Sargassum Seaweed as a Source of Anti-Inflammatory Substances and the Potential Insight of the Tropical Species: A Review

 ,
,
Abstract
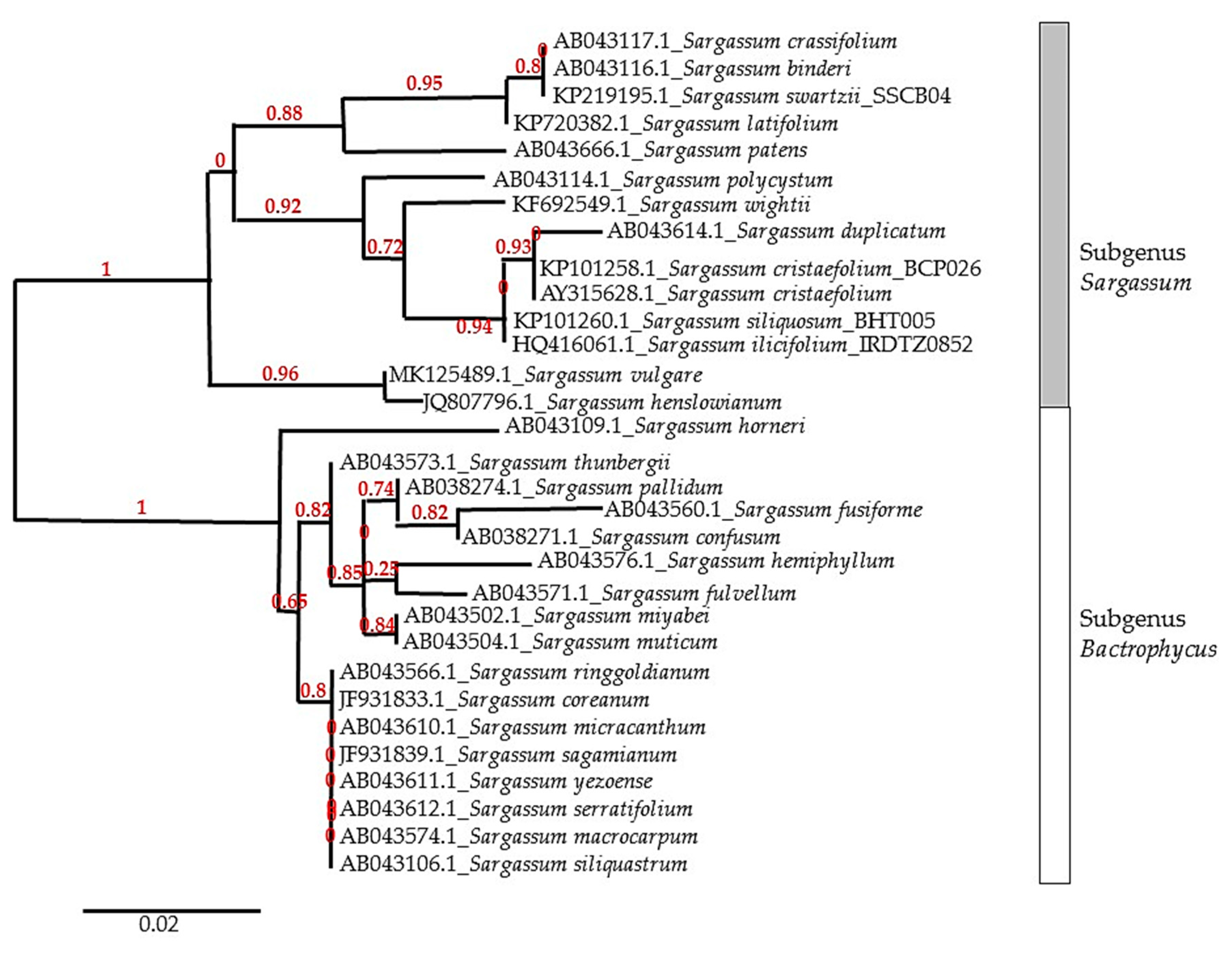
:1. Introduction
2. Sargassum Species Are a Source of Anti-Inflammatory Agents
2.1. Crude Extracts and Their Partitions
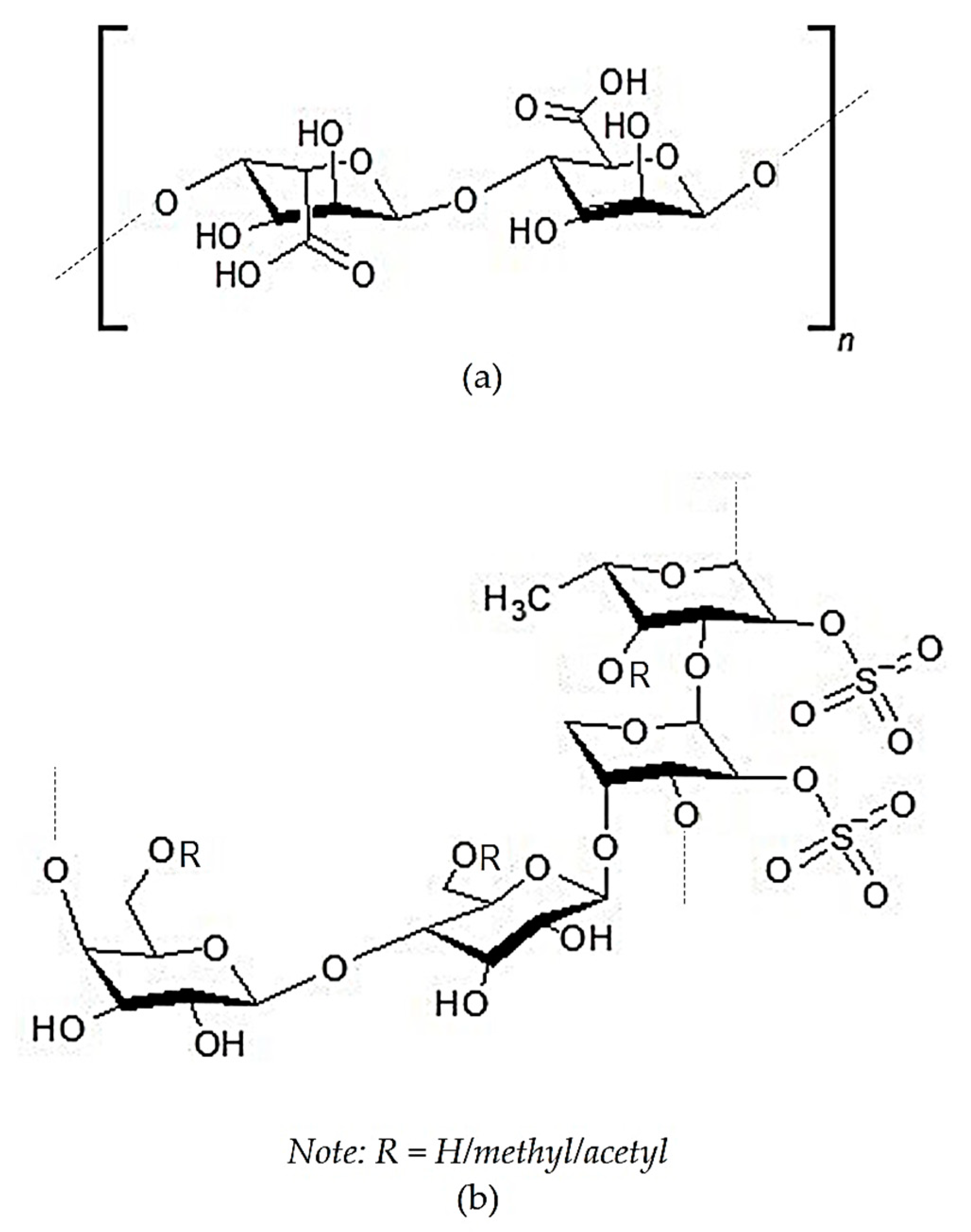
2.2. Crude Sulfated Polysaccharides
2.3. Purified Bioactive Compounds
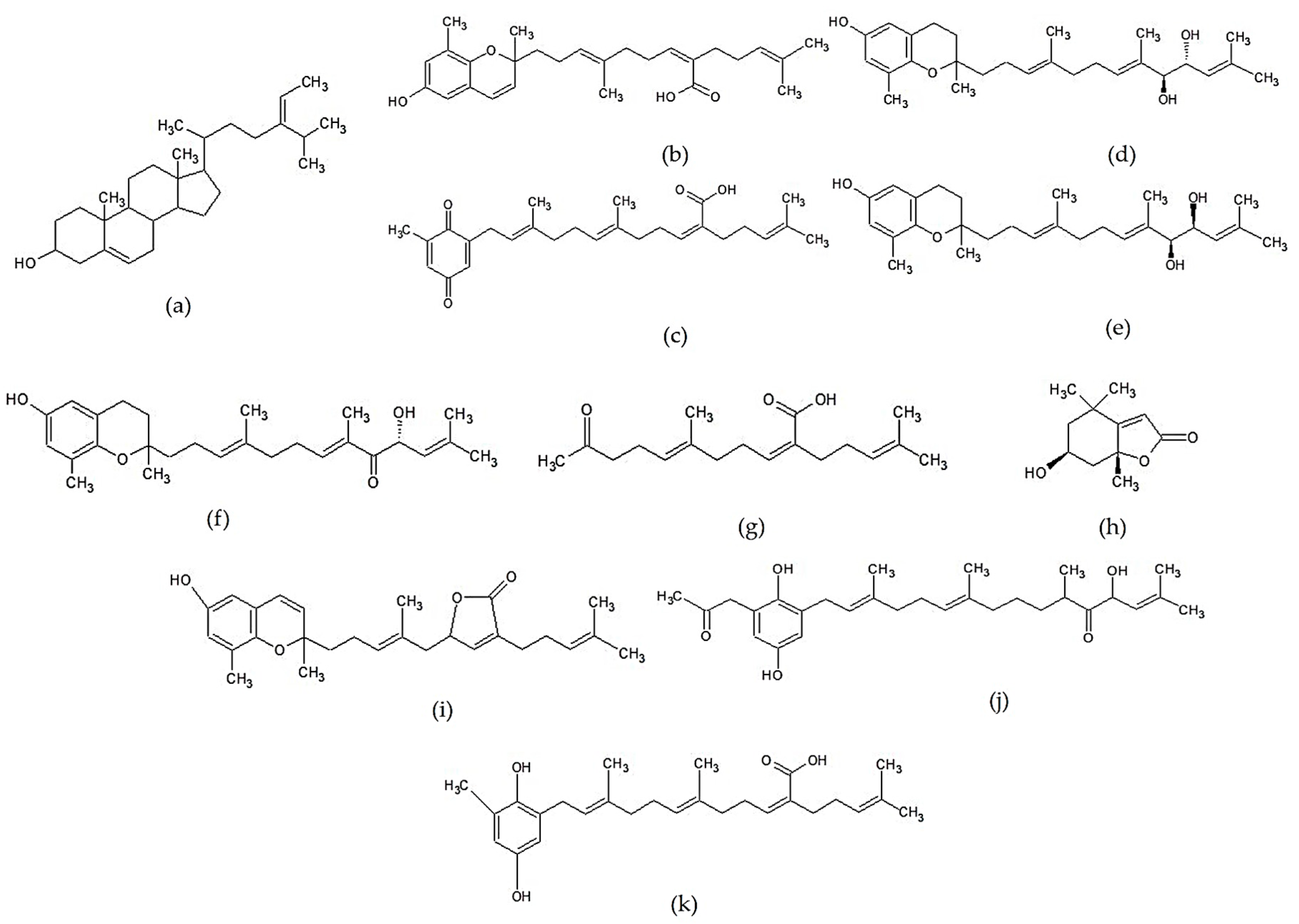
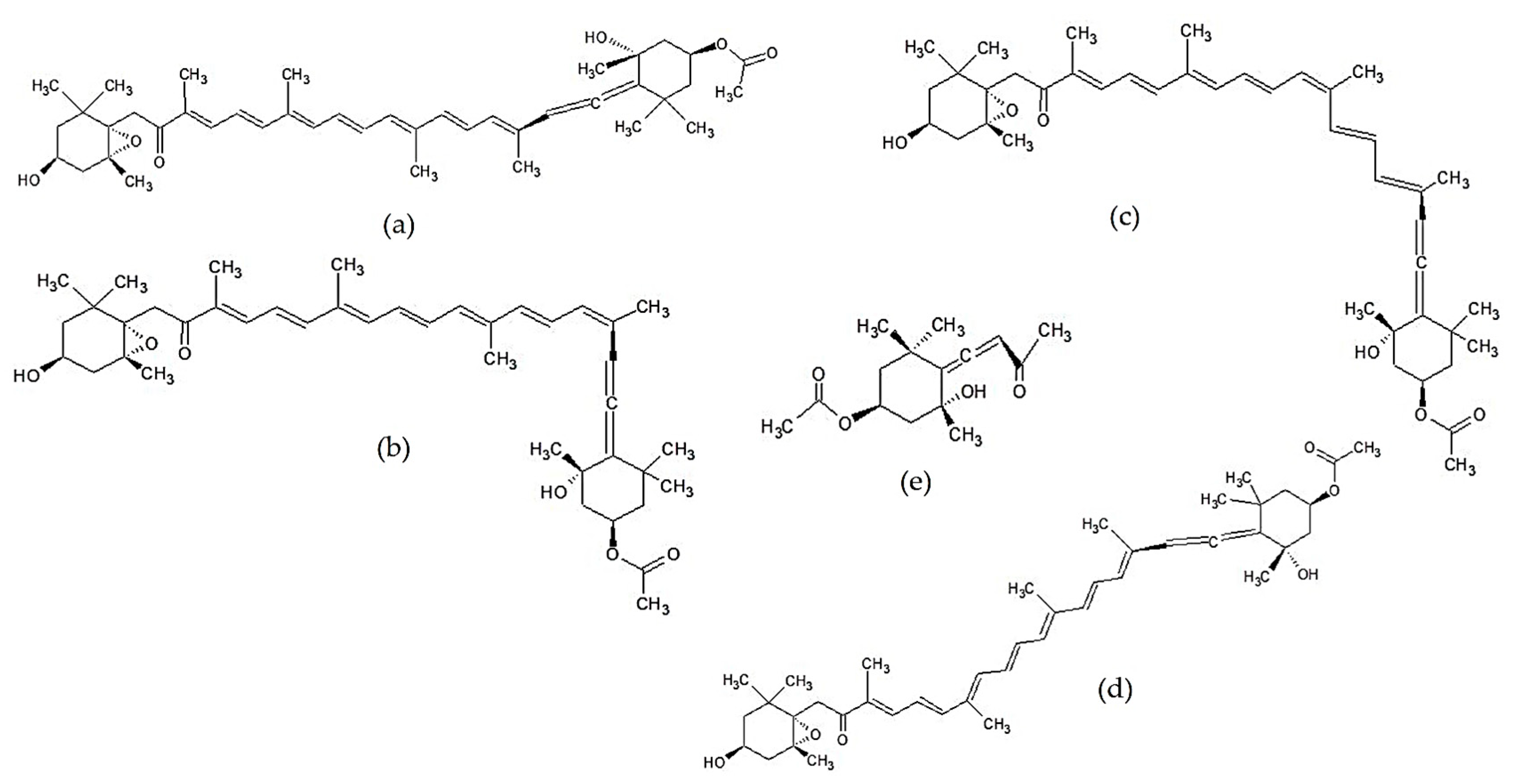
2.3.1. Terpenoids
2.3.2. Fucoxanthin and Its Derivatives
2.3.3. Other Lipid-Soluble Compounds
2.3.4. Polysaccharides
2.3.5. Phenolic Compounds
3. The Potency of Tropical Sargassum as an Anti-Inflammatory Agent
3.1. Latest Reports on Anti-Inflammatory Activity of Tropical Sargassum
3.2. Potential Anti-Inflammatory Compounds of Tropical Sargassum
3.2.1. Lipid-Soluble Bioactive Compounds
3.2.2. Water-Soluble Bioactive Compounds
4. Anti-Inflammatory Mechanisms of Bioactive Compounds of Sargassum
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Ahmed, A.U. An overview of inflammation: Mechanism and consequences. Front. Biol. China 2011, 6, 274–281. [Google Scholar]
- Kotas, M.E.; Medzhitov, R. Homeostasis, inflammation, and disease susceptibility. Cell 2015, 160, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Fernando, I.P.S.; Nah, J.-W.; Jeon, Y.-J. Potential anti-inflammatory natural products from marine algae. Environ. Toxicol. Pharmacol. 2016, 48, 22–30. [Google Scholar] [CrossRef]
- Newton, K.; Dixit, V.M. Signaling in innate immunity and inflammation. Cold Spring Harb. Perspect. Biol. 2012, 4. [Google Scholar] [CrossRef]
- Dray, A. Inflammatory mediators of pain. Br. J. Anaesth. 1995, 75, 125–131. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef]
- Fitton, J.H. Brown marine algae: A survey of therapeutic potentials. Altern. Complement. Ther. 2003, 9, 29–33. [Google Scholar] [CrossRef]
- Oh, J.-H.; Kim, J.; Lee, Y. Anti-inflammatory and anti-diabetic effects of brown seaweeds in high-fat diet-induced obese mice. Nutr. Res. Pract. 2016, 10, 42. [Google Scholar] [CrossRef]
- Husni, A.; Lailatussifa, R.; Isnansetyo, A. Sargassum hystrix as a source of functional food to improve blood biochemistry profiles of rats under stress. Prev. Nutr. Food Sci. 2019, 24, 150–158. [Google Scholar]
- Vo, T.-S.; Ngo, D.-H.; Kim, S.-K. Potential targets for anti-inflammatory and anti-allergic activities of marine algae: An overview. Inflamm. Allergy-Drug Targets 2012, 11, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Heinrich, M.; Myers, S.; Dworjanyn, S.A. Towards a better understanding of medicinal uses of the brown seaweed Sargassum in Traditional Chinese Medicine: A phytochemical and pharmacological review. J. Ethnopharmacol. 2012, 142, 591–619. [Google Scholar] [CrossRef] [PubMed]
- Jaswir, I.; Monsur, H.A. Anti-inflammatory compounds of macro algae origin: A review. J. Med. Plants Res. 2011, 5, 7146–7154. [Google Scholar]
- Susanto, E.; Fahmi, A.S.; Hosokawa, M.; Abe, M.; Miyashita, K. Lipids, fatty acids, and fucoxanthin content from temperate and tropical brown seaweeds. Aquat. Procedia 2016, 7, 66–75. [Google Scholar] [CrossRef]
- Miyashita, K.; Mikami, N.; Hosokawa, M. Chemical and nutritional characteristics of brown seaweed lipids: A review. J. Funct. Foods 2013, 5, 1507–1517. [Google Scholar] [CrossRef]
- Mattio, L.; Payri, C. Taxonomic revision of Sargassum species (Fucales, Phaeophyceae) from New Caledonia based on morphological and molecular. J. Phycol. 2009, 45, 374–1388. [Google Scholar] [CrossRef]
- Kantachumpoo, A.; Uwai, S.; Noiraksar, T.; Komatsu, T. Systematics of marine brown alga Sargassum from Thailand: A preliminary study based on morphological data and nuclear ribosomal internal transcribed spacer 2 (ITS2) sequences. Ocean Sci. J. 2015, 50, 251–262. [Google Scholar] [CrossRef]
- Mun, O.J.; Kwon, M.S.; Karadeniz, F.; Kim, M.; Lee, S.H.; Kim, Y.Y.; Seo, Y.; Jang, M.S.; Nam, K.H.; Kong, C.S. Fermentation of Sargassum thunbergii by Kimchi-Derived Lactobacillus sp. SH-1 Attenuates LPS-Stimulated Inflammatory Response Via Downregulation of JNK. J. Food Biochem. 2017, 41, 1–9. [Google Scholar] [CrossRef]
- Jayawardena, T.U.; Kim, H.S.; Sanjeewa, K.K.A.; Kim, S.Y.; Rho, J.R.; Jee, Y.; Ahn, G.; Jeon, Y.J. Sargassum horneri and isolated 6-hydroxy-4,4,7a-trimethyl-5,6,7,7a-tetrahydrobenzofuran-2(4H)-one (HTT); LPS-induced inflammation attenuation via suppressing NF-κB, MAPK and oxidative stress through Nrf2/HO-1 pathways in RAW 264.7 macrophages. Algal Res. 2019, 40, 101513. [Google Scholar] [CrossRef]
- Gwon, W.G.; Lee, S.G.; Kim, J., II; Kim, Y.M.; Kim, S.B.; Kim, H.R. Hexane fraction from the ethanolic extract of Sargassum serratifolium suppresses cell adhesion molecules via regulation of NF-κB and Nrf2 pathway in human umbilical vein endothelial cells 06 Biological Sciences 0601 Biochemistry and Cell Biology. Fish. Aquat. Sci. 2019, 22, 1–10. [Google Scholar]
- Yoon, W.J.; Ham, Y.M.; Lee, W.J.; Lee, N.H.; Hyun, C.G. Brown alga Sargassum muticum inhibits proinflammatory cytokines, iNOS, and COX-2 expression in macrophage RAW 264.7 cells. Turk. J. Biol. 2010, 34, 25–34. [Google Scholar]
- Kang, J.Y.; Khan, M.N.A.; Park, N.H.; Cho, J.Y.; Lee, M.C.; Fujii, H.; Hong, Y.K. Antipyretic, analgesic, and anti-inflammatory activities of the seaweed Sargassum fulvellum and Sargassum thunbergii in mice. J. Ethnopharmacol. 2008, 116, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Dar, A.; Baig, H.S.; Saifullah, S.M.; Ahmad, V.U.; Yasmeen, S.; Nizamuddin, M. Effect of seasonal variation on the anti-inflammatory activity of Sargassum wightii growing on the N. Arabian Sea coast of Pakistan. J. Exp. Mar. Bio. Ecol. 2007, 351, 1–9. [Google Scholar] [CrossRef]
- Lee, C.; Park, G.H.; Ahn, E.M.; Park, C.-I.; Jang, J.-H. Sargassum fulvellum protects HaCaT cells and BALB/c mice from UVB-induced proinflammatory responses. Evidence-Based Complement. Altern. Med. 2013, 2013, 1–10. [Google Scholar]
- Kim, M.-J.; Kim, K.-B.-W.-R.; Jeong, D.-H.; Ahn, D.-H. Inhibitory effect of Sargauum fulvellum ethanolic extract on LPS-induced inflammatory reaction in RAW 264.7 mouse macrophages. J. Appl. Biol. Chem. 2013, 56, 249–255. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Jayawardena, T.U.; Kim, H.S.; Lee, W.W.; Vaas, A.P.J.P.; De Silva, H.I.C.; Abayaweera, G.S.; Nanayakkara, C.M.; Abeytunga, D.T.U.; Lee, D.S.; et al. Beijing urban particulate matter-induced injury and inflammation in human lung epithelial cells and the protective effects of fucosterol from Sargassum binderi (Sonder ex J. Agardh). Environ. Res. 2019, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.; Yoon, W.J.; Ham, Y.M.; Yoon, S.A.; Kang, S.C. Anti-arthritis effect through the anti-inflammatory effect of Sargassum muticum extract in collagen-induced arthritic (CIA) Mice. Molecules 2019, 24, 276. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.; Lim, S.J.; Joung, E.J.; Lee, B.; Oh, C.W.; Kim, H.R. Meroterpenoid-rich fraction of an ethanolic extract from Sargassum serratifolium alleviates obesity and non-alcoholic fatty liver disease in high fat-fed C57BL/6J mice. J. Funct. Foods 2018, 47, 288–298. [Google Scholar] [CrossRef]
- Gwon, W.G.; Joung, E.J.; Shin, T.; Utsuki, T.; Wakamatsu, N.; Kim, H.R. Meroterpinoid-rich fraction of the ethanol extract from Sargassum serratifolium suppresses TNF-α-induced monocytes adhesion to vascular endothelium and vascular inflammation in high cholesterol-fed C57BL/6J mice. J. Funct. Foods 2018, 46, 384–393. [Google Scholar] [CrossRef]
- Karadeniz, F.; Lee, S.G.; Oh, J.H.; Kim, J.A.; Kong, C.S. Inhibition of MMP-2 and MMP-9 activities by solvent-partitioned Sargassum horneri extracts. Fish. Aquat. Sci. 2018, 21, 1–7. [Google Scholar] [CrossRef]
- Sugiura, Y.; Kinoshita, Y.; Abe, M.; Murase, N.; Tanaka, R.; Matsushita, T.; Usui, M.; Hanaoka, K.; Miyata, M. Suppressive effects of the diethyl ether fraction from a brown alga Sargassum fusiforme on allergic and inflammatory reactions. Fish. Sci. 2016, 82, 369–377. [Google Scholar] [CrossRef]
- Nomura, M.; Kamogawa, H.; Susanto, E.; Kawagoe, C.; Yasui, H.; Saga, N.; Hosokawa, M.; Miyashita, K. Seasonal variations of total lipids, fatty acid composition, and fucoxanthin contents of Sargassum horneri (Turner) and Cystoseira hakodatensis (Yendo) from the northern seashore of Japan. J. Appl. Phycol. 2013, 25, 1159–1169. [Google Scholar] [CrossRef]
- Han, Y.R.; Ali, M.Y.; Woo, M.H.; Jung, H.A.; Choi, J.S. Anti-Diabetic and Anti-Inflammatory Potential of the Edible Brown Alga Hizikia Fusiformis. J. Food Biochem. 2015, 39, 417–428. [Google Scholar] [CrossRef]
- Moni, S.S.; Alam, M.F.; Makeen, H.A.; Jabeen, A.; Sanobar, S.; Siddiqui, R.; Moochikkal, R.; Fouda, S. Therapeutic potential of oleic acid nanovesicles prepared from petroleum ether extract of Sargassum binderi in streptozotocin–induced diabetic wound in wistar rats. Trop. J. Pharm. Res. 2018, 17, 2123–2128. [Google Scholar] [CrossRef]
- Jaswir, I.; Monsur, H.A.; Simsek, S.; Amid, A.; Alam, Z.; Salleh, M.N.b.; Tawakalit, A.-H.; Octavianti, F. Cytotoxicity and inhibition of nitric oxide in lipopolysaccharide-induced mammalian cell lines by aqueous extracts of brown seaweed. J. Oleo Sci. 2014, 63, 787–794. [Google Scholar] [CrossRef]
- Monsur, A.H.; Jaswir, I.; Simsek, S.; Amid, A.; Alam, Z.; Tawakalit, A.-H. Cytotoxicity and inhibition of nitric oxide syntheses in LPS induced macrophage by water soluble fractions of brown seaweed. Food Hydrocoll. 2014, 42, 269–274. [Google Scholar] [CrossRef]
- Kim, M.; Kim, K.; Park, S.; Choi, H.; Park, S.; Kim, J.; Jang, M.; Im, M.; Ahn, D. Anti-Inflammatory effect of Sargassum patens C. Agardh ethanol extract in LPS-induced RAW264.7 cells and mouse ear edema. Korean J. Microbiol. Biotechnol. 2017, 45, 110–117. [Google Scholar] [CrossRef]
- Kim, K.-N.; Kim, J.; Yoon, W.-J.; Yang, H.-M.; Heo, S.Y.; Ko, J.-Y.; Woon Roh, S.; Jeon, Y.-J.; Kang, S.-M.; Heo, S.-J.; et al. Inhibitory effect of Sargassum patens on inflammation and melanogenesis. Int. J. Pharmacol. 2013, 9, 524–532. [Google Scholar] [CrossRef]
- Maneesh, A.; Chakraborty, K.; Makkar, F. Pharmacological activities of brown seaweed Sargassum wightii (Family Sargassaceae) using different in vitro models. Int. J. Food Prop. 2017, 20, 931–945. [Google Scholar] [CrossRef]
- Fazeela, M.B.S.; Hemalatha, S. Characterisation, in silico and in vitro determination of antidiabetic and anti inflammatory potential of ethanolic extract of Sargassum wightii. Asian J. Pharm. Clin. Res. 2017, 10, 297–301. [Google Scholar]
- Raman, R.; Parthiban, S.; Karthikeyan, S.; Muthuraman, M.S.; Sivasubramanian, A. Antimicrobial and anti-inflammatory studies on Sargassum wightii extracts. Int. J. Pharm. Pharm. Sci. 2014, 6, 614. [Google Scholar]
- Radhika, D.; Veerabahu, C.; Priya, R. Anti-inflammatory activities of some seaweed collected from The Gulf of Mannar Coast, Tuticorin, South India. Int. J. Pharma Bio Sci. 2013, 4, 39–44. [Google Scholar]
- Pramitha, V.S.; Kumari, N.S. Anti-inflammatory, anti-oxidant, phytochemical, and GC-MS analysis of marine brown macroalga, Sargassum wightii. Int. J. Pharm. Chem. Biol. Sci. 2016, 6, 7–15. [Google Scholar]
- Dhas, T.S.; Kumar, V.G.; Karthick, V.; Vasanth, K.; Singaravelu, G.; Govindaraju, K. Effect of biosynthesized gold nanoparticles by Sargassum swartzii in alloxan induced diabetic rats. Enzyme Microb. Technol. 2016, 95, 100–106. [Google Scholar] [CrossRef]
- Hong, D.D.; Hien, H.M.; Anh, H.T.L. Studies on the analgesic and anti-inflammatory activities of Sargassum swartzii (Turner) C. Agardh (Phaeophyta) and Ulva reticulata Forsskal (Chlorophyta) in experiment animal models. African J. Biotechnol. 2011, 10, 2308–2313. [Google Scholar]
- Khouzani, M.R.M.; Shushizadeh, M.R.; Kalantari, H.; Ghotrami, E.R. Hepatoprotective effect of aqueous extract of Persian Gulf brown algae Sargassum swartzii against acetaminophen-induced hepatotoxicity in mice. Jundishapur J. Nat. Pharm. Prod. 2019, 14, e77168. [Google Scholar]
- Vairappan, C.S.; Kamada, T.; Lee, W.W.; Jeon, Y.-J. Anti-inflammatory activity of halogenated secondary metabolites of Laurencia snackeyi (Weber-van Bosse) Masuda. J. Appl. Phycol. 2013. [Google Scholar] [CrossRef]
- Chamidah, A.; Prihanto, A.A.; Hardoko, H. Edema-reducing activity of Sargassum crassifolium B-glucan (laminaran) on edema-induced Rattus norvegicus. Asian J. Pharm. Clin. Res. 2017, 10, 311. [Google Scholar] [CrossRef]
- Simpi, C.; Nagathan, C.; Karajgi, S.; Kalyane, N. Evaluation of marine brown algae Sargassum ilicifolium extract for analgesic and anti-inflammatory activity. Pharmacognosy Res. 2013, 5, 146. [Google Scholar] [CrossRef]
- Ahmed, H.H.; Abdalla, M.S.; Eskander, E.F.; Al-Khadragy, M.F.; Massoud, M.N. Hypolipidemic influence of Sargassum subrepandum: Mechanism of action. Eur. Rev. Med. Pharmacol. Sci. 2012, 16, 112–120. [Google Scholar]
- Buwono, N.R.; Risjani, Y.; Arsad, S. Anti-inflammatory and analgesic activity from brown algae Sargassum polycystum. J. Pharm. Sci. Res. 2018, 10, 2092–2096. [Google Scholar]
- Raghavendran, H.B.; Sathivel, A.; Devaki, T. Defensive nature of Sargassum polycystum (Brown alga) against acetaminophen-induced toxic hepatitis in rats: Role of drug metabolizing microsomal enzyme system, tumor necrosis factor- α and fate of liver cell structural integrity. World J. Gastroenterol. 2006, 12, 3829–3834. [Google Scholar] [CrossRef] [PubMed]
- Motshakeri, M.; Ebrahimi, M.; Goh, Y.M.; Othman, H.H.; Hair-bejo, M.; Mohamed, S. Effects of brown seaweed (Sargassum polycystum) extracts on kidney, liver, and pancreas of type 2 diabetic rat model. Evid.-Based Complement. Altern. Med. 2014, 2014, 379407. [Google Scholar] [CrossRef]
- Gany, S.A.; Tan, S.C.; Gan, S.Y. Anti-neuroinflammatory properties of Malaysian brown and green seaweeds. Int. J. Ind. Manuf. Eng. 2014, 8, 1269–1275. [Google Scholar]
- Chatter, R.; Tarhouni, S.; Kharrat, R. Screening of anti-inflammatory and analgesic activities in marines macroalgae from Mediterranean Sea. Arch. Inst. Pasteur Tunis 2011, 88, 19–28. [Google Scholar]
- Shreadah, M.A.; Abd, N.M.; Moneam, E. Phytochemical and pharmacological screening of Sargassium vulgare from Suez Canal, Egypt. Food Sci. Biotechnol. 2018, 27, 963–979. [Google Scholar] [CrossRef]
- Yang, E.-J.; Moon, J.-Y.; Kim, M.-J.; Kim, D.S.; Kim, C.-S.; Lee, W.J.; Lee, N.H.; Hyun, C.-G. Inhibitory effect of Jeju endemic seaweeds on the production of pro-inflammatory mediators in mouse macrophage cell line RAW 264.7. J. Zhejiang Univ. Sci. B 2010, 11, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Kang, H. Neuroprotective Effect of Sargassum thunbergii (Mertens ex Roth) Kuntze in Activated Murine Microglial Cells. Trop. J. Pharm. Res. 2015, 14, 235. [Google Scholar]
- Kim, J.; Karadeniz, F.; Ahn, B.; Kwon, M.S.; Mun, O.; Kim, M.; Lee, S.; Yu, K.H.; Kim, Y.Y.; Kong, C. Sargassum sp. attenuates oxidative stress and suppresses lipid accumulation in vitro. J. Life Sci. 2014, 24, 274–283. [Google Scholar] [CrossRef]
- Kim, J.; Kong, C.; Seo, Y.; Kim, S. Sargassum thunbergii extract inhibits MMP-2 and -9 expressions related with ROS scavenging in HT1080 cells. Food Chem. 2010, 120, 418–425. [Google Scholar] [CrossRef]
- Khan, M.N.; Choi, J.; Lee, M.C.; Kim, E.; Nam, T.; Fujii, H.; Hong, Y.K. Anti-inflammatory activities of methanol extracts from various seaweed species. J. Environ. Biol. 2008, 29, 465–469. [Google Scholar]
- Kim, J.; Kong, C.; Kim, S. Effect of Sargassum thunbergii on ROS mediated oxidative damage and identification of polyunsaturated fatty acid components. Food Chem. Toxicol. 2010, 48, 1243–1249. [Google Scholar] [CrossRef] [PubMed]
- Gwon, W.-G.; Lee, M.-S.; Kim, J.-S.; Kim, J.-I.; Lim, C.-W.; Kim, N.-G.; Kim, H.-R. Hexane fraction from Sargassum fulvellum inhibits lipopolysaccharide-induced inducible nitric oxide synthase expression in RAW 264.7 cells via NF-κB pathways. Am. J. Chin. Med. 2013, 41, 565–584. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.-H.; Kim, K.-B.-W.-R.; Kim, M.-J.; Kang, B.-K.; Bark, S.-W.; Pak, W.-M.; Kim, B.-R.; Ahn, N.-K.; Choi, Y.-U.; Ahn, D.-H. Anti-inflammatory effect of ethanol extract from Sargassum fulvellum on lipopolysaccharide induced inflammatory responses in RAW 264.7 cells and mice ears. J. Korean Soc. Food Sci. Nutr. 2014, 43, 1158–1165. [Google Scholar] [CrossRef]
- Kang, B.; Kim, M.; Kim, K.; Ahn, D. In vivo and in vitro inhibitory activity of an ethanolic extract of Sargassum fulvellum and its component grasshopper ketone on atopic dermatitis. Int. Immunopharmacol. 2016, 40, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Joung, E.-J.; Gwon, W.-G.; Shin, T.; Jung, B.-M.; Choi, J.; Kim, H.-R. Anti-inflammatory action of the ethanolic extract from Sargassum serratifolium on lipopolysaccharide-stimulated mouse peritoneal macrophages and identification of active components. J. Appl. Phycol. 2017, 29, 563–573. [Google Scholar] [CrossRef]
- Oh, S.-J.; Joung, E.-J.; Kwon, M.-S.; Lee, B.; Utsuki, T.; Oh, C.-W.; Kim, H.-R. Anti-Inflammatory Effect of Ethanolic Extract of Sargassum serratifolium in Lipopolysaccharide-Stimulated BV2 Microglial Cells. J. Med. Food 2016, 19, 1023–1031. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Jeong, J.W.; Lee, D.S.; Yim, M.J.; Lee, J.M.; Han, M.H.; Kim, S.; Kim, H.S.; Kim, G.Y.; Park, E.K.; et al. Sargassum serratifolium extract attenuates interleukin-1β-induced oxidative stress and inflammatory response in chondrocytes by suppressing the activation of NF-κB, p38 MAPK, and PI3K/Akt. Int. J. Mol. Sci. 2018, 19, 2308. [Google Scholar] [CrossRef]
- Kim, M.-J.; Bae, N.-Y.; Kim, K.-B.-W.-R.; Park, S.-H.; Jang, M.-R.; Im, M.-H.; Ahn, D.H. Anti-inflammatory activity of ethanol extract of Sargassum miyabei Yendo via inhibition of NF-kB and MAPK activation. Microbiol. Biotechnol. Lett. 2016, 44, 442–451. [Google Scholar] [CrossRef]
- Liu, N.; Fu, X.; Duan, D.; Xu, J.; Gao, X.; Zhao, L. Evaluation of bioactivity of phenolic compounds from the brown seaweed of Sargassum fusiforme and development of their stable emulsion. J. Appl. Phycol. 2018, 30, 1955–1970. [Google Scholar] [CrossRef]
- Kang, S.y.; Kim, E.; Kang, I.; Lee, M.; Lee, Y. Anti-diabetic effects and anti-inflammatory effects of Laminaria japonica and hizikia fusiforme in skeletal muscle: In vitro and in vivo model. Nutrients 2018, 10, 491. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.S.; Mun, O.; Bae, M.J.; Lee, S.; Kim, M.; Lee, S.; Yu, K.H.; Kim, Y.Y.; Kong, C. Anti-inflammatory activity of ethanol extracts from Hizikia fusiformis fermented with lactic acid bacteria in LPS-stimulated RAW264.7 macrophages. J. Korean Soc. Food Sci. Nutr. 2015, 44, 1450–1457. [Google Scholar] [CrossRef]
- Kim, M.E.; Jung, Y.C.; Jung, I.; Lee, H.-W.; Youn, H.-Y.; Lee, J.S. Anti-inflammatory Effects of Ethanolic Extract from Sargassum horneri (Turner) C. Agardh on Lipopolysaccharide-Stimulated Macrophage Activation via NF-κB Pathway Regulation. Immunol. Invest. 2015, 44, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Lee, J.Y.; Song, D.; Pan, C.; Nho, C.W.; Kim, M.C.; Lee, E.H.; Jung, H.; Kim, H.; Kim, Y.S.; et al. Cancer chemopreventive effects of Korean seaweed extracts. Food Sci. Biotechnol. 2008, 17, 613–622. [Google Scholar]
- Sanjeewa, K.K.A.; Jayawardena, T.U.; Kim, H.S.; Kim, S.Y.; Ahn, G.; Kim, H.J.; Fu, X.; Jee, Y.; Jeon, Y.J. Ethanol extract separated from Sargassum horneri (Turner) abate LPS-induced inflammation in RAW 264.7 macrophages. Fish. Aquat. Sci. 2019, 22, 1–10. [Google Scholar] [CrossRef]
- Herath, K.H.I.N.M.; Cho, J.; Kim, A.; Kim, H.S.; Han, E.J.; Kim, H.J.; Kim, M.S.; Ahn, G.; Jeon, Y.J.; Jee, Y. Differential modulation of immune response and cytokine profiles of Sargassum horneri ethanol extract in murine spleen with or without Concanavalin A stimulation. Biomed. Pharmacother. 2019, 110, 930–942. [Google Scholar] [CrossRef]
- Jayawardena, T.U.; Asanka Sanjeewa, K.K.; Shanura Fernando, I.P.; Ryu, B.M.; Kang, M.C.; Jee, Y.; Lee, W.W.; Jeon, Y.J. Sargassum horneri (Turner) C. Agardh ethanol extract inhibits the fine dust inflammation response via activating Nrf2/HO-1 signaling in RAW 264.7 cells. BMC Complement. Altern. Med. 2018, 18, 249. [Google Scholar] [CrossRef]
- Kim, H.S.; Sanjeewa, K.K.A.; Fernando, I.P.S.; Ryu, B.; Yang, H.W.; Ahn, G.; Kang, M.C.; Heo, S.J.; Je, J.G.; Jeon, Y.J. A comparative study of Sargassum horneri Korea and China strains collected along the coast of Jeju island South Korea: Its components and bioactive properties. Algae 2018, 33, 341–349. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.A.; Fernando, I.P.S.; Kim, S.; Kim, W.; Jee, Y.; Jeon, Y. Ecklonia cava (Laminariales) and Sargassum horneri (Fucales) synergistically inhibit the lipopolysaccharide-induced inflammation via blocking NF-κB and MAPK pathways. Algae 2019, 34, 45–56. [Google Scholar] [CrossRef]
- Kang, B.-K.; Kim, K.-B.-W.-R.; Kim, M.-J.; Bark, S.-W.; Pak, W.-M.; Ahn, N.-K.; Choi, Y.-U.; Bae, N.-Y.; Park, J.-H.; Ahn, D.-H. Anti-inflammatory effect of Sargassum coreanum ethanolic extract through suppression of NF-kB pathway in LPS induced RAW264.7 cells in mice. Microbiol. Biotechnol. Lett. 2015, 43, 112–119. [Google Scholar] [CrossRef]
- Jeong, D.H.; Kang, B.K.; Kim, K.B.W.R.; Kim, M.J.; Ahn, D.H. Anti-inflammatory activity of Sargassum micracanthum water extract. J. Appl. Biol. Chem. 2014, 57, 227–234. [Google Scholar] [CrossRef]
- Jeong, D.-H.; Kim, K.-B.-W.-R.; Kim, M.-J.; Kang, B.-K.; Ahn, D.-H. Anti-inflammatory activity of ethanolic extract of Sargassum micracanthum. J. Microbiol. Biotechnol. 2013, 23, 1691–1698. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, Z.; Mathema, V.B.; Chae, D.; Yoo, E.-S.; Kang, H.-K.; Hyun, J.-W.; Lee, N.H.; Ko, M.-H.; Koh, Y.-S. Extracts of the seaweed Sargassum macrocarpum inhibit the CpG-induced inflammatory response by attenuating the NF-κB pathway. Food Sci. Biotechnol. 2014, 23, 293–297. [Google Scholar] [CrossRef]
- Cheon, J.M.; Kim, H.S.; Choi, E.O.; Kwon, D.H.; Choi, Y.H.; Kim, B.W.; Hwang, H.J. Anti-inflammatory activities of an ethanol extract of Sargassum macrocarpum in Lipopolysaccharide (LPS) stimulated RAW 264.7 macrophages. J. Life Sci. 2017, 27, 1437–1444. [Google Scholar]
- Kim, M.-J.; Kim, K.-B.-W.-R.; Jeong, D.-H.; Ahn, D.-H. Anti-inflammatory activity of ethanolic extract of Sargassum sagamianum in RAW 264.7 cells. Food Sci. Biotechnol. 2013, 22, 1113–1120. [Google Scholar] [CrossRef]
- Song, J.H.; Piao, M.J.; Han, X.; Kang, K.A.; Kang, H.K.; Yoon, W.J.; Ko, M.H.; Lee, N.H.; Lee, M.Y.; Chae, S.; et al. Anti-wrinkle effects of Sargassum muticum ethyl acetate fraction on ultraviolet B-irradiated hairless mouse skin and mechanistic evaluation in the human HaCaT keratinocyte cell line. Mol. Med. Rep. 2016, 14, 2937–2944. [Google Scholar] [CrossRef]
- Safhi, M.M.; Alam, M.F.; Sivakumar, S.M.; Anwer, T. Hepatoprotective potential of Sargassum muticum against STZ-induced diabetic liver damage in wistar rats by inhibiting cytokines and the apoptosis pathway. Anal. Cell. Pathol. 2019, 2019, 7958701. [Google Scholar] [CrossRef]
- Piao, M.J.; Kim, K.C.; Zheng, J.; Yao, C.W.; Cha, J.W.; Jin, S.; Yoon, W.J.; Kang, H.K.; Yoo, E.S.; Koh, Y.S.; et al. The ethyl acetate fraction of Sargassum muticum attenuates ultraviolet B radiation-induced apoptotic cell death via regulation of MAPK- and caspase-dependent signaling pathways in human HaCaT keratinocytes The ethyl acetate fraction of Sargassum muticum a. Pharm. Biol. 2014, 52, 1110–1118. [Google Scholar] [CrossRef]
- Na, H.-J.; Moon, P.-D.; Ko, S.-G.; Lee, H.-J.; Jung, H.-A.; Hong, S.-H.; Seo, Y.; Oh, J.-M.; Lee, B.-H.; Choi, B.-W.; et al. Sargassum hemiphyllum inhibits atopic allergic reaction via the regulation of inflammatory mediators. J. Pharmacol. Sci. 2005, 97, 219–226. [Google Scholar] [CrossRef]
- Shin, D.-B.; Han, E.-H.; Park, S.-S. Cytoprotective Effects of Phaeophyta Extracts from the Coast of Jeju Island in HT-22 Mouse Neuronal Cells. J. Korean Soc. Food Sci. Nutr. 2014, 43, 224–230. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Lee, D.; Kim, M.; Kong, C.; Yu, K.; Kim, Y.; Lee, S. Enhancement of anti-inflammatory activity by fermentation of Sargassum siliquanstrum. J. Life Sci. 2016, 26, 318–324. [Google Scholar] [CrossRef]
- Zhang, R.; Luo, W.; Bi, T.; Zhou, S. Evaluation of Antioxidant and Immunity-Enhancing Activities of Sargassum pallidum Aqueous Extract in Gastric Cancer Rats. Molecules 2012, 17, 8419–8429. [Google Scholar] [CrossRef]
- Balachandran, P.; Parthasarathy, V.; Ajay Kumar, T.V. Isolation of Compounds from Sargassum wightii by GCMS and the Molecular Docking against Anti-Inflammatory Marker COX2. Int. Lett. Chem. Phys. Astron. 2016, 63, 1–12. [Google Scholar] [CrossRef]
- Brglez Mojzer, E.; Knez Hrnčič, M.; Škerget, M.; Knez, Ž.; Bren, U. Polyphenols: Extraction methods, antioxidative action, bioavailability and anticarcinogenic effects. Molecules 2016, 21, 901. [Google Scholar] [CrossRef]
- Liu, L.; Jin, C.; Zhang, Y. Lipophilic phenolic compounds (Lipo-PCs): Emerging antioxidants applied in lipid systems. RSC Adv. 2014, 4, 2879–2891. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.A.; Fernando, I.P.S.; Kim, S.Y.; Kim, H.S.; Ahn, G.; Jee, Y.; Jeon, Y.J. In vitro and in vivo anti-inflammatory activities of high molecular weight sulfated polysaccharide; containing fucose separated from Sargassum horneri: Short communication. Int. J. Biol. Macromol. 2018, 107, 803–807. [Google Scholar] [CrossRef]
- Dore, C.M.P.G.; Faustino Alves, M.G.D.C.; Pofírio Will, L.S.E.; Costa, T.G.; Sabry, D.A.; De Souza Rêgo, L.A.R.; Accardo, C.M.; Rocha, H.A.O.; Filgueira, L.G.A.; Leite, E.L. A sulfated polysaccharide, fucans, isolated from brown algae Sargassum vulgare with anticoagulant, antithrombotic, antioxidant and anti-inflammatory effects. Carbohydr. Polym. 2013, 91, 467–475. [Google Scholar] [CrossRef]
- Hwang, P.-A.; Chien, S.-Y.; Chan, Y.-L.; Lu, M.-K.; Wu, C.-H.; Kong, Z.-L.; Wu, C.-J. Inhibition of lipopolysaccharide (LPS)-induced inflammatory responses by Sargassum hemiphyllum sulfated polysaccharide extract in RAW 264.7 macrophage cells. J. Agric. Food Chem. 2011, 59, 2062–2068. [Google Scholar] [CrossRef]
- Preetha, S.P.; Devaraj, H. Role of sulphated polysaccharides from Sargassum wightii in the control of diet-induced hyperlipidemia and associated inflammatory complications in rats. Eur. J. Inflamm. 2010, 8, 23–30. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.A.; Fernando, I.P.S.; Kim, E.-A.; Ahn, G.; Jee, Y.; Jeon, Y.-J. Anti-inflammatory activity of a sulfated polysaccharide isolated from an enzymatic digest of brown seaweed Sargassum horneri in RAW 264.7 cells. Nutr. Res. Pract. 2017, 11, 3–10. [Google Scholar] [CrossRef]
- Neelakandan, Y.; Venkatesan, A. Antinociceptive and anti-inflammatory effect of sulfated polysaccharide fractions from Sargassum wightii and Halophila ovalis in male Wistar rats. Indian J. Pharmacol. 2016, 48, 562. [Google Scholar] [CrossRef]
- Wen, Z.-S.; Xiang, X.-W.; Jin, H.-X.; Guo, X.-Y.; Liu, L.-J.; Huang, Y.-N.; OuYang, X.-K.; Qu, Y.-L. Composition and anti-inflammatory effect of polysaccharides from Sargassum horneri in RAW264.7 macrophages. Int. J. Biol. Macromol. 2016, 88, 403–413. [Google Scholar] [CrossRef]
- Wu, G.J.; Shiu, S.M.; Hsieh, M.C.; Tsai, G.J. Anti-inflammatory activity of a sulfated polysaccharide from the brown alga Sargassum cristaefolium. Food Hydrocoll. 2016, 53, 16–23. [Google Scholar] [CrossRef]
- Hwang, P.-A.; Hung, Y.-L.; Chien, S.-Y. Inhibitory activity of Sargassum hemiphyllum sulfated polysaccharide in arachidonic acid-induced animal models of inflammation. J. Food Drug Anal. 2015, 23, 49–56. [Google Scholar] [CrossRef]
- Lavanya, R.; Seethalakshmi, S.; Gopal, V.; Chamundeeswari, D. Effect of crude sulphated polysaccharide from marine brown algae in TPA induced inflammation on poly morphonuclear leukocytes. Int. J. Pharm. Pharm. Sci. 2015, 7, 100–102. [Google Scholar]
- Raafat, E.M.; Gamal-Eldeen, A.M.; El-Hussieny, E.A.; Ahmed, E.F.; Eissa, A.A. Polysaccharide extracts of the brown alga Sargassum asperifolium possess in vitro cancer chemopreventive properties. Nat. Prod. Res. 2014, 28, 2304–2311. [Google Scholar] [CrossRef]
- Neelakandan, Y.; Kanmani, P.; Satishkumar, R.; Paari, A.; Pattukumar, V.; Arul, V. Antinociceptive and Anti-Inflammatory activities of Sargassum wightii and Halophila ovalis sulfated polysaccharides in experimental animal models. J. Med. Food 2013, 16, 740–748. [Google Scholar]
- Fernando, I.P.S.; Asanka Sanjeewa, K.K.; Samarakoon, K.W.; Kim, H.-S.; Gunasekara, U.K.D.S.S.; Park, Y.-J.; Abeytunga, D.T.U.; Lee, W.W.; Jeon, Y.-J. The potential of fucoidans from Chnoospora minima and Sargassum polycystum in cosmetics: Antioxidant, anti-inflammatory, skin-whitening, and antiwrinkle activities. J. Appl. Phycol. 2018, 30, 3223–3232. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Sanjeewa, K.K.A.; Samarakoon, K.W. Antioxidant and anti-inflammatory functionality of ten Sri Lankan seaweed extracts obtained by carbohydrase assisted extraction. Food Sci. Biotechnol. 2018, 27, 1761–1769. [Google Scholar] [CrossRef]
- Gamal-Eldeen, A.M.; Ahmed, E.F.; Abo-Zeid, M.A. In vitro cancer chemopreventive properties of polysaccharide extract from the brown alga, Sargassum latifolium. Food Chem. Toxicol. 2009, 47, 1378–1384. [Google Scholar] [CrossRef]
- Vasquez, R.D.; Garcia-Meim, R.S.; Ramos, J.D.A. Polysaccharide extract from Sargassum siliquosum J.G. Agardh modulates production of pro-inflammatory cytokines in LPS-induced PBMC and delays coagulation time in-vitro. J. Harmon. Res. Pharm. 2014, 3, 101–112. [Google Scholar]
- Vasquez, R.D.; Ramos, J.D.A. Immunomodulatory effects of sulfated polysaccharides from Sargassum siliquosum J.G Agardh on peripheral blood mononuclear cells. Int. J. Res. Phytochem. Pharmacol. 2012, 22, 55–63. [Google Scholar]
- Vasquez, R.D.; Ramos, J.D.A.; Bernal, S.D. Chemopreventive properties of sulfated polysaccharide extracts from Sargassum siliquosum J.G. Agardh (Sargassaceae). Int. J. Pharma Bio Sci. 2012, 3, 333–345. [Google Scholar]
- Senni, K.; Gueniche, F.; Foucault-Bertaud, A.; Igondjo-Tchen, S.; Fioretti, F.; Colliec-Jouault, S.; Durand, P.; Guezennec, J.; Godeau, G.; Letourneur, D. Fucoidan a sulfated polysaccharide from brown algae is a potent modulator of connective tissue proteolysis. Arch. Biochem. Biophys. 2006, 445, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Deniaud-Bouët, E.; Hardouin, K.; Potin, P.; Kloareg, B.; Hervé, C. A review about brown algal cell walls and fucose-containing sulfated polysaccharides: Cell wall context, biomedical properties and key research challenges. Carbohydr. Polym. 2017, 175, 395–408. [Google Scholar] [CrossRef]
- Yoon, W.-J.; Heo, S.-J.; Han, S.-C.; Lee, H.-J.; Kang, G.-J.; Kang, H.-K.; Hyun, J.-W.; Koh, Y.-S.; Yoo, E.-S. Anti-inflammatory effect of sargachromanol G isolated from Sargassum siliquastrum in RAW 264.7 cells. Arch. Pharm. Res. 2012, 35, 1421–1430. [Google Scholar] [CrossRef]
- Heo, S.-J.; Yoon, W.-J.; Kim, K.-N.; Oh, C.; Choi, Y.-U.; Yoon, K.-T.; Kang, D.-H.; Qian, Z.-J.; Choi, I.-W.; Jung, W.-K. Anti-inflammatory effect of fucoxanthin derivatives isolated from Sargassum siliquastrum in lipopolysaccharide-stimulated RAW 264.7 macrophage. Food Chem. Toxicol. 2012, 50, 3336–3342. [Google Scholar] [CrossRef]
- Lee, J.-H.; Ko, J.-Y.; Samarakoon, K.; Oh, J.-Y.; Heo, S.-J.; Kim, C.-Y.; Nah, J.-W.; Jang, M.-K.; Lee, J.-S.; Jeon, Y.-J. Preparative isolation of sargachromanol E from Sargassum siliquastrum by centrifugal partition chromatography and its anti-inflammatory activity. Food Chem. Toxicol. 2013, 62, 54–60. [Google Scholar] [CrossRef]
- Jeon, Y.; Jung, Y.; Cheol, M.; Cheol, H.; Sung, K.; Kee, Y.; Kim, S. Sargahydroquinoic acid inhibits TNF a -induced AP-1 and NF-kB signaling in HaCaT cells through PPAR a activation. Biochem. Biophys. Res. Commun. 2014, 450, 1553–1559. [Google Scholar] [CrossRef]
- Kim, J.; Ahn, B.; Kong, C.; Kim, S. Protective effect of chromene isolated from Sargassum horneri against UV-A-induced damage in skin dermal fibroblasts. Exp. Dermatol. 2012, 21, 630–631. [Google Scholar] [CrossRef]
- Fernando, P.; Piao, M.J.; Hewage, S.; Kang, H.K.; Yoo, E.S.; Koh, Y.S.; Ko, M.H.; Ko, C.S.; Byeon, S.H.; Mun, S.R.; et al. Photo-protective effect of sargachromenol against UVB radiation-induced damage through modulating cellular antioxidant systems and apoptosis in human keratinocytes. Environ. Toxicol. Pharmacol. 2016, 43, 112–119. [Google Scholar] [CrossRef]
- Kim, E.A.; Kim, S.Y.; Kim, J.; Oh, J.Y.; Kim, H.S.; Yoon, W.J.; Kang, D.H.; Heo, S.J. Tuberatolide B isolated from Sargassum macrocarpum inhibited LPS-stimulated inflammatory response via MAPKs and NF-κB signaling pathway in RAW264.7 cells and zebrafish model. J. Funct. Foods 2019, 52, 109–115. [Google Scholar] [CrossRef]
- Gwon, W.-G.; Joung, E.-J.; Kwon, M.-S.; Lim, S.-J.; Utsuki, T.; Kim, H.-R. Sargachromenol protects against vascular inflammation by preventing TNF-α-induced monocyte adhesion to primary endothelial cells via inhibition of NF-κB activation. Int. Immunopharmacol. 2017, 42, 81–89. [Google Scholar] [CrossRef]
- Gwon, W.-G.; Lee, B.; Joung, E.-J.; Choi, M.-W.; Yoon, N.; Shin, T.; Oh, C.-W.; Kim, H.-R. Sargaquinoic acid inhibits TNF-α-induced NF-κB signaling, thereby contributing to decreased monocyte adhesion to human umbilical vein endothelial cells (HUVECs). J. Agric. Food Chem. 2015, 63, 9053–9061. [Google Scholar] [CrossRef]
- Ham, Y.M.; Yoon, W.J.; Lee, W.J.; Kim, S.C.; Baik, J.S.; Kim, J.H.; Lee, G.S.; Lee, N.H.; Hyun, C.G. Anti-inflammatory effects of isoketocharbroic acid from brown alga, Sargassum Micracanthum. EXCLI J. 2015, 14, 1116–1121. [Google Scholar]
- Yang, E.-J.; Ham, Y.M.; Yang, K.-W.; Lee, N.H.; Hyun, C.-G. Sargachromenol from Sargassum micracanthum inhibits the lipopolysaccharide-induced production of inflammatory mediators in RAW 264.7 macrophages. Sci. World J. 2013, 2013, 1–6. [Google Scholar]
- Heo, S.-J.; Jang, J.; Ye, B.-R.; Kim, M.-S.; Yoon, W.-J.; Oh, C.; Kang, D.-H.; Lee, J.-H.; Kang, M.-C.; Jeon, Y.-J.; et al. Chromene suppresses the activation of inflammatory mediators in lipopolysaccharide-stimulated RAW 264.7 cells. Food Chem. Toxicol. 2014, 67, 169–175. [Google Scholar] [CrossRef]
- Sah, S.K.; Kim, B.-H.; Park, G.-T.; Kim, S.; Hwa Jang, K.; Eun Jeon, J.; Shin, J.; Kim, T.-Y. Novel isonahocol E3 exhibits anti-inflammatory and anti-angiogenic effects in endothelin-1-stimulated human keratinocytes. Eur. J. Pharmacol. 2013, 720, 205–211. [Google Scholar] [CrossRef]
- Kang, G.-J.; Han, S.-C.; Yoon, W.-J.; Koh, Y.-S.; Hyun, J.-W.; Kang, H.-K.; Youl Cho, J.; Yoo, E.-S. Sargaquinoic acid isolated from Sargassum siliquastrum inhibits lipopolysaccharide-induced nitric oxide production in macrophages via modulation of nuclear factor-κB and c-Jun N -terminal kinase pathways. Immunopharmacol. Immunotoxicol. 2013, 35, 80–87. [Google Scholar] [CrossRef]
- Kim, E.A.; Kim, S.Y.; Ye, B.R.; Kim, J.; Ko, S.C.; Lee, W.W.; Kim, K.N.; Choi, I.W.; Jung, W.K.; Heo, S.J. Anti-inflammatory effect of Apo-9′-fucoxanthinone via inhibition of MAPKs and NF-kB signaling pathway in LPS-stimulated RAW 264.7 macrophages and zebrafish model. Int. Immunopharmacol. 2018, 59, 339–346. [Google Scholar] [CrossRef]
- Han, S.-C.; Kang, N.-J.; Yoon, W.-J.; Kim, S.; Na, M.-C.; Koh, Y.-S.; Hyun, J.-W.; Lee, N.-H.; Ko, M.-H.; Kang, H.-K.; et al. External application of apo-9-fucoxanthinone, isolated from Sargassum muticum, suppresses inflammatory responses in a mouse model of atopic dermatitis. Toxicol. Res. 2016, 32, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Chae, D.; Manzoor, Z.; Kim, S.; Kim, S.; Oh, T.-H.; Yoo, E.-S.; Kang, H.-K.; Hyun, J.-W.; Lee, N.; Ko, M.-H.; et al. Apo-9′-fucoxanthinone, isolated from Sargassum muticum, inhibits CpG-Induced inflammatory response by attenuating the mitogen-activated protein kinase pathway. Mar. Drugs 2013, 11, 3272–3287. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.-J.; Ham, Y.; Lee, W.; Lee, N.; Hyun, C.-G. Anti-inflammatory effects of apo-9′-fucoxanthinone from the brown alga, Sargassum muticum. DARU J. Pharm. Sci. 2013, 21, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Sun, P.; Li, Y.; Wu, G.; Nie, J. Structural characterization of a polysaccharide from Sargassum henslowianum, and its immunomodulatory effect on gastric cancer rat. Int. J. Biol. Macromol. 2018, 108, 1120–1127. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Jayawardena, T.U.; Sanjeewa, K.K.A.; Wang, L.; Jeon, Y.-J.; Lee, W.W. Anti-inflammatory potential of alginic acid from Sargassum horneri against urban aerosol-induced inflammatory responses in keratinocytes and macrophages. Ecotoxicol. Environ. Saf. 2018, 160, 24–31. [Google Scholar] [CrossRef]
- Maneesh, A.; Chakraborty, K. Pharmacological potential of sulfated polygalactopyranosyl-fucopyranan from the brown seaweed Sargassum wightii. J. Appl. Phycol. 2018, 30, 1971–1988. [Google Scholar] [CrossRef]
- Hwang, P.A.; Phan, N.N.; Lu, W.J.; Hieu, B.T.N.; Lin, Y.C. Low-molecular-weight fucoidan and high-stability fucoxanthin from brown seaweed exert prebiotics and anti-inflammatory activities in Caco-2 cells. Food Nutr. Res. 2016, 60, 1–9. [Google Scholar] [CrossRef]
- Sarithakumari, C.H.; Renju, G.L.; Kurup, G.M. Anti-inflammatory and antioxidant potential of alginic acid isolated from the marine algae, Sargassum wightii on adjuvant-induced arthritic rats. Inflammopharmacology 2013, 21, 261–268. [Google Scholar] [CrossRef]
- Sarithakumari, C.H.; Kurup, G.M. Alginic acid isolated from Sargassum wightii exhibits anti-inflammatory potential on type II collagen induced arthritis in experimental animals. Int. Immunopharmacol. 2013, 17, 1108–1115. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.A.; Jayawardena, T.U.; Kim, S.; Kim, H.; Ahn, G.; Kim, J.; Jeon, Y. Fucoidan isolated from invasive Sargassum horneri inhibit LPS-induced inflammation via blocking NF-κB and MAPK pathways. Algal Res. 2019, 41, 101561l. [Google Scholar] [CrossRef]
- Lin, H.V.; Tsou, Y.; Chen, Y.; Lu, W.; Hwang, P. Effects of low-molecular-weight fucoidan and high stability fucoxanthin on glucose homeostasis, lipid metabolism, and liver function in a mouse model of type II diabetes. Mar. Drugs 2017, 15, 113. [Google Scholar] [CrossRef] [PubMed]
- Casas, M.P.; Rodríguez-Hermida, V.; Pérez-Larrán, P.; Conde, E.; Liveri, M.T.; Ribeiro, D.; Fernandes, E.; Domínguez, H. In vitro bioactive properties of phlorotannins recovered from hydrothermal treatment of Sargassum muticum. Sep. Purif. Technol. 2016, 167, 117–126. [Google Scholar] [CrossRef]
- Lopes, G.; Sousa, C.; Silva, L.R.; Pinto, E.; Andrade, P.B.; Bernardo, J.; Mouga, T.; Valentão, P. Can phlorotannins purified extracts constitute a novel pharmacological alternative for microbial infections with associated inflammatory conditions? PLoS ONE 2012, 7, e31145. [Google Scholar] [CrossRef] [PubMed]
- Maneesh, A.; Chakraborty, K. Unprecedented antioxidative and anti-inflammatory aryl polyketides from the brown seaweed Sargassum wightii. Food Res. Int. 2017, 100, 640–649. [Google Scholar] [CrossRef]
- Kim, M.; Jeong, S.-M.; Kang, B.-K.; Kim, K.B.W.R.; Ahn, D. Anti-inflammatory effects of grasshopperketone from Sargassum fulvellum ethanol extract on lipopolysaccharide-induced inflammatory responses in RAW 264.7 cells. J. Microbiol. Biotechnol. 2019, 29, 820–826. [Google Scholar] [CrossRef]
- Balboa, E.M.; Conde, E.; Moure, A.; Falqué, E.; Domínguez, H. In vitro antioxidant properties of crude extracts and compounds from brown algae. Food Chem. 2013, 138, 1764–1785. [Google Scholar] [CrossRef]
- Yoon, W.-J.; Heo, S.-J.; Han, S.-C.; Lee, H.-J.; Kang, G.-J.; Yang, E.-J.; Park, S.-S.; Kang, H.-K.; Yoo, E.-S. Sargachromanol G regulates the expression of osteoclastogenic factors in human osteoblast-like MG-63 cells. Food Chem. Toxicol. 2012, 50, 3273–3279. [Google Scholar] [CrossRef]
- Chen, Z.; Xu, Y.; Liu, T.; Zhang, L.; Liu, H.; Guan, H. Comparative studies on the characteristic fatty acid profiles of four different Chinese medicinal Sargassum seaweeds by GC-MS and chemometrics. Mar. Drugs 2016, 14, 68. [Google Scholar] [CrossRef]
- Narayan, B.; Hosakawa, M.; Miyashita, K. Growth inhibition of human pro-myelocytic leukemia (HL-60) cells by lipid extracts of marine alga Sargassum marginatum (Fucales, Phaeophyta) harvested off Goa (west coast of India) with special reference to fatty acid composition. Indian J. Mar. Sci. 2004, 33, 355–360. [Google Scholar]
- Narayan, B.; Miyashita, K.; Hosokawa, M. Comparative evaluation of fatty acid composition of different Sargassum (Fucales, Phaeophyta) species harvested from temperate and tropical waters. J. Aquat. Food Prod. Technol. 2004, 13, 41–52. [Google Scholar]
- Sanina, N.M.; Goncharova, S.N.; Kostetsky, E.Y. Fatty acid composition of individual polar lipid classes from marine macrophytes. Phytochemistry 2004, 65, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Hasumi, K.; Peng, H.; Hu, X.; Wang, X.; Bao, B. Fibrinolytic Compounds Isolated from a Brown Alga, Sargassum fulvellum. Mar. Drugs 2009, 7, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Heras, B.; Hortelano, S. Molecular Basis of the Anti-Inflammatory Effects of Terpenoids. Inflamm. Allergy-Drug Targets 2009, 8, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Su, J.-H.; Dai, C.-F.; Huang, H.-H.; Wu, Y.-C.; Sung, P.-J.; Hsu, C.-H.; Sheu, J.-H. Terpenoid-related metabolites from a formosan soft coral Nephthea chabrolii. Chem. Pharm. Bull. (Tokyo) 2007, 55, 594–597. [Google Scholar] [CrossRef] [PubMed]
- Grabarczyk, M.; Wińska, K.; Mączka, W.; Potaniec, B.; Anioł, M. Loliolide—The most ubiquitous lactone. Folia Biol. Oecologica 2015, 11, 1–8. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information PubChem Database. Fucosterol, CID=5281326. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Fucosterol (accessed on 19 July 2019).
- National Center for Biotechnology Information PubChem Database. Sargachromanol D, CID=11247397. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Sargachromanol-D (accessed on 19 July 2019).
- National Center for Biotechnology Information PubChem Database. Sargachromanol E, CID=11339386. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Sargachromanol-E (accessed on 19 July 2019).
- ChemSpider Search and Share Chemistry Sargachromanol G. Available online: http://www.chemspider.com/Chemical-Structure.9582569.html (accessed on 19 July 2019).
- National Center for Biotechnology Information PubChem Database. Loliolide, CID=100332. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Loliolide (accessed on 19 July 2019).
- National Center for Biotechnology Information PubChem Database. Tuberatolide B, CID=50993746. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Tuberatolide-B (accessed on 19 July 2019).
- National Center for Biotechnology Information PubChem Database. Sargahydroquinoic acid, CID=10202734. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Sargahydroquinoic-acid (accessed on 13 August 2019).
- Boileau, A.C.; Merchen, N.R.; Wasson, K.; Atkinson, C.A.; Erdman, J.W. Cis-lycopene is more bioavailable than trans-lycopene in vitro and in vivo in lymph-cannulated ferrets. J. Nutr. 1999, 129, 1176–1181. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, Y.; Sashima, T.; Hosokawa, M.; Miyashita, K. Comparative evaluation of growth inhibitory effect of stereoisomers of fucoxanthin in human cancer cell lines. J. Funct. Foods 2009, 1, 88–97. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information PubChem Database. Fucoxanthin, CID=5281239. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Fucoxanthin (accessed on 19 July 2019).
- ChemSpider Search and Share Chemistry Apo-9-Fucoxanthinone. Available online: http://www.chemspider.com/Chemical-Structure.10200226.html (accessed on 19 July 2019).
- Daley, D.K.; Brown, K.J.; Badal, S. Fungal Metabolites; Elsevier Inc.: Amsterdam, The Netherlands, 2016; ISBN 9780128020999. [Google Scholar]
- National Center for Biotechnology Information PubChem Database. CID=13922639. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/13922639 (accessed on 13 August 2019).
- Ale, M.T.; Mikkelsen, J.D.; Meyer, A.S. Important determinants for fucoidan bioactivity: A critical review of structure-function relations and extraction methods for fucose-containing sulfated polysaccharides from brown seaweeds. Mar. Drugs 2011, 9, 2106–2130. [Google Scholar] [CrossRef]
- Park, S.-B.; Chun, K.-R.; Jung, Y.-M.; Kim, J.-K.; Suk, K.; Lee, W.-H. The differential effect of high and low molecular weight fucoidans on the severity of collagen-induced arthritis in mice. Phyther. Res. 2010, 24, 1384–1391. [Google Scholar] [CrossRef]
- Sinurat, E.; Saepudin, E.; Peranginangin, R.; Hudiyono, S. Immunostimulatory activity of brown seaweed-derived fucoidans at different molecular weights and purity levels towards white spot syndrome virus (WSSV) in shrimp litopenaeus vannamei. J. Appl. Pharm. Sci. 2016, 6, 082–091. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information PubChem Database. Alginic acid, CID=6850754. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Alginic-acid (accessed on 19 July 2019).
- Li, Y.; Fu, X.; Duan, D.; Liu, X.; Xu, J.; Gao, X. Extraction and identification of phlorotannins from the brown alga, Sargassum fusiforme (Harvey) Setchell. Mar. Drugs 2017, 15, 49. [Google Scholar] [CrossRef] [PubMed]
- Vijayan, R.; Chitra, L.; Penislusshiyan, S.; Palvannan, T. Exploring bioactive fraction of Sargassum wightii: In vitro elucidation of angiotensin-i-converting enzyme inhibition and antioxidant potential. Int. J. Food Prop. 2018, 21, 674–684. [Google Scholar] [CrossRef]
- Glombitza, K.W.; Schmidt, A. Trihydroxyphlorethols from the brown alga Carpophyllum angustifolium. Phytochemistry 1999, 51, 1095–1100. [Google Scholar] [CrossRef]
- Saso, L.; Valentini, G.; Casini, M.L.; Grippa, E.; Gatto, M.T.; Leone, M.G.; Silvestrini, B. Inhibition of heat-induced denaturation of albumin by nonsteroidal antiinflammatory drugs (NSAIDs): Pharmacological implications. Arch. Pharm. Res. 2001, 24, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, Y.; Kobayashi, M. Interaction of antiinflammatory drugs with serum proteins, especially with some biologically active proteins. J. Pharm. Pharmacol. 1968, 20, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Silvestrini, B.; Gugiielmotti, A.; Saso, L.; Cheng, C. Van Changes in concanavalin-A reactive proteins in inflammatory disorders for the control rats. Clin. Chem. 1989, 35, 2207–2211. [Google Scholar]
- Thinakaran, T.; Balamurugan, M.; Sivakumar, K. Screening of phycochemical constituents qualitatively and quantitatively certain seaweeds from Gulf of Mannar biosphere reserve. Int. Res. J. Pharm. 2012, 3, 261–265. [Google Scholar]
- Van Ginneken, V.J.T.; Helsper, J.P.F.G.; De Visser, W.; Van Keulen, H.; Brandenburg, W.A. Polyunsaturated fatty acids in various macroalgal species from north Atlantic and tropical seas. Lipids Health Dis. 2011, 10, 104–111. [Google Scholar] [CrossRef]
- Terasaki, M.; Miyashita, K.; Narayan, B.; Kawagoe, C.; Hosokawa, M.; Ito, A.; Kumon, H. Spatial and seasonal variations in the biofunctional lipid substances (fucoxanthin and fucosterol) of the laboratory-grown edible Japanese seaweed (Sargassum horneri Turner) cultured in the open sea. Saudi J. Biol. Sci. 2016, 24, 1475–1482. [Google Scholar] [CrossRef]
- Airanthi, M.K.W.A.; Sasaki, N.; Iwasaki, S.; Baba, N.; Abe, M.; Hosokawa, M.; Miyashita, K. Effect of brown seaweed lipids on fatty acid composition and lipid hydroperoxide levels of mouse liver. J. Agric. Food Chem. 2011, 59, 4156–4163. [Google Scholar] [CrossRef]
- Mori, K.; Ooi, T.; Hiraoka, M.; Oka, N.; Hamada, H.; Tamura, M.; Kusumi, T. Fucoxanthin and its metabolites in edible brown algae cultivated in deep seawater. Mar. Drugs 2008, 2, 63–72. [Google Scholar] [CrossRef]
- Lann, K.L.; Ferret, C.; Vanmee, E.; Spagnol, C.; Lhuillery, M.; Payri, C.; Stiger-Pouvreau, V. Total phenolic, size-fractionated phenolics and fucoxanthin content of tropical Sargassaceae (Fucales, Phaeophyceae) from the South Pacific Ocean: Spatial and specific variability. Phycol. Res. 2012, 60, 37–50. [Google Scholar] [CrossRef]
- Mouritsen, O.G.; Bagatolli, L.A.; Duelund, L.; Garvik, O.; Ipsen, J.H.; Simonsen, A.C. Effects of seaweed sterols fucosterol and desmosterol on lipid membranes. Chem. Phys. Lipids 2017, 205, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Fleury, B.G.; Pereira, M.V.G.; Da Silva, J.R.P.; Kaisin, M.; Teixeira, V.L.; Kelecom, A. Sterols from brazilian marine brown algae. Phytochemistry 1994, 37, 1447–1449. [Google Scholar] [CrossRef]
- Chkhikvishvili, I.D.; Ramazanov, Z.M. Phenolic substances of brown algae and their antioxidant activity. Appl. Biochem. Microbiol. 2000, 36, 289–291. [Google Scholar] [CrossRef]
- Targett, N.M.; Coen, L.D.; Boettcher, A.A.; Tanner, C.E. Biogeographic comparisons of marine algal polyphenolics: Evidence against a latitudinal trend. Oecologia 1992, 89, 464–470. [Google Scholar] [CrossRef]
- Bolser, R.C.; Hay, M.E. Are tropical plants better defended? Palatability and defenses of temperate vs. tropical seaweeds. Ecology 1996, 77, 2269–2286. [Google Scholar] [CrossRef]
- Audibert, L.; Fauchon, M.; Blanc, N.; Hauchard, D.; Ar Gall, E. Phenolic compounds in the brown seaweed Ascophyllum nodosum: Distribution and radical-scavenging activities. Phytochem. Anal. 2010, 21, 399–405. [Google Scholar] [CrossRef]
- Abdelkafi-Koubaa, Z.; Drira, N.; Khemakhem, B.; Mejdoub, H.; Marrakchi, N.; Chakroun, M.; El Abed, H. Antioxidant, Anti-Inflammatory, and Antitumoral Effects of Aqueous Ethanolic Extract from Phoenix dactylifera L. Parthenocarpic Dates. Biomed Res. Int. 2018, 2018, 1–7. [Google Scholar]
- Kwak, C.S.; Son, D.; Chung, Y.S.; Kwon, Y.H. Antioxidant activity and anti-inflammatory activity of ethanol extract and fractions of Doenjang in LPS-stimulated RAW 264.7 macrophages. Nutr. Res. Pract. 2015, 9, 569–578. [Google Scholar] [CrossRef]
- Mhadhebi, L.; Mhadhebi, A.; Robert, J.; Bouraoui, A. Antioxidant, anti-inflammatory and antiproliferative effects of aqueous extracts of three mediterranean brown seaweeds of the Genus Cystoseira. Iran. J. Pharm. Res. 2014, 13, 207–220. [Google Scholar] [PubMed]
- Zhang, L.; Ravipati, A.S.; Koyyalamudi, S.R.; Jeong, S.C.; Reddy, N.; Smith, P.T.; Bartlett, J.; Shanmugam, K.; Münch, G.; Wu, M.J. Antioxidant and anti-inflammatory activities of selected medicinal plants containing phenolic and flavonoid compounds. J. Agric. Food Chem. 2011, 59, 12361–12367. [Google Scholar] [CrossRef] [PubMed]
- Chale-Dzul, J.; Freile-Pelegrín, Y.; Robledo, D.; Moo-Puc, R. Protective effect of fucoidans from tropical seaweeds against oxidative stress in HepG2 cells. J. Appl. Phycol. 2017, 29, 2229–2238. [Google Scholar] [CrossRef]
- Sugiono Widjanarko, S.B.; Soehono, L.A. Extraction optimization by response surface methodology and characterization of Fucoidanfrom brown seaweed Sargassum polycystum. Int. J. ChemTech Res. 2014, 6, 195–205. [Google Scholar]
- Wang, C.Y.; Wu, T.C.; Hsieh, S.L.; Tsai, Y.H.; Yeh, C.W.; Huang, C.Y. Antioxidant activity and growth inhibition of human colon cancer cells by crude and purified fucoidan preparations extracted from Sargassum cristaefolium. J. Food Drug Anal. 2015, 23, 766–777. [Google Scholar] [CrossRef]
- Vijayabaskar, P.; Vaseela, N.; Thirumaran, G. Potential antibacterial and antioxidant properties of a sulfated polysaccharide from the brown marine algae Sargassum swartzii. Chin. J. Nat. Med. 2012, 10, 421–428. [Google Scholar] [CrossRef]
- Bruhn, A.; Janicek, T.; Manns, D.; Nielsen, M.M.; Balsby, T.J.S.; Meyer, A.S.; Rasmussen, M.B.; Hou, X.; Saake, B.; Göke, C.; et al. Crude fucoidan content in two North Atlantic kelp species, Saccharina latissima and Laminaria digitata—Seasonal variation and impact of environmental factors. J. Appl. Phycol. 2017, 29, 3121–3137. [Google Scholar] [CrossRef]
- Kokilam, G.; Vasuki, S.; Sajitha, N. Biochemical composition, alginic acid yield and antioxidant activity of brown seaweeds from Mandapam Region, Gulf of Mannar. J. Appl. Pharm. Sci. 2013, 3, 99–104. [Google Scholar]
- Aponte de Otaola, N.E.; Diaz-Piferrer, M.; Graham, H.D. Seasonal variations and anatomical distribution of alginic acid in Sargassum spp. found along the coasts of Puerto Rico. J. Agric. Puerto Rico 1983, 67, 464–475. [Google Scholar]
- Ehrig, K.; Alban, S. Sulfated galactofucan from the brown alga saccharina latissima-variability of yield, structural composition and bioactivity. Mar. Drugs 2015, 13, 76–101. [Google Scholar] [CrossRef]
- So, M.-J.; Kim, B.-K.; Choi, M.-J.; Park, K.-Y.; Rhee, S.-H.; Cho, E.-J. Protective activity of fucoidan and alginic acid against free radical-induced oxidative stress under in vitro and cellular system. J. Food Sci. Nutr. 2007, 12, 191–196. [Google Scholar] [CrossRef]
- Galis, Z.S.; Sukhova, G.K.; Lark, M.W.; Libby, P. Increased expression of matrix metalloproteinases and matrix degrading activity in vulnerable regions of human atherosclerotic plaques. J. Clin. Invest. 1994, 94, 2493–2503. [Google Scholar] [CrossRef]
- Berry, C.E.; Hare, J.M. Xanthine oxidoreductase and cardiovascular disease: Molecular mechanisms and pathophysiological implications. J. Physiol. 2004, 555, 589–606. [Google Scholar] [CrossRef]
- Folmer, F.; Jaspars, M.; Dicato, M.; Diederich, M. Marine natural products as targeted modulators of the transcription factor NF-κB. Biochem. Pharmacol. 2008, 75, 603–617. [Google Scholar] [CrossRef]
- Chen, H.; Tan, H.; Yang, J.; Wei, Y.; Hu, T. Sargassum polysaccharide inhibits inflammatory response in PCV2 infected-RAW264.7 cells by regulating histone acetylation. Carbohydr. Polym. 2018, 200, 633–640. [Google Scholar] [CrossRef]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell. Mol. Life Sci. 2016, 73, 3221–3247. [Google Scholar] [CrossRef]
- Lee, S.; Yeon, S.L.; Sang, H.J.; Sam, S.K.; Kuk, H.S. Anti-oxidant activities of fucosterol from the marine algae Pelvetia siliquosa. Arch. Pharm. Res. 2003, 26, 719–722. [Google Scholar] [CrossRef]
- Makarenkova, I.D.; Logunov, D.Y.; Tukhvatulin, A.I.; Semenova, I.B.; Besednova, N.N.; Zvyagintseva, T.N. Interactions between sulfated polysaccharides from sea brown algae and toll-like receptors on HEK293 eukaryotic cells in vitro. Bull. Exp. Biol. Med. 2012, 154, 241–244. [Google Scholar] [CrossRef]
- Russell, F.D.; Bürgin-Maunder, C.S. Distinguishing health benefits of eicosapentaenoic and docosahexaenoic acids. Mar. Drugs 2012, 10, 2535–2559. [Google Scholar] [CrossRef]
- Tian, Y.; Katsuki, A.; Romanazzi, D.; Miller, M.R.; Adams, S.L.; Miyashita, K.; Hosokawa, M. Docosapentaenoic Acid (22:5n-3) Downregulates mRNA Expression of Pro-inflammatory Factors in LPS-activated Murine Macrophage Like RAW264.7 Cells. J. Oleo Sci. 2017, 66, 1149–1156. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Observed Response | Tested Compound | Model | Ref. |
|---|---|---|---|---|
| Subgenus Sargassum | ||||
| S. binderi |
| Phyto-oleic acid nanovesicles (PONVs) made by petroleum ether extract 1; water extract 2; and ethanolic precipitate of water extract 3 | Wounded streptozotocin-induced diabetic rats 1; and lipopolysaccharide (LPS)-induced RAW 264.7 2,3 | 1 [34] * 2 [35] * 3 [36] * |
| S. patens |
| Ethanol extract 1,2 | LPS-induced RAW 264.7 1,2; and croton oil-induced rat ear edema 1 | 1 [37] 2 [38] |
| S. wightii |
| Methanol-ethyl acetate extract 1; chloroform extract 1,3; ethanol extract 2,3; hexane extract 3, 6; methanol extract 4,5,6; butanol extract 4; and ethyl-acetate extract 6 | In vitro inhibition of 5-LOX, COX-1, and COX-2 1; albumin denaturation inhibition 2,6; carrageenan-induced rat paw edema 3,4,5; and RBC membrane stabilization and proteinase inhibition 6 | 1 [39] * 2 [40] * 3 [41] * 4 [23] 5 [42] * 6 [43] * |
| S. swartzii |
| Gold nanoparticles of water extract 1; methanol extract 2; water extract 3; and diethyl ether fraction of methanol extract 4 | Alloxan-induced diabetic Wistar rats 1; carrageenan-induced rat paw edema and peritonitis 2; acetaminophen-induced hepatotoxicity in mice 3; and LPS-induced RAW 264.7 4 | 1 [44] * 2 [45] * 3 [46] 4 [47] * |
| S. crassifolium | Decreased rat paw edema volume | β-glucan extract yielded from acid and ultrasound methods | Carrageenan-induced rat (Rattus novergicus) paw edema | [48] * |
| S. ilicifolium | Decreased paw edema volume | Methanol extract; | Carrageenan-induced rat paw edema | [49] * |
| S. duplicatum | Decreased NO production 1,2 | Water extract 1; and ethanolic precipitate of water extract 2 | LPS-induced RAW 264.7 1,2 | 1 [35] * 2 [36] * |
| S. subrepandum | Improved plasma lipid profile (plasma cholesterol, triglycerides, low density lipoprotein/LDL, and high density lipoprotein/HDL), decreased serum malondialdehyde (MDA), NO, TNF-α, and leptin production, and increased serum adiponectin level | Methanol extract (100 mg/kg) | Atherogenic diet-induced female Sprague Dawley rats | [50] |
| S. polycystum |
| Hexane fraction from methanol extract 1; ethanol extract 2,3; water extract 3; hexane, dichloromethane, and methanol extract 4 | Carrageenan-induced rat paw edema 1; acetaminophen-induced hepatoxicity in rat 2; high calorie diet and low dose streptozotocin-induced type II diabetes 3; and LPS-induced C8B4 microglia cells 4 | 1 [51] * 2 [52] * 3 [53] * 4 [54] * |
| S. vulgare |
| Methanol extract 1; ethyl acetate extract 2 | Carrageenan-induced rat paw edema 1; and in vitro inhibition to COX-1 and COX-2 enzymes 2 | 1 [55] 2 [56] |
| Subgenus Bactrophycus | ||||
| S. thunbergii |
| Ethanolic extract of fermented samples and its fractions 1; ethyl acetate extract 2; extract of dichloromethane, ethanol, and water 3; ethanol 70% extract 4; ethanol extract 5; and methanol extract 6,7,8 | LPS-induced RAW 264.7 1,2,5; rat ear edema 3; LPS-induced BV-2 microglial cells 4; H2O2-induced RAW 264.7 5; phorbol 12-myristate 13 acetate (PMA)-induced HT1080 6; PMA-induced mouse ear edema and erythema 7; and TNF-α stimulated human monocytic leukemia 8 | 1 [18] 2 [57] 3 [22] 4 [58] 5 [59] 6 [60] 7 [61] 8 [62] |
| S. fulvellum |
| Ethyl acetate fraction from ethanol extract 1, hexane fraction 2; ethanol extract 3,4,7; dichloromethane extract 4; and water extract 4,5; ethanol precipitate of water extract 6; and methanol extract 8 | Ultraviolet B (UVB)-induced HaCaT keratinocytes and BALB/c mice 1; LPS-induced RAW 264.7 2,3,5, 6; rat ear edema 3,4; dinitrochlorobenzene (DNCB)-induced atopic dermatitis (AD)-like skin lesions in BALB/c mice 7; and PMA-induced mouse ear edema and erythema 8 | 1 [24] 2 [63] 3 [64] 4 [22] 5 [35] * 6 [36] * 7 [65] 8 [61] |
| S. serratifolium |
| Ethanol extract 1,2,4; hexane fraction from ethanolic extract 3; meroterpenoid-rich extract from hexane fraction 5,6 | LPS-induced mouse peritoneal macrophage 1; LPS-induced BV-2 microglial cells and LPS-induced rat hippocampus cells 2; TNF-α-induced human umbilical vein endothelial cells (HUVECs) 3,6; IL-1β-treated SW1353 human chondrocytes 4; high fat (HF)-fed C57BL/6J mice 5; and high cholesterol diet (HCD)-fed C57BL/6J mice 6 | 1 [66] 2 [67] 3 [20] 4 [68] 5 [28] 6 [29] |
| S. miyabei |
| Ethanol extract | LPS-induced RAW 264.7 and rat ear edema | [69] |
| S. fusiforme |
| Administration of diethyl ether fraction percutaneously and orally 1; ethyl acetate fraction from ethanol 30% extract 2; water extract 3; methanol extract and its fractions (dichloromethane, ethyl acetate, n-butanol, water) 4; ethanol extract of fermented and non-fermented sample 5; and methanol extract 6 | Rat ear edema and RBL cells 1, LPS-induced RAW 264.7 2,4,5; tert-butyl hydroperoxide (t-BHP)-induced RAW 264.7 RAW 264.7 4; LPS-induced C2C12 myotube cells 3; and PMA-induced mouse ear edema and erythema 6 | 1 [31] 2 [70] 3 [71] 4 [33] 5 [72] 6 [61] |
| S. horneri |
| Ethanol 70% extract 1,4,5; ethanol 85% extract 2; ethanol extract 3,6,7,10 and its fraction 3,6; methanol 80% extract and its fraction 8; combination of Ecklonia cava and S. horneri ethanol 70% extracts 9; and methanol extract 11 | LPS-induced RAW 264.7 1,2,4,6,8,9,10; PMA-induced HT1080 fibrosarcoma 3; concanavalin A-induced rat splenocytes 5; fine dust (FD)-induced RAW 264.7 7; and PMA-induced mouse ear edema and erythema 11 | 1 [73] 2 [74] 3 [30] 4 [75] 5 [76] 6 [19] 7 [77] 8 [78] 9 [79] 10 [59] 11 [61] |
| S. coreanum |
| Ethanol extract | LPS-induced RAW 264.7 and rat ear edema | [80] |
| S. ringgoldianum | Suppression of mouse ear edema and erythema formation | Methanol extract | PMA-induced mouse ear edema and erythema | [61] |
| S. micracanthum |
| Water extract 1; and ethanol extract 2 | LPS-induced RAW 264.7 1; and croton oil-induced rat ear edema 2 | 1 [81] 2 [82] |
| S. macrocarpum |
| Ethanol 70% extract 1,2 | CpG-DNA-induced bone marrow-derived macrophages (BMDMs) and bone marrow-derived dendritic cells (BMDCs) from C57BL/6 mice 1; and LPS-induced RAW 264.7 2 | 1 [83] 2 [84] |
| S. sagamianum |
| Ethanol extract 1; and methanol extract 2 | LPS-induced RAW 264.7 and rat ear edema 1; and PMA-induced mouse ear edema and erythema 2 | 1 [85] 2 [61] |
| S. muticum |
| Ethanol 80% extract and its fraction (hexane, CH2Cl2, EtOAc, BuOH, and water) 1; ethanol 70% extract 2; ethyl acetate fraction of ethanol extract 3,5; and methanol extract 4 | LPS-induced RAW 264.7 1; Collagen-induced arthritis DBA/1J mice 2; UVB-induced HaCaT keratinocytes 3,5; and STZ-induced hepatic injury in Wistar rats 4 | 1 [21] 2 [27] 3 [86] 4 [87] 5 [88] |
| S. hemiphyllum |
| Methanol extract 1,2; and ethanol extract 3 | PMA-induced rat peritoneal mastocyte, A23187-induced HMC-1/human mast cell, and TNF-α-induced 293T cells 1; β-amyloid protein (Aβ)-induced HT-22 mouse neuronal cells 2; and LPS and H2O2-induced RAW 264.7 3 | 1 [89] 2 [90] 3 [59] |
| S. confusum |
| Methanol extract 1,2 | β-amyloid protein (Aβ)-induced HT-22 mouse neuronal cells 1; and PMA-induced mouse ear edema and erythema 2 | 1 [90] 2 [61] |
| S. siliquastrum | Suppression of NO and iNOS production | Aqueous extract of sample fermented by Lactobacillus sp. SH-1 | LPS-induced RAW 264.7 | [91] |
| S. pallidum |
| Water extract | N-methyl-N′-nitro-nitrosoguanidine (MNNG)-induced gastric cancer rats | [92] |
| Sample | Observed Response | Tested Compound | Model | Ref |
|---|---|---|---|---|
| Subgenus Sargassum | ||||
| S. wightii |
| CSP 2,3; and fraction of CSP (Fr IV) 1 | Carrageenan-induced rat paw edema, carrageenan-induced peritonitis, and Freund’s adjuvant-induced arthritis 1,2; and hypercholesterol diet-induced rat dyslipidemia 3 | 1 [101] * 2 [107] * 3 [99] * |
| S. cristaefolium |
| CSP and its fraction (1193.2, 864.4, 386.1 kDa, 55.9, 15.4, and 1.9 kDa) | LPS-induced RAW 264.7 | [103] |
| S. ilicifolium |
| CSP | TPA (12- O- Techanoyl 13—Myristate)-induced polymorphonuclear leukocytes (PMNL) | [105] * |
| S. asperifolium | Decreased production of NO and TNF-α | CSP | LPS-induced lymph macrophage | [106] |
| S. vulgare | Suppressed paw edema formation | CSP | Carrageenan-induced rat paw edema | [97] * |
| S. polycystum | Decreased production of NO 1,2, PGE2, TNF-α, IL-1β, and IL-6 1 | CSP 1,2 | LPS-induced RAW 264.7 | 1 [108] * 2 [109] * |
| S. latifolium | Decreased production of NO, TNF-α, and COX-2 | Different fraction of water-soluble polysaccharide extracts (not only sulfated form) | LPS-induced RAW 264.7 | [110] |
| S. siliquosum |
| Different fraction of water-soluble polysaccharide extracts (not only sulfated form) 1,2,3 | LPS-induced peripheral blood mononuclear cells 1,2; and LPS-stimulated promyelocyticleukemic cells 3 | 1 [111] * 2 [112] * 3 [113] * |
| Subgenus Bactrophycus | ||||
| S. horneri |
| Fraction of CSP yielded from membrane filtration (<5 kDa (f1), 5–10 kDa (f2), 10–30 kDa (f3), and >30 kDa (f4)) 1; and CSP resulted from Celluclast enzyme digestion 2, CSP and its fractions (Q Sepharose Fast Flow column) 3 | LPS-induced RAW 264.7 1,2,3; and LPS-induced zebra fish 1 | 1 [96] 2 [100] 3 [102] |
| S. hemiphyllum |
| CSP | Arachidonic acid-induced rat ear edema 1; and LPS-induced RAW 264.7 2 | 1 [104] 2 [98] |
| Compound | Source | Modulation of Inflammation | Ref |
|---|---|---|---|
| Terpenoid group | |||
| Fucosterol | S. binderi | Suppression of COX-2, PGE2, TNF-α, and IL-6 production via the inhibition of NF-kB activation and MAPK group phosphorylation | [26]* |
| Sargachromenol | S. serratifolium | Suppression of adhesion molecules (VCAM-1, and ICAM-1) and chemotactic cytokine (MCP-1) production via inhibition of IKK-β - Ikβ phosphorylation, and NF-kB nuclear translocation in TNF-α-induced HUVECs | [123] |
| S. micracanthum | Suppression of pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6), PGE2, NO, COX-2, and iNOS production via inhibition of Ikβ degradation in LPS-induced RAW 264.7 | [126] | |
| S. horneri | Suppression of MMP-1, -2, and -9 via inhibition of AP-1 activation (c-Jun and c-Fos) in UVA-induced human derman fibroblast | [120] | |
| S. macrocarpum | Inhibition of JNK and ERK phosphorylation and increased ROS scavenging activity in UVB-induced HaCaT keratinocytes | [121] | |
| Sargaquinoic acid | S. serratifolium | Suppression of adhesion molecules (VCAM-1, and ICAM-1) and chemotactic cytokine (MCP-1, and IL-8) production via inhibition of Ikβ degradation in TNF-α-induced HUVECs | [124] |
| S. siliquastrum | Suppression of iNOS and NO production via inhibition of Ikβ degradation, NF-kB nuclear translocation, and JNK1/2 phosphorylation in LPS-induced RAW 264.7 | [129] | |
| Sargahydroquinoic acid | S. yezoense | Suppression of MMP-2/-9 expression via inhibition of NF-kB nuclear translocation, Ikβ degradation, and AP-1 activation in TNF-α stimulated HaCaT cells | [119] |
| Sargachromanol D | S. siliquastrum | Suppression of pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6), PGE2, NO, COX-2, and iNOS production via inhibition of p65 and Ikβ-α phosphorylation in LPS-induced RAW 264.7 | [127] |
| Sargachromanol E | S. siliquastrum | Suppression of pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6), PGE2, NO, COX-2, and iNOS production via inhibition of MAPKs group phosphorylation (JNK, ERK, and p38) LPS-induced RAW 264.7 | [118] |
| Sargachromanol G | S. siliquastrum | Suppression of pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6), PGE2, NO, COX-2, and iNOS production via inhibition of IkB-α, NF-κB (p65 and p50), and MAPK (ERK1/2, JNK, and p38) phosphorylation in LPS-induced RAW 264.7 | [116] |
| Suppression of osteoclastogenic factor (PGE2, COX-2, IL-6, OPG, and RANKL) via inhibition of IkB-α, NF-κB (p65 and p50), and MAPKs (ERK1/2, JNK, and p38) phosphorylation in IL-1β-induced MG-63 osteoblast cells | [147] | ||
| Isoketochabrolic acid (IKCA) | S. micracanthum | Suppression of pro-inflammatory cytokines (TNF-α, IL-6, and IL-1β), PGE2, NO, COX-2, and iNOS production in LPS-induced RAW 264.7 | [125] |
| Tuberatolide B | S. macrocarpum | Suppression of NO, PGE2, IL-6, IL-1β, iNOS, and COX-2 production via inhibition of NF-κB (p65) and MAPK (ERK1/2, JNK, and p38) phosphorylation, and IkB degradation LPS-induced RAW 264.7 | [122] |
| Isonahocol E3 | S. siliquastrum | Suppression of IL-6, IL-8, and TNF-α production, and MMP gene expression via inhibition of ERK phosphorylation in ET-1-induced human keratinocytes | [128] |
| Loliolide | S. horneri | Suppression of NO production in LPS-induced RAW 264.7 | [19] |
| Carotenoid group | |||
| Fucoxanthin | S. siliquastrum | Suppression of pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6), PGE2, NO, COX-2, and iNOS production in LPS-induced RAW 264.7 | [117] |
| Apo-9′-fucoxanthinone | S. muticum | Suppression of NO 1,2, PGE2, proinflammatory cytokines (TNF-α, IL-6, and IL-1β), iNOS, and COX-2 production via inhibition of NF-κB (p65) and MAPK (ERK1/2, JNK, and p38) phosphorylation, and IkB degradation in LPS-induced RAW 264.7 1 | [130]1; [27]2 |
| Suppression of NO and PGE2 production via inhibition of Ikβ degradation in LPS-induced RAW 264.7 | [133] | ||
| Suppression of pro-inflammatory cytokines (IL-12 p40, TNF-α, and IL-6) and iNOS production via inhibition of ERK phosphorylation and AP-1 translocation in CpG DNA-induced BMDMs (bone marrow-derived macrophages) and BMDC (bone marrow-derived dendritic cells) | [132] | ||
| Suppression of IgE, IL-4, interferon- gamma, and TNF-α production, and lymph node size in atopic dermatitis rats | [131] | ||
| Other group | |||
| Aryl polyketide lactone | S. wightii | Direct inhibition of 5-LOX, COX-2, and COX-1 enzymes (in vitro) | [144] * |
| Grasshopper ketone | S. fulvellum | Suppression of pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6), NO, COX-2, and iNOS production via inhibition of p65 NF-κB nuclear translocation and MAPK (ERK1/2, JNK, and p38) phosphorylation in LPS-induced RAW 264.7 Suppression of IFN-γ and IL-4 production in concanavalin-A-induced BALB/c mice splenocytes | [145] [65] |
| Compound | Source | Modulation of Inflammation | Ref |
|---|---|---|---|
| Polysaccharides | |||
| Purified FCSPs (fucoidan) | S. wightii | Direct inhibition of 5-LOX, COX-2, and COX-1 enzymes (in vitro). | [136] * |
| S. henslowianum | Increased secretion of anti-inflammatory cytokines (IL-2, IL-4, and IL-10) and suppression of pro-inflammatory cytokines (IL-6 and TNF-α) production in MNNG-induced gastric cancer rats. | [134] | |
| S. hemiphyllum | A combination of oligofucoidan (LMF) and fucoxanthin resulted in enhancement of the intestinal epithelial barrier and immune function against LPS stimulation through suppression of IL-1β and TNF-α production and increased secretion of IL-10 and IFN-γ in CaCo2 cells co-cultured with B. lactis 1 A combination of oligofucoidan (LMF) and fucoxanthin resulted in enhancement of adiponectin production, and decreased production of TNF-α and IL-6 in type II diabetes mouse model 2 | [137]1 [141]2 | |
| S. horneri | Decreased NO production (IC50 = 40 μg/mL) via inhibition of the NF-κB and MAPK (ERK and p38) signaling pathways in LPS-stimulated RAW 264.7 cells. Decreased heart-beating rate, cell death, ROS, and NO levels in LPS-exposed zebrafish embryos. | [140] | |
| Alginic acid | S. horneri | Suppression of PGE2, proinflammatory cytokines (TNF-α, IL-6, and IL-1β), and COX-2 production via inhibition of NF-κB (p65) nuclear translocation and MAPK (ERK1/2, JNK, and p38) phosphorylation in CFD (Chinese fine dust)-induced HaCaT keratinocytes. | [135] |
| S. wightii | Suppression of COX-2, 5-LOX, MPO, xanthine oxidase (XO), ceruloplasmin, rheumatoid factor, CRP, pro-inflammatory cytokines, and lysosomal enzymes in type-2 collagen-induced rat arthritis. | [139] * | |
| Suppression of COX-2, 5-LOX, MPO, XO, ceruloplasmin, rheumatoid factor, and CRP production, and enhancement of antioxidant enzymes activity in Freund’s complete adjuvant-induced rat arthritis. | [138] * | ||
| Phenolic compounds | |||
| Phlorotannin | S. muticum | Suppression of ROS production in PMA-induced neutrophil and suppression of PGE2, COX-1, and COX-2 expression in A23187-induced erythrocytes. | [142] |
| S. vulgare | Suppression of NO production in LPS-induced RAW 264.7 and direct scavenging of NO in a cell-free system. | [143] | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saraswati; Giriwono, P.E.; Iskandriati, D.; Tan, C.P.; Andarwulan, N. Sargassum Seaweed as a Source of Anti-Inflammatory Substances and the Potential Insight of the Tropical Species: A Review. Mar. Drugs 2019, 17, 590. https://doi.org/10.3390/md17100590
Saraswati, Giriwono PE, Iskandriati D, Tan CP, Andarwulan N. Sargassum Seaweed as a Source of Anti-Inflammatory Substances and the Potential Insight of the Tropical Species: A Review. Marine Drugs. 2019; 17(10):590. https://doi.org/10.3390/md17100590
Chicago/Turabian StyleSaraswati, Puspo Edi Giriwono, Diah Iskandriati, Chin Ping Tan, and Nuri Andarwulan. 2019. "Sargassum Seaweed as a Source of Anti-Inflammatory Substances and the Potential Insight of the Tropical Species: A Review" Marine Drugs 17, no. 10: 590. https://doi.org/10.3390/md17100590