Ectoine from Bacterial and Algal Origin Is a Compatible Solute in Microalgae


Abstract
:1. Introduction
2. Results and Discussion
2.1. Thalassiosira weissflogii
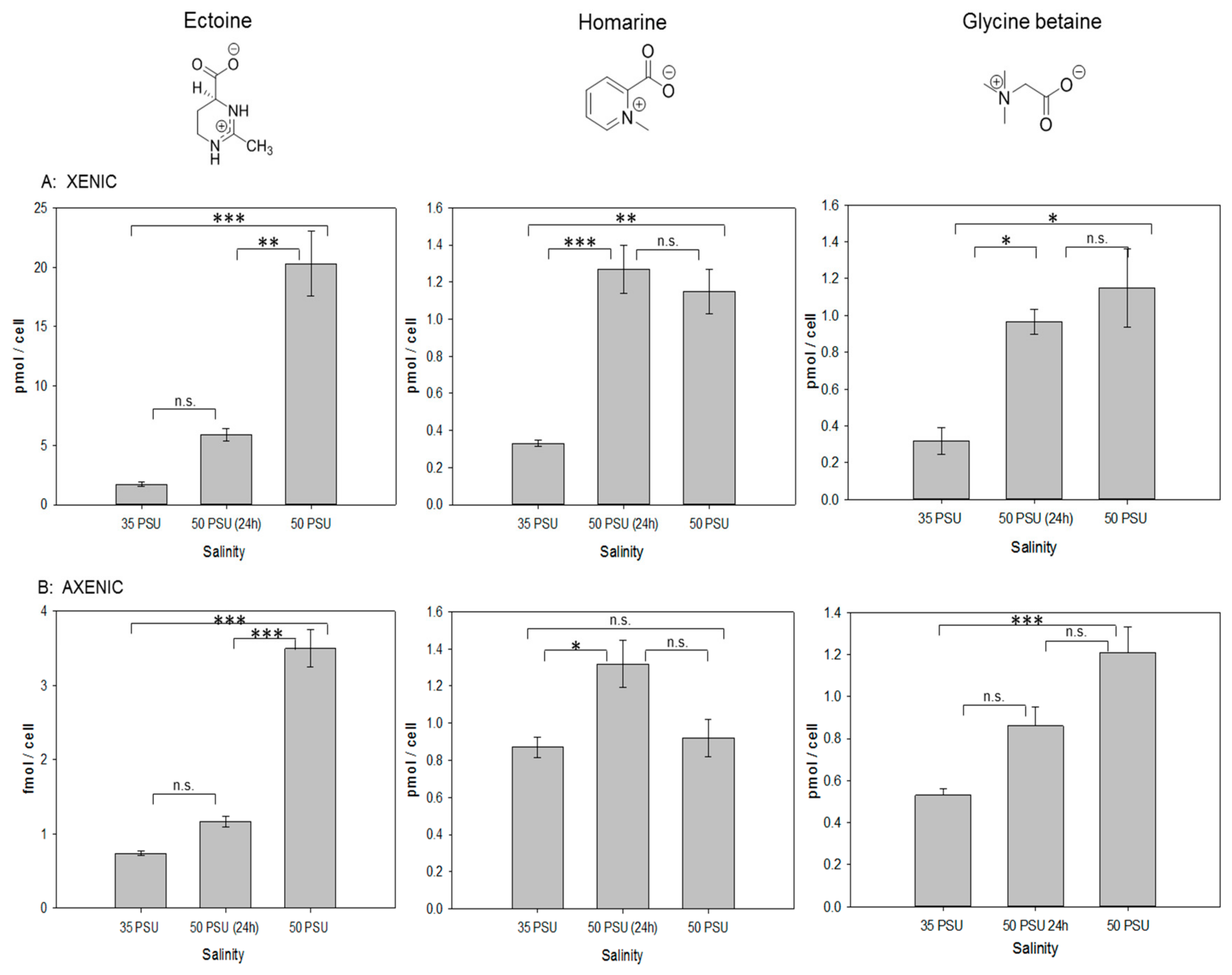
2.2. Response to Osmotic Stress of T. weissflogii
2.3. Ectoine Uptake by T. weissflogii
2.4. Ectoine in Phaeodactylum tricornutum
2.5. Ectoine in Other Microalgae
3. Materials and Methods
3.1. Cultivation of Microalgae
3.2. Salinity Treatment
3.3. Cell Counting and Size Measurement
3.4. Sample Preparation
3.5. Equipment
3.6. Osmolyte Analysis
3.7. Uptake of Labeled Ectoine by Marine Diatoms
3.8. Homology Search
3.9. Synthesis of D3-ectoine
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Moran, M.A.; Durham, B.P. Sulfur metabolites in the pelagic ocean. Nat. Rev. Microbiol. 2019, 17, 665–678. [Google Scholar] [CrossRef] [PubMed]
- Gebser, B.; Pohnert, G. Synchronized regulation of different zwitterionic metabolites in the osmoadaption of phytoplankton. Mar. Drugs 2013, 11, 2168–2182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoch, D.C. Dimethylsulfoniopropionate: Its sources, role in the marine food web, and biological degradation to dimethylsulfide. Appl. Environ. Microbiol. 2002, 68, 5804–5815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thume, K.; Gebser, B.; Chen, L.; Meyer, N.; Kieber, D.J.; Pohnert, G. The metabolite dimethylsulfoxonium propionate extends the marine organosulfur cycle. Nature 2018, 563, 412–415. [Google Scholar] [CrossRef]
- Curson, A.R.J.; Todd, J.D.; Sullivan, M.J.; Johnston, A.W.B. Catabolism of dimethylsulphoniopropionate: Microorganisms, enzymes and genes. Nat. Rev. Microbiol. 2011, 9, 849–859. [Google Scholar] [CrossRef]
- Smyth, K.; Elliott, M. Effects of changing salinity on the ecology of the marine environment. In Stressors in the Marine Environment; Solan, M., Whiteley, N., Eds.; Oxford University Press: Oxford, UK, 2016; pp. 161–174. [Google Scholar] [CrossRef]
- Spielmeyer, A.; Gebser, B.; Pohnert, G. Investigations of the uptake of dimethylsulfoniopropionate by phytoplankton. ChemBioChem 2011, 12, 2276–2279. [Google Scholar] [CrossRef]
- Dickschat, J.S.; Rabe, P.; Citron, C.A. The chemical biology of dimethylsulfoniopropionate. Org. Biomol. Chem. 2015, 13, 1954–1968. [Google Scholar] [CrossRef]
- Keller, M.D.; Kiene, R.P.; Matrai, P.A.; Bellows, W.K. Production of glycine betaine and dimethylsulphoniopropionate in marine phytoplankton. II. N-limited chemostat cultures. Mar. Biol. 1999, 135, 249–257. [Google Scholar] [CrossRef]
- Spielmeyer, A.; Pohnert, G. Direct quantification of dimethylsulfoniopropionate (DMSP) with hydrophilic interaction liquid chromatography/mass spectrometry. J. Chromatogr. B 2010, 878, 3238–3242. [Google Scholar] [CrossRef]
- Spielmeyer, A.; Gebser, B.; Pohnert, G. Dimethylsulfide sources from microalgae: Improvement and application of a derivatization-based method for the determination of dimethylsulfoniopropionate and other zwitterionic osmolytes in phytoplankton. Mar. Chem. 2011, 124, 48–56. [Google Scholar] [CrossRef]
- Zhao, P.P.; Gu, W.; Wu, S.; Huang, A.; He, L.; Xie, X.; Gao, S.; Zhang, B.; Niu, J.; Lin, A.P.; et al. Silicon enhances the growth of Phaeodactylum tricornutum Bohlin under green light and low temperature. Sci. Rep. 2014, 4, 3958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bussard, A.; Corre, E.; Hubas, C.; Duvernois-Berthet, E.; Le Corguille, G.; Jourdren, L.; Coulpier, F.; Claquin, P.; Lopez, P.J. Physiological adjustments and transcriptome reprogramming are involved in the acclimation to salinity gradients in diatoms. Environ. Microbiol. 2017, 19, 909–925. [Google Scholar] [CrossRef] [PubMed]
- Garcia, N.; Lopez-Elias, J.A.; Miranda, A.; Martinez-Porchas, M.; Huerta, N.; Garcia, A. Effect of salinity on growth and chemical composition of the diatom Thalassiosira weissflogii at three culture phases. Lat. Am. J. Aquat. Res. 2012, 40, 435–440. [Google Scholar] [CrossRef]
- Ito, T.; Asano, Y.; Tanaka, Y.; Takabe, T. Regulation of biosynthesis of dimethylsulfoniopropionate and its uptake in sterile mutant of Ulva pertusa (Chlorophyta). J. Phycol. 2011, 47, 517–523. [Google Scholar] [CrossRef]
- Takagi, M.; Karseno; Yoshida, T. Effect of salt concentration on intracellular accumulation of lipids and triacylglyceride in marine microalgae Dunaliella cells. J. Biosci. Bioeng. 2006, 101, 223–226. [Google Scholar] [CrossRef] [Green Version]
- Keller, M.D.; Kiene, R.P.; Matrai, P.A.; Bellows, W.K. Production of glycine betaine and dimethylsulphoniopropionate in marine phytoplankton. I. Batch cultures. Mar. Biol. 1999, 135, 237–248. [Google Scholar] [CrossRef]
- Dickson, D.M.J.; Kirst, G.O. The role of b-dimethylsulphoniopropionate, glycine betaine and homarine in the osmoacclimation of Platymonas subcordiformis. Planta 1986, 167, 536–543. [Google Scholar] [CrossRef]
- Galinski, E.A.; Pfeiffer, H.-P.; Trüper, H.G. 1,4,5,6-Tetrahydro-2-methyl-4-pyrimidinecarboxylic acid. A novel cyclic amino acid from halophilic phototrophic bacteria of the genus Ectothiorhodospira. Eur. J. Biochem. 1985, 149, 135–139. [Google Scholar] [CrossRef]
- Kolp, S.; Pietsch, M.; Galinski, E.A.; Gutschow, M. Compatible solutes as protectants for zymogens against proteolysis. Biochim. Biophys. Acta 2006, 1764, 1234–1242. [Google Scholar] [CrossRef]
- Kunte, H.J.; Lentzen, G.; Galinski, E.A. Industrial production of the cell protectant ectoine: Protection mechanisms, processes and products. Curr. Biotechnol. 2014, 3, 10–25. [Google Scholar] [CrossRef] [Green Version]
- Waditee-Sirisattha, R.; Kageyama, H.; Takabe, T. Halophilic microorganism resources and their applications in industrial and environmental biotechnology. AIMS Microbiol. 2016, 2, 42–54. [Google Scholar] [CrossRef]
- Weinisch, L.; Kuhner, S.; Roth, R.; Grimm, M.; Roth, T.; Netz, D.J.A.; Pierik, A.J.; Filker, S. Identification of osmoadaptive strategies in the halophile, heterotrophic ciliate Schmidingerothrix salinarum. PLoS Biol. 2018, 16, e2003892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinisch, L.; Kirchner, I.; Grimm, M.; Kuhner, S.; Pierik, J.J.; Rossello-Mora, R.; Filker, S. Glycine betaine and ectoine are the major compatible solutes used by four different halophilic heterotrophic ciliates. Microb. Ecol. 2019, 77, 317–331. [Google Scholar] [CrossRef]
- Harding, T.; Roger, A.J.; Simpson, A.G.B. Adaptations to high salt in a halophilic protist: Differential expression and gene acquisitions through duplications and gene transfers. Front. Microbiol. 2017, 8, 944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widderich, N.; Czech, L.; Elling, F.J.; Könneke, M.; Stöveken, N.; Pittelkow, M.; Riclea, R.; Dickschat, J.S.; Heider, J.; Bremer, E. Strangers in the archaeal world: Osmostress-responsive biosynthesis of ectoine and hydroxyectoine by the marine thaumarchaeon Nitrosopumilus maritimus. Environ. Microbiol. 2016, 18, 1227–1248. [Google Scholar] [CrossRef] [PubMed]
- Landa, M.; Burns, A.S.; Roth, S.J.; Moran, M.A. Bacterial transcriptome remodeling during sequential co-culture with a marine dinoflagellate and diatom. ISME J. 2017, 11, 2677–2690. [Google Scholar] [CrossRef] [Green Version]
- Dickson, D.M.J.; Kirst, G.O. Osmotic adjustment in marine eukaryotic algae—The role of inorganic-ions, quaternary ammonium, tertiary sulfonium and carbohydrate solutes. I. Diatoms and a Rhodophyte. New Phytol. 1987, 106, 645–655. [Google Scholar] [CrossRef]
- Kiene, R.P.; Williams, L.P.H.; Walker, J.E. Seawater microorganisms have a high affinity glycine betaine uptake system which also recognizes dimethylsulfoniopropionate. Aquat. Microb. Ecol. 1998, 15, 39–51. [Google Scholar] [CrossRef]
- Van Bergeijk, S.A.; Van der Zee, C.; Stal, L.J. Uptake and excretion of dimethylsulphoniopropionate is driven by salinity changes in the marine benthic diatom Cylindrotheca closterium. Eur. J. Phycol. 2003, 38, 341–349. [Google Scholar] [CrossRef] [Green Version]
- Torstensson, A.; Young, J.N.; Carlson, L.T.; Ingalls, A.E.; Deming, J.W. Use of exogenous glycine betaine and its precursor choline as osmoprotectants in Antarctic sea-ice diatoms. J. Phycol. 2019, 55, 663–675. [Google Scholar] [CrossRef]
- Czech, L.; Bremer, E. With a pinch of extra salt—Did predatory protists steal genes from their food? PLoS Biol. 2018, 16, e2005163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onraedt, A.; De Mey, M.; Walcarius, B.; Soetaert, W.; Vandamme, E.J. Transport kinetics of ectoine, an osmolyte produced by Brevibacterium epidermis. Biotechnol. Lett. 2006, 28, 1741–1747. [Google Scholar] [CrossRef] [PubMed]
- Czech, L.; Hermann, L.; Stoveken, N.; Richter, A.A.; Hoppner, A.; Smits, S.H.J.; Heider, J.; Bremer, E. Role of the extremolytes ectoine and hydroxyectoine as stress protectants and nutrients: Genetics, phylogenomics, biochemistry, and structural analysis. Genes 2018, 9, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grammann, K.; Volke, A.; Kunte, H.J. New type of osmoregulated solute transporter identified in halophilic members of the Bacteria domain: TRAP transporter TeaABC mediates uptake of ectoine and hydroxyectoine in Halomonas elongata DSM 2581T. J. Bacteriol. 2002, 184, 3078–3085. [Google Scholar] [CrossRef] [Green Version]
- Kuhlmann, A.U.; Hoffmann, T.; Bursy, J.; Jebbar, M.; Bremer, E. Ectoine and hydroxyectoine as protectants against osmotic and cold stress: Uptake through the SigB-controlled betaine-choline- carnitine transporter-type carrier EctT from Virgibacillus pantothenticus. J. Bacteriol. 2011, 193, 4699–4708. [Google Scholar] [CrossRef] [Green Version]
- Kiene, R.P.; Linn, L.J.; Bruton, J.A. New and important roles for DMSP in marine microbial communities. J. Sea Res. 2000, 43, 209–224. [Google Scholar] [CrossRef]
- Simó, R. Production of atmospheric sulfur by oceanic plankton: Biogeochemical, ecological and evolutionary links. Trends Ecol. Evolut. 2001, 16, 287–294. [Google Scholar] [CrossRef]
- Maier, I.; Calenberg, M. Effect of extracellular Ca2+ and Ca2+-antagonists on the movement and chemoorientation of male gametes of Ectocarpus siliculosus (Phaeophyceae). Bot. Acta 1994, 107, 451–458. [Google Scholar] [CrossRef]
- Hillebrand, H.; Durselen, C.D.; Kirschtel, D.; Pollingher, U.; Zohary, T. Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 1999, 35, 403–424. [Google Scholar] [CrossRef]
- Himdi-Kabbab, S.; Lavrador, K.; Bazureau, J.P.; Hamelin, J. Synthesis of 1,4,5,6-Tetrahydro 2-Methyl 4-Pyrimidine Carboxylic Acid: Osmoprotector Amino Acid. Synth. Commun. 1995, 25, 2223–2227. [Google Scholar] [CrossRef]
- Shortreed, M.R.; Lamos, S.M.; Frey, B.L.; Phillips, M.F.; Patel, M.; Belshaw, P.J.; Smith, L.M. Ionizable isotopic labeling reagent for relative quantification of amine metabolites by mass spectrometry. Anal. Chem. 2006, 78, 6398–6403. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lenght (µm) | Width (µm) | Cell Volume (µm3) | Standard Deviation | Difference (Δ%) | One Way ANOVA | |
|---|---|---|---|---|---|---|
| T. weissflogii | ||||||
| 35 PSU | 188 | 95.1 | 1.32 × 106 | ±2.59 × 105 | ||
| 50 PSU (24 h) | 227 | 85.7 | 1.23 × 106 | ±1.74 × 105 | −6.82% | n.s. |
| 50 PSU | 207 | 88.9 | 1.29 × 106 | ±3.12 × 105 | −2.27% | n.s. |
| P. tricornutum | ||||||
| 35 PSU | 21.6 | 11.8 | 131 | ±39.1 | ||
| 50 PSU (24 h) | 20.7 | 10.4 | 113 | ±29 | −13.70% | n.s. |
| 50 PSU | 22.4 | 13.6 | 159 | ±33.8 | 21.40% | p < 0.01 |
| Species | GBT | DMSA | Gonyol | n DMSP (fmol per cell) | n DMSOP (fmol per cell) | n Ectoine (fmol per cell) |
|---|---|---|---|---|---|---|
| P. minimum | + | + | + | 304.5 ± 61.2 | 3.66 ± 1.23 | 141.67 ± 13.43 |
| P. parvum | + | - | + | 16.2 ± 4.4 | 0.029 ± 0.005 | 7.85 ± 1.61 |
| S. costatum | + | - | - | 6.56 ± 2.06 | 0.029 ± 0.005 | 5.42 ± 1.89 |
| E. huxleyi | + | - | + | 4.83 + 0.57 | 0.029 ± 0.013 | 2.26 ± 0.45 |
| I. galbana | + | - | + | 4.69 + 0.27 | 0.017 ± 0.003 | 2.62 ± 0.53 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fenizia, S.; Thume, K.; Wirgenings, M.; Pohnert, G. Ectoine from Bacterial and Algal Origin Is a Compatible Solute in Microalgae. Mar. Drugs 2020, 18, 42. https://doi.org/10.3390/md18010042
Fenizia S, Thume K, Wirgenings M, Pohnert G. Ectoine from Bacterial and Algal Origin Is a Compatible Solute in Microalgae. Marine Drugs. 2020; 18(1):42. https://doi.org/10.3390/md18010042
Chicago/Turabian StyleFenizia, Simona, Kathleen Thume, Marino Wirgenings, and Georg Pohnert. 2020. "Ectoine from Bacterial and Algal Origin Is a Compatible Solute in Microalgae" Marine Drugs 18, no. 1: 42. https://doi.org/10.3390/md18010042