Antibacterial Activity of Chromomycins from a Marine-Derived Streptomyces microflavus

 and
and
Abstract
:1. Introduction
2. Results
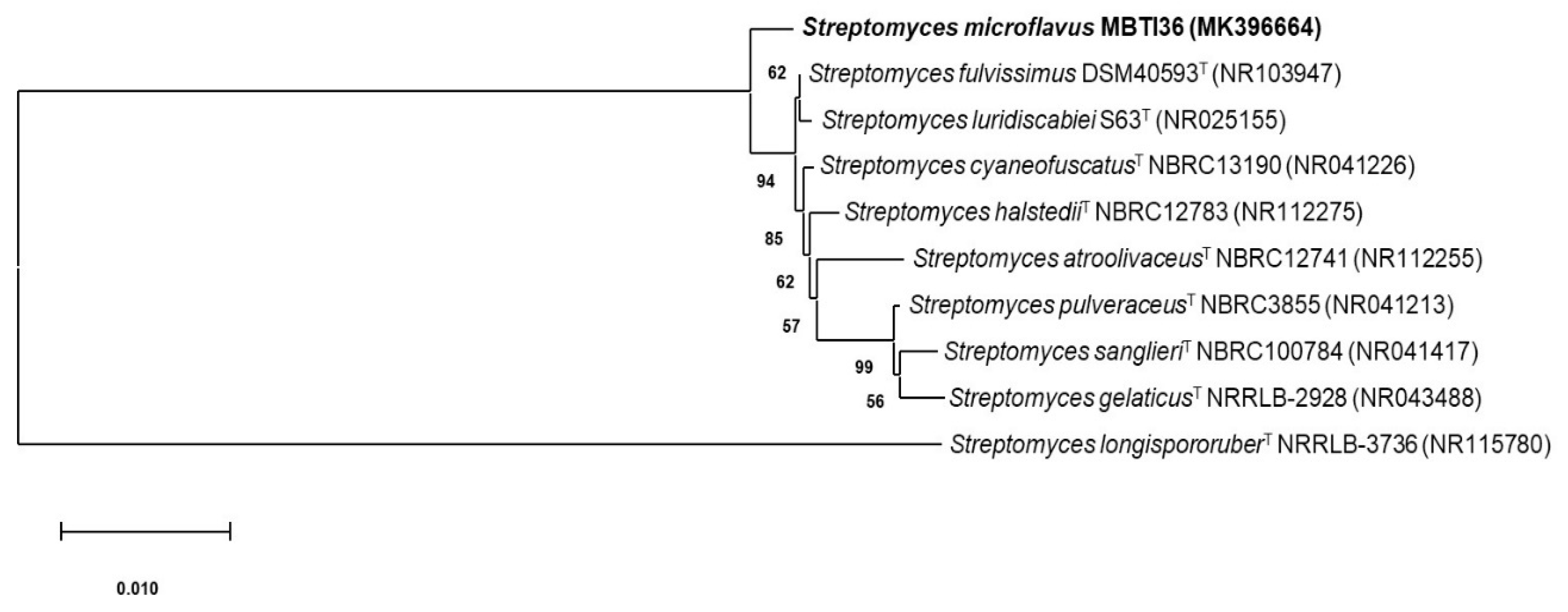
2.1. Taxonomy and Phylogenetic Analysis of MBTI36
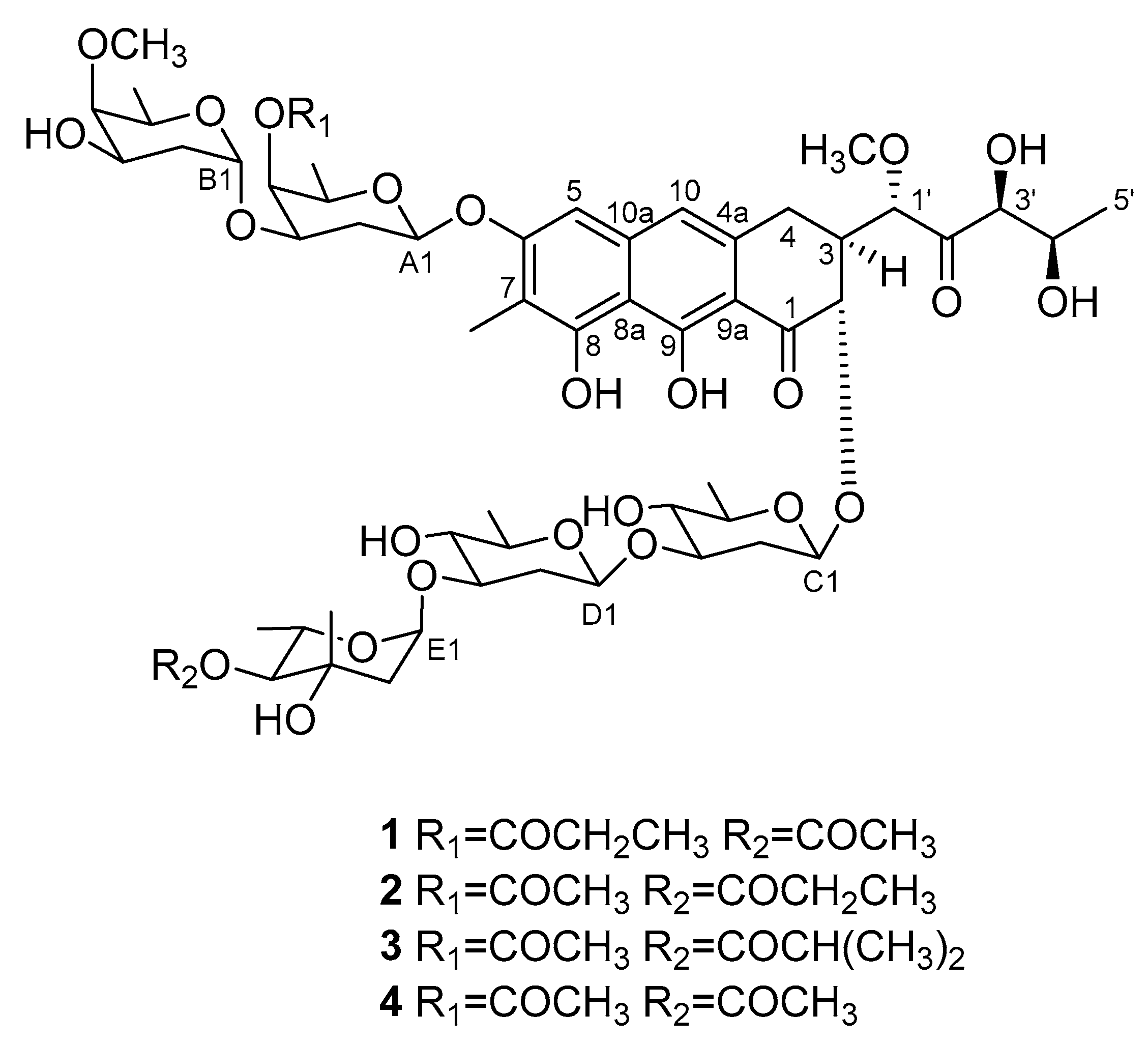
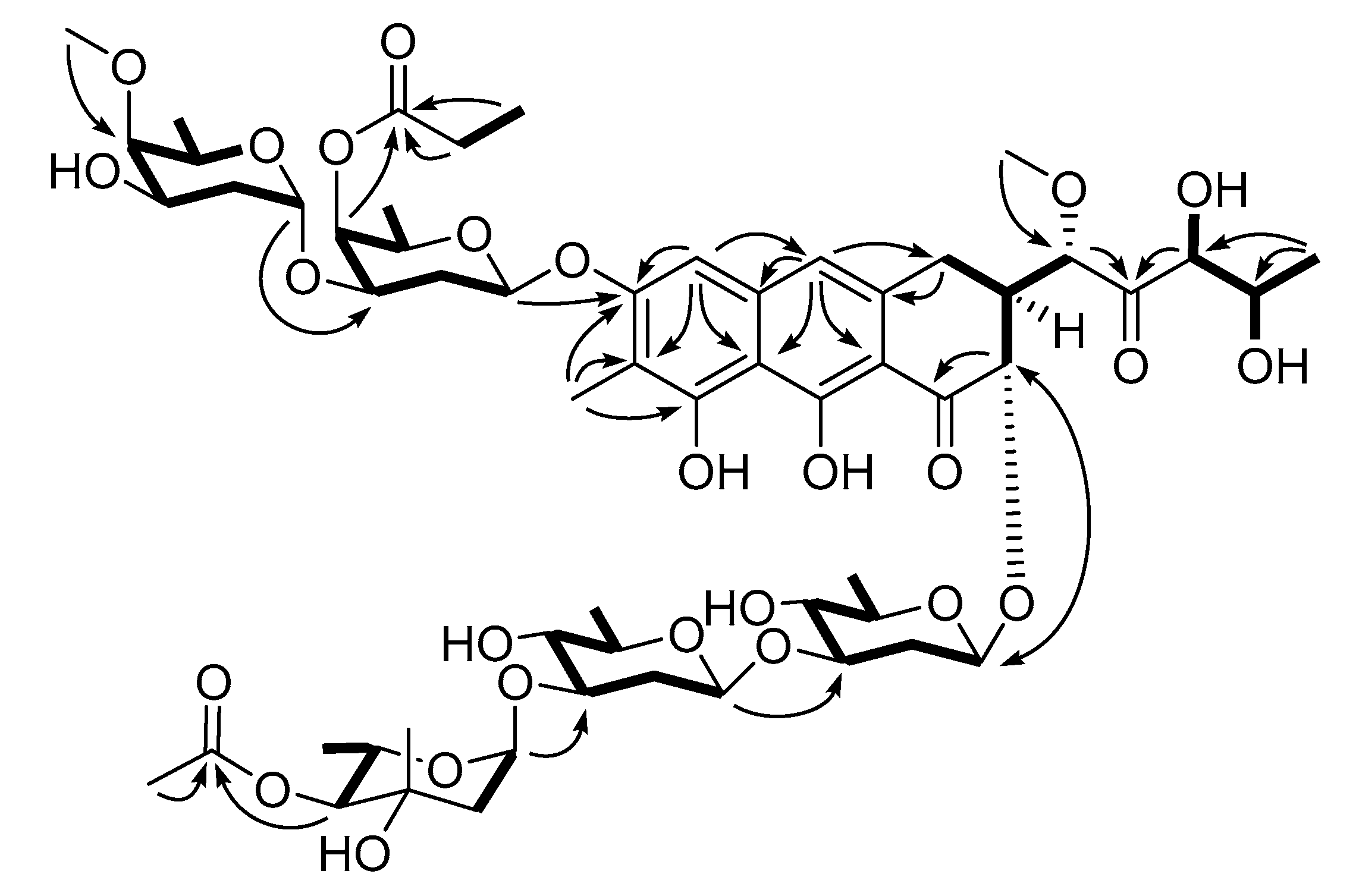
2.2. Isolation and Structural Elucidation of Compounds 1–4
2.3. Antimicrobial Activity of Compounds 1–4
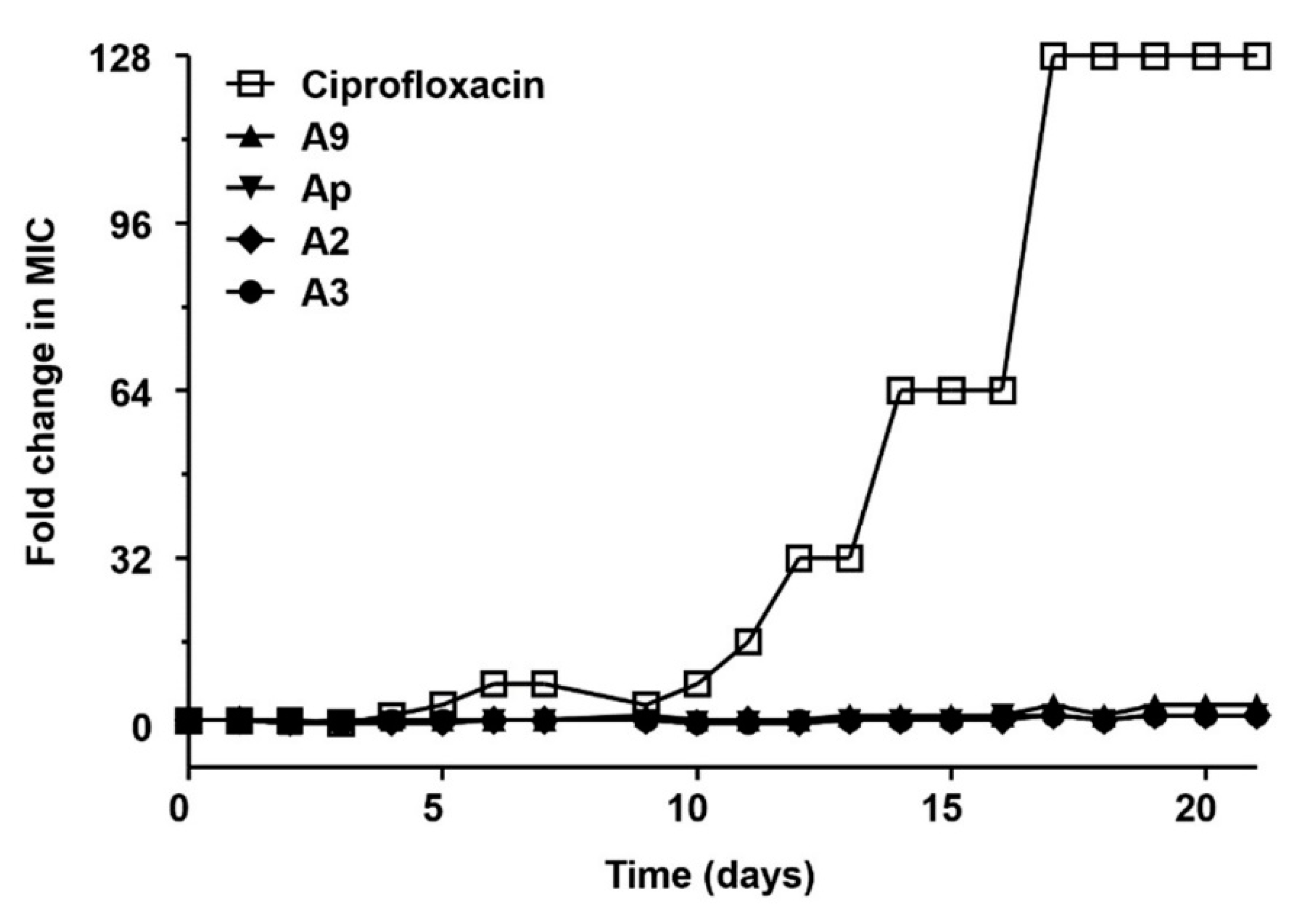
2.4. Multi-Step Resistance Development
3. Discussion
4. Materials and Methods
4.1. General Experimental Equipments
4.2. Taxonomic Identification of the Chromomycin Producing Microorganism
4.3. Cultivation
4.4. Extraction and Isolation
4.5. Antibacterial Activity Assays
4.6. Antifungal Activity Assays
4.7. Multi-Step Resistance Development Assays
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kupferschmidt, K. Resistance fighters. Science 2016, 352, 758–761. [Google Scholar] [CrossRef] [Green Version]
- Holmes, A.H.; Moore, L.S.; Sundsfjord, A.; Steinbakk, M.; Regmi, S.; Karkey, A.; Guerin, P.J.; Piddock, L.J. Understanding the mechanisms and drivers of antimicrobial resistance. Lancet 2016, 387, 176–187. [Google Scholar] [CrossRef]
- Hutchings, M.I.; Truman, A.W.; Wilkinson, B. Antibiotics: Past, present and future. Curr. Opin. Microbiol. 2019, 51, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Bérdy, J. Thoughts and facts about antibiotics: Where we are now and where we are heading. J. Antibiot. (Tokyo) 2012, 65, 385–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramani, R.; Aalbersberg, W. Marine actinomycetes: An ongoing source of novel bioactive metabolites. Microbiol. Res. 2012, 167, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Bull, A.T.; Ward, A.C.; Goodfellow, M. Search and discovery strategies for biotechnology: The paradigm shift. Microbiol. Mol. Biol. Rev. 2000, 64, 573–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramesh, S.; Mathivanan, N. Screening of marine actinomycetes isolated from the Bay of Bengal, India for antimicrobial activity and industrial enzymes. World J. Microbiol. Biotechnol. 2009, 25, 2103–2111. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2019, 36, 122–173. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Okamura, N.; Utagawa, K.; Ito, Y.; Watanabe, M. Studies on the antitumor activity of chromomycin A3. Sci. Rep. Res. Inst. Tohoku. Univ. Med. 1960, 9, 224–232. [Google Scholar]
- Lombó, F.; Menéndez, N.; Salas, J.A.; Méndez, C. The aureolic acid family of antitumor compounds: Structure, mode of action, biosynthesis, and novel derivatives. Appl. Microbiol. Biotechnol. 2006, 73, 1–14. [Google Scholar] [CrossRef]
- Ogawa, H.; Yamashita, Y.; Katahira, R.; Chiba, S.; Iwasaki, T.; Ashizawa, T.; Nakano, H. UCH9, a new antitumor antibiotic produced by Streptomyces: I. Producing organism, fermentation, isolation and biological activities. J. Antibiot. 1998, 51, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Pinto, F.C.L.; Silveira, E.R.; Vasconcelos, A.L.C.; Florêncio, K.G.D.; Oliveira, F.A.S.; Sahm, B.B.; Costa-Lotufo, L.V.; Bauermeister, A.; Lopes, N.P.; Wilke, D.V.; et al. Dextrorotatory chromomycins from the marine Streptomyces sp. associated to Palythoa caribaeorum. J. Braz. Chem. Soc. 2020, 31, 143–152. [Google Scholar] [CrossRef]
- Mir, M.A.; Majee, S.; Das, S.; Dasgupta, D. Association of chromatin with anticancer antibiotics, mithramycin and chromomycin A3. Bioorg. Med. Chem. 2003, 11, 2791–2801. [Google Scholar] [CrossRef]
- Hu, Y.; Espindola, A.P.D.M.; Stewart, N.A.; Wei, S.; Posner, B.A.; MacMillan, J.B. Chromomycin SA analogs from a marine-derived Streptomyces sp. Bioorg. Med. Chem. 2011, 19, 5183–5189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, N.; Rutigliano, C.; Passadore, M.; Tomassetti, M.; Pippo, L.; Mischiati, C.; Feriotto, G.; Gambari, R. Targeting of the HIV-1 long terminal repeat with chromomycin potentiates the inhibitory effects of a triplex-forming oligonucleotide on Sp1-DNA interactions and in vitro transcription. Biochem. J. 1997, 326, 919–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barceló, F.; Ortiz-Lombardía, M.; Martorell, M.; Oliver, M.; Méndez, C.; Salas, J.A.; Portugal, J. DNA binding characteristics of mithramycin and chromomycin analogues obtained by combinatorial biosynthesis. Biochemistry 2010, 49, 10543–10552. [Google Scholar] [CrossRef] [PubMed]
- Zihlif, M.; Catchpoole, D.R.; Stewart, B.W.; Wakelin, L.P. Effects of DNA minor groove binding agents on global gene expression. Cancer Genom. Proteom. 2010, 7, 323–330. [Google Scholar]
- Murase, H.; Noguchi, T.; Sasaki, S. Evaluation of simultaneous binding of chromomycin A3 to the multiple sites of DNA by the new restriction enzyme assay. Bioorg. Med. Chem. Lett. 2018, 28, 1832–1835. [Google Scholar] [CrossRef] [PubMed]
- Norio, S.; Eiji, H.; Takashi, S.; Mitsuko, A.; Kimiaki, M. Chromomycin Ap. Japan Patent JP 52102202, 23 February 1976. [Google Scholar]
- Yoshimura, Y.; Koenuma, M.; Matsumoto, K.; Tori, K.; Terui, Y. NMR studies of chromomycins, olivomycins, and their derivatives. J. Antibiot. 1988, 41, 53–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, M.; Kawamatsu, Y.; Kawashima, K.; Shinohara, M.; Tanaka, K.; Tatsuoka, S.; Nakanishi, K. Chromomycin A2, A3 and A4. Tetrahedron 1967, 23, 421–437. [Google Scholar] [CrossRef]
- Toume, K.; Tsukahara, K.; Ito, H.; Ariai, M.A.; Ishibashi, M. Chromomycins A2 and A3 from marine actinomycetes with TRAIL resistance-overcoming and Wnt signal inhibitory activities. Mar. Drugs 2014, 12, 3466–3476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, C.; Kosowska-Shick, K.; McGhee, P.; Dewasse, B.; Beachel, L.; Appelbaum, P.C. Resistance selection studies comparing the activity of razupenem (PTZ601) to vancomycin and linezolid against eight methicillin-resistant and two methicillin-susceptible Staphylococcus aureus strains. Antimicrob. Agents Chemother. 2009, 53, 3118–3121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosowska-Shick, K.; Clark, C.; Pankuch, G.A.; McGhee, P.; Dewasse, B.; Beachel, L.; Appelbaum, P.C. Activity of telavancin against staphylococci and enterococci determined by MIC and resistance selection studies. Antimicrob. Agents Chemother. 2009, 53, 4217–4224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, E.Y.; Trucksis, M.; Hooper, D.C. Quinolone resistance mutations in topoisomerase IV: Relationship to the flqA locus and genetic evidence that topoisomerase IV is the primary target and DNA gyrase is the secondary target of fluoroquinolones in Staphylococcus aureus. Antimicrob. Agents Chemother. 1996, 40, 1881–1888. [Google Scholar] [CrossRef] [Green Version]
- Ferrero, L.; Cameron, B.; Crouzet, J. Analysis of gyrA and grlA mutations in stepwise-selected ciprofloxacin-resistant mutants of Staphylococcus aureus. Antimicrob. Agents Chemother. 1995, 39, 1554–1558. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 11th ed.; M07; CLSI: Wayne, PA, USA, 2018. [Google Scholar]
- Clinical and Laboratory Standards Institute. Reference Methods for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi, 3rd ed.; M38; CLSI: Wayne, PA, USA, 2017. [Google Scholar]
- Farrell, D.J.; Robbins, M.; Rhys-Williams, W.; Love, W.G. Investigation of the potential for mutational resistance to XF-73, retapamulin, mupirocin, fusidic acid, daptomycin, and vancomycin in methicillin-resistant Staphylococcus aureus isolates during a 55-passage study. Antimicrob. Agents Chemother. 2011, 55, 1177–1181. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δC, Type | δH (J in Hz) | Position | δC, Type | δH (J in Hz) |
|---|---|---|---|---|---|
| 1 | 202.3, C | 4-O-methyl-d-oliose (Sugar B) | |||
| 2 | 76.1, CH | 4.72, d (11.4) | B1 | 95.4, CH | 5.11, br s |
| 3 | 44.0, CH | 2.60, m | B2 | 33.7, CH2 | 1.76, m; 1.73, m |
| 4 | 27.1, CH2 | 3.10, m | B3 | 66.0, CH | 3.96, m |
| 2.67, dd (16.6, 3.5) | B4 | 81.7, CH | 3.22, d (2.8) | ||
| 5 | 101.0, CH | 6.64, s | B5 | 66.9, CH | 3.87, q (6.6) |
| 6 | 159.9, C | B6 | 17.4, CH | 1.28, d (6.3) | |
| 7 | 111.9, C | B4-OCH3 | 62.6, CH3 | 3.60, s | |
| 8 | 156.3, C | d-olivose (Sugar C) | |||
| 9 | 165.5, C | C1 | 100.5, CH | 5.10, dd (9.7, 1.3) | |
| 10 | 117.2, CH | 6.75, s | C2 | 37.7, CH2 | 2.48, m; 1.70, m |
| 4a | 134.8, C | C3 | 82.5, CH | 3.61, m | |
| 8a | 108.3, C | C4 | 75.3, CH | 3.12, m | |
| 9a | 108.3, C | C5 | 72.3, CH | 3.38, m | |
| 10a | 138.6, C | C6 | 18.2, CH3 | 1.35, d (5.6) | |
| 7-CH3 | 8.4, CH3 | 8.40, s | d-olivose (Sugar D) | ||
| 8-OH | 9.81, s | D1 | 99.9, CH | 4.60, dd (9.6, 1.7) | |
| 1′ | 82.0, CH | 4.70, d (1.5) | D2 | 37.3, CH2 | 2.28, m; 1.70, m |
| 2′ | 211.3, C | D3 | 80.9, CH | 3.50, m | |
| 3′ | 78.3, C | 4.22, br s | D4 | 75.4, CH | 3.12, m |
| 4′ | 68.1, C | 4.36, m | D5 | 72.5, CH | 3.30, m |
| 5′ | 20.8, CH3 | 1.37, d (5.6) | D6 | 18.0, CH3 | 1.24, d (6.1) |
| 1′-OCH3 | 59.9, CH3 | 3.52, s | l-chromose (Sugar E) | ||
| 4-O-propioyl-d-oliose (Sugar A) | E1 | 97.3, CH | 5.02, dd (3.6, 1.8) | ||
| A1 | 97.6, C | 5.21, dd (9.7, 2.0) | E2 | 43.9, CH2 | 2.04, m; 2.00, m |
| A2 | 33.2, CH2 | 2.19, m; 2.05, m | E3 | 70.8, C | |
| A3 | 70.2, CH | 3.98, m | E4 | 79.9, CH | 4.61, d (9.3) |
| A4 | 67.2, CH | 5.18, d (2.9) | E5 | 67.2, CH | 3.98, m |
| A5 | 70.0 | 3.82, q (6.5) | E6 | 18.0, CH3 | 1.38, d (5.4) |
| A6 | 17.0, C | 1.28, d (6.3) | E3-CH3 | 23.2, CH3 | 1.35, s |
| COCH2CH3 | 174.6, C | COCH3 | 171.7, C | ||
| COCH2CH3 | 27.8, CH2 | 2.46, q (7.5) | COCH3 | 21.1, CH3 | 2.14, s |
| COCH2CH3 | 9.7, CH3 | 1.19, t (7.5) | |||
| Compound | MIC (μg/mL) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Gram (+) Bacteria | Gram (−) Bacteria | Fungi | ||||||||
| A | B | C | D | E | F | G | H | I | J | |
| 1 | 0.03 | 0.5 | 0.13 | 0.5 | >128 | >128 | >128 | >128 | >128 | >128 |
| 2 | 0.13 | 0.5 | 0.13 | 1 | >128 | >128 | >128 | >128 | >128 | >128 |
| 3 | 0.06 | 0.5 | 0.06 | 0.5 | >128 | >128 | >128 | >128 | >128 | >128 |
| 4 | 0.13 | 0.5 | 0.13 | 0.5 | >128 | >128 | >128 | >128 | >128 | >128 |
| Ampicillin | 0.06 | 0.5 | 0.25 | 0.25 | 128 | 32 | ||||
| Tetracycline | 0.06 | 0.13 | 0.25 | 0.25 | 0.5 | 0.5 | ||||
| Amphotericin B | 0.5 | 1 | 1 | 1 | ||||||
| Microorganism | MIC (μg/mL) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Dap | Van | Pla | Lin | Cip | 1 | 2 | 3 | 4 | |
| CCARM0027 a | 8 | 0.5 | 4 | 2 | 0.25 | 0.13 | 0.13 | 0.06 | 0.13 |
| CCARM0204 a | 2 | 0.25 | 4 | 1 | 0.25 | 0.06 | 0.06 | 0.03 | 0.06 |
| CCARM0205 a | 1 | 0.13 | 2 | 1 | 0.25 | 0.06 | 0.13 | 0.06 | 0.06 |
| CCARM3640 a | 8 | 0.25 | 4 | 2 | 0.25 | 0.13 | 0.25 | 0.06 | 0.13 |
| CCARM3089 b | >32 | 1 | 8 | 2 | >32 | 0.13 | 0.25 | 0.13 | 0.13 |
| CCARM3090 b | >32 | 1 | 8 | 1 | >32 | 0.13 | 0.25 | 0.13 | 0.13 |
| CCARM3634 b | >32 | 0.5 | 8 | 2 | >32 | 0.13 | 0.13 | 0.06 | 0.13 |
| CCARM3635 b | >32 | 1 | 8 | 2 | >32 | 0.13 | 0.06 | 0.06 | 0.13 |
| ATCC43300 b | >32 | 1 | 4 | 2 | 0.25 | 0.13 | 0.13 | 0.06 | 0.13 |
| ATCC700787 b | >32 | 2 | 8 | 2 | 0.13 | 0.13 | 0.25 | 0.25 | 0.13 |
| ATCC700788 b | >32 | 2 | 8 | 2 | 16 | 0.13 | 0.25 | 0.13 | 0.13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, E.; Kwon, O.-S.; Chung, B.; Lee, J.; Sun, J.; Shin, J.; Oh, K.-B. Antibacterial Activity of Chromomycins from a Marine-Derived Streptomyces microflavus. Mar. Drugs 2020, 18, 522. https://doi.org/10.3390/md18100522
Cho E, Kwon O-S, Chung B, Lee J, Sun J, Shin J, Oh K-B. Antibacterial Activity of Chromomycins from a Marine-Derived Streptomyces microflavus. Marine Drugs. 2020; 18(10):522. https://doi.org/10.3390/md18100522
Chicago/Turabian StyleCho, Eunji, Oh-Seok Kwon, Beomkoo Chung, Jayho Lee, Jeongyoon Sun, Jongheon Shin, and Ki-Bong Oh. 2020. "Antibacterial Activity of Chromomycins from a Marine-Derived Streptomyces microflavus" Marine Drugs 18, no. 10: 522. https://doi.org/10.3390/md18100522