line
3.16. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brennan, L.; Owende, P. Biofuels from microalgae—A review of technologies for production, processing, and extractions of biofuels and co-products. Renew. Sustain. Energy Rev. 2010, 14, 557–577. [Google Scholar] [CrossRef]
- Vásquez-Piñeros, M.A.; Rondón-Barragan, I.S.; Eslava-Mocha, P.R.; Marina, B. Inmunoestimulantes en teleosteos: Probióticos, β-glucanos y LPS. Orinoquia 2012, 16, 46–62. [Google Scholar] [CrossRef]
- Huang, G.; Chen, F.; Wei, D.; Zhang, X.; Chen, G. Biodiesel production by microalgal biotechnology. Appl. Energy 2010, 87, 38–46. [Google Scholar] [CrossRef]
- Acién, F.; Gómez-Serrano, C.; Morales-Amaral, M.; Fernández-Sevilla, J.; Molina-Grima, E. Wastewater treatment using microalgae: How realistic a contribution might it be to significant urban wastewater treatment? Appl. Microbiol. Biotechnol. 2016, 100, 9013–9022. [Google Scholar] [CrossRef] [PubMed]
- Pérez-García, O.; Escalante, F.M.E.; De-Bashan, L.E.; Bashan, Y. Heterotrophic cultures of microalgae: Metabolism and potential products. Water Res. 2011, 45, 11–36. [Google Scholar] [CrossRef] [PubMed]
- Morales-Sánchez, D.; Martinez-Rodriguez, O.A.; Kyndt, J.; Martinez, A. Heterotrophic growth of microalgae: Metabolic aspects. World J. Microbiol. Biotechnol. 2015, 31, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Morales-Sánchez, D.; Martinez-Rodriguez, O.A.; Martinez, A. Heterotrophic cultivation of microalgae: Production of metabolites of commercial interest. J. Chem. Technol. Biotechnol. 2017, 92, 925–936. [Google Scholar] [CrossRef]
- Borowitzka, M.A. Algal Physiology and Large-Scale Outdoor Cultures of Microalgae. In The Physiology of Microalgae: Developments in Applied Phycology; Borowitzka, M., Beardall, M., Rave, J., Eds.; Springer: Cham, Switzerland, 2016; Volume 6, pp. 601–652. [Google Scholar]
- Velea, S.; Oancea, F.; Fischer, F. Heterotrophic and mixotrophic microalgae cultivation. In Microalgae-Based Biofuels and Bioproducts; Gonzalez-Fernandez, C., Muñoz, R., Eds.; Woodhead Publishing Elsevier: Kindlington, UK, 2017; pp. 45–65. [Google Scholar]
- Barros, A.; Pereira, H.; Campos, J.; Marques, A.; Varela, J.; Silva, J. Heterotrophy as a tool to overcome the long and costly autotrophic scale-up process for large scale production of microalgae. Sci. Rep. 2019, 9, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Miao, X.; Wu, Q. High quality biodiesel production from a microalga Chlorella protothecoides by heterotrophic growth in fermenters. J. Biotechnol. 2006, 126, 499–507. [Google Scholar] [CrossRef]
- Azma, M.; Shamzi Mohamed, M.; Mohamad, R.; Abdul Rahim, R.; Ariff, A.B. Improvement of medium composition for heterotrophic cultivation of green microalgae, Tetraselmis suecica, using response surface methodology. Biochem. Eng. J. 2011, 53, 187–195. [Google Scholar] [CrossRef]
- Leyva, L.A.; Bashan, Y.; Mendoza, A.; de-Bashan, L.E. Accumulation fatty acids of in Chlorella vulgaris under heterotrophic conditions in relation to activity of acetyl-CoA carboxylase, temperature, and co-immobilization with Azospirillum brasilense. Naturwissenschaften 2014, 101, 819–830. [Google Scholar] [CrossRef] [PubMed]
- Gladue, R.M.; Maxey, J.E. Microalgal feeds for aquaculture. J. Appl. Phycol. 1994, 6, 131–141. [Google Scholar] [CrossRef]
- Choix, F.J.; De-Bashan, L.E.; Bashan, Y. Enhanced accumulation of starch and total carbohydrates in alginate-immobilized Chlorella spp. induced by Azospirillum brasilense: II. Heterotrophic conditions. Enzym. Microb. Technol. 2012, 51, 300–309. [Google Scholar] [CrossRef]
- El-Sheekh, M.M.; Bedaiwy, M.Y.; Osman, M.E.; Ismail, M.M. Mixotrophic and heterotrophic growth of some microalgae using extract of fungal-treated wheat bran. Int. J. Recycl. Org. Waste Agric. 2012, 1, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Sloth, J.K.; Wiebe, M.G.; Eriksen, N.T. Accumulation of phycocyanin in heterotrophic and mixotrophic cultures of the acidophilic red alga Galdieria sulphuraria. Enzym. Microb. Technol. 2006, 38, 168–175. [Google Scholar] [CrossRef]
- Graverholt, O.S.; Eriksen, N.T. Heterotrophic high-cell-density fed-batch and continuous-flow cultures of Galdieria sulphuraria and production of phycocyanin. Biotechnol. Bioprocess. Eng. 2007, 77, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.Y.; Shi, C.L.; Shi, X.M. Modeling of lutein production by heterotrophic Chlorella in batch and fed-batch cultures. World J. Microbiol. Biotechnol. 2007, 23, 1233–1238. [Google Scholar] [CrossRef]
- Day, J.; Edwards, A.; Rodgers, G. Development of an industrial-scale process for the heterotrophic production of a microalgal mollusc feed. Bioresour. Technol. 1991, 38, 245–249. [Google Scholar] [CrossRef]
- Azma, M.; Mohamad, R.; Rahim, R.A.; Ariff, A.B. Improved Protocol for the Preparation of Tetraselmis suecica Axenic Culture and Adaptation to Heterotrophic Cultivation. Open Biotechnol. J. 2010, 4, 36–46. [Google Scholar] [CrossRef]
- Austin, B.; Day, J.G. Inhibition of prawn pathogenic Vibrio spp. by a commercial spray-dried preparation of Tetraselmis suecica. Aquaculture 1990, 90, 389–392. [Google Scholar] [CrossRef]
- Austin, B.; Baudet, E.; Stobie, M. Inhibition of bacterial fish pathogens by Tetraselmis suecica. J. Fish. Dis. 1992, 15, 55–61. [Google Scholar] [CrossRef]
- Irianto, A.; Austin, B. Probiotics in aquaculture. J. Fish. Dis. 2002, 25, 633–642. [Google Scholar] [CrossRef]
- Carballo-Cárdenas, E.C.; Tuan, M.; Janssen, M.; Wijffels, R.H. Vitamin E (a-tocopherol) production by the marine microalgae Dunaliella tertiolecta and Tetraselmis suecica in batch cultivation. Biomol. Eng. 2003, 20, 139–147. [Google Scholar] [CrossRef]
- Guzman-Murillo, M.; Ascencio, F. Anti-adhesive activity of sulphated exopolysaccharides of microalgae on attachment of red sore disease-associated bacteria and Helicobacter pylori to tissue culture cells. Lett. Appl. Microbiol. 2000, 30, 473–478. [Google Scholar] [CrossRef]
- Pontis, H.G. Case Study: Polysaccharides. In Methods for Analysis of Carbohydrate Metabolism in Photosynthetic Organisms: Plants, Green Algae and Cyanobacteria; Pontis, H., Ed.; Elsevier Academic Press: Cambridge, MA, USA, 2016; pp. 137–149. [Google Scholar]
- De Jesus Raposo, M.F.; de Morais, A.M.; de Morais, R.M. Bioactivity and Applications of Polysaccharides from Marine Microalgae. In Polysaccharides; Ramawat, K., Mérillon, J., Eds.; Springer: Cham, Switzerland, 2015; pp. 1683–1727. [Google Scholar]
- Rupérez, P.; Ahrazem, O.; Leal, J.A. Potential antioxidant capacity of sulfated polysaccharides from the edible marine brown seaweed Fucus vesiculosus. J. Agric. Food Chem. 2002, 50, 840–845. [Google Scholar] [CrossRef]
- Mendiola, J.A.; Jaime, L.; Santoyo, S.; Reglero, G.; Cifuentes, A.; Ibañez, E.; Señoráns, F.J. Screening of functional compounds in supercritical fluid extracts from Spirulina platensis. Food Chem. 2007, 102, 1357–1367. [Google Scholar] [CrossRef]
- Abdala Díaz, R.T.; Casas Arrojo, V.; Arrojo Agudo, M.A.; Cárdenas, C.; Dobretsov, S.; Figueroa, F.L. Immunomodulatory and Antioxidant Activities of Sulfated Polysaccharides from Laminaria ochroleuca, Porphyra umbilicalis, and Gelidium corneum. Mar. Biotechnol. 2019, 21, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Gardeva, E.; Toshkova, R.; Minkova, K.; Gigova, L. Cancer Protective Action of Polysaccharide, Derived from Red Microalga Porphyridium cruentum-A Biological Background. Biotechnol. Biotechnol Equip. 2009, 23, 783–787. [Google Scholar] [CrossRef]
- Day, J.; Tsavalos, A. An investigation of the heterotrophic culture of the green alga Tetraselmis. J. Appl. Phycol. 1996, 8, 73–77. [Google Scholar] [CrossRef]
- Walne, P. Studies on food value of nineteen genera of algae to juvenile bivalvies of the genera Ostrea. Fish. Invest. Lond. Ser. 1970, 1–62. [Google Scholar]
- Razaghi, A.; Godhe, A.; Albers, E. Effects of nitrogen on growth and carbohydrate formation in Porphyridium cruentum. Cent. Eur. J. Biol 2014, 9, 156–162. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhou, W.; Gao, C.; Lan, K.; Gao, Y.; Wu, Q. Biodiesel production from Jerusalem artichoke (Helianthus tuberosus L.) tuber by heterotrophic microalgae Chlorella protothecoides. J. Chem. Technol. Biotechnol. 2009, 84, 777–781. [Google Scholar] [CrossRef]
- Cid, A.; Abalde, J.; Herrero, C. Crecimiento y composición bioquímica de la microalga marina Tetraselmis suecica en cultivos mixotróficos con distintos azúcares y aminoácidos. Cah. Biol. Mar. 1992, 33, 169–178. [Google Scholar]
- Canelli, G.; Neutsch, L.; Carpine, R.; Tevere, S.; Giuffrida, F.; Rohfritsch, Z.; Dionisi, F.; Bolten, C.; Mathys, A. Chlorella vulgaris in a heterotrophic bioprocess: Study of the lipid bioaccessibility and oxidative stability. Algal Res. 2020, 45, 101754. [Google Scholar] [CrossRef]
- Morales-Sánchez, D.; Tinoco-Valencia, R.; Kyndt, J.; Martinez, A. Heterotrophic growth of Neochloris oleoabundans using glucose as a carbon source. Biotechnol. Biofuels 2013, 6, 100. [Google Scholar] [CrossRef] [PubMed]
- Leman, J. Oleaginous microorganisms: An assessment of the potential. In Advances in Applied Microbiology; Neidleman, S., Laskin, A., Eds.; Elsevier Academic Press: Cambridge, MA, USA, 1997; Volume 43, pp. 195–243. [Google Scholar]
- Garcia-Ferris, C.; Rios, A.; Ascaso, C.; Moreno, J. Correlated biochemical and ultrastructural changes in nitrogen-starved Euglena gracilis. J. Phycol. 1996, 32, 953–963. [Google Scholar] [CrossRef]
- Lillo, C.; Lea, U.; Ruoff, P. Nutrient depletion as a key factor for manipulating gene expression and product formation in different branches of the flavonoid pathway. Plant Cell Environ. 2008, 31, 587–601. [Google Scholar] [CrossRef]
- Quiñones-Galvez, J.; Hernández de la Torre, M.; Quirós Molina, Y.; Capdesuñer Ruiz, Y.; Trujillo Sánchez, R. Factors controlling phenol content on Theobroma cacao callus culture. Cultiv. Trop. 2016, 37, 118–126. [Google Scholar]
- Shetty, V.; Sibi, G. Relationship Between Total Phenolics Content and Antioxidant Activities of Microalgae Under Autotrophic, Heterotrophic and Mixotrophic Growth. J. Food Resourc. Sci. 2015, 4, 1–9. [Google Scholar]
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef]
- Lippemeier, S.; Klaus, R.H.; Vanselow, K.H.; Hartig, P.; Colijn, F. In-line recording of PAM fluorescence of phytoplankton cultures as a new tool for studying effects of fluctuating nutrient supply on photosynthesis. Eur. J. Phycol. 2001, 36, 89–100. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive Oxygen Species: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goiris, K.; Van Colen, W.; Wilches, I.; León-Tamariz, F.; De Cooman, L.; Muylaert, K. Impact of nutrient stress on antioxidant production in three species of microalgae. Algal Res. 2015, 7, 51–57. [Google Scholar] [CrossRef]
- Alam, M.N.; Bristi, N.J.; Rafiquzzaman, M. Review on in vivo and in vitro methods evaluation of antioxidant activity. Saudi Pharm. J. 2013, 21, 143–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.; Kumari, P.; Trivedi, N.; Shukla, M.K.; Gupta, V.; Reddy, C.R.K.; Jha, B. Minerals, PUFAs and antioxidant properties of some tropical seaweeds from Saurashtra coast of India. J. App.l Phycol. 2011, 23, 797–810. [Google Scholar] [CrossRef]
- Surveswaran, S.; Cai, Y.Z.; Corke, H.; Sun, M. Systematic evaluation of natural phenolic antioxidants from 133 Indian medicinal plants. Food Chem. 2007, 102, 938–953. [Google Scholar] [CrossRef]
- Jiménez-Escrig, A.; Jiménez-Jiménez, I.; Pulido, R.; Saura-Calixto, F. Antioxidant activity of fresh and processed edible seaweeds. J. Sci. Food Agric. 2001, 81, 530–534. [Google Scholar] [CrossRef]
- Kováčik, J.; Klejdus, B.; Bačkor, M. Physiological Responses of Scenedesmus quadricauda (Chlorophyceae) to UV-A and UV-C Light. Photochem. Photobiol. 2010, 86, 612–616. [Google Scholar] [CrossRef]
- Young, A.J. Factors that affect the carotenoid composition of higher plants and algae. In Carotenoids in Photosynthesis, 1st ed.; Young, A.J., Britton, G., Eds.; Springer: Dordrecht, The Netherlands, 1993; pp. 160–205. [Google Scholar]
- Lapointe, B.E.; Duke, C.S. Biochemical strategies for growth of Gracilaria tikvahiae (Rhodophyta) in relation to light intensity and nitrogen availability. J. Phycol. 1984, 20, 488–495. [Google Scholar] [CrossRef]
- Dogra, B.; Amna, S.; Il Park, Y.; Park, J.K. Biochemical Properties of Water Soluble Polysaccharides from Photosynthetic Marine Microalgae Tetraselmis Species. Macromol. Res. 2017, 25, 172–179. [Google Scholar] [CrossRef]
- Kashif, S.A.; Hwang, Y.J.; Park, J.K. Potent biomedical applications of isolated polysaccharides from marine microalgae Tetraselmis species. Bioproc. Biosyst. Eng. 2018, 41, 1611–1620. [Google Scholar] [CrossRef]
- Kirst, C.O. Salinity tolerance of eukaryotic marine algae. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 21–53. [Google Scholar] [CrossRef]
- Díaz Bayona, K.C.; Garcés, L. Effect of different media on exopolysaccharide and biomass production by the green microalga Botryococcus braunii. J. Appl. Phycol. 2014, 26, 2087–2095. [Google Scholar] [CrossRef]
- Sun, L.; Wang, C.; Shi, Q.; Ma, C. Preparation of different molecular weight polysaccharides from Porphyridium cruentum and their antioxidant activities. Int. J. Biol. Macromol. 2009, 45, 42–47. [Google Scholar] [CrossRef]
- Abdala, R.; Chabrillón, M.; Cabello-Pasini, A.; Gómez-Pinchetti, J.; Figueroa, F. Characterization of polysaccharides from Hypnea spinella (Gigartinales) and Halopithys incurva (Ceramiales) and their effect on RAW 264.7 macrophage activity. J. Appl. Phycol. 2011, 23, 523–528. [Google Scholar]
- Fernando, I.P.S.; Sanjeewa, K.K.A.; Samarakoon, K.W.; Lee, W.W.; Kim, H.S.; Kim, E.A.; Gunasekara, U.K.D.S.S.; Abeytunga, D.T.U.; Nanayakkara, C.; de Silva, E.D.; et al. FTIR characterization and antioxidant activity of water soluble crude polysaccharides of Sri Lankan marine algae. Algae 2017, 2017, 75–86. [Google Scholar] [CrossRef] [Green Version]
- Parages, M.; Rico, R.; Abdala, R.; Chabrillón, M.; Sotiroudis, T.; Jiménez, C. Acidic polysaccharides of Arthrospira (Spirulina) platensis induce the synthesis of TNF-α in RAW macrophages. J. Appl. Phycol. 2012, 24, 1537–1546. [Google Scholar] [CrossRef]
- Cabassi, F.; Casu, B.; Perlin, A.S. Infrared absorption and raman scattering of sulfate groups of heparin and related glycosaminoglycans in aqueous solution. Carbohydr. Res. 1978, 63, 1–11. [Google Scholar] [CrossRef]
- Pereira, L.; Amado, A.M.; Critchley, A.T.; van de Velde, F.; Ribeiro-Claro, P.J.A. Identification of selected seaweed polysaccharides (phycocolloids) by vibrational spectroscopy (FTIR-ATR and FT-Raman). Food Hydrocoll. 2009, 23, 1903–1909. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L.; Gheda, S.F.; Ribeiro-Claro, P.J.A. Analysis by Vibrational Spectroscopy of Seaweed Polysaccharides with Potential Use in Food, Pharmaceutical, and Cosmetic Industries. Int. J. Carbohydr. Chem. 2013, 2013, 1–7. [Google Scholar] [CrossRef]
- Pereira, L.; Sousa, A.; Coelho, H.; Amado, A.M.; Ribeiro-Claro, P.J.A. Use of FTIR, FT-Raman and 13 C-NMR spectroscopy for identification of some seaweed phycocolloids. Biomol. Eng. 2003, 20, 223–228. [Google Scholar] [CrossRef] [Green Version]
- Meng, Y.; Yao, C.; Xue, S.; Yang, H. Application of fourier transform infrared (FT-IR) spectroscopy in determination of microalgal compositions. Bioresour. Technol. 2014, 151, 347–354. [Google Scholar] [CrossRef]
- Kermanshahi-Pour, A.; Sommer, T.J.; Anastas, P.T.; Zimmerman, J.B. Enzymatic and acid hydrolysis of Tetraselmis suecica for polysaccharide characterization. Bioresour. Technol. 2014, 173, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Becker, B.; Hard, K.; Melkonian, M.; Kamerling, J.P.G.; Vliegenthart, J.F. Identification of 3-deoxy-manno-2-octulosonic acid, 3-deoxy-5-O-methyl-rnanno-2-octulosonic acid and 3-deoxy-lyxo-2-heptulosaric acid in the cell wall (theca) of the green alga Tetraselmis striata Butcher (Prasinophyceae). Eur. J. Biochem. 1989, 182, 153–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, B.; Lommerse, J.P.M.; Melkonian, M.; Kamerling, J.P.; Vliegenthart, J.F.G. The structure of an acidic trisaccharide component from a cell wall polysaccharide preparation of the green alga Tetraselmis striata Butcher. Carbohydr. Res. 1995, 267, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Becker, B.; Melkonian, M.; Kamerling, J.P. The cell wall (theca) of Tetraselmis striata (Chlorophyta): Macromolecular compositionand structural elementsof the complex polysaccharides. J. Phycol. 1998, 34, 779–787. [Google Scholar] [CrossRef]
- Xiao, R.; Zheng, Y. Overview of microalgal extracellular polymeric substances (EPS) and their applications. Biotechnol. Adv. 2016, 34, 1225–1244. [Google Scholar] [CrossRef]
- Chen, X.; Song, L.; Wang, H.; Liu, S.; Yu, H.; Wang, X.; Li, R.; Liu, T.; Li, P. Partial Characterization, the Immune Modulation and Anticancer Activities of Sulfated Polysaccharides from Filamentous Microalgae Tribonema sp. Molecules 2019, 24, 322. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Wan, H.; Wang, R.; Hao, D. Sulfated polysaccharides from Phaeodactylum tricornutum: Isolation, structural characteristics, and inhibiting HepG2 growth activity in vitro. PeerJ. 2019, 7, e6409. [Google Scholar] [CrossRef] [Green Version]
- Abd El Baky, H.; Hanna El Baz, K.; El-Latife, S. Induction of Sulfated Polysaccharides in Spirulina platensis as Response to Nitrogen Concentration and its Biological Evaluation. J. Aquac. Res. Dev. 2013, 5, 1–8. [Google Scholar]
- Sun, L.; Wang, L.; Zhou, Y. Immunomodulation and antitumor activities of different-molecular-weight polysaccharides from Porphyridium cruentum. Carbohydr. Polym. 2012, 87, 1206–1210. [Google Scholar] [CrossRef]
- Gardeva, E.; Toshkova, R.; Yossifova, L.; Minkova, K.; Gigova, L. Cytotoxic and apoptogenic potential of red microalgal polysaccharides. Biotechnol. Biotechnol. Equip. 2012, 26, 3167–3172. [Google Scholar] [CrossRef]
- Yao, W.Z.; Veeraperumal, S.; Qiu, H.M.; Chen, X.Q.; Cheong, K.L. Anti-cancer effects of Porphyra haitanensis polysaccharides on human colon cancer cells via cell cycle arrest and apoptosis without causing adverse effects in vitro. 3 Biotech. 2020, 10, 386. [Google Scholar] [CrossRef] [PubMed]
- Nani, B.D.; Franchin, M.; Lazarini, J.G.; Freires, I.A.; da Cunha, M.G.; Bueno-Silva, B.; de Alencar, S.M.; Murata, R.M.; Rosalen, P.L. Isoflavonoids from Brazilian red propolis down-regulate the expression of cancer-related target proteins: A pharmacogenomic analysis. Phytother. Res. 2018, 32, 750–754. [Google Scholar] [CrossRef] [PubMed]
- Guillard, R. Culture of phytoplankton for feeding marine invertebrate animals. In Culture of Marine Invertebrates, 1st ed.; Smith, W., Chanley, M., Eds.; Springer: Boston, MA, USA, 1975; pp. 29–60. [Google Scholar]
- Sun, Y.; Wang, H.; Guo, G.; Pu, Y.; Yan, B. The isolation and antioxidant activity of polysaccharides from the marine microalgae Isochrysis galbana. Carbohydr. Polym. 2014, 113, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Morris Quevedo, H.J.; Martínez Manrique, C.E.; Abdala Díaz, R.T.; Cobas Pupo, G. Evidencias preliminares de la actividad inmunomoduladora de la fracción polisacárida de origen marino Pc-1. Rev. Cuba. Oncol. 2000, 16, 171–177. [Google Scholar]
- Arredondo-Vega, B.O.; Voltolina, D. Concentración, recuento celular y tasa de crecimiento. In Métodos y Herramientas Analíticas en la Evaluación de la Biomasa Microalgal, 1st ed.; Arredondo, B.O., Voltolina, D., Eds.; CIBNOR: La Paz, B.C.S., Mexico, 2007; pp. 21–29. [Google Scholar]
- Lourenço, S.O.; Barbarino, E.; Lavín, P.L.; Lanfer Marquez, U.M.; Aidar, E. Distribution of intracellular nitrogen in marine microalgae: Calculation of new nitrogen-to-protein conversion factors. Eur. J. Phycol. 2004, 39, 17–32. [Google Scholar] [CrossRef]
- Folch, J.; Less, M.; Sloane Stanley, G.H. A Simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Biochem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Folin, O.; Ciocalteau, V. On tyrosine and tryptophane determinations in proteins. J. Biol. Chem. 1927, 73, 627–648. [Google Scholar]
- Ritchie, R.J. Consistent sets of spectrophotometric chlorophyll equations for acetone, methanol and ethanol solvents. Photosynth. Res. 2006, 89, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, R.J. Universal chlorophyll equations for estimating chlorophylls a, b, c, and d and total chlorophylls in natural assemblages of photosynthetic organisms using acetone, methanol, or ethanol solvents. Photosynthetica 2008, 46, 115–126. [Google Scholar] [CrossRef]
- Strickland, J.D.H.; Parsons, T.R. A Practical Handbook of Seawater Analysis, 2nd ed.; Fisheries Research Board of Canada: Ottawa, ON, Canada, 2008; pp. 1–310. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Bio. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Vijayabaskar, P.; Vaseela, N. In vitro antioxidant properties of sulfated polysaccharide from brown marine algae Sargassum tenerrimum. Asian Pac. J. Trop. Dis. 2012, S890–S896. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. Lebensm. Wiss. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Underwood, A.J. Experiments in Ecology: Their Logical Design and Interpretation Using Analysis of Variance, 1st ed.; Cambridge University Press: Cambridge, UK, 1997; pp. 1–522. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population Parameter | Autotrophic | Heterotrophic |
|---|---|---|
| Cell density (cell mL−1) | 2.0 × 106 ± 1.6 × 104 | 3.6 × 107 ± 7.7 × 104 * |
| Cell concentration (g L−1) | 5.5 ± 0.5 | 10.2 ± 0.7 * |
| Specific growth rate (µ) (d−1) | 0.3 ± 0.2 | 0.3 ± 0.2 |
| Biovolume (µm3 mL−1) | 8.8 × 108 ± 2.8 × 108 | 1.2 × 109 ± 3.0 × 108 * |
| Cell volume (µm3) | 521 ± 33 * | 30 ± 2 |
| Elemental Analysis (%) | Autotrophic | Heterotrophic |
|---|---|---|
| TC | 25.7 ± 0.4 | 31.0 ± 0.1 * |
| TN | 3.5 ± 0.2 | 4.3 ± 0.1 * |
| C/N | 7.3 ± 0.2 | 7.2 ± 0.1 |
| Biochemical Composition | Autotrophic | Heterotrophic |
|---|---|---|
| Proteins | 16.76 ± 0.40 | 20.78 ± 0.14 * |
| Lipids | 6.13 ± 0.12 | 7.96 ± 0.10 * |
| Carbohydrates | 24.31 ± 0.32 | 28.18 ± 0.37 * |
| Ash | 34.88 ± 0.08 | 33.07 ± 1.30 |
| Moisture | 17.93 ± 0.52 * | 10.01 ± 1.16 |
| Culture | Phenols (mg Eq Phloroglucinol) | Antioxidant Activity | |
|---|---|---|---|
| ABTS (µmol TE g−1 DW) | DPPH (µmol TE g−1 DW) | ||
| Autotrophic | 3.88 ± 0.03 | 24.25 ± 0.70 | 3.49 ± 0.61 |
| Heterotrophic | 5.56 ± 0.10 * | 80.17 ± 0.95 * | 6.35 ± 0.91 * |
| Culture | Exopolysaccharide Type | % TC | % TN | C/N | % S |
|---|---|---|---|---|---|
| Autotrophic | Total | 9.60 ± 0.10 c | 0.53 ± 0.01 d | 18.28 ± 0.17 a | 0.00 |
| Autotrophic | Acid | 9.02 ± 0.08 d | 0.71 ± 0.01 b | 12.75 ± 0.10 c | 0.00 |
| Heterotrophic | Total | 9.84 ± 0.07 b | 0.66 ± 0.02 c | 14.91 ± 0.10 b | 0.33 ± 0.08 b |
| Heterotrophic | Acid | 11.96 ± 0.09 a | 0.80 ± 0.01 a | 14.88 ± 0.12 b | 3.47 ± 0.10 a |
| Monosaccharide | Autotrophic Total (%) | Autotrophic Acid (%) | Heterotrophic Total (%) | Heterotrophic Acid (%) |
|---|---|---|---|---|
| Arabinose | 5.23 | - | - | - |
| Ribose | 0.83 | 0.65 | 1.22 | 0.33 |
| Rhamnose | - | - | 1.36 | - |
| Fucose | - | - | 0.38 | 0.35 |
| Xylose | - | 3.03 | - | 0.30 |
| Mannose | 6.64 | 1.57 | 36.15 | 34.49 |
| Galacturonic acid | <0.10 | 2.93 | 2.45 | 2.95 |
| Galactopyranoside | 5.11 | 27.06 | 6.76 | 8.14 |
| Galactose | 25.27 | 9.96 | 3.00 | 2.93 |
| Glucose | 35.46 | 34.70 | 23.32 | 37.57 |
| Glucuronic acid | 21.47 | 20.10 | 25.36 | 22.94 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parra-Riofrío, G.; García-Márquez, J.; Casas-Arrojo, V.; Uribe-Tapia, E.; Abdala-Díaz, R.T. Antioxidant and Cytotoxic Effects on Tumor Cells of Exopolysaccharides from Tetraselmis suecica (Kylin) Butcher Grown Under Autotrophic and Heterotrophic Conditions. Mar. Drugs 2020, 18, 534. https://doi.org/10.3390/md18110534
Parra-Riofrío G, García-Márquez J, Casas-Arrojo V, Uribe-Tapia E, Abdala-Díaz RT. Antioxidant and Cytotoxic Effects on Tumor Cells of Exopolysaccharides from Tetraselmis suecica (Kylin) Butcher Grown Under Autotrophic and Heterotrophic Conditions. Marine Drugs. 2020; 18(11):534. https://doi.org/10.3390/md18110534
Chicago/Turabian StyleParra-Riofrío, Geovanna, Jorge García-Márquez, Virginia Casas-Arrojo, Eduardo Uribe-Tapia, and Roberto Teófilo Abdala-Díaz. 2020. "Antioxidant and Cytotoxic Effects on Tumor Cells of Exopolysaccharides from Tetraselmis suecica (Kylin) Butcher Grown Under Autotrophic and Heterotrophic Conditions" Marine Drugs 18, no. 11: 534. https://doi.org/10.3390/md18110534