Biocatalysis of Fucodian in Undaria pinnatifida Sporophyll Using Bifidobacterium longum RD47 for Production of Prebiotic Fucosylated Oligosaccharide


{kind=link}
{kind=link}
Abstract
:1. Introduction
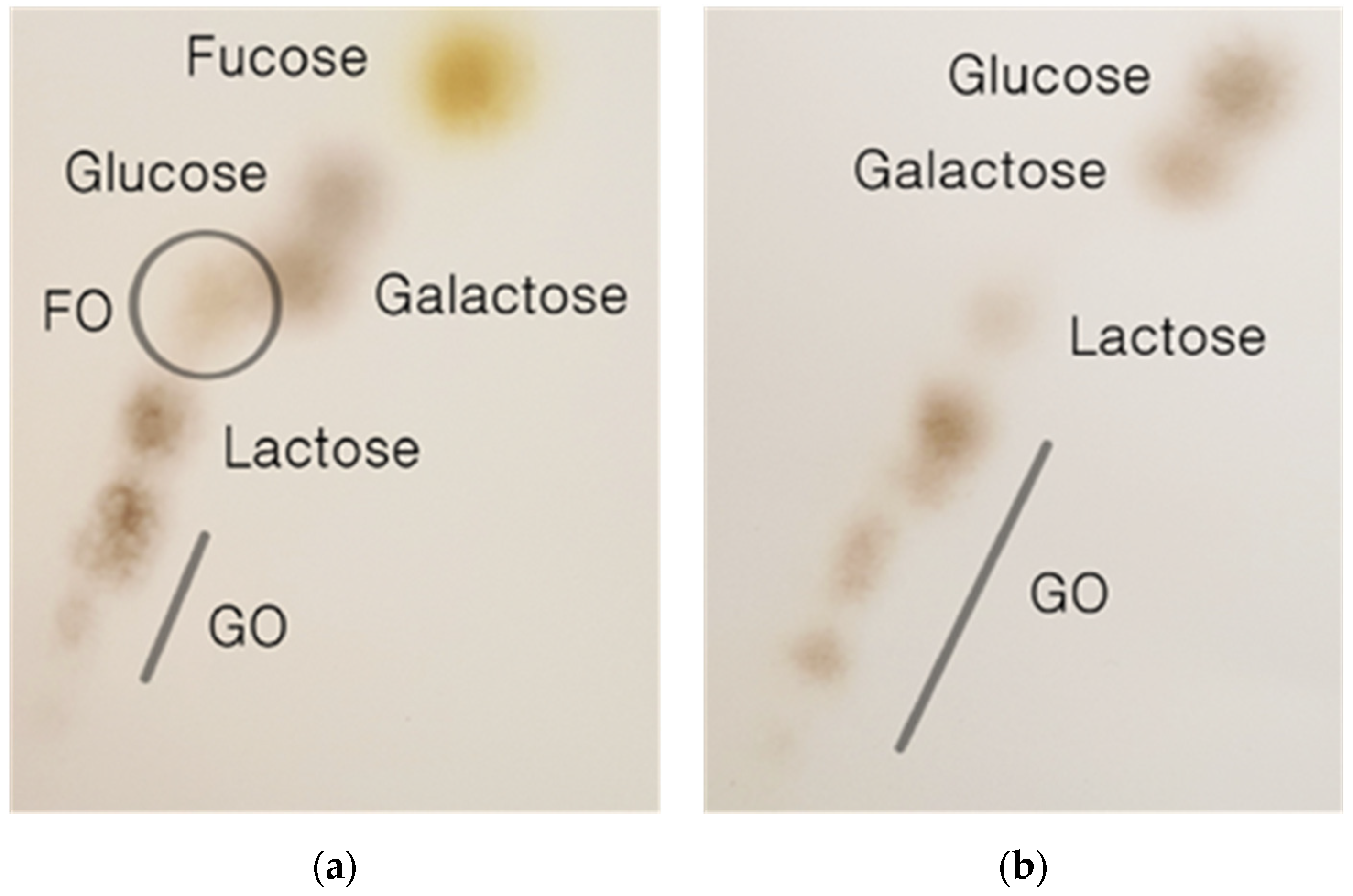
2. Results and Discussion
2.1. Extraction and Purification of Monosaccharides from U. pinnatifida Sporophyll
2.2. Synthesis of Fucosylated Oligosaccharide Using β-Galactosidase of B. longum RD47
3. Materials and Methods
3.1. Extraction of Crude Fucoidan from U. pinnatifida Sporophyll
3.2. Preparation of Purified Fucoidan Hydrolysate from Crude Fucoidan
3.3. Evaluation of Monosaccharide Composition of Fucoidan Hydrolysate by Bio-LC
3.4. Synthesis of Fucosylated Oligosaccharide Using B. longum RD47
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Becker, D.J.; Lowe, J.B. Fucose: Biosynthesis and biological function in mammals. Glycobiology 2003, 13, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef]
- Berteau, O.; Mulloy, B. Sulfated fucans, fresh perspectives: Structures, functions, and biological properties of sulfated fucans and an overview of enzymes active toward this class of polysaccharide. Glycobiology 2003, 13, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Mak, W.; Wang, S.K.; Liu, T.; Hamid, N.; Li, Y.; Lu, J.; White, W.L. Anti-proliferation potential and content of fucoidan extracted from sporophyll of New Zealand Undaria pinnatifida. Front. Nutr. 2014, 1. [Google Scholar] [CrossRef] [PubMed]
- Chizhov, A.O.; Dell, A.; Morris, H.R.; Haslam, S.M.; McDowell, R.A.; Shashkov, A.S.; Nifant’ev, N.E.; Khatuntseva, E.A.; Usov, A.I. A study of fucoidan from the brown seaweed Chorda filum. Carbohydr. Res. 1999, 320, 108–119. [Google Scholar] [CrossRef]
- Sakai, T.; Ishizuka, K.; Shimanaka, K.; Ikai, K.; Kato, I. Structures of oligosaccharides derived from Cladosiphon okamuranus fucoidan by digestion with marine bacterial enzymes. Mar. Biotechnol. 2003, 5, 536–544. [Google Scholar] [PubMed]
- Bilan, M.I.; Grachev, A.A.; Ustuzhanina, N.E.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. A highly regular fraction of a fucoidan from the brown seaweed Fucus distichus L. Carbohydr. Res. 2004, 339, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Nishino, T.; Nishioka, C.; Ura, H.; Nagumo, T. Isolation and partial characterization of a noval amino sugar-containing fucan sulfate from commercial Fucus vesiculosus fucoidan. Carbohydr. Res. 1994, 255, 213–224. [Google Scholar] [CrossRef]
- Synytsya, A.; Kim, W.J.; Kim, S.M.; Pohl, R.; Synytsya, A.; Kvasnička, F.; Čopíková, J.; Il Park, Y. Structure and antitumour activity of fucoidan isolated from sporophyll of Korean brown seaweed Undaria pinnatifida. Carbohydr. Polym. 2010, 81, 41–48. [Google Scholar] [CrossRef]
- Kim, W.J.; Park, J.W.; Park, J.K.; Choi, D.J.; II Park, Y. Purification and characterization of a fucoidanase (FNase S) from a marine bacterium Sphingomonas paucimobilis PF-1. Mar. Drugs 2015, 13, 4398–4417. [Google Scholar] [CrossRef]
- Blank, D.; Dotz, V.; Geyer, R.; Kunz, C. Human milk oligosaccharides and lewis blood group: Individual high-throughput sample profiling to enhance conclusions from functional studies. Adv. Nutr. 2012, 3, 440S–449S. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zheng, Y.; Wang, J.; Ma, S.; Yu, Y.; White, W.L.; Yang, S.; Yang, F.; Lu, J. Fucoidan extracted from Undaria pinnatifida: Source for nutraceuticals/functional foods. Mar. Drugs 2018, 16, 321. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, P.; Warren, C.D.; Altaye, M.; Morrow, A.L.; Ruiz-Palacios, G.; Pickering, L.K.; Newburg, D.S. Fucosylated human milk oligosaccharides vary between individuals and over the course of lactation. Glycobiology 2001, 11, 365–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.T.; Chen, C.; Kling, D.E.; Liu, B.; McCoy, J.M.; Merighi, M.; Heidtman, M.; Newburg, D.S. The principal fucosylated oligosaccharides of human milk exhibit prebiotic properties on cultured infant microbiota. Glycobiology 2012, 23, 169–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bode, L. Human milk oligosaccharides: Prebiotics and beyond. Nutr. Rev. 2009, 67, S183–S191. [Google Scholar] [CrossRef] [PubMed]
- Sampedro, J.; Gianzo, C.; Iglesias, N.; Guitian, E.; Revilla, G.; Zarra, I. Atbgal10 is the main xyloglucan β-galactosidase in arabidopsis, and its absence results in unusual xyloglucan subunits and growth defects. Plant Physiol. 2012, 158, 1146–1157. [Google Scholar] [CrossRef] [PubMed]
- Bakunina, I.; Slepchenko, L.; Anastyuk, S.; Isakov, V.; Likhatskaya, G.; Kim, N.; Tekutyeva, L.; Son, O.; Balabanova, L. Characterization of properties and transglycosylation abilities of recombinant α-galactosidase from cold-adapted marine bacterium pseudoalteromonas KMM 701 and its C494N and D451A mutants. Mar. Drugs 2018, 16, 349. [Google Scholar] [CrossRef]
- Hwang, J. Synthesis of fucosylated oligosaccharides using crude enzyme extracts from Bifidobacterium longum Subsp. longum RD 47. Master’s Thesis, Seoul National University, Seoul, South Korea, February 2014. [Google Scholar]
- Lane, J. ZuChem Picks up SBIR Grant for Fermentative Production Methods for l-Fucose: Biofuels Digest. Available online: http://www.biofuelsdigest.com/bdigest/2016/03/20/zuchem-picks-up-sbir-grant-for-fermentative-production-methods-for-l-fucose/ (accessed on 12 December 2018).
- Nayar, S.; Bott, K. Current Status of Global Cultivated Seaweed Production and Markets. World Aquacult. 2014, 45, 32–37. [Google Scholar]
- Kim, W.J.; Kim, S.M.; Kim, H.G.; Oh, H.R.; Lee, K.B.; Lee, Y.K.; II Park, Y. Purification and anticoagulant activity of a fucoidan from Korean Undaria pinnatifida sporophyll. Algae 2007, 22, 247–252. [Google Scholar] [CrossRef]
- Skriptsova, A.V.; Shevchenko, N.M.; Zvyagintseva, T.N.; Imbs, T.I. Monthly changes in the content and monosaccharide composition of fucoidan from Undaria pinnatifida (Laminariales, Phaeophyta). J. Appl. Phycol. 2009, 22, 79–86. [Google Scholar] [CrossRef]
- Chin, Y.W.; Seo, N.; Kim, J.H.; Seo, J.H. Metabolic engineering of Escherichia coli to produce 2′-fucosyllactose via salvage pathway of guanosine 5′-diphosphate (GDP)-l-fucose. Biotechnol. Bioeng. 2016, 113, 2443–2452. [Google Scholar] [CrossRef] [PubMed]
- Intanon, M.; Arreola, S.L.; Pham, N.H.; Kneifel, W.; Haltrich, D.; Nguyen, T.H. Nature and biosynthesis of galacto-oligosaccharides related to oligosaccharides in human breast milk. FEMS Microbiol. Lett. 2014, 353, 89–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.H.; Pathanibul, P.; Quarterman, J.; Jo, J.H.; Han, N.S.; Miller, M.J.; Jin, Y.S.; Seo, J.H. Whole cell biosynthesis of a functional oligosaccharide, 2′-fucosyllactose, using engineered Escherichia coli. Microb. Cell Fact. 2012, 11, 48. [Google Scholar] [CrossRef] [PubMed]
- New York Times. Stop Bashing G.M.O. Foods, More Than 100 Nobel Laureates Say. Available online: https://www.nytimes.com/2016/07/01/us/stop-bashing-gmo-foods-more-than-100-nobel-laureates-say.html?action=click&module=RelatedCoverage&pgtype=Article®ion=Footer (accessed on 14 December 2018).
- Roberts, R.J. The Nobel Laureates’ Campaign Supporting GMOs. J. Innov. Knowl. 2018, 3, 61–65. [Google Scholar] [CrossRef]
- Li, Y.; Ku, S.; Park, M.S.; Li, Z.; Ji, G.E. Acceleration of aglycone isoflavone and aminobutyric acid production from doenjang using whole-cell biocatalysis accompanied by protease treatment. J. Microbiol. Biotechnol. 2017, 27, 1952–1960. [Google Scholar] [CrossRef] [PubMed]
- Ku, S. Finding and producing probiotic glycosylases for the biocatalysis of ginsenosides: A mini review. Molecules 2016, 21, 645. [Google Scholar] [CrossRef] [PubMed]
- Lucht, J.M. Public acceptance of plant biotechnology and GM crops. Viruses 2015, 7, 4254–4281. [Google Scholar] [CrossRef]
- Wang, J.; Wang, F.; Yun, H.; Zhang, H.; Zhang, Q. Effect and mechanism of fucoidan derivatives from Laminaria japonica in experimental adenine-induced chronic kidney disease. J. Ethnopharmacol. 2012, 139, 807–813. [Google Scholar] [CrossRef]
- Ahn, H.J.; You, H.J.; Park, M.S.; Johnston, T.V.; Ku, S.; Ji, G.E. Biocatalysis of platycoside E and platycodin D3 using fungal extracellular Β-glucosidase responsible for rapid platycodin D production. Int. J. Mol. Sci. 2018, 19, 2671. [Google Scholar] [CrossRef]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.M.; Oh, S.Y.; Johnston, T.V.; Ku, S.; Ji, G.E. Biocatalysis of Fucodian in Undaria pinnatifida Sporophyll Using Bifidobacterium longum RD47 for Production of Prebiotic Fucosylated Oligosaccharide. Mar. Drugs 2019, 17, 117. https://doi.org/10.3390/md17020117
Lee JM, Oh SY, Johnston TV, Ku S, Ji GE. Biocatalysis of Fucodian in Undaria pinnatifida Sporophyll Using Bifidobacterium longum RD47 for Production of Prebiotic Fucosylated Oligosaccharide. Marine Drugs. 2019; 17(2):117. https://doi.org/10.3390/md17020117
Chicago/Turabian StyleLee, Jeong Min, So Young Oh, Tony V. Johnston, Seockmo Ku, and Geun Eog Ji. 2019. "Biocatalysis of Fucodian in Undaria pinnatifida Sporophyll Using Bifidobacterium longum RD47 for Production of Prebiotic Fucosylated Oligosaccharide" Marine Drugs 17, no. 2: 117. https://doi.org/10.3390/md17020117